0022-538X/90/062876-08$02.00/0
Copyright C 1990, AmericanSociety forMicrobiology
Plasmid Origin of Replication of Herpesvirus Papio:
DNA
Sequence
and
Enhancer
Function
DANIEL D. LOEB,t NANCY SCHWAB SUNG, RICKL. PESANO, CONNIE J. SEXTON, CLYDE HUTCHISON III, ANDJOSEPH S. PAGANO*
Department
ofMicrobiology and Immunology and the Lineberger Cancer
ResearchCenter,
University of
NorthCarolina,
Chapel Hill, North Carolina 27599-7295
Received 16 October1989/Accepted 19 March 1990
Herpesvirus
papio
(HVP)
isalymphotropic
virus of baboons which is related toEpstein-Barr
virus(EBV)
and
produces latent infection.
Thenucleotidesequenceof the5,775-base-pair (bp)
EcoRIKfragment
ofHVP,
which
haspreviously
been showntoconfer theability
toreplicate autonomously,
has been determined. Within this DNAfragment
isaregion
which bears structural andsequencesimilarity
totheori-P
region
of EBV.The HVPori-P region
hasa10-
by 26-bp tandem
arraywhich
isrelatedtothe 20-by 30-bp
tandemarrayfromthe EBV oni-Pregion.
InHVP
there isanintervening region of
764bp
followedby
fivepartial copies
of the26-bp
monomer.
Both
theEBV
and HVP3' regions
have the potentialtoformdyad
structureswhich, however,
differ in arrangement. We also demonstrate that atranscriptional
enhancer whichrequires
transactivationby
avirus-encoded factor
ispresentin theHVP ori-P.The
episomal form of the Epstein-Barr
virus(EBV)
ge-nome is
thought
tobe the molecular
basis for latent EBVinfection (1, 25, 26, 28), but the
mechanism of thepersis-tence of the
episome
incells
was untilrecently largely
unknown.
EBVepisomes
areunit-length supercoiled
intra-cellular EBVgenomesfound innon-virus-producing
cellsatcopy numbers that are both limited and fixed in successive cell
generations (19, 24, 28). The episomes
areintranuclear
and inanucleosomal
arrangement(37)
andarethought
tobereplicated by host rather than
viral polymerase (5, 28).
Transcripts found spanning the fused genomic termini in
infected
cells indicate that
atleast
one EBV gene inthe
episomal
form istranscribed
(17).
Replication of EBV episomes requires
twocis-acting
elements
inthe BamHI-C region
of the genome,which
constitutetheplasmid origin
ofreplication (ori-P)
(35, 38, 40,
41). The cis-acting elements consist of
aseries
of
repeatunits located atthe
5' endof ori-P separated by
intervening
sequencefrom
an areaof dyad
symmetrylocated
at the 3' end(20),
wherereplication
is initiated(8). Transactivation
by
the EBNA-1protein
isrequired
for theepisomal
mainte-nancefunction of
ori-P, aswell
asfor
atranscriptional
enhanceractivity that
has been identified within the 5' elementof ori-P
(34).
We have
analyzed herpesvirus papio (HVP),
alymphotro-pic
virusof baboons
that is abletoimmortalize both human
and
baboon
Blymphocytes
invitro (31),
togeneralize these
mechanismsof
episomal maintenance
toanother virus.
HVP is similarstructurally
andbiologically
toEBV, but
onthe
genomic
level theviruses share
only 40% homology (7,
10,18).
Aregion
in HVPwith
sequenceand functional
homol-ogyto the EBV
ori-P
hasbeen
identified which requires
atrans-acting function analogous
tothat found in
EBV (29,30).
Thistransactivation
canbe supplied
by either EBNA-1 or HVPprotein, presumably
HVP nuclearantigen
(27).Conversely,
constructscontaining the
EBVori-P
canbe
maintained
inHVP-infected cells, although HVP nuclear
* Corresponding author.
t Present address:DepartmentofMicrobiologyandImmunology, University of California, San Francisco, CA 94143.
antigen
and EBNA-1 are not identicalproteins (6). We
therefore sequenced the HVP ori-Pto
identify
andcomparethe essential conserved features of the two
origins
ofrepli-cation. We
also
demonstratethat
atranscriptional
enhancer,
requiring
transactivationby
a virus-encodedfactor, exists
within the HVP ori-P.
MATERIALS
AND METHODSSequencing
of the5,775-base-pair (bp)
HVP EcoRI Kfragment.
The HVP EcoRI Kfragment
was subclonedand
sequenced
as describedby
Bankieretal.(3).
The sequencewas
compiled
andanalyzed
with programs describedby
Staden
(38).
Cell lines.
B95-8,
anEBV-productive
marmosetB-cellline
(21); Raji,
ahuman Burkitt'slymphoma-derived
linelatently
infected withEBV; Loukes,
an EBVgenome-negative
hu-manB-cellline;
and594-S(32),
ababoon B-cell lineproduc-tively
infected withHVP,
were maintainedat37°C
inRPMI 1640 mediumsupplemented
with10% fetal calfserum, 100U ofpenicillin
perml,
and100,ug
ofstreptomycin
perml ina5%
CO2
environment.DNAtransfections. Plasmid DNAwas
amplified
in Esche-richia coli andpurified
overtwosequential cesium
chloridegradients.
For eachsample,
10,ug
ofplasmid
DNA waselectroporated (23)
into107
cellsat1,500
V. Cellswerethensuspended
in 10 ml of RPMI 1640 mediumsupplemented
with
10% fetal calf
serumandincubated for 48 h
at37°C
in5%
CO2-Chloramphenicol acetyltransferase (CAT)
assays. Extracts wereprepared by washing cell pellets twice
inphosphate-buffered saline
solution, suspending
them in200
p.1of 0.25 MTris
hydrochloride (pH 7.5), and
freeze-thawing four
times.Protein
determinations
weremade
by using the Bio-Rad
Laboratoriesprotein
assay. Allsamples
ineach
assay con-tainedequal
amounts ofprotein from
extracts of107
cells. Reactions were carried out asdescribed
previously (9) by
incubating
eachsample with acetyl
coenzyme A and[14C]chloramphenicol
at37°C
for 60min. Acetylated
reac-tionproducts
wereseparated by
thin-layer chromatography,
visualizedby autoradiography,
andquantitated
by
scintilla-tion
counting.
2876on November 10, 2019 by guest
http://jvi.asm.org/
HVP ori-P 2877
A.
II I l
1000 2000 3000 4000 5000 5775
BNRF1 EBER
1 2
- V
Mat
-4 iB.
10 x 26 bp HVP II IfII
J-20x30bp
764bp
5x 16-18bp
67bp
ififliflliIllll EI-Mm
4x15-18 bp
C.
HVP5'monomer consensus
EBV5'monomer consensus
axisofsymmetry
FIG. 1. HVPand EBV ori-P regions. (A) Map ofsequencefeatures in the HVP EcoRI K fragment. The following featuresareincommon
with EBV. Open reading frames and direction oftranscriptionaredenoted byblackarrows. Bases 1to326correspondtothe 3'endofthe
EBV BNRF1reading frame. Bases 3770to4285correspondtothe EBVBCRF1 reading frame. EBER 1 and 2aredenoted bygrayarrows
andarefoundatbases 1235to1424and 1549to1717,respectively. The ori-P regionencompassespositions 2084to3220. (B)Organization ofori-Pregions of EBV and HVP. The ori-P of each virus has the generalstructureofanarrayofmonomerunits, followed byanintervening sequenceofvariable length, followed by fourorfive partial copies of themonomerunit which has thepotentialtoformalargedyad. HVP
hasanadditional twopartial copies of the monomerunitatthe 5' end of the intervening sequence. (C) Comparison of 5'array monomer
consensussequencesof EBV and HVP. SequencecommontoEBVand HVP is underlined (match in 16 of 19 bp).Arrowindicatesdyad of
symmetry in eachmonomerunit. HVPconsensus sequence wasgenerated by considering only the sequences in the 5' arrays. The EBV
consensus sequenceis takenfromFig. 6 of Lupton and Levine (20).
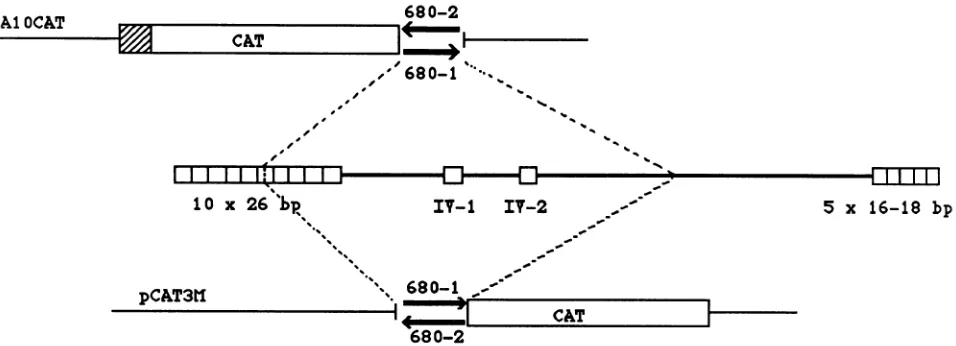
Plasmid
constructions.
pA1OCAT-680-1
andpA1OCAT-680-2 were
generated by cloning
a680-bp DNA fragment from
theHVPori-Pregion, containing
fiveof the
repeatunits
as well as twoimperfect
repeat unitsfrom the
intervening
sequence, inboth
orientations,
intothe
BamHI siteof
pA1OCAT
(see Fig. 4).
ThepA1OCAT
vectorcontains the
CAT geneunder the control of the simian virus 40
(SV40)
early
promoterbutlacks
theSV40 enhancer
sequences(16).
pCAT3M680-1
andpCAT3M680-2
weregenerated by
clon-ing
thesame680-bp fragment,
inbothorientations,
intotheBglII
site of thepromoterless
CATvectorpCAT3M (see Fig.
4) (15).
Asecondconstructcontaining
theentire motifof 10 repeatunits was alsocloned
intopA1OCAT.
RESULTS
Nucleotide sequence of the HVP EcoRI K
fragment.
Thenucleotide
sequence of the HVP EcoRI Kfragment
wasdetermined. It is
5,775 bp
inlength
and ishomologous
tosequences within the EBV BamHI C
fragment
(nucleotides
5367to 11635 of the EBV
sequence).
Within thisregion
of the EBV genome there are five notable features: the 3'portion
of the BNRF1reading frame;
the entire BCRF1reading frame;
twopolymerase
IIIRNAs,
EBER 1 and2;
and the
ori-P
region (Fig. 1A
andB).
Allof these featuresare also found in the HVP EcoRI Kfragment.
TheHVP EBER 1 and 2 are 78 and 66%homologous,
respectively,
to the EBV EBER sequencereported by
Howe and Shu(11).
Although
the HVP EcoRI Kfragment
and itscorresponding
EBV sequence are
homologous
for most oftheirlengths,
especially
in these functional domains, there areregions
inwhich there is no obvious sequence
homology.
Table 1containsasummary ofthedegreeof
homology
between theEBV and HVPsequences.
Sequence
ofthe HVPoni-Pregion
andcomparison
with theor P BCRFI
EBV VOL. 64,1990
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.69.557.72.445.2]TABLE 1. Sequence homology between the HVP EcoRI K fragment and the homologous region in EBV as a function
of nucleotideposition
HVP positions" EBVpositionsb % Homologyc EBVfeatured
1-326 5367-5692 73 BNRF1
327-754 5693-6153 70 3' to BNRF1
755-1050 6154-6406 NSH
1051-1234 6407-6628 56 5' to EBER 1
1235-1424 6629-6795 80 EBER 1
1425-1548 6796-6955 60 Intervening
1549-1717 6956-7128 66 EBER2
1718-1810 7129-7206 54 3' to EBER2
1811-1990 7207-7302 NSH
1991-2083 7303-7449 58 5' to array
2084-2347 7450-8032 84 10-by 26-bparray
2348-2621 8033-8554 NSH Intervening
2622-3090 8555-9032 50 Intervening
3134-3224 9033-9133 70 3' dyadmotif
3225-3611 9134-9515 NSH
3612-3769 9516-9674 64 5' toBCRF1
3770-4285 9675-10187 82 BCRF1
4286-4595 10188-10491 73 3'to BCRF1 4596-4895 10492-10765 NSH
4896-577 10766-11635 69
aReferstothe nucleotide coordinate within theHVP
EcoRI
Kfragment.bRefers to the nucleotide ccordinate within the published B95-8 EBV sequence(2).
cHomologyvalueswithinthe 10-by 26-bparrayreferto amatchin16of19 bp between theconsensus EBVandconsensus HVPmonomersequences. NSH,Nosignificanthomologyfound.
dFeatures arethosepreviously describedintheEBVB95-8sequence.
EBV
ori-P.
Although the general format of the HVP ori-P is
similar
to
that
of the EBV ori-P, there are considerable
differences
between the two ori-Ps on the nucleotide level.
To
facilitate the description of the HVP ori-P region, a brief
review
of
the
structural features of the EBV ori-P is in order.
The 5'
end of the EBV ori-P region has 20 copies of a 30-bp
unit
in a tandem array.
Approximately 987 bp downstream of
this
20-
by 30-bp tandem array are four partial copies of the
30-bp
monomer.
Dyad structures can be proposed in this
downstream
region, with monomers and surrounding
se-quence
contributing to the structure (14, 20, 33).
The HVP ori-P
is similar in its general structure, with the
following
specific differences (Fig. 1B). The 5' end of the
HVP ori-P
region has 10 imperfect and/or partial copies of a
26-bp unit
in tandemarray.This26-bp
monomeris related tothe
EBV30-bp
monomer(Fig.
1C;
seebelow).
Approxi-mately 764 bp downstream
of the 10-by 26-bp
array arefivepartial copies
of the26-bp
unit. Theintervening 764-bp
sequence harbors twoimperfect copies
of the26-bp
mono-mer,
which
arenotfound in the EBVori-P.
As inEBV,
the3'
sequence of the HVPori-P
has thepotential
to form alarge dyad (80 bp),
but unlike inEBV, only
monomersequence contributes to the
proposed dyad
structure(Fig.
2).
Comparison
of the EBV and HVPori-P repeat
units. The30-bp
EBV repeat monomer and the HVP26-bp
repeat monomer arehomologous
for 19 bp. Acomparison
of aconsensus sequenceof the repeat unitof each virusreveals a match in 16 of 19
bp (Fig. 1C).
Ifthe first base of theseconsensussequences is
discarded,
animperfect
9-by 9-bp
dyad
of symmetry can beformed,
aspreviously
notedfor EBV. Whenonly
the HVP repeat unit isconsidered,
thisdyad
can be extended to animperfect
13-by 13-bp dyad,
which comprises
the entire26-bp
unit. In EBV thedyad
cannotbe extended. Thus in the HVP
ori-P,
all 26bp
ofeachmonomeric
unit canform
animperfect dyad,
whereas in EBVonly
18bp
ofeach30-bp
monomeric unitcanform adyad.
Examination of
the patternof
polymorphisms
within the individual EBV repeat units revealsanimperfect duplication
of aputative
ancestral 10-by 30-bp
arrayleading
to thepresent-day 20- by 30-bp
array. Inthis
respect, the 10-by
26-bp
HVP arraywould
be more similar to the ancestral EBVarray in size.Toward
defining
arecognition
site forthe HVPtrans-acting
protein. The
HVPori-P, like the
EBV ori-P,requires
acti-vation by
atrans-acting protein. Since transactivation
ap-pears tobebrought
aboutby actual binding of
EBNA-1 to therepeatunits(22, 33),
thevariationstolerated
within the repeat monomers may be useful in furtherdefining
thesequence that the HVP transactivator
recognizes.
Whenonly
the5'tandemarrayof each virusisconsidered,
19bp
in themonomerunitis sharedby
bothviruses(Fig. 3).
Addi-tionally,
theportion
of the monomer unit that is commonbetween the 5' and 3'arrayswithin
both
viruses is the last 18bp
of the above-mentioned 19bp.
This18-bp
sequence ispalindromic
and contains the12-bp
coreEBNA-1-binding
sequence(12).
Aconsensussequence,GGRTAGYMTRYR
CTRYCC,
inwhich R isapurine,
Yisapyrimidine,
and MEBV
A
B
C DACCCTTTTAC TAACCCTAAT TCGATAGCAT ATGCTTCCCG TTGGGTAACA TATGCTATTGAATTAGGGTTAGTCTGGATA GTATATACTA CTACCCGGGA AGCATATGCT ACCCGTTTAGGG
11II llllll} 1l 1 I1 l IIIlllI IlllI ll lI 1H1I 11 lI lllI ll 11 II
GGACAGCACATACTGCCTCG CAGATAGCAT ATACCGCCGG CATGATAGCA TATGCTACCC AG... ...ATGGATA GCATACGCTA CCTCCCGGGT AGTATTTGCT ATCCGGGTAAGG
3113
3134
3155
3177
3198
HVP
FIG. 2. Sequencecomparison of the 3' dyadregions of HVP and EBV. The upper nucleotide sequence is EBV; the lower sequence is HVP. Labeledoverlined and underlined regions (A to D) indicate copies of the monomer unit. The regions ofdyad symmetry are indicated byarrows.Although other EBV dyad structures are possible, that proposed by Lupton and Levine (20) is shown here. The indicated HVP
dyad
is the largestamong several that can be formed.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.61.545.551.681.2]HVP ori-P 2879
EBV
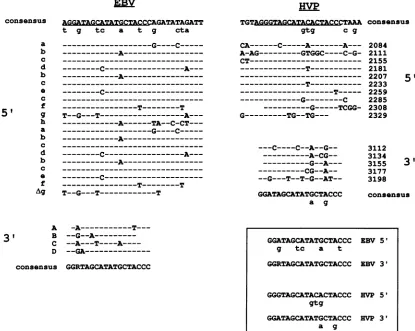
COnSenSUS A&GGZTAGCATATGCTACCCGATATAGATT
t g tc a t g cta
HVP
TGTAGGGTAGCATACACTACCCTAAA
gtg C g
consensus
---G----C---
---A---
---C---A---
A---c-
---T---T
T--G---T---A--- ---A---TA--C-CT---
---G----C---
---A--- C- ---
---T---T
T--G---T---T
CA---C---A---A--- A-AG---GTGGC----C-G- CT---
---T---__________________________
---T---
-_______---T---G---C ---G---TCGG-
G---TG--TG--- C----C--A--G-- ---A-CG-- ---G--A---
CG--A--
--G---T--T-G--AT--a g
A
-A---T---31 B
--G--A---C --A---T----A----D
--GA---consensus GGRTAGCATATGCTACCC
2084 2111 2155 2181 2207 2233 2259 2285 2308 2329
3112 3134 3155 3177 3198
5'
3'
consensus
FIG. 3. Comparisonof themonomer sequencesinthe HVP and EBVori-Pregions. Theconsensus sequenceis generated by noting the
mostcommon nucleotideatthatposition. Aconsensus wasgeneratedforthe5' and 3'arrays for each virus. Nonconsensus nucleotides
appearing inmore thanonemonomeraredenoted by lowercase letters beneath theconsensusnucleotides. Aligned belowthe consensus
sequencearetheindividual complete and partialmonomer sequences,withdifferences indicated.Namesappearnexttoeachmonomer.HVP
monomers wereidentified by searching the HVPEcoRI Kfragmentforamatchin 9 of 12 bp by thesequenceTAGCATATGCTA. Allbut
themonomeratposition 3113 wereidentified by this method. The 3113monomer wasvisuallyidentified. EBVmonomers weretaken from
Fig. 6 ofLupton and Levine (20). (Inset) Alignment ofcommonnucleotides intheconsensus sequencesfrom the 5' and 3'arraysisshown.
Minorvariantsaredenoted by lowercase letters beneath therespectiveconsensus sequence.Rindicatesapurine (AorG).
is A or
C,
canbe
derived from the EBV
and HVP 18-bppalindrome.
Demonstration of
atranscriptional
enhancerelement within the HVPori-P. The
EBVori-P
contains
atranscriptional
enhancer(34, 41). To determine whether enhancer activity
could
also be
localized
tothe 5'
repeatelement of the
HVPori-P,
constructsweremade containing HVP repeated units
linked
tothe
CAT
reportergene.A 680-bp
fragment
contain-ing five of
the
repeatunits and the
twoimperfect units from
the
intervening
sequence wascloned in both orientations
downstream of the CATgene in
the
pAjOCAT
vector(Fig.
4).
This680-bp fragment
wasable toenhance CAT
expres-sion
from the
SV40 early
promoterby
approximately 12-fold
in594-Scells
expressing
HVPgeneproducts and by 9-fold
in B95-8cellsexpressing
EBVgeneproducts
butwasunable
toenhance expression in EBV-
orHVP-negative cells (Fig.
5 andTable2).
To
demonstrate that the
enhancement observed
was notdueto promoter
activity intrinsic
to the680-bp
fragment, it
was
cloned
inboth
orientations into
thepromoterless
CAT
vectorpCAT3M (Fig. 4). No
enhancement
wasobserved in
anyofthe cell lines tested.
Representative
assaysareshown
inFig.
5.Additional
pAjOCAT
constructsconsisting of
a1,750-bp
[image:4.612.99.514.76.407.2]fragment (HVP EcoRI-K positions 1688
to2438
in both
orientations), which included all 10
repeatunits in the motif
but
omittedthe 2
repeatsinthe
intervening
sequence, were also tested. Whenthey
were assayedsimultaneously in
either
EBV-infected
orHVP-infected
cells, the CAT
activity
TABLE 2. Quantitation of enhancement of CATexpression in different celllines
Virusa Foldenhancementb Cellline
EBV HVP 680-1 680-2
Loukes - - 1.7(8) 0.7(7)
B95-8 + - 9.3(6) 8.9(5)
594-S - + 13.9 (4) 11.8(5)
aThe presence (+)orabsence(-) of viralgenomesin each cell line is
indicated.
bRatio of the percent acetylation permicrogram ofextract in samples containing the680-bpinserttothepercentacetylation in samples containing the pA1oCATvectoralone, expressed as the average of the numberof determinations indicated inparentheses. 680-1 and 680-2designatethetwo orientationsof theinsert.
a
b
c
d b
c
0
c
f
5'
gh
a
b
c
d b
c
0
f
Ag
GGATAGCATATGCTACCC EBV 5'
g tc a t
GGRTAGCATATGCTACCC EBV 31
GGGTAGCATACACTACCC EVP 51
gtg
GGATAGCATATGCTACCC HVP 31
a g
VOL.
64,
1990on November 10, 2019 by guest
http://jvi.asm.org/
2880 LOEB ET AL.
680-2
CAT
l~~-w.
Il
,'
680-1
vf %.
1 0 x 26
bhp
IVr-1
IV-2
5
ITIT-1
5 X 16-18
bP
\
Is_680-1
-I
[image:5.612.73.551.73.246.2]680-2
FIG. 4. Plasmid constructions containing a portion of the HVP repeat motif.
pAjOCAT-680-1
andpAjOCAT-680-2
indicate the twoorientations inwhich the 680-bp fragmentwascloned into the BamHI sitedirectlydownstream of the CATgenein thevector
pAjOCAT.
Theposition of the SV40 earlypromoteris indicated(0). pCAT3M680-1and pCAT3M680-2indicate thesame680-bp fragmentclonedinboth
orientationsdirectlyupstreamof the CATgenein thepromoterlessCATexpressionvectorpCAT3M.Theregionof the HVPori-Pincluded
inthe680-bp fragment is shown (- --).
of
these
constructswasequivalent
toorstrongerthan that ofthe
original
constructshown in Fig.
4(data
notshown).
DISCUSSION
The HVP
ori-P, first
definedasthe functionalanalog
of the EBVori-P(29), is
shown in this work
tobe similar
structur-ally
tothe EBV
ori-P.Both ori-Ps have the
samegeneral
format andmanyspecial features
incommon onthe basis ofsequence.
However,
there aredifferences between the twoori-Ps
that mayimply
what isnecessary for function.The HVP
ori-P
consists
in essenceof
twoelements,
a5'array
consisting of
10 tandemcopies
ofa26-bp
unitand,
approximately 750 bp downstream
in the genome,5
addi-tional imperfect and partial copies of the
monomerunit.
EBV ori-Phas the
samegeneral
structure,although
it islarger. EBV
ori-Phas
20tandem copies of
a30-bp
monomerunit, which is related
tothe 26-bp HVP
monomerunit.
Approximately 1,000 bp downstream of this
arrayin EBV
arefouradditional imperfect and partial copies of the 30-bp
monomer
unit. Dyad
structures canbe proposed in this 3'
region (13), which have been hypothesized
tobe important
in ori-Pfunction
(20). Similar although
notidentical
dyad
structures can
be proposed for the 3' region of the HVP
ori-P.A
comparison of the
twoviral
ori-Pregions leads
us to proposethatthe
following general features
areimportant for
function. The 5'tandem
arraydoes
notneed
tobe
larger
than 10 monomerunits,
asevidenced
by the size of
the HVP 5' tandem array.This
conclusion is corroborated by
recentwork
(4, 39) in which deletion analysis of the
EBV 5'tandem
arrayindicates
that
atleast six
or seven monomerunits
areneeded for
ori-Pfunction.
However,since the
monomersactcooperatively
ratherthan additively
tobind
EBNA-1pro-tein, less than
aminimum
number of binding sites does notsimply reduce efficiency but abolishes
function.The
intervening
sequencebetween the
5'tandem
array and the3'
arrayis
thought
tobe
of lesser importance
in the EBVori-P, because
ori-Pfunction is
preservedin
constructsin which this sequence
is largely
deleted.Presumably,
theintervening
DNAplays
aminimal role in
ori-Pfunction
bycontributing
to the overallstructural
arrangement,rather
than
having
a direct sequence effect. The sequences ofthe HVP and EBVintervening
DNAarequite
different,
except for theincreasing homology (50
to60%)
of the last 400basesapproaching
the 3'dyad.
The3'arraysor
dyad regions
of EBV andHVP have four and fivepartial copies
of themonomerunits, respectively.
The
partial copies
arehomologous
toeach other. Thisregion
has
recently
beenproposed
to functionasthe site of initia-tion ofplasmid replication
in EBV(8),
andwe cansafely
saythat
only partial copies
of themonomerunitareneededfor function. The role ofdyad
formation is lessclear, although
we
have shown
that theareainwhich
thesestructuresform isspared
of nucleosomesduring
reassembly
invitro,
whichmayfacilitate initiation of
replication
inlatent infection(36).
Both EBV and HVP have the
potential
to from these structures.We have also demonstrated the existence ofa
transcrip-tional enhancer element within the ori-Pregion
of HVP. Enhancement was observed in the presence of fiveperfect
andtwo
imperfect
repeatsof themonomerunit,
which is the minimal number ofcopies required
for enhancement in EBV(4, 39).
Inotherconstructscontaining
10repeats,the level of enhancement wasgreater. Enhanceractivity
was mostpro-nounced in the HVP-infected cell line
594-S,
which wasexpected
since these cellspresumably produce
anauthentic HVP transactivatoranalogous
butnotidenticaltoEBNA-1. Enhancementwas alsoobserved,
to a lesserdegree,
in the EBV-infected cell lineB95-8, suggesting
that the HVP enhancer can be transactivatedby
an EBV geneproduct.
However,
wehave not identified the transactivatorgene orgenes, norhavewe
explored
whetherlymphoid
cell factorsmight
contributeto HVPenhancer function.Our results
clearly
demonstrate that the HVPori-P,
like its EBV counterpart,requires
transactivationby
factors present in both EBV- and HVP-infected cells for both enhancer andreplicative functions (29). Since binding
of the EBNA-1protein
tothe EBV repeatmonomersappearstobeinseparable
from the transactivatorfunction,
and since an EBNA-1 fusionprotein
bindsspecifically
tothe HVPori-P(C.
J.Sexton,
J.Griffith,
and J. S.Pagano, unpublished
data),
itseemsprobable
thatthetransactivator in HVP isanpCAT3tl
siro&.a
I
J. VIROL.
pAl
OCATCAT
on November 10, 2019 by guest
http://jvi.asm.org/
HVP ori-P 2881
A
\H
~o
0oc
<
U) < <: <
cr 0 0
aL aL a) a
N
- 0
00 c
4: Cc (D
0 2 2
C) CY) CO
Q - HQ
-cr 0 0 0
al a. aL a
0 0
< (D CD3
02
E2
C') CO) C)
> F-
F-) < < <:
mr
C0
03 0. a aL Ca
e
594
S
H erC HtC HC)
> 0 0 0
~~1
T
u 4 4 .4
a:
< a <a t:L Q. Q
3
B958
cl
o o
> Cz ~~~~~oC
0 Iw 0
> 0 C~( (0
Hn r- - H
,c < <: <
ca. a z
-LOUKES
C~ 0C"
o o
<;7
0(0C 0(0-'
4-li, --- < <
MV,a, e
'.111S
i,4
t
*.e.
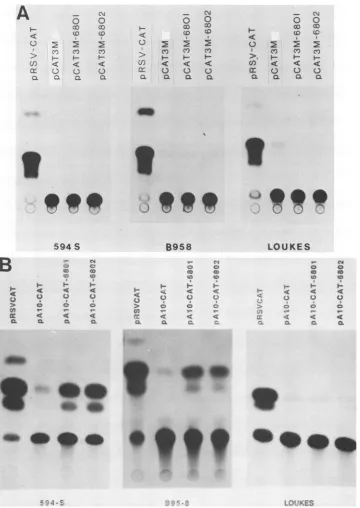
FIG. 5. HVPenhancer function shown through CAT expression in different cell lines.(A)Assayforpromoteractivity.pCAT3M680-1 and pCAT3M680-2weretransfected into the cell lines shown and assayed for CAT activity. (B)Assayfor enhancer activity.
pA1OCAT-680-1
andpA1OCAT-680-2
weretransfected into the cell lines shown and assayed for CAT activity. pRSV-CAT containstheCATgeneunder thecontrolofthe RSVenhancer.
pA1OCAT
contains the CATgeneunder the controloftheSV40earlypromoterbut lacks enhancersequences.EBNA-1-like protein. EBNA-1 is likely
tobe
alessefficient
transactivator of the HVP enhancer,
and there is evidence that the EBNA-1fusion protein binds
tothe HVP
ori-Pwith
a lower
specificity (Sexton
etal., unpublished data). This
decrease in
efficiency
maybe
accounted for by subtle
differences
inthe
binding domains of the
twoproteins
orby
minor variations in
the binding sites between the
twoviral
ori-Ps.
Although
theconsensusbinding
sequencefor
EBNA-1has been
defined
as a12-bp palindrome (33),
ourexamina-tion of the sequence of the repeat monomers revealed an
18-bp palindromic
sequence conserved between the two viruses. The12-bp EBNA-1-binding
sequence is located within this 18bp.
In
EBV, only
19bp
of therepeatmonomeris conserved in both the 5' and 3' repeat elements. This19-bp
sequence encompasses an18-bp palindrome
conserved in EBV and HVP. Conservation of the18-bp palindrome
inboth the EBV 3'dyad
and the HVPori-P
seems to indicate that thea_.
VOL.64, 1990
40
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.131.491.75.583.2]sequenceflanking the12-bp
EBNA-1-binding
site may play a role in recognition by EBNA-1 or an EBNA-1-like HVPprotein.
The different variations toleratedin the
repeat monomers within the HVPand EBVori-Ps
may correlatewith
the different binding andtransactivation efficiencies
of
the
twoviral proteins.
Thus, the HVP
ori-P
exhibits all of the structural and functional components of the EBV ori-P, although the initiation and termination points for DNA replication have yet tobe demonstrated in HVP.ACKNOWLEDGMENTS
This work was supported by Public Health Service grant 2POlCA19014 from the National Institutesof Health.
We thankTerri Snodgrassfor making two of the HVP enhancer constructs. We are grateful to Shannon Kenney and Nancy Raab-Traub for helpful discussions.
LITERATURE CITED
1. Adams, A., and T. Lindahl. 1975. Epstein-Barr virus genomes with properties of circularDNAmolecules incarrier cells. Proc. Natl. Acad. Sci. USA 72:1477-1481.
2. Baer, R., A. T. Bankier, M. D. Biggin, P. L. Deininger, P. J. Farrell, T. J. Gibson, G. Hatful, G. S. Stachwell, S. C. Hudson, C. Sequin, P.S. Tuffnell, and B. G. Barrell. 1984. Organization of the B95-8 Epstein-Barr virus genome. Nature (London) 310:207-211.
3. Bankier, A. T., K. M. Weston, and B. G.Barrell. 1987. Random cloning and sequencing by the M13/dideoxynucleotide chain termination method.Methods Enzymol. 155:51-93.
4. Chittenden, T., S. Lupton, and A. J. Levine. 1989. Functional limits of oriP, the Epstein-Barr virus plasmid origin of replica-tion. J. Virol.
63:3016-3025.
5. Colby, B., J. Shaw, G. B. Elion, and J. S. Pagano. 1980. The effect of acyclovir [9-(2-hydroxyethoxymethyl)guanine] on Ep-stein-Barr virus DNA replication. J. Virol.34:560-568. 6. Dillner, J., H. Rabin, N. Letvin, W. Henle, G. Henle, and G.
Klein. 1987. NuclearDNA-bindingproteins determined by the Epstein-Barr virus-related simian lymphotropic herpesviruses H. gorilla, H. pan, H. pongo and H. papio. J. Gen. Virol. 68:1587-1596.
7. Falk, L., F.Deinhardt, M.Nonoyama,L. G.Wolfe, C.Bergholz, B. Lapin, L. Yakovleva, V. Agrba, G. Henle, and W. Henle. 1976. Properties of ababoon lymphotropic herpesvirus related to Epstein-Barr virus.Int. J.Cancer18:798-807.
8. Gahn, T. A., and C. L. Schildkraut. 1989. The Epstein-Barr virus origin of plasmid replication, oriP, contains both the initiation and termination sites of DNA replication. Cell 58: 527-535.
9. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which express chloramphenicol acetyl-transferase in mammaliancells. Mol. Cell. Biol.2:1044-1051. 10. Heller, M., P. Gerber, and E. Kieff. 1981. Herpesvirus papio
DNA is similar in organization to Epstein-Barr virus DNA. J. Virol. 37:698-709.
11. Howe, J.G., and M.-D. Shu. 1988. Isolation and characteriza-tionof the genes for two small RNAs ofherpesvirus papio and their comparison with Epstein-Barr virus-encoded EBER RNAs. J. Virol. 62:2790-2798.
12. Jones, C. H., S. D.Hayward, andD. R.Rawlins. 1989. Interac-tion oflymphocyte-derived Epstein-Barr virus nuclear antigen EBNA-1 with itsDNA-binding sites. J. Virol.63:101-110. 13. Karlin,S.1986. Significant potential secondary structures in the
Epstein-Barr virus genome. Proc. Natl. Acad. Sci. USA 83: 6915-6919.
14. Karlin, S., and B. E. Blaisdell. 1987. Amodel for the develop-ment ofthe tandem repeat units in the EBV ori-Pregion and a discussion of their possiblefunction. J.Mol. Evol. 25:215-229. 15. Laimins, L. A., P. Gruss, R. R. Pozatti, and G. Khoury. 1984. Characterization of enhancer elements in the long terminal
repeat of Moloneymurinesarcomavirus. J.Virol. 49:183-189. 16. Laimins, L. A., G. Khoury, C. Gorman, B. Howard, and P. Cruss. 1982.Host-specific activation oftranscription by tandem repeats from simian virus 40 and Moloney murine sarcoma
virus. Proc. Natl.Acad. Sci.USA79:6453-6457.
17. Laux, G., M. Perricaudet, and P. J. Farrell. 1988. A spliced Epstein-Barr virus geneexpressed in immortalizedlymphocytes iscreated bycircularization of the linear viral genome. EMBO J.7:769-774.
18. Lee, Y.,M.Nonoyama, and H. Rabin. 1981. Colinear relation-ships ofherpesvirus papio DNA to Epstein-Barrvirus DNA. Virology110:248-252.
19. Lindahl, T., A. Adams, G. Bjursell, G. W. Bornkamm, C. Kascha-Dierich, and U. Jehn. 1976. Covalently closed circular duplex DNAofEpstein-Barr virus in humanlymphoidcell lines. J.Mol. Biol. 102:511-530.
20. Lupton, S., and A. Levine. 1985.Mappinggenetic elements of Epstein-Barr virus that facilitate extrachromosomalpersistence of Epstein-Barr virus-derived plasmids in human cells. Mol. Cell. Biol. 5:2533-2542.
21. Miller, G., T.Shope, H. Lisco, D.Stitt,and M. Lipman.1972. Epstein-Barr virus: transformation, cytopathic changes, and viral antigens in squirrel monkey and marmoset
leukocytes.
Proc.Natl. Acad. Sci. USA69:383-387.
22. Milman, G., and E. S. Hwang.1987.Epstein-Barrvirus nuclear antigen forms a complex that binds with
high
concentration dependence to a single DNA-binding site. J. Virol. 61:465-471.23. Neumann, E., M. Schaefer-Ridder, Y. Wang, and P. H. Hof-schneider. 1982. Gene transfersinto mouse lymphomacells
by
electroporation inhigh electric fields. EMBO J. 1:841-845. 24. Nilsson, K., E. Klein, G. Klein, W.Henle, andG. Henle. 1971.
Theestablishment oflymphoblastoid lines from adultand fetal human lymphoid tissue and its dependence on EBV. Int. J. Cancer8:443-450.
25. Nonoyama, M., and J. S. Pagano. 1971. Detection of
Epstein-Barr viral genome in non-productive cells. Nature
(London)
NewBiol. 233:103-106.
26. Nonoyama, M., andJ. S. Pagano. 1972.
Separation
ofEpstein-Barrvirus DNA from large chromosomal DNA in non-virus-producing cells. Nature (London)New Biol. 238:169-171. 27. Ohno, S., J. Luka, L.Falk, and G. Klein. 1977. Detectionofa
nuclear, EBNA-typeantigen in apparently
Epstein-Barr
virus-negativeHerpesvirus Papio(HVP)-transformed
lymphoid
lines by the acid-fixed nuclearbinding
technique.
Int. J. Cancer 21:941.28. Pagano, J. S. 1979. TheEpstein-Barr virus
plasmid,
p. 235-248. InD. J. Cummings,P.Borst,I. B.David, S.M.Weissman,
and C. F. Fox (ed.), Extrachromosomal DNA(ISBN
0-12-198780-9). Academic Press, Inc., New York.29. Pesano, R. L., and J. S. Pagano. 1986.
Herpesvirus
papio
containsaplasmidorigin ofreplicationthat actsin cis
interspe-cies withan Epstein-Barr virus
trans-acting
function. J. Virol. 60:1159-1162.30. Pesano, R. L., and J.S. Pagano. 1987.
trans-acting
functionthat maintains Epstein-Barr virus episomes actsinterspecies
on aherpesvirus papio putative
cis-acting
origin
ofreplication,
p. 397-414. In R. Gallo (ed.), Virusesand humancancer. Alan R. Liss, Inc., New York.31. Rabin, H., R. Neubauer, and R. Hopkins. 1978. Studies of Epstein-Barr (EBV)-like viruses of Old World nonhuman
pri-mates, p. 205-208. In P. Bentbelzen
(ed.),
Advances in compar-ative leukemia research. Elsevier BiomedicalPress,
Amster-dam.32. Rabin, H., R. H. Neubauer, R. F. Hopkins, E. K. Dzhikidze, Z. V. Shevtsova, and B. A. Lapin. 1977.
Transforming
activity
and antigenicity of an Epstein-Barr
(EBV)-like
virus from lymphoblastoid cell lines of baboons withlymphoid
disease. Intervirology 8:240-249.33. Rawlins, D., G. Milman, S. D. Hayward, and G. S. Hayward. 1985.Sequence-specificDNA-binding ofthe
Epstein-Barr
virus nuclear antigen (EBNA-1) to clustered sites in theplasmid
maintenanceregion. Cell42:859-868.
on November 10, 2019 by guest
http://jvi.asm.org/
HVP ori-P 2883 34. Reisman, D., and B. Sugden. 1986. trans activation of an
Epstein-Barr viraltranscriptional enhancer by the Epstein-Barr viral nuclear antigen 1. Mol. Cell. Biol. 6:3838-3846.
35. Reisman,D., J. Yates, and B.Sugden. 1985. Aputative originof replication of plasmids derived from Epstein-Barr virus is composed of two cis-acting components. Mol. Cell. Biol. 5: 1822-1832.
36. Sexton, C.J., and J.S.Pagano. 1989.Analysis of the Epstein-Barr virusorigin of plasmidreplication (oriP) revealsan areaof
nucleosome sparing thatspans the 3' dyad. J. Virol. 63:5505-5508.
37. Shaw, J., L. Levinger, and C. Carter, Jr. 1979. Nucleosomal
structureof Epstein-Barr virus DNA intransformed cell lines. J. Virol. 29:657-665.
38. Staden, R. 1982. Automation of the computerhandling ofgel reading dataproduced by the shotgun method of DNA
sequenc-ing. Nucleic Acids Res. 10:4731-4751.
39. Wysokenski, D. A., and J. L. Yates. 1989. Multiple EBNA1-binding sites are required to form an EBNA1-dependent
en-hancer andtoactivateaminimalreplicative origin within oriP of
Epstein-Barr virus. J. Virol. 63:2657-2666.
40. Yates, J., N. Warren, D. Reisman, and B. Sugden. 1984. A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infected cells. Proc. Natl. Acad. Sci. USA 81:3806-3810. 41. Yates,J., N. Warren, and B.Sugden. 1985. Stablereplication of
plasmids derived from Epstein-Barr virus in various mammalian cells. Nature(London) 313:812-815.
VOL.64, 1990