JOURNAL OFVIROLOGY, Feb. p. Vol. 64, No. 2 0022-538X/90/020890-12$02.00/0
CopyrightC 1990, AmericanSociety forMicrobiology
Molecular
Characterization
of
an
Attenuated
Human
Immunodeficiency Virus Type
2 Isolate
PRASANNA
KUMAR,'
HUXIONGHUI,'
JOHN C.KAPPES,1
BETH S. HAGGARTY,2JAMES A. HOXIE,2 SURESH K.ARYA,3 GEORGE M. SHAW,"5 ANDBEATRICE H.HAHN'.4*
Department of
Medicine,'
Departmentof Microbiology,4 andDepartmentof Biochemistry,5 University of Alabama atBirmingham, University Station, Birmingham,Alabama35294; Hematology andOncology Section,Department of
Medicine, Hospital ofthe University of
Pennsylvania,
Philadelphia, Pennsylvania191042;
andLaboratory ofTumor Cell Biology, National Cancer Institute, Bethesda, Maryland208953
Received13July1989/Accepted6October 1989
Naturallyoccurring strains of humanimmunodeficiencyvirus(HIV)canvaryconsiderably in their invitro biological properties,and such differencesmayalso be reflected in their in vivopathogenesis.In anattempt to definegenetic determinants of viralpathogenicity, wehavemolecularly cloned, sequenced,andcharacterized anattenuated isolate of HIV type2(HIV-2/ST)that differs fromprototypeHIV-2 strains in itsinabilitytofuse with and kill susceptible CD4-bearing target cells. A proviral clone, termed JSP4-27, was identified to be transfection competent and to fully exhibit the noncytopathic and nonfusogenic properties of its parental isolate. Nucleotide sequence analysis of this clone revealed a genomic organization very similar to that of cytopathic HIV-2 strains and an overallnucleotide sequence homology of 88 to 90%. Amino acid sequence comparison confirmedtheintegrityof allmajorviralgeneproductsinJSP4-27but identified two aminoacid sequence substitutions in itsenvelope fusion region. Toinvestigate whether these mutations wereresponsible forthenonfusogenic phenotype ofJSP4-27, weamplffied,cloned,andsequenced theenvelopefusionregions of four additional HIV-2/ST strains, two of which representedin vitro-generated, fusogenicand cytopathic variants of HIV-2/ST. The analysis showed that all HIV-2/ST strains examined, including the fusogenic variants, contained thesameamino acid sequencechanges.On the basis of thesefindings,weconclude thatthe attenuatedphenotypeofJSP4-27, and that of its parental virus,is not due to a direct alteration of theenvelope fusion domain. Our results alsoshow, for the firsttime,thatindividualreplication-competent proviralclones canberepresentative ofattenuatedstrains ofHIV.
Humanimmunodeficiency virus types 1 and 2 (HIV-1 and HIV-2) represent two distinct groups of HIV known to cause
acquired immunodeficiency syndrome (AIDS) in infected individuals (3, 11, 12, 21, 40). Whereas HIV-1 is the caus-ative agentof epidemic AIDSworldwide,HIV-2 appears to be geographically restricted to West Africa (4, 7, 13, 26, 29). Numerous isolates of HIV-1 and HIV-2 have been obtained, and their biological and molecular properties have been
characterized (1, 3, 11, 22, 40, 51). Nucleotide sequence
analysisshows that HIV-2 is only distantly related to HIV-1
(22) and ismoreclosely related to two primate retroviruses,
simian immunodeficiency virus (SIV) strains SIVMAC and
SIVSM, which cause an AIDS-like disease in captive macaques(8, 14, 25). Although genetically divergent, proto-type HIV-1 and HIV-2 isolates have very similar biological properties, including a propensity for rapid genetic change
(17, 42, 51), a similar host cell tropism, a considerable cytopathic effect on T-cell cultures and peripheral blood mononuclear cells in vitro, and the ability to form syncytia with CD4-bearing target cells (11, 40). In fact, the majority of HIV-1 and HIV-2 strains isolated from patients with
immu-nodeficiency disease have been shown to cause cell fusion and the formation of multinucleated giant cells in culture. This represents a hallmark of productive viral infection and accountsfor the profound cytopathic effect of HIV in vitro
(23, 37, 49).
In contrast to these viruses, we and others have recently
isolated less pathogenic strains of HIV-1 and HIV-2 that exhibit markedly different biological properties (2, 9, 16, 32,
*Correspondingauthor.
34,47). Theseparticular isolatescauselittle or nocelldeath
in susceptible target cells, fail to induce cell fusion with
CD4-bearing immortalized T-cell lines, exhibit arestricted host cell tropism with a preference for peripheral blood
mononuclear cells or macrophages, and are often derived fromasymptomatic individuals. Although their in vitro
bio-logical differencesarewelldocumented, thegeneticchanges
responsible for their attenuated phenotype are not under-stood. Toelucidate determinantsof HIV pathogenicity, we have begun to molecularly dissect a previously reported,
nonfusogenic and noncytopathicHIV-2isolate, termed
HIV-2/ST,thatwasobtainedfromahealthy Senegaleseprostitute (32). Although this virus replicated to high titers in tissue
culture, itinfectedcellsat aslowerratethandid cytopathic
strainsofHIV-1and HIV-2andcaused little or no cell killing and fusion. This was the case despite the fact that its externalenvelope glycoproteinwascleaved correctly,
trans-portedto the cell surface, and shown to bind to a specific
epitope onCD4, which was recognized by OKT4a but not OKT4antibodies(32). HIV-2/STtherefore appeared to bind totheCD4 molecule analogous to other HIVs, but it failed to
fusewithCD4-bearing target cells, suggesting that its
infec-tivitywasgreatly retarded at the level of cell entry (32).
Since HIVisolates generally represent complex mixtures of genotypically distinct viruses and since the biological
phenotype ofany HIV culture depends on the sum of the
properties of each genotypic variant (21a, 42), we first
attempted to isolate amolecularclone that was both
trans-fectioncompetentandrepresentative of the in vitro proper-ties ofits parental virus. We therefore obtained three
full-length proviral clones (AJSP4-27, AJSP4-32, and XJSP4-34) 890
on November 10, 2019 by guest
http://jvi.asm.org/
A Xbal Xbal
J1XI
Xbal
Xbal
---atil X I digest Z
=I_ S mmY000-////m IAI -_
partialXbaldigest subcloninginto pSP65
(Xbal)
Xbal Xbal
J1A
XJSP4-27
(Xbal) Xbal
!o
pJSP4-27B
Syncytia Formation Assay C Western Blot
JSP SL1
r* ; wr (
I
t
*O L^. $* , fHIV-2/ST
pJSP4-27 HIV-2/RODpSL1
gp43-_ .-.
_0'-gp3
z p24 -wi-I
-p24human
anti-HIV-2serum
FIG. 1. (A)Construction ofareplication-competent HIV-2/ST plasmid clone. AJSP4-27 (32) was partially cleaved withXbaItoremove bacteriophagearms (double lines) andflanking cellular sequences (hatched lines), and the resulting 14-kilobase-pair provirus-containing fragmentwassubsequentlysubclonedinto pSP65. (B and C) Biological comparison of HIV-2/ST- andHIV-2/ROD-derived,genetically pure viral strains. CEMx174cultures,productivelyinfected(>90%)withJSP4-27 (HIV-2/ST) andSL1 (HIV-2/ROD),respectively, wereexamined in syncytium formation assays and by Western blot analysis. (B) No syncytium formation was observed upon cocultivation of JSP4-27-producingCEMx174 cells with uninfected CEM cells,whereas numerous andlarge syncytia were generated uponcocultivation of thesameuninfected CEM cells with SL1-infected CEMx174cells (identicalresults were obtained with H9, SupTl, andCEMx174cells). Syncytium formation was monitored 18 h after cocultivation. (C) Western blot analysis of cell-free virions derived from these same transfection-derived cultures demonstrated differencesinthe sizes ofthe JSP andSL1transmembraneenvelope glycoproteins.
from a genomic library of a biologically cloned high-pro-ducercellline,termedST/B12,andsubsequentlytransfected them into the neoplastic T-cell lines SupTl (46) and CEMx174 (27, 44). Reverse transcriptase activity was de-tected in supernatantsofcultures transfected with XJSP4-27 asearly as 5days posttransfection, whereas XJSP4-32-and
AJSP4-34-transfected cultures revealed no signs of viral replication, indicatingthat theseproviruseswerereplication defective. Immunofluorescence analysis further confirmed the presence of virus-expressing cells in AJSP4-27-trans-fected cultures but failed to
identify
virus-mediated cellfusion.Westernblot
(immunoblot)
analysis
ofpurified
JSP4-27 virions demonstrated a protein profile similar tothat of theparental HIV-2/STvirus. Tofacilitatesubsequent trans-fection experiments and to allow the direct
comparison
of JSP4-27 with othertransfection-competent
HIV-2plasmid
constructs,wesubclonedthe
proviral
insert of XJSP4-27 into theplasmidvectorpSP65(Fig. 1A).To test whether the transfection-derived JSP4-27 virions wereinfectious,filtered supernatantsof
plasmid-transfected
cultures were transmitted to uninfected
SupTl
andCEMx174 cells. The results showed thatcell-free transmis-sionofJSP4-27virionswasreadilyandreproducibly demon-strable. However, infection and spreadin culture, particu-larlyin SupTl cells,occurred slowlyand with considerable
delay. These results were confirmed and extended in
com-parative studies with a transfection-derived,
cytopathic
HIV-2/ROD strain, termed SL1 (36). Whereas transfection
of the SLi
provirus
resulted in >90% infectedSupTl
or CEMx174 cultures within 3 to 4days
posttransfection,
JSP4-27-transfectedcultures reachedonly 10%infectivity
in the same time period, which indicated the same reducedability to spread in culture that had been observed for the
parental HIV-2/ST strain (32).
Similarly,
the transfection-derived JSP4-27 cultures did notformsyncytia
upon cocul-tivation with severalCD4-bearing
T-celllines,
including
SupTl, CEM, H9, andCEMx174,
whereas SLi-infectedcultures
produced
numerousandlarge syncytia,
aswellas aprofound
cytopathic
effect,
with thesesametargetcells(Fig.
1B). These datathus showed that the JSP4-27provirus
wasreplicationcompetentandinfectiousandexhibitedthesame
NOTES 891
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.94.526.68.417.2]892 NOTES
-->R... . . 1~~~~~~~~~~~~~~~~~~~~~~~~~~I00.
AGTCGCTCTGCGGAGAGGCTGGCAGATTGAGCCCTGGGAGGTTCTCTCCAGCACTAGCAGGTAGAGCCTGGGTGTTCCCTGCTAGACTCTCACCAGTGCTTGGCCGGCACTGGGCAGACG
R<---->U5. . 200 . .GCTCCACGCTTGCTTGCTTAAAAGACCTCTTAATAAAGCTGCCAGTTAGAAGCAAGTTAAGTGTGTGCTCCCATCTCTCCTAGTCGCCGCCTGGTCATTCGGTGTTCATCTAAAGTAACA
U5 <--I PBS . .
AGACCCTGGTCTGTTAGGACCCTTTCTGCTTTGGGAAACCAAGGCAGGAAAATCCCTAGCAGGT,TGGCGCCCGAACAGGGACTTGAAGAAGACTGAGAAGCCTTGGAACACGGCTGAGTG
400 , *
AAGGCAGTAAGGGCGGCAGGAACAAACCACGACGGAGTGCTCCTAGAAAAGCGCAGGCCGAGGTACCAAGGGCGGCGTGTGGAGCGGGAGTGAAAGAGGCCTCCGGGTGAAGGTAAGTGC
500 . 600
CTACACCAAATACAGTAGCCAGAAGGGCTTGTTATCCTACCTTTAGACGGGTAGAAGATTGTGGGAGATGGGCGCGAGAAACTCCGTCTTGAGAGGGAAAAAAGCAGACGAATTAGAAAA
gag >
MetGlyAlaArgAsnSerValLeuArgGlyLysLysAlaAspGluLeuGluLy
700GATTAGGTTACGGCCCGGCGGAAAGAAAAAATATAGGCTAAAACATATTGTGTGGGCAGCGAATGAATTGGACAGATTCGGATTGGCAGAGAGCCTGTTGGAGTCAAAAGAGGGTTGCCA
sIleArgLeuArgProGlyGlyLysLysLysTyrArgLeuLysHisIeValTrpAlaAlaAsnGluLeuAspArgPheGlyLeuAlaGluSerLeuLeuGluSerLysGluGlyCysGI
800
9 00 . .
AAAAATTCTTACAGTTTTAGATCCATTAGTACCGACAGGGTCAGAAAATTTAAAAAGCCTTTTTAATACTGTCTGCGTCATTTGGTGTATACACGCAGAAGAGAAAGCGAAAGATACTGA
nLysIleLeuThrValLeuAspProLeuValProThrGlySerGluAsnLeuLysSerLeuPheAsnThrValCysValIleTrpCysIleHisAlaGluGluLysAiaLysAspThrG1
900
AGAAGCAAAACAAAAGGTACAGAGACATCTAGTGGCAGAAACAAAAACTACAGAAAAAATGCCAAGTACAAGTAGACCAACAGCACCACCTAGCGGGAACGGAGGAAACTTCCCCGTACA
uGluAlaLysGlnLysValGlnArgHisLeuValAlaGluThrLysThrThrGluLysMetProSerThrSerArgProThrAlaProProSerGlyAsnGlyGlyAsnPheProValG1
11000
ACAAGTGGCCGGCAACTATACCCATGTGCCACTAAGTCCCCGAACCCTAAATGCTTGGGTAAAACTAGTAGAGGAAAAGAAGTTCGGGGCAGAAGTAGTGCCAGGATTTCAGGCACTCTC
nGlnValAlaGlyAsnTyrThrHisValProLeuSerProArgThrLeuAsnAlaTrpValLysLeuValGluGluLysLysPheGlyAlaGluValValProGlyPheGlnAlaLeuSe
.1100 . . . .150 . . .10
AGAAGGCTGCACGCCCTATGATATTAATCAAATGCTTAATTGTGTGGGCGACCATCAAGCAGCTATGCAAATAATCAGGGAAATTATTAATGAAGAAGCAGCAGATTGGGACGCACAACA
rGlu'GlyCysThrProTyrAspIleAsnGlnMetLeuAsnCysVa1GlyAspHisGlnAlaAlaMetGlnIleIleArgGlulleIleAsnGluGluAlaAlaAspTrepAs'AuGllnYHi
1300
CCCAATACCAGGCCCCTTACCAGCGGGGCAGCTCAGGGAGCCAAGGGGATCTGACATAGCAGGGACAACAAGCACAGTAGAAGAGCAGATCCAGTGGATGTTTAGGCCACAAAATCCTGT
sProIleProGlyProLeuProAlaGlyGlnLeuArgGluProArgGlySerAspIleAlaGlyThrThrSerThrValGluGluGlnlleGlnTrpMetPheArgProGlnAsnProVa
24000
.. . . . .~~210 . 140 ...
ACCAGTAGGAAGCATCTATAGAAGATGGATCCAGATAGGGCTACAGAAGTGCGTCAGGATGTACAACCCAACCAACATCCTAGACATAAAACAGGGACCAAAGGAGCCATTCCAGAGTTA
lProValGlySerIleTyrArgArgTrpIleGGnIleGlyLeuGlnLysCysValArgMetTyrAsnProThrAsnIleLeuAspIleLysGlnGlyProLysGluProPheGlnSerTy
2200 .~150
. 150. . 2500.
TGTAGATAGATTCTACAAGAGCTTGAGGGCAGAACAAACAGATCCAGCAGTAAAAAATTGGATGACCCAAACACTGCTAGTGCAGAATGCCAACCCAGACTGTAAGTTAGTACTAAAAGG
rValAspArgPheTyrLysSerLeuArgAlaGluGlnThrAspProAlaValLysAsnTrpMetThrGlnThrLeuLeuValGlnAsnAlaAsnProAspCysLysLeuValLeuLysG1
. 01600. 2
ACTAGGGATAAATCCTACCTTAGAAGAAATGCTAACCGCCTGTCAGGGGGTAGGTGGACCAGGCCAGAAAGCCAGATTAATGGCAGAAGCCTTAAAGGAGGCCATGGCACCAGCCCCCAT
yLeuGlyIleAsnProThrLeuGluGluMetLeuThrAlaCysGlnGlyValGlyGlyProGlyGlnLysAlaArgLeuMetAlaGluAlaLeuLysGluAlaMetAlaProAlaProI1
1
1700 ...1800
CCCATTTGCAGCAGCCCAACAGAGAAGGACAATTAAGTGCTGGAATTGCGGAAAGGAAGGGCACTCGGCAAGACAATGCCGAGCACCTAGAAGACAAGGCTGCTGGAAATGTGGCAAGGC
pol> LysThrArgLeuLeuGluMetTrpGlnGly
eProPheAlaAlaAlaGlnGlnArgArgThreIeLysCysTrpAsnCysGlyLysGluGlyHisSerAlaArgGlnCysArgAlaProArgArgGlnGlyCysTrpLysCysGlyLysAi
1900AGGACACATCATGGCAAAATGCCCAGAAAGACAGGCGG GTTTTTTAGGGTTGGGCCCATGGGGAAAGAAGCCCCGCAATTTC CCTGTGGCCCAAATC CCGCAGGGGCTGACACCAACAGC
ArgThrHisHisGlyLysMetProArgLysThrGlyGlyPhePheArgValGlyProMetGlyLysGluAlaProGlnPheProCysGlyProAsnProAlaGlyAlaAspThrAsnSer
aGlyHisIleMetAlaLysCysProGluArgGlnAlaGlyPheLeuGlyLeuGlyProTrpGlyLysLysProArgAsnPheProValAlaGlnlleProGlnGlyLeuThrProThrA1
... . . ~~~~~~2000~ ~...~ ~ ~ ~
ACCCCCGATAGACCCAGTAGAGGACCTACTAGAGAAGTACATGCAGCAAGGGAAAAGGCAGAGAGAGCAGAGAGAGAGGCCATACAAAGAAGTGACAGAGGACTTCCTGCAGCTCGAGAA
ThrProAspArgProSerArgGlyProThrArgGlUValHisAlaAlaArgGluLysAlaGluArgAlaGluArgGluAlaIleGlnArgSerAspArgGlyLeuProAlaAlaArgGlu
aProProIleAspProValGluAspLeuLeuGluLysTyrMetGlnGInGlyLysArgGInArgGluGInArgGluArgProTyrLysGluValThrGluAspPheLeuGlnLeuGluLy
.. . . . ~~~~~~2100~ ~...~
ACAAGAGACACCATGCAGAGAGACGACAGAGGACTTGCTGCACCTCAATTCTCTCTTTGGAAAAGACCAGTAGTCACAGCACATGTTGAGGGCCAGCCAGTAGAAGTTTTGCTAGACACA
ThrArgAspThrMetGlnArgAspAspArgGlyLeuAlaAlaProGlnPheSerLeuTrpLysArgProValValThrAlaHisValGluGlyGlnProValGluValLeuLeuAspThr
sGlnGluThrProCysArgGluThrThrGluAspLeuLeuHisLeuAsnSerLeuPheGlyLysAspGln***
. . . ~~~~~2200
GGGGCTGACGACTCAATAGTAGCAGGCGTAGAGTTAGGGAGCAATTATAGTCCAAAGATAGTAGGGGGAATAGGGGGATTCATAAATACCAAAGAATATAAAAATGTAGAAATAAGAGTA
GlyAlaAspAspSerIleValAlaGlyValGluLeuGlySerAsnTyrSerProLysIleValGlyGlyIleGlyGlyPheIleAsnThrLysGluTyrLysAsnValGluIleArgVae
2
230 0 .,...20024
TTAAATAAAAGAGTAAGAGCCACCATAATGACAGGTGATACCCCAATCAACATTTTTGGCAGAAACATTCTGACAGCCTTAGGCATGTCATTAAATCTACCAGTCGCCAAGATAGAACCA
LeuAsnLysArgValArgAlaThrIleMetThrGlyAspThrProu leAsnIlePheGlyArgAsnIleLeuThrAlaLeuGlyMetSerLeuAsnLeuProValAlaLysIleGluPro
... . . ~~~~~~~~~~~~~~2500~. ~
ATAAAAATAATGCTGAAGCCAGGAAAGGATGGACCAAAACTGAGACAATGGCCCTTAACAAAAGAAAAAATAGAGGCACTAAAAGAGATCTGTGAGAAAATGGAAAGAGAGGGCCAGCTA IleLysIleMetLeuLysProGlyLysAspGlyproLysLeuArgGlnTrpProLeuThrLysGluLysIleGluAlaLeuLysG(lrleCysGluLysMetGluArgGluGlyGlnLeu
.~~~~...200~ ~ ~ ~ ~ ~ ~ ~ ~ ~~~~~ ~6 ..~ ~ ~ ~~~~~~~~
GAGGAGGCACCTCCAACTAATCCTTATAATACCCCCACATTTGCAATCAAGAAAAAGGACAAAAACAAATGGAGAATGCTAATAGATTTTAGAGAACTAAACAAGGTAACTCAAGACTTC GluGluAlaProProThrAsnProTyrAsnThrProThrPheAlaIleLysLysLysAspLysAsnLysTrpArgMetLeuIleAspPheArgGluLeuAsnLysValThrGlnAspPhe
FIG. 2. Completenucleotide sequence of the HIV-2/ST proviralgenome. Shownare9,672bpofnucleotide sequence and thededuced amino acid sequencesof thecorrespondingviralproteins.The sequencestarts atthe 5'capsiteand ends with the 3' polyadenylationsite of theviral RNA. Theprimer-bindingsite(PBS;complementarytotheLys-tRNA),polypurinetract(PPT),andshortinvertedrepeatsthat
flank
the LTRsare underlined. Core enhancer sequences(E), Spl-bindingsites(Spl),theTATAbox(TATAA),and thepolyadenylation
signal
(AATAAA)areshown. The U3-Rand R-U5boundaries,aswellasthesplicedonor(SD)andspliceacceptor(SA) sites,havebeendetermined inanalogywithHIV-2/ROD(22).Thevpropenreadingframecontainsaprematurein-frame TAAstopcodonatposition5777(***).Sequence analysiswasperformedbythechemicaldegradationmethod of Maxam andGilbert(38)aswellasbythedideoxynucleotide-chaintermination method ofSangeretal.(45).ThenucleotidesequenceofJSP4-27 has beensubmittedtotheAIDSSequenceDataBase,LosAlamosNational Laboratories,aswellas totheGenBankandEMBLlibraries.
nonfusogenic
andnoncytopathic properties
aspreviously
9,672 basepairs (bp) long
and exhibits an overallgenomic
described for the
parental
virus(32).organization
of 5'long
terminal repeat(LTR)-gag-pol-cen-Having
identified andcharacterized thebiological
features tralregion-env-nef-3'
LTR,which isidenticaltothatof other of a molecular clone ofHIV-2/ST, we nextsequenced
itscytopathic
HIV-2 andSIVMAC proviruses.
It contains all entire genome. Thecomplete
nucleotide sequence of themajor
openreading
framescharacteristic forHIV-2, includ-JSP4-27provirus
isdepicted
inFig.
2. The viral genome ising
vpx, which is present in HIV-2 andSIVMAC
but not inon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.65.558.66.572.2]* . . . . 2700 . . .
ACAGAAATCCAGTTAGGAATTCCACACCCAGCAGGACTAGCCAAGAAGAAACGAATTACTGTCCTAGATGTAGGGGATGCTTACTTTTCCATACCACTACATGAGGATTTTAGACAGTAT
ThrGlulleGlnLeuGlyIleProHisProAlaGlyLeuAlaLysLysLysArgIleThrValLeuAspValGlyAspAlaTyrPheSerIleProLeuHisGluAspPheArgGlnTyr
* . . 2800 . . . .
ACTGCATTTACTCTACCATCAATAAACAATGCTGAACCAGGAAAAAGATACATATATAAAGTCTCACCACAGGGATGGAAGGGATCACCAGCAATTTTTCAGTACACAATGAGGCAGGTC ThrAlaPheThrLeuProSerIleAsnAsnAlaGluProGlyLysArgTyrIleTyrLysValSerProGlnGlyTrpLysGlySerProAlaIlePheGlnTyrThrMetArgGlnVa1
* 2900 . . . 2900 3000
TTAGAACCATTCAGAAAAGCAAACCCGGATATCATTCTCATTCAGTACATGGATGATATCTTGATAGCCAGCGACAGGACAGATTTAGAACATGACAGAGTGGTTCTGCAGCTAAAGGAA LeuGluProPheArgLysAlaAsnProAspIleIleLeuIleGlnTyrMetAspAspIleLeuIleAlaSerAspArgThrAspLeuGluHisAspArgValValLeuGlnLeuLysGlu
* . . . 3100
CTTCTAAATGGCCTGGGATTTTCCACCCCAGATGAGAAGTTCCAAAAAGACCCTCCATACCAATGGATGGGCTATGAACTGTGGCCAACTAAATGGAAGCTGCAAAGAATACAATTGCCC LeuLeuAsnGlyLeuGlyPheSerThrProAspGluLysPheGlnLysAspProProTyrGInTrpMetGlyTyrGluLeuTrpProThrLysTrpLysLeuGlnArgIleGlnLeuPro
* . . . 3200 . .
CAAAAGGAAGTATGGACAGTCAATGACATCCAAAAACTGGTGGGTGTCCTAAATTGGGCAGCACAAATCTACCCAGGGATAAAGACCAGAAACTTATGTAGGTTAATCAGAGGAAAAATG GlnLysGluValTrpThrValAsnAspIleGlnLysLeuValGlyValLeuAsnTrpAlaAlaGlnIleTyrProGlyIleLysThrArgAsnLeuCysArgLeuIleArgGlyLysMet
** . ~~~~~~~~3300***~ ~
ACACTCACAGAAGAGGTACAGTGGACAGAATTAGCAGAAGCGGAACTAGAAGAAAACAAAATCATCTTAAGCCAGGAACAAGAAGGATGCTATTACCAAGAGGAAAAGGAGCTAGAAGCA ThrLeuThrGluGluValGlnTrpThrGluLeuAlaGluAlaGluLeuGluGluAsnLysIleIleLeuSerGlnGluGlnGluGlyCysTyrTyrGlnGluGluLysGluLeuGluAla
* . ~~~~~3400 .*****
* . 3 0 . . . .370
ACAGTCCAAAAAGATCAAGACAATCAGTGGACATATAAGATACACCAGGGAGGAAAAATTCTAAAAGTAGGAAAATATGCAAAGGTAAAAAATACCCACACCAACGGAGTCAGACTCCTA ThrValGlnLysAspGlnAspAsnGlnTrpThrTyrLysIleHisGlnGlyGlyLysIleLeuLysValGlyLysTyrAlaLysValLysAsnThrHisThrAsnGlyValArgLeuLeu
3500
..3.. . . . 38600.
GCACAAGTAGTTCAAAAAATAGGAAAAGAAGCACTAGTCATTTGGGGAC-GAATACCAAAATTTCACCTACCAGTAGAAAGAGATACCTGGGAACAGTGGTGGGATAACTACTGGCAAGTG
AlaGlnValValGlnLysIleGlyLysGluAlaLeuValIleTrpGlyArgIleProLysPheHisLeuProValGluArgAspThrTrpGluGlnTrpTrpAspAsnTyrTrpGlnVa1
* . . 4000 . . . .3700
ACATGGATCCCAGACTGGGACTTCATATCTACCCCGCCACTGGTCAGATTAGTATTTAACCTGGTGAAAGATCCCATACTAGGCGCAGAAACCTTCTACACAGATGGATCCTGCAATAAG
ThrTr,pIleProAspTrpAspPheIleSerThrProProLeuValArgLeuValPheAsnLeuValLysAspProIleLeuGlyAlaGluThrPheTyrThrAspGlySerCysAsnLys
* . .*. . . .*380043008.
CAATCAAGAGAAGGAAAAGCAGGATACATAACAGATAGAGGAAGAGACAAGGTGAGGCTATTAGAGCAAACCACCAATCAGCAAGCAGAATTAGAAGCCTTTGCGATGGCAGTAACAGAC
GlnSerArgGluGlyLysAlaGlyTyrIleThrAspArgGlyArgAspLysValArgLeuLeuGluGlnThrThrAsnGlnGlnAlaGluLeuGluAlaPheAlaMetAlaValThrAsp
* . . . 3900. .00
TCAGGTCCAAAGGCCAACATTATAGTAGACTCACAATATGTAATGGGAATAGTAGCAGGCCAACCAACAGAGTCAGAGAGTAAAATAGTAAATCAAATCATAGAAGAAATGATAAAAAAG SerGlyProLysAlaAsnIleIleValAspSerGlnTyrValMetGlyIleValAlaGlyGlnProThrGluSerGluSerLysIleValAsnGlnIleIleGluGluMetIleLysLys
* *400. 4500
4. . 46000 . .
* 4700 . . . 4800
CAGGAGGAACATGAAAAATATCATAGCAATGTAAAAGAACTATCCCATAAATTTGGACTGCCCAAATTAGTGGCAAGACAAATAGTAAACACATGCACCCAATGTCAGCAGAAAGGGGAG
GlnGluGluHisGluLysTyrHisSerAsnValLysGluLeuSerHisLysPheGlyLeuProLysLeuValAlaArgGlnIleValAsnThrCysThrGlnCysGlnGlnLysGlyGlu
* . . . 4300
GCTATACATGGGCAAGTAAATGCAGAATTAGGCACTTGGCAAATGGACTGCACACACTTAGAAGGAAAAATCATTATAGTAGCAGTACATGTTGCAAGTGGATTTATAGAAGCAGAAGTC
AlaIleHisGlyGlnValAsnAlaGluLeuGlyThrTrpGlnMetAspCysThrHisLeuGluGlyLysIleIleIleValAlaValHisValAlaSerGlyPheIleGluAlaGluVa1
* . . . 54400 .
ATCCCACAGGAATCAGGAAGGCAAACGGCACTCTTCCTACTAAAACTGGCCAGTAGGTGGCCAATAACACATTTGCACACAGACAATGGTGCCAACTTCACTTCACAGGAAGTAAAGATG
IleProGlnGluSerGlyArgGlnThrAlaLeuPheLeuLeuLysLeuAlaSerArgTrpProIleThrHisLeuHisThrAspAsnGlyAlaAsnPheThrSerGlnGluValLysMet
* . . . . 4500 . . .
GTGGCATGGTGGATAGGTATAGAACAATCCTTCGGAGTACCTTACAATCCACAAAGCCAAGGAGTAGTGGAAGCAATGAATCACCACCTAAAAAATCAGATAAGCAGAATTAGAGAGCAG
ValAlaTrpTrpIleGlyIleGluGlnSerPheGlyValProTyrAsnProGlnSerGlnGlyValValGluAlaMetAsnHisHisLeuLysAsnGlnlleSerArgIleArgGluGln
* 5300 . 4600 . . . 5
GCAAACACAGTAGAAACAATAGTACTAATGGCAGTTCATTGCATGAATTTTAAAAGGAGGGGAGGAATAGGGGATATGACCCCAGCAGAAAGACTAATCAATATGGTCACTGCAGAACAG
AlaAsnThrValGluThrIleValLeuMetAlaValHisCysMetAsnPheLysArgArgGlyGlyIleGlyAspMetThrProAlaGluArgLeuIleAsnMetValThrAlaleGlGn
44700 . .F 2-Continued800. 4 GAAATACAATTCCTCCAAGCAAAAAATTCAAAATTACAAAATTTTCGGGTCTATTTCAGAGAAGGCAGAGATCAGCTGTGGAAAGGACCTGGGGAACTACTGTGGAAGGGGGACGGAGCA
GluIleGlnPheLeuGlnAlaLysAsnSerLysLeuGlnAsnPheArgValTyrPheArgGluGlyArgAspGlnLeuTrpLysGlyProGlyGluLeuLeuTrpLysGlyAspGlyAla
... . . ~~~~~~~~~~~~~4900~ ~. ~ GTCATAGTCAAGGTAGGGGCTGACATAAAAATAATACCAAGAAGGAAAGCTAAGATCATCAAAGACTATGGAGGAAGGCAAGAGATGGATAGCGGTTCCAACTTGGAGGGTGCCAGGGAG
ValIleValLysValGlyAlaAspIleLysIleIleProArgArgLysAlaLysIleIleLysAspTyrGlyGlyArgGlnGluMetAspSerGlySerAsnLeuGluGlyAlaArgGlu
vif > MetGluGluGlyLysArgTrpIleAlaValProThrTrpArgValProGlyAr
... . . ~~~~~~~~5000...~ ~ ~ ~
GATGGAGAGGTGGCATAGCCTTATCAAGTATCTAAAATACAGAACAGGAGATCTAGAGAAGGTGTGCTATGTTCCCCACCATAAGGTGGGATGGGCGTGGTGGACTTGCAGCAGGGTAAT
AspG1yG1uVa1A1a***
gMetGluArgTrpHisSerLeuIleLysTyrLeuLysTyrArgThrGlyAspLeuGluLysValCysTyrValProHisHisLysValGlyTrpAlaTrpTrpThrCysSerArgValIo
.. . . . ~~~~~~5100~ ~...~
ATTCCCATTAAAAGGAGAAAGTCATCTGGAGATACAGGCATACTGGAACCTAACACCAGAAAAAGGATGGCTCTCCTCCTATTCAGTAAGACTAACTTGGTATACAGAAAAATTCTGGAC
ePheProLeuLysGlyGluSerHisLeuGluIleGlnAlaTyrTrpAsnLeuThrProGluLysGlyTrpLeuSerSerTyrSerValArgLeuThrTrpTyrThrGluLysPheTrpTh
AGATGTTACCCCAGACTGTGCGGACTCCCTAATACATAGCACTTATTTCTCTTGCTTTACGGCAGGCGAAGTAAGAAGAGCCATCAGAGGGGAAAAGCTATTATCCTGCTGCAACTACCC
rAspValThrProAspCysAlaAspSerLeuIleHisSerThrTyrPheSerCysPheThrAlaGlyGluValArgArgAlaIleArgGlyGluLysLeuLeuSerCysCysAsnTyrPr
553 00 . . 5004
CCAAGCCCATAAGTACCAGGTACCGTCACTCCAGTTTCTGGCCTTAGTGGTAGTGCAACAAAATGGCAGGCCCCAGAGAGACAATACCACCAGGAAACAGTGGCGAAGAAACTATCGGAG
oGlnAlaHisLysTyrGlnValProSerLeuGlnPheLeuAlaLeuValValValGlnGlnAsnGlyArgProGlnArgAspAsnThrThrArgLysGlnTrpArgArgAsnTyrArgAr
vpx > MetAlaGlyProArgGluThrIleProProGlyAsnSerGlyGluGluThrIleGlyG
FIG. 2-Continued.
HIV-1 (18, 24, 30, 50),and vpr, which ispresent in HIV-1, similar
length,
and therearenomajor
deletionsorinsertionsHIV-2,and
SIVMAc
butnotinSIVAGM
(19).Like other HIV thatwoulddistinguish
itfrom the LTRs of othercytopathic
and SIV
proviruses,
HIV-2/ST is flankedby
LTRsequences HIV-2 strains (datanotshown).thatareknown to
regulate
viral geneexpression. Sequence
Comparison
of the deduced amino acid sequences of thecomparison
with other HIV-2 LTRs showed thatregulatory
HIV-2/STreading
framessuggested
that with theexception
elements, suchasthe TATAbox, thepolyadenylation
site, of the vprgene,they
allencodedfull-length
and functional core enhancer sequences,Spl-binding
sites,
and the tat- proteins. This openreading
frame wasfound to containanresponsiveregion, areall presentinHIV-2/STand thattheir in-frame TAA stop codon that truncates the vpr protein sequences are highly conserved. The HIV-2/ST LTR is of
prematurely
after the first 32 amino acid residues. Since theon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.63.558.65.613.2]894
5500
.. . . * . .~~560 . . ..
AGGCCTTCGAGTGGCTAGA'CAGGACGGTAGAAGCCATAAACAGAGAGGCAGTGAACCACCTGCCCCGAGAGCTTATTTTCCAGGTGTGGCAAAGGTCCTGGAGATACTGGCATGATGAAC
gGlyLeuArgValAlaArgGlnAspGlyArgSerHisLysGlnArgGlySerGluProProAlaProArgAlaTyrPheProGlyValAlaLysValLeuGluIleLeuAla*** luAlaPheGluTrpLeuAspArgThrValGluAlaIleAsnArgGluAlaValAsnHisLeuProArgGluLeuIlePheGlnvalTrpGlnArgSerTrpArgTyrTrpHlisAspGluG
5600
AAGGAATGTCAATAAGTTA'CACAAAGTATAGATATTTGTGCCTAATGCAGAAAGCTATGTTCATACATTCTAAGAGAGGGTGCACTTGCCTGGGGGGAGGACATGGGCCGGGAGGATGGA
lnGlyMetSerIleSerTyrThrLysTyrArgTyrLeuCysLeuMetGlnLysAlaMetPheIleHisSerLysArgGlyCysThrCysLeuGlyGlyGlyHLsGlyProGlyGlyTrpA
5700
GATCAGGACCTCCCCCTC CTC CCCCTCCAGGTCTAGTCTAATGACTGAAGCACCAACAGAGTCTCCCCCGGAGGATAGGACCCCAC CGAG GGAG CCAGGGGATGAGTGGGTAATAGAAAC
rgSerGlyProProProProProProProGlyLeuVal***
vpr > MetThrGluAlaProThrGluSerProProGluAspArgThrProProArgGluProGlyAspGluTrpValIleGluTh
5800
CCTGAGAGAGATAAAATAAGAAGCTTTAAAGCACTTTGACCCTCGCTTGCTAATTACTCTTGGCAACTATATCTATGCTAGACATGGAGACACCCTTGAAGGCGCCAGAGGGCTCATTAG
rLeuArgGluIleLysEndGluAlaLeuLysHisPheAspProArgLeuLeulleThrLeuGlyAsnTyrIleTyrAlaArgHisGlyAspThrLeuGluGlyAlaArgGlyLeuIleAr
eDv > MetCysGlyArgAsnGlnLeuPheVallaSerLeuLeuAlaSAlaCySUItat > MetGluThrProLeuLysAlaProGluGlySerLeuG
5900 . . . 6000
GATCCTACAACGAGCCCTCCTCTTGCACTTCAGAGCAGGATGCGGCCGCTCAAGGATTGGTCAGCCCAGGGGACGAAATCCTTTATCAGCTATACCAACCCCTAGAGGCATGCGATAACA
gIleLeuGlnArgAlaLeuLeuLeuHisPheArgAlaGlyCysGlyArgSerArgIleGlyGlnProArgGlyArgAsnProLeuSerAlaIleProThrProArgGlyMetArg*** lySerTyrAsnGluProSerSerCysThrSerGluGlnAspAlaAlaAlaGlnGlyLeuValSerProGlyAspGluIleLeuTyrGlnLeuTyrGlnProLeuGluAlaCysAspAsnL
6100
AATGTTACTGTAAAAAGTGCTGCTACCATTGCCAGATGTGTTTTTTAAACAAGGGGCTCGGGATATGGTATGAACGAAAGGGCAGAAGAAGAAGAACTCCGAAGAAAACTAAGGCTCATT
ysCysTyrCysLysLysCysCysTyrHisCysGlnMetCysPheLeuAsnLysGlyLeuGlyIleTrpTyrGluArgLysGlyArgArgArgArgThrProLysLysThrLysAlaHisS
rev > MetAsnGluArgAlaGluGluGluGluLe-uArgTAriIyvsLeuArgLeuIle
SD . 70 . . 6200
CGTCTTCTGCATCAGACAAGTGAGTAAGATGTGTGGTAGGAATCAACTATTTGTTGCCAGCTTGCTAGCTAGTGCTTGCTTAATATATTGCGTCCAATATGTGACTGTTTTCTATGGCGT
erSerSerAlaSerAspLys ArgLeuLeuHisGlnThrAsn
env > MetCysGlyArgAsnGlnLeuPheValAlaSerLeuLeuAlaSerAlaCysLeuIleTyrCysValGlnTyrValThrValPheTyrGlyVa
6300
GCCCGTGTGGAGAAATGCATCCATTCCCCTCTTTTGTGCAACTAAAAATAGAGATACTTGGGGAACCATACAGTGCTTGCCAGACAATGATGACTATCAGGAAATAGCTTTAAATGTGAC
lProValTrpArgAsnAlaSerIleProLeuPheCysAlaThrLysAsnArgAspThrTrpGlyThrIleGlnCysLeuProAspAsnAspAspTyrGlnGluIleAlaLeuAsnValTh
6400
AGAGGCCTTCGACGCATGGAATAATACAGTAACAGAACAAGCAGTAGAAGATGTCTGGAGTCTATTTGAGACATCAATAAAACCATGCGTCAAACTAACACCCTTATGTGTAGCAATGCG
rGluAlaPheAspAlaTrpAsnAsnThrValThrGluGlnAlavalGluAspValTrpSerLeuPheGluThrSerIleLysProCysValLysLeuThrProLeuCysValAlaMetAr
6500 . . . .6600
TTGTAACAGCACAACTGCAAAAAACACAACCTCCACACCAACAACCACCACAACAGCAAACACAACAATAGGAGAGAATTCTTCATGCATACGCACAGACAACTGCACAGGGTTGGGAGA
gCysAsnSerThrThrAlaLysAsnThrThrSerThrProThrThrThrThrThrAlaAsnThrThrIleGlyGluAsnSerSerCysIleArgThrAspAsnCysThrGlyLeuGlyG1
6700
AGAAGAGATGG TC GACTGTCAGTTCAATATGACAGGATTAGAGAGG GA TAAGAAAAAAC TATATAATGAAACATGG TACTCAAAAGATG TAG TC TGTGAATCAAATGACAC CAAGAAAGA
uGluGluMetValAspCysGlnPheAsnMetThrGlyLeuGluArgAspLysLysLysLeuTyrAsnGluThrTrpTyrSerLysAspValValCysGluSerAsnAspThrLysLysGI
6800
GAAAACATGTTACATGAACCACTGCAACACATCAGTCATCACAGAGTCATGTGACAAGCACTATTGGGATACTATGAGGTTTAGATATTGTGCACCACCGGGTTTTGCCCTGCTAAGATG
uLysThrCysTyrMetAsnHisCysAsnThrSerValIleThrGluSerCysAspLysHisTyrTrpAspThrMetArgPheArgTyrCysAlaProProGlyPheAlaLeuLeuArgCy
6900
CAATGATACCAATTATTCAGGCTTTGAGCCCAATTGTTCTAAGGTAGTAGCTGCTACATGTACAAGGATGATGGAAACGCAAACCTCCACTTGGTTTGGCTTTAATGGCACCAGGGCAGA
sAsnAspThrAsnTyrSerGlyPheGluProAsnCysSerLysValValAlaAlaThrCysThrArgMetMetGluThrGlnThrSerThrTrpPheGlyPheAsnGlyThrArgAlaG1
7000
AAATAGAACATATATCTATTGGCATGGTAGGGATAATAGAACCATCATTAGCTTAAACAAGTTTTATAATCTCACCGTACATTGTAAGAGGCCAGGAAACAAGACAGTTGTACCAATAAC
uAsnArgThrTyrIleTyrTrpHisGlyArgAspAsnArgThrIleIleSerLeuAsnLysPheTyrAsnLeuThrValHisCysLysArgProGlyAsnLysThrValValProIleTh
710 . . . . 8 .0
ACTCATGTCAGGGTTAGTGTTTCACTCCCAGCCAATCAATAGAAGACCCAGGCAAGCATGGTGCTGGTTCAAAGGCGAGTGGAAGGAAGCCATGAAGGAGGTGAAGCTAACCCTTGCAAA
rLeuMetSerGlyLeuValPheHisSerGlnProIleAsnArgArgProArgGlnAlaTrpCysTrpPheLysGlyGluTrpLysGluAlaMetLysGluValLysLeuThrLeuAlaLy
7300
ACATCCCAGGTATAAAGGAACCAACGACACAGAAAAAATTCGTTTTATAGCGCTAGGAGAACGCTCAGACCCAGAAGTGGCATACATGTGGACTAACTGCAGAGGAGAATTTCTCTACTG
sHisProArgTyrLysGlyThrAsnAspThrGluLysIleArgPheIleAlaLeuGlyGluArgSerAspProGluVawAlaTyrMetTrpThrAsnCysArgGlyGluPheLeuTyrCy
7400
CAATATGACTTGGTTCCTCAATTGGGTAGAAAACAGAACGAATCAGACACAGCACAATTATGTGCCATGCCATATAAAGCAAATAATTAATACCTGGCACAAGGTAGGGAAAAATGTATA
sAsnMetThrTrpPheLeuAsnTrpValGluAsnArgThrAsnGlnThrGlnHisAsnTyrValProCysHisIleLysGlnIleIleAsnThrTrpHisLysValGlyLysAsnValTy
7500
TTTGCCTCCTAGGGAAGGACAGTTAACCTGCAACTCTACAGTGACCAGCATAATTGCTAACATTGACGGAGGAGAGAACCAGACAAATATTACCTTTAGTGCAGAGGTGGCAGAACTATA
rLeuProProArgGluGlyGlnLeuThrCysAsnSerThrValThrSerIleIleAlaAsnIleAspGlyGlyGluAsnGlnThrAsnIleThrPheSerAlaGluValAlaGluLeuTy
7600
CCGATTAGAATTGGGGGATTATAAATTGATAGAAGTAACACCAATTGGCTTTGCACCTACACCAGTAAAAAGATACTCCTCTGCTCCAGTGAGGAATAAAAGAGGTGTATTCGTGCTAGG
rArgLeuGluLeuGlyAspTyrLysLeuIleGluValThrProIleGlyPheAlaProThrProValLysArgTyrSerSerAlaProValArgAsnLysArgGlyValPheValLeuGe
7f7d00fe5 o n f o 7800
GTTCTTAGGTTTTCTCACGACAGCAGGAGCTGCAATGGGCGCGGCGTCCTTGACGCTGTCGGCTCAGTCTCGGACTTTATTGGCCGGGATAGTGCAGCAACAGCAACAGCTGTTGGACGT
yPheLeuGlyPheLeuThrThrAlaGlyAlaAlaMetGlyAlaAlaSerLeuThrLeuSerAlaGlnSerArgThrLeuLeuAlaGlyIleValGlnGlnGlnGlnGlnLeuLeuAspVa
7900
GGTCAAGAGACAACAAGAAATGTTGCGACTGACCGTCTGGGGAAcAAAAAATCTCCAGGCAAGAGTCACTGCTATCGAGAAATACTTAAAGGACCAGGCGCAACTAAATTCATGGGGATG
lValLysArgGlnGlnGluMetLeuArgLeuThrValTrpGlyThrLysAsnLeuGlnAlaArgValThrAlaIleGluLysTyrLeuLysAspGlnAlaGlnLeuAsnSerTrpGlyCy
8000
TGCGTCTAGACAAGTCTGCCACACTACTGTACCATGGGTAAATGACACCTTAACGCCTGATTGGAACAACATGACATGGCAGGAATGGGAGCAACGAATCCGCAACCTAGAGGCAAATAT
[image:5.612.64.556.67.616.2]sAlaSerArgGlnValCysHisThrThrValProTrpValAsnAspThrLeuThrProAspTrpAsnAsnMetThrTrpGlnGluTrpGluGlnArgIleArgAsnLeuGluAlaAsnI1
FIG. 2-Continued.
JSP4-27
provirus
isfully replication
competent, it can beunlikely
thatthe lackofafunctionalvprgeneinHIV-2/ST is concluded that the vpr geneproduct
is notrequired
for inresponsible
for its attenuatedphenotype.
vitro
replication
of HIV-2. This conclusionwasconfirmedby
Pairwisesequencealignments
of JSP4-27 with othercyto-the
biological analysis
of a secondvpr-deficient
HIV-2 pathic strains of HIV-2similarly
revealednogenetic
featuresprovirus
independently
constructed inourlaboratory
(J. C.unique
to HIV-2/ST.Comparison
of 2/ST andHIV-Kappes
and B. H.Hahn,unpublished data)
aswellasby
the 2/ROD demonstrated an overall sequencedivergence
offindings
of others (15). Moreover, sincevpr-deficient
pro- 11%,which is within theexpected
rangeofgenetic
variabil-viruses of HIV-2 are also
cytopathic
andfusogenic,
it isity
observed amonggeographicallydistantisolates of HIV-2on November 10, 2019 by guest
http://jvi.asm.org/
* . . . . 8100
CAGTGAAAGTTTAGAACAGGCACAAATCCAGCAAGAAAAGAACATGTATGAACTACAAAAATTAAATAGCTGGGATGTTTTTGGCAACTGGTTTGATTTAACCTCCTGGATCAAATATAT
eSerGluSerLeuGluGlnAlaGlnIleGlnGlnGluLysAsnMetTyrGluLeuGlnLysLeuAsnSerTrpAspValPheGlyAsnTrpPheAspLeuThrSerTrpIleLysTyrI1
eGlnTyrGlyValTyrIleValValGlyIleIleValLeuArgIleValIleTyrValValGlnMetLeuSerArgLeuArgLysGlyTyrArgProValPheSerSerProProAlaTy
SA 8300 . . . 00
CTTCCAACAGATCCATATCCACAAGGACCGGGAACAGCCAGCCAGAGAAGAAACAGAAGAAGACGTTGGAAACAGCGTTGGAGACAATTGGTGGCCCTGGCCGATAAGATATATACATTT
rPheGlnGlnIleHisIleHisLysAspArgGluGlnProAlaArgGluGluThrGluGluAspValGlyAsnSerValGlyAspAsnTrpTrpProTrpProIleArgTyrIleHisPh
tat > SerIleSerThrArgThrGlyAsnSerGlnProGluLysLysGlnLysLysThrLeuGluThrAlaLeuGluThrIleGlyGlyProGlyArg***
rev > ProTyrProGlnGlyProGlyThrAlaSerGlnArgArgAsnArgArgArgArgTrpLysGlnArgTrpArgGlnLeuValAlaLeuAlaAspLysIleTyrThrPhe
* . . . . 8700 . . . 8500
CCTGATCCGCCAGCTGATTCGCCTCTTGAACAGACTATACAACATCTGCAGGGACTTACTATCCAGGAGCTTCCAGACCCTCCAACTAATCTCCCAGAGTCTTCGGAGAGCATTGACAGC
eLeuTleArgGlnLeuIleArgLeuLeuAsnArgLeuTyrAsnIleCysArgAspLeuLeuSerArgSerPheGlnThrLeuGlnLeuIleSerGlnSerLeuArgArgAlaLeuThrA1 ProAspProProAlaAspSerProLeuGluGlnThrIleGlnHisLeuGlnGlyLeuThrIleGlnGluLeuProAspProProThrAsnLeuProGluSerSerGluSerIleAspSer
. . 8800 . . . 8600..8.
AGTCAGAGACTGGCTGAGATTTAACACAGCCTACCTGCAATATGGGGGCGAGTGGATCCAAGAAGCGTTCCGAGCCTTCGCGAGGGCTACGGGAGAGACTCTTACAAACGCCTGGAGAGG
aValArgAspTrpLeuArgPheAsnThrAlaTyrLeuGlnTyrGlyGlyGluTrpIleGlnGluAlaPheArgAlaPheAlaArgAlaThrGlyGluThrLeuThrAsnAlaTrpArgG1 SerG1nArgLeuA1aG1uIee***
nef > MetGlyAlaSerGlySerLysLysArgSerGluProSerArgGlyLeuArgGluArgLeuLeuGlnThrProGlyGluA
* . . . . 870091000. . .
CTTCTGGGGGACACTGGGA'CAAATTGGGAGGGGAATACTTGCAGTCCCAAGAAGGATCAGGCAGGGGGCAGAAATCGCCCTCCTGTGAGGGACGGCGGTATCAACAGGGAGATTTTATGA
. . .8800 .. 9300. . . .
ATACCCCATGGAGAGCCCCAGCAGAAGGGGAGAAAGGCTCGTACAAGCAACAAAATATGGATGATGTAGATTCAGATGATGATGACCTAGTAGGGGTCCCTGTCACACCAAGAGTACCAT
snThrProTrpArgAlaProAlaGluGlyysGluGlySerTyrLysGlnGlnAsnMetAspAspValAspSerAspAspAspAspLeuValGlyValProValThrProArgValProL
* U +R8900 . P PT . -->U. . . 9000
TAAGAGAAATGACATATAGGTTGGCAAGAGATATGTCACATTTGATAAAAGAAAAGGGGGGACTGGAAGGGCTGTATTACAGTGATAGGAGACGTAGAGTCCTAGACATATACTTAGAAA
euArgGluMetThrTyrArgLeuAlaArgAspMetSerHisLeuIleLysGluLysGlyGlyLeuGluGlyLeuTyrTyrSerAspArgArgArgArgValLeuAspIleTyrLeuGluL
9 100
AGGAAGAGGGAATAATTGGAGACTGGCAGAACTATACTCATGGACCAGGAGTAAGGTATCCAAAGTTCTTTGGGTGGTTATGGAAGCTAGTACCAGTAGATGTCCCACAAGAGGGAGATG
ysGluGluGlyIleIleGlyAspTrpGlnAsnTyrThrHisGlyProGlyValArgTyrProLysPhePheGlyTrpLeuTrpLysLeuValProValAspValProGlnGluGlyAspA FIG
....2-Continued.20
ACAGTGAGACTCAC TG CTTAGTG CATC CAGCACAAACAAGCAGGTTTGATGACC CGCATGGAGAAACATTAGTTTG GAGGTTTGAC CCCAC GCTAGCTTTTAGC TAC GAG GCC TTTATTC
spSerGluThrHisCysLeuValHisProAlaGlnThrSerArgPheAspAspProHisGlyGluThrLeuValTrpArgPheAspProThrLeuAlaPheSerTyrGluAlaPheIleA
.. . . . 9~~~~~~~~~~~~9300 ...
GATACCCAGAGGAGTTTGGGTACAAGTCAGGCCTGCCAGAGGATGAATGGAAGGCAAGACTGAAAGCAAGAGGGATACCGTTTAGCTAAAAACAGGAACAGCTATACTTGGTCAGGGCAG
rgTyrProGluGluPheGlyTyrLysSerGlyLeuProGluAspGluTrpLysAlaArgLeuLysAlaArgGlyIleProPheSer***
. . . ~~~~E9400 . E . Spl Spl Spl..
GAAGTAACTAACAGAAAACAGCTGAGACTGCAGGGACTTTCCAGAAGGGGCTGTTACCAGGGGAGGGACATGGGAGGAGCCGGTGGGGAACGCCCTCATACTTTCTGTATAAATGTACCC
U3<--+ ->R . 9600
GCTACTCGCATTGTATTCAGTCGCTCTGCGGAGAGGCTGGCAGATTGAGCCCTGGGAGGTTCTCTCCAGCACTAGCAGGTAGAGCCTGGGTGTTCCCTGCTAGACTCTCACCAGTGCTTG
GCCGGCACTGGGCAGACGGCTCCACGCTTGCTTGCTTAAAAGACCTCTTAATAAAGCTGCCAGTTAGAAGCA
FIG. 2Continued.
(Tables
1 and2).
Three otherrecently
reported
HIV-2closely
related to the Gambian isolate HIV-2/1SY, whichstrains, HIV-2/ISY,
derived fromaGambian individual with shared 90% of its nucleotide sequence withHIV-2/ST. AIDS (1, 17),HIV-2/NIHz,
derived from a patient with Sinceinfectivity, syncytium formation, andcellfusionareAIDS from Guinea Bissau(51),andHIV-2/GH,derivedfrom viralproperties that aremediated by theviral envgene,we a patient with AIDS from Ghana (28; A. Hasegawa, H. examined this gene in particular with respect to sequence
Tsujimoto,N. Maki, K. Ishikawa,T. Miura,M. Fukasawa, differences unique to HIV-2/ST. Alignment of the deduced K. Miki,and M. Hayami, AIDSRes. Hum. Retroviruses,in HIV-2/ST env sequence with those of six other
cytopathic
press),differfromHIV-2/ROD (Cape VerdeIslands[11,22]) and fusogenic HIV-2 and SIV strains is shown in Fig. 3. [image:6.612.67.553.70.417.2]by11, 12, and12%, respectively. Among all of thesestrains, Overall,thesizes of the variousenvsequencescomparedare the Senegalese HIV-2/ST isolate was found to be most approximately the same. In contrast to other HIV-2 and
TABLE 1. Nucleotide and amino acidsequencedivergence amongHIV-2andSIVstrainsa
Enveiop.ArrnoAcidSequenc Divergece
A HIV428T0 f4V-2IOD HIV-2IISYA HIV2NH HIV-H2/H SIVMAA SIVWSM
HIV-2/ST 19% 17% 18% 16% 28% 28%
HIV-2/ROD 11% 0% 19% 18% 28% 28%o
HIV-2/1SY 10% 11% 20% 19% 30% 29%
HIV-2/NIHZ 12% 12%o 13% 19% 28% 28%
HIV-2/GH 11% 12% 12% 15% 29% E- 29%
SI/MAC142 23% 23% 24% 26% 24% 19%
SIV/SM 23% 22% 23% 23% 23% 15%
TotW Nuclooide SequenceDivergence
aThe percentnucleotide sequence divergencebetween HIV-2/ST(JSP4-27), HIV-2/ROD (22), HIV-2/ISY (17), HIV-2/GH (Hasegawaet al., in press),
SIVMAC14 (8),andSIVsM(25)is shownalongwith thepercentaminoacidsequencedivergenceof theirenvelope glycoproteins.Sequenceswerealignedpairwise, usingthekicrogeniecomputersoftware(Beckman).
on November 10, 2019 by guest
http://jvi.asm.org/
896 NOTES
TABLE 2. Sequencehomologiesamongvirus-specificgenesof three HIV-2proviruses
%Homology
Openreadingframe ST/ROD ST/ISY ROD/ISY
Nucleotide Amino acid Nucleotide Amino acid Nucleotide Amino acid
LTR 91.5 91.4 91.8
gag 91.1 92.0 90.7 89.1 90.3 89.5
po/ 91.2 91.4 91.2 91.4 91.2 91.7
vif 91.5 88.4 92.4 91.2 92.2 85.6
vpx 89.3 85.7 92.6 90.2 88.4 87.5
vpr 89.9a 80.0a 89.8a 84.8a 93.3 89.5
tat 86.7 75.4 88.7 78.5 90.5 80.8
rev 82.0 85.0 NDb NDb NDb NDb
env 85.5 81.4 86.0 83.0 84.8 80.4
nef 86.1 78.9 85.8 77.7 87.0 78.5
Overall%homology 89.5 89.9 89.3
aAnin-framestopcodon ispresentintheHIV-2/STvpr openreadingframe.
bMeaningful comparisonof theHIV-2/ISYrevgene(17) withthecorrespondingrevgenesofHIV-2/ST(JSP4-27)and HIV-2/ROD(22)wasnotpossiblebecause ofconsiderablelengthdifferencesbetween theirsequences.ND,Notdone.
128
:ST...TAKNTTS.T HIV-2/ST
S--ESS-GN----KS HIV-2/ROD
. NAS-ESA HIV-2/ISY
.... RNM-.... HIV-2/NIHz - .N---..- HIV-2/GH -GCLG---LI-I--L-VYG---T . A---T---.T---G--S-L---S----E--- I----QS----IT----KSETDRWGL-K-S-.
-GCLG---LI-L--V-VLE-C ---K---T.---T---.S-L-I---D---I----N---. 1--I.K-ETDRWGL-GNAG
** S* * * * * * 256
PTTTTTANTT...IGENSS.CIRTDNCTGLGEEEEMVD_CQF_GLERDKKKLYETWYSKDVVCES.NDTKKEKTCYMNHC2TSVITESCDKHYWDTMRFRYCAPPGFALLRCNDTNYSGFEP
TS----TP-DQEQE. ...-S-DT.P-A-A---S---TIN---Q---TN-S-.NQTQ---AI---Y---A-VA--SPSGP...DMINDTDP--QLN--S--R--D--E
.---L----Q-S---D-S-.DR-R---A---V---TW-GR-.D-.QNIT...-INDT-.HA-A--- K----I----S---R-Q-T-A--- D.N-TS.SQSK---A---A-.-GS--GMSE...-N-. T-. PSYS---K--I-N---Y---Q---N-TKDGKNR---AIK---Y---T.I--A-P-SAPVSEKIDMVNET-S--AQN---EQ-Q-IS-K-T----K---T-E---T-L---QG-S-DN-SR ---Q---I---.Y---. M-T.---AIT--ATPSVAENVINE-NP--KNNS-A--EQ-P-IG-K .---E---.R-LI--QS.
ANES-SK---H---Q---AlI---Y---S--L--A-* S * * 384
urcsv^rvwMr.TnvAvW t:F"n AFtYR TYWH GRnDRITITISLNRFYNLTVHCKRP GKTVVPITLMSr.LV FH S -OPTINRRPROAWCWFKGEWKEAMKEVKLTLAKH PRYKCT-NDTE KTIR
SIV/MAC SIV/SM
HIV-2/ST HIV-2/ROD HIV-2/ISY
HIV-2/NIHz
HIV-2/GH SIV/MAC SIV/SM
MR-R--E----L-V-I---.---.
-E--K---E-S--K-IQ---E--V---T--.---RN-S HIV-2/ROD .---N--N HIV-2/ISY IRSR--N-K HIV-2/NIHz
.---KN-N HIV-2/GH .-N-D--N SIV/MAC
.---R--N SIV/SM
* * * * 512
FIALGERSDPEVAYMWTNCRGEFLYCIXTWFLNWVENR... ....TNQ.. TQHNYVPCHIKQIINTWHKVGKNVYLPPREGQLTCNSTVTSIIANID.GGENQTNITFSAEVAELYRLELGDYKL
-A-P-KG---.I.--. .
K-HR--A---R---E-S---WQNN---T-PEKD---RK----...-G--- E---E-S-E---VD-D-R---K-P-RG---T---S--- -G-.G...K-R--A--R-R---R----L---E---.A-D ---A---TKP-RG---A--- ---. VNS---LT-P.GGG----TF---K-N---D-D...V-T-RPKERHRR ---R
---D---L---WTDG---S--M---extracellular domain <--I--> transmembrane domain * * 640
IEVTPIGFAPTPVKRYSSAPV.RNKRGVFVLGFLGFLTTAGAAMGAASLTLSAQSRTLLAGIVQQQQQLLDVVKRQQEMLRLTVWGTKNLQARVTAIEKYLKDOAOLNSWGCASRQVCHTTVPWVNDT
-- -- __ __ _tWV___ ---A---a---u---_____ __ ---___-___ ---_---_ ---__ --_____ -_______
V---AE---0G.-V-I ---S---HQ. -
---RE---V-I
----L---D----TTGGTS-
--I----L---S-R--TTTGAS-.---S HIV-2/ROD
HIV-2/ISY HIV-2/NIHz ---S HIV-2/GH -P-AS SIV/MAC P-E- SIV/SM
768
---G-I---G---N---G-S
.-L--G-I---W---D---t:_T ---n---C
HIV-2/ST HIV-2/ROD HIV-2/ISY
HIV-2/NIHz
HIV-2/GH SIV/MAC SIV/SM
* 893
SVGDNWWPWPIRYIHFLIRQLIRLLNRLYNICRDLLSRSFQTLQLISQSLRRALTAVRDWLRFNTAYLQYGGEWIQEAFRAFAR. ATGETLTNAWRGFWGTLGQIGRGILAVPRRIRQGAEIALL HIV-2/ST
NG--RY---A- ---T---S---L---CY-N...LR--F ----C---Q-A--.--R---AG-C--L-RV-ER--- HIV-2/ROD D--SRS---E---L---T----S---LYLI--PL...LKA---C---Q-L--.V-R----S-G-SL--A--R--- HIV-2/ISY
NG--RS----A---L---TG---AISPI--P-F---Q----I---.LKA---C---Q-L--. T-R---AG-G-DL-RA-QR---L--- HIV-2/NIHZ DD-YDL----N---HL-T---TG--K---TNSP-HR----....N---I---LKA---K.T-RN---AS--G-LCAAVQRV--- HIV-2/GH
GG-NSS---Q-E---TW-FSN--T----AY-I--P-L-R-SAT-RR--EV--TELT---WSYFH--VQ-GW-S--.---AG---DL-E--RRG--W---I---L-LT-- SIV/MAC RG-SRS---Q---TW-FSS---W-L--Y-I--PVL---STT-QR--EVI-II---- WRYF---WV-WWKL-.R---AS--GDI-E---RV---I---L-LT-- SIV/SM
FIG. 3. Alignmentof the deducedamino acidsequencesof the envelopeopenreadingframes of HIV-2/ST (JSP4-27),HIV-2/ROD(22),
HIV-2/ISY(17),HIV-2/NIHz(51),HIV-2/GH(Hasegawaetal.,in press), SIVMAC (8), and SIVsM(25). The positionofthepresumedprimary
envelopeprecursor cleavage site, as identified for HIV-2/ROD(22),is shown. Symbols: *, cysteine residues conservedamongall seven
envelopesequences;*,nonconserved cysteine residues;#,in-framestopcodonspresentin thesequencesof SIVMAC andHIV-2/GH;*, gaps
introduced for optimal sequence alignment. Potential N-linked glycosylation sites (NXS/T) in the HIV-2/ST envelope sequence are
underlined.
-M.. .
-S-. K
HIV-2/ST HIV-2/ROD HIV-2/ISY HIV-2/NIHz HIV-2/GH SIV/MAC SIV/SM
HIV-2/ST HIV-21/ST
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.65.559.261.670.2]SIVMAC isolates, JSP4-27 contains no in-frame stop codon in its transmembrane envelope domain (Fig. 3). This is consis-tent with the presence of a 43-kilodalton rather than a 32-kilodalton transmembrane glycoprotein on Western blots of JSP4-27-derived virions (Fig. 1C) and is distinct from the protein profile of the HIV-2/ST parental strain, which com-prises a mixture of viruses with both full-length and trun-cated transmembrane proteins (32). Pairwise sequence align-ment shows that the HIV-2/ST env sequence differs from those of other HIV-2 and SIV envelopes to the same degree as they differ from each other, with amino acid sequences varying between 16 and 30% (Table 1). Of 32 cysteine residues, 25 are conserved among all viruses, which indi-cates a highly conserved envelope structure. In addition, HIV-2/ST contains 28 potential N-linked glycosylation sites that are arranged in a
pattern
similar to that of other viruses and that also include one highly conserved glycosylation site previously shown to be critical for HIV-1 infectivity (48). Finally, the HIV-2/ST env gene contains highly variable regions that correspond closely in distribution and size to similar hypervariable regions in the other env gene se-quences.Although a three-dimensional structure has not been de-termined for any HIV or SIV envelope glycoproteins, there are certain envelope domains whose functions have been characterized by mutagenesis analysis. These include the putative CD4-binding domain
(33,
35), the envelope precur-sor cleavage site (39), and the viral fusion sequence (5, 6, 20, 33). Since sequence changes in any one of these domains could alter the fusogenic properties of a virus (33), we analyzed the envelope sequence of HIV-2/ST for particular mutations in these areas. No changes or only conservative amino acid changes were found in an envelope domain of JSP4-27 that corresponds to the HIV-1 envelope region previously identified to be involved in CD4 binding (35). HIV-2/ST also contained an apparently functional primary envelope precursor cleavage site, with a recognition se-quence (RNKR) identical to that of three other fusogenic HIV-2 or SIV strains (Fig. 3). In contrast to these isolates, however, HIV-2/ST was found to differ in 2 of 16 highly conserved amino acid residues at the N terminus of the transmembrane envelope glycoprotein which, as shown by site-directed mutagenesis, contains the viral fusion domain (5). The mutations include an alanine-to-threonine change involving amino acid residue 517 (position 12 after the cleavage site) and a serine-to-alanine change involving amino acid residue 521 (position 16 after the cleavage site). Only one other fusogenic HIV-2 strain, HIV-2/ISY, con-tained these same changes. However, this strain exhibited three additional mutations in this same envelope area (Fig. 3). Since the fusion domain is generally highly conserved among cytopathic HIV and SIV isolates (5), we considered the possibility that the nonfusogenic properties of HIV-2/ST resulted from these mutations.To determine whether the observed amino acid substitu-tions in the HIV-2/ST envelope fusion region were likely responsible for the impaired cytopathic properties of this virus, we examined two fusogenic variants of HIV-2/ST, termed ST/24.1C and ST/24.2C (Fig. 4). Both fusogenic strains were originally derived from a biologically cloned subculture of HIV-2/ST, termed ST/24, that produced non-cytopathic and nonfusogenic virions biologically indistin-guishable from those of the parental HIV-2/ST isolate (32). After serial cell-free transmissions of ST/24 supernatant to uninfected
SupTl
cells, large and numerous syncytia were observed on two independent occasions, which indicatedtheparentalisolate
TM 42kD
IST/24.1C |ST/24.2C
[image:8.612.321.558.72.219.2]TM 3OkD TM 3OkD
FIG. 4. Diagram of the generation offusogenic and cytopathic variants of HIV-2/ST showing the origins of biologically distinct HIV-2/ST substrains. A total offive subcultures were established from the original HIV-2/ST isolate by limiting dilution cloning. All biologically cloned subcultures, the transfection-derived JSP4-27 cell line, and the parental bulk culture produce nonfusogenic and noncytopathic progeny virus ( E ). Two fusogenic and cytopathic variants ST/24.1C and ST/24.2C were generated by repeated cell-freepassageofST/24 supernatants to uninfected SupTl cells (_).
The size of the envelope transmembrane glycoprotein (TM) for virions derived from each culture is shown. kD, Kilodaltons.
emergence offusogenic progeny virus in the culture. Two cell lines were subsequently established (ST/24.1C and ST/ 24.2C) and shown to produce virions with fusogenic and
cytopathic properties similar to those of prototype HIV-1 and HIV-2 isolates (J. A. Hoxie et al., manuscript in
preparation). Moreover, these cell lines were confirmed to beinfectedwith HIV-2/ST by Southern blot analysis,which revealed no changes in their BamHI, NheI, HindlIl, and
PstI
cleavage patternscompared withST/24. Toidentifythe molecular basisforthe phenotypical change in these variants and to determine whether a direct mutation of the viralfusion sequence had occurred, we amplified the
envelope
fusion domain of these cultures by using the polymerase
chain reaction (PCR) (43).
Two oligonucleotide primers (30-mers) were designed to allow the amplification ofa 544-bp envelope fragment from virus-infected cellular DNA, which included the putative
precursorcleavagesite aswell astheenvelope fusion
region
(Fig. 5 and 6). Both primers were synthesized according to the JSP4-27 sequence; however, sequence changes were introduced toaccommodate aBamHI site in the 5' amplimer
and a
PstI
site in the 3' amplimer (primer 1, 5'-AGAATTGGGGGATCCTAAATTGATAGAAGT-3';
primer 2,5'-GCTATTTAATTTCTGCAGTTCATACATGTT-3').
Total genomic DNAsofST/24, ST/24.1C,and ST/24.2C,aswellas DNA ofST/B12 as a control, wereamplified by using these primers. A100-,u
sample of reaction mixture contained 10 mM Tris hydrochloride (pH 8.3), 50 mM KCl, 1.5 mMMgCl2,
0.01% gelatin, 200,uM
deoxynucleotidetriphos-phates, 10 pmol of each primer, 2.5 U of Taq polymerase,
and 1
,ug
of high-molecular-weight DNA. Samples were subjectedto 45 amplification cyclesconsisting
ofa denatur-ing step at94°C
for 90 s, aprimer-annealing step at50°C for 90 s,andaprimer extension stepat72°C for 135s.Amplifiedenvelopefragments were purified, cleaved withBamHI and
PstI,
andsubsequently cloned into M13. Tenindividual M13 clones per amplified DNApreparation
were then isolated,and each clone was sequenced in the region, which
corre-NOTES 897
on November 10, 2019 by guest
http://jvi.asm.org/
898 NOTES
CC
AGAATTGGGGGATTATAAATTGATAGAAGTAACACCAATTGGCTTTGCACCTACACCAGTA Primer 1 LysLeuIleGluValThrProIleGlyPheAlaProThrProVa1
---
C---T---Ser
---C---T---Ser
SerIle
---C---T---Ser
---C---T---Ser
cleavage s*it
AAAAGATACTCCTCTGCTCCAGTGAGGAATAAAAGAGGTGTATTCGTGCTAGGGTTCTTA LysArgTyrSerSerAlaProValArgAsnLysArgGlyValPheValLeuGlyPheLeu
---
G---G---G---G---__
---G---G---__
---G---G---__
---
G---G---GGTTTTCTCACGACAGCAGGACCIGCAATGGGCGCGGCGTCC TTGACGCTGTCGGCTCAG
GlyPheLeuTbrThrAlaGlyAlaAlaMetGlyAlaAlaSerLeuThrLeuSerAlaGln
I?CR 113
Sequence Clones ST/B12
ST/24
ST/24. 1C
ST/24. IC
ST/24.2C
ST/24.2C
ST/B12 ST/24 ST/24.IC ST/24.IC ST/24 .2C ST/24.2C
ST/B12 ST/24 ST/24.1C
ST/24.1C ST/24.2C ST/24.2C
10/10 10/10
1/9
8/9
5/10
5/10
10/10 10/10 1/9 8/9 5/10 5/10
10/10
10/10 1/9 8/9 5/10 5/10
JSP4-27
SerArgThrLeuLeuAlaGlyIleValGlnGlnGlnGlnGlnLeuLeuAspValValLys
Ser
PatI
C G
[image:9.612.127.498.70.427.2]JSP4-27 AGACAA...//...AACATGTATGAACTACAAAAATTAAATAGC ArgGln Primer 2
FIG. 5. Nucleotidesequence variationin thefusiondomain ofcytopathicand noncytopathicHIV-2/STstrains. A stretch of 230bpof
PCR-amplified envelopesequenceiscomparedbetweenJSP4-27and four other HIV-2/ST strains. The boundaries of theamplifiedfragments
areindicated. Thesequencesof bothprimers usedtoamplifythegenomicDNAofST/B12,ST/24, ST/24.1C,andST/24.2Careunderlined, and the basepair changes thatwereintroducedtogenerateBamHI and PstIcloningsitesareindicated.Nucleotidesubstitutionsareshown
withrespecttotheJSP4-27sequence,and amino acidsequencechangesuniquetothecytopathicandfusogenicST/24 strainsarein boldface.
Asterisksmark thethreonine and alanine substitutionspreviouslyidentifiedtodistinguishJSP4-27 frommostothercytopathicHIV-2strains.
Thenumber of M13 clonesanalyzedperHIV-2/ST strain islisted,withfrequencies referringtotheproportionof clones that haveidentical
sequences.
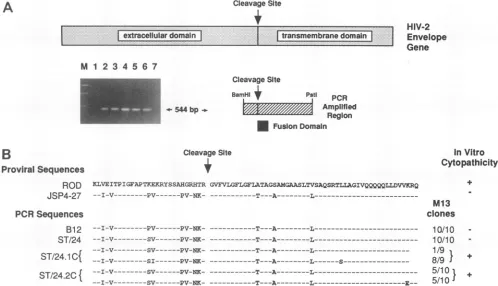
spondedtothe Nterminus of thetransmembrane envelope domain. Alignment ofthesesequencesis shown inFig. 5.
All 10 M13 clones derived from ST/B12 contained
se-quencesidenticaltothatof JSP4-27, which indicated that the PCRamplification procedurewasreliable and didnotcause
frequent misincorporations of nucleotides in this particular DNAtemplate. Sequencecomparison of amplified fragments fromcell lineST/24demonstratednodifferencesamongthe
individual M13 clones but revealed four-nucleotide point mutations between these ST/24 sequences andthe JSP4-27
reference sequence. In fact, all ST/24-derived strains,
in-cluding the cytopathic and fusogenic ones, exhibited these
same four-nucleotide sequence differences as well as the
threonine and alanine substitutions previously identified in the envelope fusion region of JSP4-27. The results thus confirmed thattheobserved fusionsequencemutationswere
representative of all HIV-2/ST strains regardless of origin andbiological phenotype. Interestingly, 8 of9 M13 clones representing ST/24.1C and 5 of10 clones representing ST/
24.2Ccontainedadditional point mutations, which predicted threeamino acidsequencechangeswithrespect totheST/24
sequence(Fig. 6). The presence of these mutations within theamplified material identifies the ST/24.1C andST/24.2C viral strains as mixtures that comprise the parental ST/24 virus aswellasadditional genotypicvariants. These newly
generated, genotypicalvariants mustberesponsible for the phenotypical changes seen in the ST/24.1C and ST/24.2C cultures. However, the biologically significant changes ap-pear notto occur in the envelope fusion domain of these variants.
While thesestudieswereinprogress, thebiological
prop-erties of the HIV-2/ST envelope gene products were also analyzedinaeucaryoticexpressionsystem(M. J. Mulligan, P. Kumar, H. Hui, R. J. Owens, G. D. Ritter, Jr., B. H. Hahn, and R. W. Compans, submitted for publication). Vaccinia virus-expressed JSP4-27 envelope glycoproteins
were compared withthose ofprototype HIV-1 and HIV-2 isolates. Whereas theprocessing, expression, andtransport tothecell surface appeared tobeunaltered, vaccinia virus-produced JSP4-27 envelope glycoproteins failed to form syncytiawithCD4-bearingHeLacells. Sincenoother HIV-2 proteinswereproducedin thissystem,theseresults strongly
JSP4 -27
JSP4-27
JSP4-27
ST/B12 ST/24 ST/24.1C
ST/24.IC
ST/24.2C
ST/24.2C Glu
10/10 10/10 1/9 8/9
5/10 5/10
10/10 10/10 5/10 5/10 ST/B12
ST/24 ST/24.2C
ST/24.2C
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 64, 1990
A
CleavageSite+
extracellular
domain.:transmemb.ane.domai
~~~~~~~.
...HIV-2
Envelope
Gene
M 1 2 3 4 5 6 7
544 bp-i
Cleavage Site
BomHll
+
Pstl PCRAmplified
Region
Fusion Domain
B
Proviral Sequences ROD
JSP4-27
PCRSequences
B12
ST/24 ST/24.1 C{
Cleavage Site
+
KLVEITPIGFAPTKF.RYSSAHGRHTR GVFVIGFLGFLATAGSAMGAASLTVSAQSRTLLAGIVQQQQQLLDVVKRQ
--I-V- PV---PV-NK --T---A-
L---I-V- PV---PV-NK LT---A-L --I-V --- PV-NK- ---T---A---- -L---I-V- SV-- PV-NK- -- -T---A---
L---I-V- SI---PV-NK-
---T---A---L---S-ST/24.2C{
--I-V
SV---PV-NK
-T---A---L---I-V---SV-- PV-NK- ---T---A----L---
E--InVitro
Cytopathicity
M13 clones
10/10 10/10 1/9 8/9 }
5/10
}
5/10I+
FIG. 6. Amino acidsequencevariation inthefusiondomain ofcytopathic and noncytopathic HIV-2/ST strains. (A) Relativelocationsof
thePCR-amplifiedenvelopefragmentsin thecontext of the entire HIV-2 envelope open reading frame. Amplification products: Lanes: 1 and 7, uninfected peripheral blood lymphocyte DNA (negative control); 2, ST/B12; 3, ST/24; 4, ST/24.1C; 5, ST/24.2C; 6, SupTl/LK001 (HIV-2/ST-infected positivecontrol cell line). (B) Alignment of the deduced amino acid sequences of the amplified fusion regions to the corresponding sequences ofHIV-2/ROD andJSP4-27. The number of M13 clones analyzed per HIV-2/ST strain is listed, with frequencies referringtothe proportion of clones that have identical sequences.
suggested that the JSP4-27 envelope glycoproteins were
primarily responsible for the nonfusogenic and
noncyto-pathic phenotype ofthis strain (Mulligan etal., submitted).
Onthebasis of theseresults as well as thesequence data, we
concludethat thenonfusogenic andattenuatedproperties of
HIV-2/ST are indeed afunction ofits envelope gene prod-ucts, although the causative genetic defect appears not to
involve mutations withintheenvelopefusion domain. Thereare several mechanisms otherthan adirect
altera-tion of the fusionsequence thatcould resultinthebiological changes observed in HIV-2/ST. These include mutations that affect envelope-CD4 interactions, mutations that cause
differencesinenvelopeglycosylation,mutations thatrequire additional cell surface molecules to facilitate virus-cell
fu-sion, and mutations that reduce the stability of envelope glycoprotein complexes on the cell surface. Infact, several
naturallyoccurring aswell asgenetically engineered
immu-nodeficiency viruses are altered in theirfusogenic or
cyto-pathic properties because ofoneofthesemechanisms. For
example, possible differences in the bindingaffinityofHIV and SIV envelope glycoproteins to the CD4 receptor have beensuggestedbythefindingthat25-foldmoresoluble CD4
is necessary to block infectivity ofprototype HIV-2
com-pared with HIV-1 isolates (10). It is
possible
that theHIV-2/ST envelope glycoprotein binds the CD4 molecule with an even lower affinity, which would be expected to influence subsequent steps of viral entry, including mem-brane fusion andpenetration. Anothermechanismknownto causeattenuation of virulence in
naturally
occurring
retro-viruses involves differences in posttranslational modifica-tions of envelope glycoproteins. Poss and co-workers showed that the pathogenic determinants of an immunode-ficiency-causing feline leukemia virus were dependent on the
processing of particular envelope oligosaccharides (41). Since HIV-2/ST differs in number and distribution of its potential N-linked envelope glycosylation sites from other cytopathic HIV-2 strains andsince sizedifferences between the exterior envelopeglycoproteins offusogenic and nonfu-sogenic HIV-2/ST strains have been observed (J. A. Hoxie,
personalcommunication),abiologically significant changein the sugarcomposition of the HIV-2/STenvelope cannotbe excluded.Finally,arequirement of accessory molecules for virus-cellfusion represents still anotherpotentialmechanism to influence retroviral cytopathicity. Studies involving SIVMAC recently revealed that this virus has a restricted host cellrange thatcomprises onlyasubset ofCD4+ T-cell lines(27,31). Although highlyinfectious andcytopathicfor HUT78 and H9 cells, SIVMAC does not fuse with
CD4-bearingSupTlcells.Moreover, SIVMACinfectsSupTlcells
onlywithconsiderabledelay. It istherefore conceivable that
SIVMAC requiresasurfacemolecule(s)inadditiontoCD4 to establishaproductiveinfection in certain human T-cell lines. Since its infection kinetics and lack ofcytopathic effect in
SupTlcells very much resemble those ofHIV-2/ST,it is not unreasonable to speculate that HIV-2/ST similarly requires
anadditional cell surfacemolecule(s)forefficient cell fusion orpenetration.
Theavailabilityofcytopathicvariants ofHIV-2/ST will be
NOTES 899
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.612.63.561.76.362.2]900 NOTES
instrumental for future experiments designed to define the exactmolecular determinants involved inHIV-2/ST attenu-ation. Molecular clones representing the fusogenic and
cy-topathic HIV-2/STstrainsareexpected toexhibit muchless genetic divergencewithrespecttoJSP4-27 than dounrelated HIV-2 proviruses such as HIV-2/ROD and HIV-2/ISY.
Therefore, acomparativesequenceanalysisismorelikelyto identify biologicallyimportantdifferences, and the
construc-tion of chimeras between attenuated and cytopathic clones will be greatly facilitated. The fact that cytopathic and fusogenicST/24mutantsevolved bycell-free passage ontwo independent occasions indicates the presence of strong
selective pressures for cytopathic and fusogenic viruses in
vitro. It is possiblethat similarpressuresare alsopresent in vivowhichmayfavor theemergenceofmorevirulentstrains
in certain HIV-infected individuals overtime (2, 9, 47).
Wethank Gerald Myers and his co-workers for assistance with the envelope amino acid sequence alignments, Masanori Hayami
and Genoveffa Franchini for supplying HIV-2 proviral sequences
priortopublication,JereSegrest for helpfuldiscussions, and A. Jan
Nicholson for carefulpreparation of the manuscript.
Thisworkwassupported bygrantsfrom theNationalInstitutesof Health, the U.S. Army Medical ResearchAcquisition Activity, the
Lifeand HealthInsurance Medical ResearchFund, and theCarter/ Wallace Corp.B.H.H. isaSpecial Fellow andJ.A.H.isaScholarof
theLeukemia Society of America. G.M.S. is PEW Scholar of the
Biomedical Sciences.
LITERATURECITED
1. Albert,J.,U. Bredberg, F.Chiodi,B. Bottiger,E.M.Fenyo,E.
Norrby,and G. Biberfeld. 1987. Anewhuman retrovirusisolate
of West African origin (SBL-6669) and its relationship to
HTLV-IV, LAV-II, and HTLV-IIIB. AIDS Res. Hum.
Retro-viruses3:3-10.
2. Asjo, B., J. Albert, A. Karlsson, L. Morfeldt-Mamson, G.
Biberfeld, K. Lidman, and E. M. Fenyo. 1986. Replicative
propertiesofhuman immunodeficiency virus from patients with
varying severityofHIV infection. Lancetii:660-662.
3. Barre-Sinoussi, F., J. C.Chermann, F. Rey, M. T. Nugeyre, S.
Chamaret, J. Gruest, C. Dauguet, C. Axler-Blin, F. Vezinet.
Brun, C. Rouzioux, W. Rozenbaum, and L. Montagnier. 1983.
Isolation ofaT-lymphotropic retrovirus froma patientat risk
for acquired immune deficiency syndrome (AIDS). Science
220:868-870.
4. Berkelman,R.L., W. L.Heyward, J. K.Stehr-Green,and J.W.
Curran. 1989. Epidemiology ofhumanimmunodeficiency virus
infection and acquired immunodeficiency syndrome. Am. J.
Med. 86:761-770.
5. Bosch, M. L., P. L. Earl, K. Fargnoli, S. Picciafuoco, F.
Giombini,F. Wong-Staal, andG. Franchini. 1989. Identification
of the fusion peptide of primate immunodeficiency viruses.
Science 244:694-697.
6. Brasseur, R., B. Cornet, A. Burny, M. Vandenbranden, and
J. M. Ruysschaert. 1988. Mode of insertion into a lipid mem-braneofthe N-terminal HIVgp4lpeptide segment. AIDSRes.
Hum. Retroviruses4:83-90.
7. Centers for Disease Control. 1988. AIDS due to HIV-2
infec-tion-NewJersey. Morbid. Mortal. Weekly Rep. 37:33-35.
8. Chakrabarti, L., M. Guyader, M. Alizon, M. D. Daniel, R. C.
Desrosiers,P. Tiollais, and P. Sonigo. 1987. Sequenceofsimian
immunodeficiency virus from macaqueand its relationship to
other human and simian retroviruses. Nature (London) 328:
543-547.
9. Cheng-Mayer, C., D. Seto, M. Tateno, andJ. A. Levy. 1988.
Biologic featuresofHIV-1 that correlate with virulencein the
host.Science240:80-82.
10. Clapham,P. R., J.N. Weber, D. Whitby, K. McIntosh, A. G.
Dalgleish,P. J. Maddon, K. C.Deen, R. W. Sweet, and R. A.
Weiss. 1989. Soluble CD4 blocks the infectivity of diverse
strains of HIV and SIVfor
T-cells
and monocytes but notfor brain and musclecells. Nature (London) 337:368-370. 11. Clavel, F., D. Guetard, F. Brun-Vezinet, S. Chamaret, M. A.Rey, M. 0. Santos-Ferreira, A. G. Laurent, C. Dauguet, C.
Katlama,
C.Rouzioux,D.Klatzmann,J.Champalimaud, and L. Montagnier. 1986. Isolation of a new human retrovirus from West African patients with AIDS. Science 233:343-346. 12. Clavel, F.,K.Mansinho, S.Chamaret,D.Guetard,V.Favier,J.Nina, M. 0. Santos-Ferreira, J. L. Champalimaur, and L. Montagnier. 1987. Humanimmunodeficiency virustype2 infec-tion associated with AIDS in West Africa. N. Engl. J. Med. 316:1180-1185.
13. Cortes, E., R. Detels, D. Aboulafia, X. L. Li, T. Moudgil, M. Alam, C. Bonecker, A. Gonzaga, L. Oyafuso, M. Tondo, C. Boite, N.
Hammershlak,
C. Capitani, D. J. Slamon, and D. D. Ho. 1989. HIV-1, HIV-2, and HTLV-1 infection in high-risk groups in Brazil. N. Engl. J. Med. 320:953-958.14. Daniel, M. D., N. L. Letvin, N. W. King, M. Kannagi, P. K. Sehgal, R. D. Hunt,P.J. Kanki,M.Essex,and R. C. Desrosiers. 1985. Isolation of T-cell tropic
HTLV-III-like
retrovirus from macaques. Science228:1201-1204.15. Dedera, D., W. Hu, N. V. Heyden, and L. Ratner. 1989. Viral protein R of human immunodeficiency virus types 1 and 2 is dispensable for replication and cytopathogenicity in lymphoid cells. J. Virol. 63:3205-3208.
16. Evans, L. A., J.Moreau, K. Odehouri, H. Legg,A.Barboza, C. Cheng-Mayer, and J. A. Levy. 1988. Characterization of a
noncytopathic HIV-2 strain with unusual effects on CD4 expres-sion. Science 240:1522-1525.
17. Franchini, G., K. A. Fargnoli, F. Giombini, L. Jagodzinski, A. De Rossi, M. Bosch, G. Biberfeld, E. M. Fenyo, J. Albert, R. C. Gallo, and F. Wong-Staal. 1989. Molecular andbiological char-acterization of a replication competent human immunodefi-ciency type 2 (HIV-2) proviral clone. Proc.
Natl.
Acad. Sci. USA 86:2433-2437.18. Franchini, G., J. R. Rusche, T. J.
O'Keeffe,
and F.Wong-Staal. 1988. The human immunodeficiency virus type 2 (HIV-2) con-tains a novel gene encoding a 16 kD protein associated with mature virions. AIDS Res. Hum. Retroviruses 4:243-250. 19. Fukasawa, M., T. Miura, A. Hasegawa, S. Morikawa, H.Tsuji-moto, K. Miki, T. Kitamura, and M. Hayami. 1988. Sequence of simian immunodeficiency virus from African green monkey, a new member of the HIV/SIV group. Nature (London) 333: 457-461.
20. Gallaher, W. R. 1987. Detection of a fusion peptide sequence in the transmembrane protein of human immunodeficiency virus. Cell 50:327-328.
21. Gallo, R. C., S. Z. Salahuddin, M. Popovic, G. M. Shearer, M. Kaplan, B. F. Haynes, T. J. Palker, R.
Redfield,
J. Oleske, B. Safai, G.White, P. Foster, and P. D. Markham. 1984. Frequent detection and isolation of cytopathic retroviruses(HTLV-III)
from patients with AIDS and at risk for AIDS. Science 224: 500-503.21a.Goodenow, M., T. Huet, W. Saurin, S. Kwok, J. Sninsky, and S. Wain-Hobson. 1989. HIV-1 isolates are rapidly evolving quasi-species: evidence for viral mixtures and preferred nucleotide substitutions. J. AIDS 2:344-352.
22. Guyader, M., M. Emerman, P. Sonigo, F. Clavel, L. Montagnier, and M. Alizon. 1987. Genome organization and transactivation of the human immunodeficiency virus type 2. Nature (London) 326:662-669.
23. Haseltine, J. G., and J. Sodroski. 1987. Cell membrane fusion mediated by the envelope glycoprotein as the primary effector of AIDS virus cytopathicity, p. 47-56. In J. C. Gluckman and E. Vilmer (ed.), Acquired immunodeficiency syndrome. Elsevier Science Publishing, Inc., New York.
24. Henderson, L. E., R. C. Sowder, T. D. Copeland, R. E. Ben-veniste, and S. Oroszlan. 1988. Isolation and characterization of a novel protein (X-ORF product) from SIV and HIV-2. Science 241:199-201.
25. Hirsch, V. M., R. A. Olmsted, M. Murphey-Corb, R. H. Purcell, and P. R. Johnson. 1989. AnAfricanprimate lentivirus
(SIVsm)
closely related to HIV-2. Nature (London) 339:389-392.