Copyright © 1998, American Society for Microbiology
Rescue and Autonomous Replication of Adeno-Associated
Virus Type 2 Genomes Containing Rep-Binding Site
Mutations in the Viral p5 Promoter
XU-SHAN WANG1,2,3ANDARUN SRIVASTAVA1,2,3,4*Department of Microbiology and Immunology,1Walther Oncology Center,2and Division of Hematology/Oncology,
Department of Medicine,4Indiana University School of Medicine, and Walther Cancer Institute,3
Indianapolis, Indiana 46202
Received 3 December 1997/Accepted 11 February 1998
The Rep proteins encoded by the adeno-associated virus type 2 (AAV) play a crucial role in the rescue, replication, and integration of the viral genome. In the absence of a helper virus, little expression of the AAV Rep proteins occurs, and the AAV genome fails to undergo DNA replication. Since previous studies have established that expression of the Rep78 and Rep68 proteins from the viral p5 promoter is controlled by the Rep-binding site (RBS) and the YY1 factor-binding site (YBS), we constructed a number of recombinant AAV plasmids containing mutations and/or deletions of the RBS and the YBS in the p5 promoter. These plasmids were transfected in HeLa or 293 cells and analyzed for the potential to undergo AAV DNA rescue and replication. Our studies revealed that (i) a low-level rescue and autonomous replication of the wild-type AAV genome occurred in 293 but not in HeLa cells; (ii) mutations in the RBS resulted in augmented expression from the p5 promoter, leading to more efficient rescue and/or replication of the AAV genome in 293 but not in HeLa cells; (iii) little rescue and/or replication occurred from plasmids containing mutations in the YBS alone in the absence of coinfection with adenovirus; (iv) expression of the adenovirus E1A gene products was insufficient to mediate rescue and/or replication of the AAV genome in HeLa cells; (v) autonomously replicated AAV genomes in 293 cells were successfully encapsidated in mature progeny virions that were biologically active in secondary infection of HeLa cells in the presence of adenovirus; and (vi) stable transfection of recombinant AAV plasmids containing a gene for resistance to neomycin significantly affected stable integration only in 293 cells, pre-sumably because rescue and autonomous replication of the AAV genome from these plasmids occurred in 293 cells but not in HeLa or KB cells. These data suggest that in the absence of adenovirus, the AAV Rep protein-RBS interaction plays a dominant role in down-regulating viral gene expression from the p5 promoter and that perturbation in this interaction is sufficient to confer autonomous replication competence to AAV in 293 cells.
The adeno-associated virus type 2 (AAV), a nonpathogenic human parvovirus, contains a single-stranded DNA genome of 4,680 nucleotides (55). Optimal replication of the wild-type (wt) AAV genome requires coinfection with a helper virus, such as adenovirus or herpesvirus (2–5). In the absence of a helper virus, the wt AAV genome integrates into the host chromosomal DNA in a site-specific manner to establish a latent infection (7, 17–20, 48). When a latently infected cell is subsequently infected with a helper virus, the integrated wt AAV genome undergoes rescue and proceeds through a nor-mal productive infection (31, 32). The AAV genome can also be rescued from recombinant plasmids containing the wt viral genome by transfecting the plasmid DNA into adenovirus-infected human cells (44, 47). Thus, recombinant plasmids have proven to be a useful model system with which to study the molecular events involved in rescue and replication of the latent proviral AAV genome (10, 44–46, 56, 59–61). Two se-quences in the wt AAV genome are essential for viral DNA replication. The first is the viral origin of DNA replication, which consists of a 145-nucleotide inverted terminal repeat
(ITR) sequence, the terminal 125 nucleotides of which form a hairpin palindrome that is used as a primer for initiation of viral DNA replication (9, 26, 54). The second is the viral rep gene, which codes for four viral nonstructural proteins (Rep) that are synthesized from a single open reading frame by the use of alternate promoters and splicing (54). Rep78 and Rep68 are expressed from a promoter at map unit 5 (p5), and Rep52 and Rep40 are derived from expression from a promoter at map unit 19 (p19) in the viral genome (3, 4, 30, 54). The Rep proteins have multiple functions and are involved in rescue, replication, encapsidation, and integration of the AAV ge-nome as well as in regulation of the viral gene expression (12, 13, 24–30, 35–38, 49, 52, 53, 56, 63, 64). In the absence of adenovirus, the Rep proteins repress the production of the p5 and p19 transcripts, but in the presence of adenovirus, the Rep proteins simultaneously activate and repress the AAV p5 pro-moter and activate expression from the p19 propro-moter in the AAV genome (37, 38). Previous studies have shown that Rep78 and Rep68 specifically bind to the cruciform structures of the AAV ITRs (1, 14). The Rep proteins also possess a site-specific and strand-specific endonuclease activity which specifically cleaves at the terminal resolution site (trs) within the ITR sequences (15, 16). Both the secondary structure el-ement of the ITR and a specific sequence at trs are required for the recognition and efficient cleavage by the Rep proteins (52, 53), although a low-level cleavage can occur in the absence of the secondary structure (27). However, recent in vitro studies * Corresponding author. Mailing address: Department of
Microbi-ology and ImmunMicrobi-ology, 635 Barnhill Dr., Medical Science Building, Room 231-B, Indiana University School of Medicine, Indianapolis, IN 46202-5120. Phone: (317) 274-2194. Fax: (317) 274-4090. E-mail: [email protected].
4811
on November 9, 2019 by guest
http://jvi.asm.org/
have documented that the purified Rep68 protein binds not only to the ITR but also to linear DNA sequences, such as the A sequence, and p5 and p19 promoter sequences (24, 27, 37, 62). The Rep-binding site (RBS) in the p5 promoter between the TATA box and the transcription start site is responsible for the repression of expression of Rep68 (24, 37). In addition to this RBS, the binding site for yet another transcription factor, YY1 (58) (the YY1 factor-binding site [YBS]), in the p5 pro-moter is also responsible for regulation of expression of Rep proteins (6, 50, 51). Although the RBS in the p5 promoter is believed to be involved in rep gene expression (24, 35) and AAV DNA integration (8), it is unclear whether Rep-mediated binding to these sequences is required for viral DNA replica-tion. Similarly, the binding of YY1 to the p5 promoter and its consequences for AAV DNA replication in the absence of a helper virus have not been rigorously examined.
In this report, we document that the wt AAV genome un-dergoes rescue and autonomous replication in 293 but not in HeLa cells and that autonomous replication of the AAV ge-nome correlates well with expression from the viral promoters in 293 cells. Interestingly, however, expression of the adenovi-rus E1A proteins is insufficient to mediate rescue and replica-tion of the AAV genome in HeLa cells. The autonomously replicated AAV genomes in 293 cells are successfully encap-sidated in mature progeny virions that are biologically active. These studies yield insights into the complex interaction of Rep proteins with the viral genome leading to down-regulation of gene expression from the p5 promoter and indicate that perturbation in this interaction is sufficient to confer autono-mous replication competence to AAV in 293 cells.
MATERIALS AND METHODS
Cells, viruses, and plasmids.Human cervical carcinoma cell line HeLa and
human nasopharyngeal carcinoma cell line KB were provided by Asok C. Antony (Indiana University School of Medicine, Indianapolis). Human embryonic kid-ney cell line 293 was obtained from the American Type Culture Collection (Rockville, Md.). Cells were maintained as monolayer cultures in Iscove’s mod-ified Dulbecco’s medium supplemented with 10% fetal bovine serum, penicillin, and streptomycin as previously described (33, 34). AAV and the human adeno-virus type 2 (Ad2) adeno-virus stocks were obtained from Kenneth I. Berns, Cornell University Medical College, New York, N.Y., and Kenneth H. Fife, Indiana University School of Medicine, respectively, and propagated as previously de-scribed (21). The recombinant AAV plasmid pSub201 (45) was supplied by Richard J. Samulski, University of North Carolina, Chapel Hill.
Construction of recombinant AAV plasmids. Standard cloning techniques
were used for constructing all recombinant plasmids (43). First, a plasmid des-ignated pXS-26B was constructed by replacing the right XbaI site in plasmid pSub201 with a synthetic oligonucleotide linker sequence (59-CTAGGGAATT CC-39) containing an EcoRI site. To construct plasmids pXS-70A, pXS-70B, and pXS-70C, plasmid pSub201 was digested with Eco31I and KpnI. The left half of the AAV genome fragment was isolated and ligated with synthetic oligonucleo-tide linker sequences (59-CCGAGATCTCGATCAGGGTCTCC-39, 59-CCGAG TGAGCACGCAGGGTTTAA-39, and 59-CCGAGATCTCGTCAGGGTTTA A-39, respectively) and then inserted into plasmid pXS-24, described previously (60), which was digested with AvaI and KpnI. The resulting plasmids, designated pXS-69A, pXS-69B, and pXS-69C, respectively, were digested with XbaI and
HindIII, and these fragments were used to replace the XbaI-HindIII fragment in
plasmid pXS-26B. Plasmid pXS-48B was constructed by replacing the
XbaI-HindIII fragment of 26B, using the XbaI-HindIII fragment from
pXS-47, also described previously (62). The wt AAV coding sequences flanked by the Ad5 ITRs were cloned between the BalI sites in plasmid pSub201 to generate plasmid pXS-37. A DNA fragment containing the herpesvirus thymidine kinase (TK) promoter-driven gene for resistance to neomycin (TK-neor) isolated from
plasmid pTwu.G1 (33) was inserted at the ClaI site in plasmid pXS-37 to gen-erate the plasmid pXS-38. Similarly, the TK-neorfragment was inserted at the ClaI site in plasmid pXS-18, lacking the AAV D sequences described previously
(59), to generate the plasmid pXS-39. Plasmid pXS-40 was generated by inserting the TK-neorfragment at the ClaI site in plasmid pSub201.
Southern blot analysis for AAV DNA rescue and replication.DNA-mediated
transfections were carried out in triplicate by the calcium phosphate coprecipi-tation method (43) with 10mg of each plasmid per 100-mm-diameter dish of 70% confluent 293 cells. At various times posttransfection, low-MrDNA samples were
isolated by the procedure described by Hirt (11), digested extensively with DpnI, and analyzed on Southern blots by using the32P-labeled AAV DNA probe as
previously described (33, 34). HeLa cells were transfected by using Lipofectin as recommended by the vendor (Boehringer Mannheim Biochemicals, Indianapo-lis, Ind.).
Northern blot analysis for AAV gene expression.Cells were either mock
transfected or transfected in triplicate with equivalent amounts of various re-combinant AAV plasmids by using Lipofectin as described above. Forty-eight hours posttransfection, total cellular RNA was isolated from two-thirds of the cells and analyzed on Northern blots, using two32P-labeled DNA probes, one
specific for the AAV and one specific for the glyceraldehyde phosphate dehy-drogenase (GAPDH) cDNA sequence. The remainder one-third of the cells were used to isolate low-MrDNA for analyses of plasmid uptake on quantitative
DNA slot blots by using the32P-labeled AAV DNA probe as previously
de-scribed (22).
Encapsidation and biological activity of AAV progeny virions. Equivalent
amounts of plasmids pXS-70A and pSub201 were transfected in 293 cells as previously described (59–62). Approximately 72 h posttransfection, cells were harvested and progeny virions were purified by one round of CsCl equilibrium density gradient followed by exhaustive digestion with DNase I as previously described (21). These viral stocks were used in secondary infections of Ad2-infected HeLa cells, and low-MrDNA samples isolated at various times
postin-fection were analyzed on Southern blots using32P-labeled AAV probe as
pre-viously described (33, 34).
Stable transduction assays.Equivalent numbers of HeLa, KB, and 293 cells
were transfected in triplicate with 10 mg each of the recombinant plasmids pXS-38, pXS-39, and pXS-40, separately. Forty-eight hours posttransfection, G418 was added at a final active concentration of 400mg/ml, and G418-resistant colonies were enumerated 14 days later as previously described (33, 41).
RESULTS
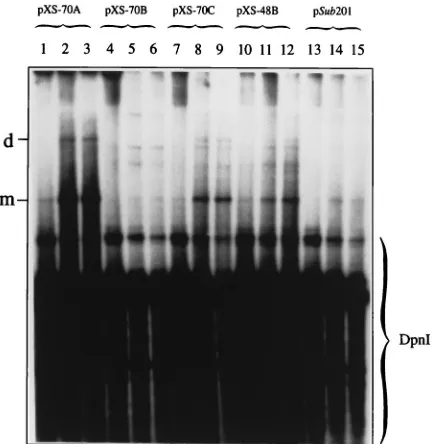
AAV DNA sequences undergo rescue from recombinant plasmids followed by autonomous replication in human 293 but not in HeLa cells. Previous studies have shown that the RBS in the p5 promoter is involved in the repression of ex-pression of Rep78 and Rep68 proteins in the absence of the helper virus and that the YBS is also involved in the regulation of Rep protein expression (6, 24, 27). Further analyses re-vealed that mutations in the RBS provide partial relief from down-regulation by Rep68, and mutations in the YBS reduced the p5 mRNA levels in 293 cells (24, 37). Since these studies were confined to analyses of gene expression in the absence of AAV DNA replication in experiments involving plasmid trans-fections, we examined the effects of these mutations on viral DNA replication in the context of the AAV genome. We created a number of mutations in the RBS and the YBS within the p5 promoter of the wt AAV genome, which are schemat-ically depicted in Fig. 1. Briefly, plasmid pSub201 contains the wt AAV genome (45). Plasmids pXS-70A and pXS-70B con-tain mutations in the RBS and the YBS, respectively, in the viral p5 promoter. In plasmid pXS-70C, both RBS and YBS have been mutated; in plasmid pXS-48B, both sites have been deleted. Equivalent amounts of these plasmids were trans-fected separately into 293 cells in the absence of adenovirus, and low-MrDNA samples were isolated at various times post-transfection, digested extensively with DpnI, and analyzed on Southern blots by using the 32P-labeled AAV DNA probe. These results are shown in Fig. 2. The roughly equivalent hybridization intensities of DpnI fragments generated from each plasmid, observed even at much shorter autoradiographic exposures, indicate that the amounts of the input plasmids were approximately the same. It is interesting that a low-level rescue and replication of the AAV genome from plasmid pSub201 occurred in 293 cells, as evidenced by time-dependent accumulation of DpnI-resistant AAV monomeric and dimeric replicative DNA intermediates. The efficiency of rescue and/or replication of the AAV genome in 293 cells transfected with plasmid pXS-70A containing a mutation in the RBS was sig-nificantly increased. These replicative DNA intermediates dis-appeared when the low-MrDNA samples were digested exten-sively with DpnI, which digests input plasmid DNA but not DNA replicated in human cells, and DpnII, which digests DNA
on November 9, 2019 by guest
http://jvi.asm.org/
replicated in human cells (data not shown). These results in-dicate that AAV DNA containing a mutation in the RBS can undergo rescue from the recombinant plasmid pXS-70A fol-lowed by a full round of autonomous replication in 293 cells in the absence of adenovirus, albeit at several orders of magni-tude less than that observed from the plasmid pSub201 in the presence of adenovirus. The efficiency of rescue and replica-tion of the AAV genome from plasmids 70C and
pXS-48B, containing a mutation and a deletion in the RBS and the YBS, respectively, was higher than that from plasmid pSub201 but less than that from plasmid pXS-70A. Little rescue and/or replication of the AAV genome occurred from the plasmid pXS-70B, which contained a mutation in the YBS. Interest-ingly, however, when these studies were carried out with HeLa cells under identical conditions, the AAV genome failed to undergo rescue and replication, even from the plasmid pXS-70A (data not shown, but see later). Abundant rescue and/or replication of the AAV genome from each of the plasmids occurred in the presence of adenovirus in 293 (Fig. 3) as well as HeLa (data not shown) cells. These results demonstrate that the RBS mediates repression of replication and that the YBS promotes AAV DNA replication in the absence of adenovirus in 293 cells.
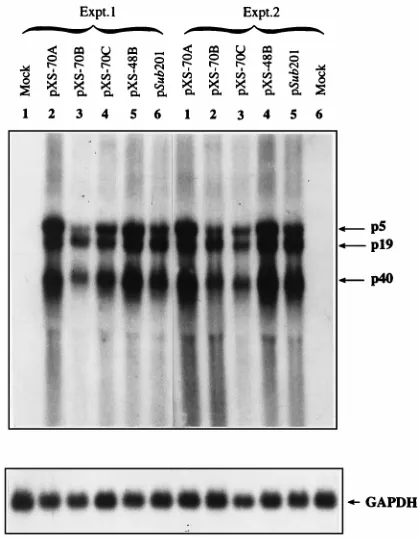
[image:3.612.55.288.63.224.2]Autonomous replication of the AAV genomes correlates with expression of the rep genes.It was next of interest to investigate whether mutations in the RBS and the YBS also had an effect on rep gene expression. Each of the recombinant AAV plas-mids was transfected in triplicate into 293 cells in the absence of adenovirus. Forty-eight hours posttransfection, total RNA and low-MrDNA were isolated and analyzed on Northern and quantitative DNA slot blots, respectively, using the32P-labeled AAV DNA as a probe. Such blots from two separate experi-ments are shown in Fig. 4 and 5. Approximately equivalent hybridization intensities of RNA samples with the GAPDH probe (Fig. 4) and that of low-MrDNA with the AAV probe (Fig. 5) indicate that RNA loads and transfection efficiency of each of the recombinant plasmids were roughly the same. A number of conclusions can be drawn from these data. First, the presence of the RBS represses the expression of rep genes, especially the p5 transcripts. For example, in cells transfected with plasmids pXS-70A, pXS-70C, or pXS-48B, in each of FIG. 1. Schematic structure of the AAV genome containing various
muta-tions and/or delemuta-tions in the RBS and the YBS in the viral p5 promoter. Muta-tions and deleMuta-tions are denoted by the prefixes “m” and “d.” Asterisks indicate the deleted nucleotides. The AAV p5 promoter is represented by the closed circle, and the RNA start site is indicated by the arrow. The hairpin (HP) and the D sequences that constitute the viral ITRs are represented by open and closed rectangles, respectively. Each of the recombinant plasmids was constructed as described in Materials and Methods.
FIG. 2. Southern blot analyses of rescue and replication of AAV genomes containing various mutations and/or deletions in the RBS and YBS in 293 cells in the absence of adenovirus. Ten micrograms of each indicated plasmid DNA was transfected separately in approximately 70% confluent 293 cells in a 10-cm-diameter dish, and low-MrDNA isolated at 24 h (lanes 1, 4, 7, 10, and 13), 48 h
[image:3.612.319.538.67.293.2](lanes 2, 5, 8, 11, and 14), and 72 h (lanes 3, 6, 9, 12, and 15) posttransfection was digested with DpnI and analyzed on Southern blots, using an AAV-specific DNA probe. Autoradiography was performed at270°C for 16 h. m and d denote the monomeric and dimeric replicative forms of the AAV genome.
FIG. 3. Southern blot analyses of rescue and replication of AAV genomes containing various mutations and/or deletions in the RBS and YBS in adenovi-rus-infected 293 cells. Ten micrograms of each indicated plasmid DNA was transfected separately in approximately 70% confluent 293 cells, and low-Mr
DNA isolated at 24 h (lanes 1, 4, 7, 10, and 13), 48 h (lanes 2, 5, 8, 11, and 14), and 72 h (lanes 3, 6, 9, 12, and 15) posttransfection was digested with DpnI and analyzed on Southern blots, using an AAV-specific DNA probe as described in the legend to Fig. 2. Autoradiography was performed at room temperature for 15 min. m and d denote the monomeric and replicative forms of the AAV genome.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.63.280.426.648.2]which there are mutations or deletions in the RBS, the level of p5 transcripts, and specifically the ratio of p5 transcripts to p19 or p40 transcripts, is significantly increased. Interestingly, mu-tations in the YBS repress the levels of p5 transcripts. Second, the effects of the RBS and the YBS on p5 transcripts appear to be independent since the level of p5 transcript is highest in cells transfected with plasmid pXS-70A, the lowest with plas-mid pXS-70B, and intermediate with plasplas-mid pXS-70C. Thus, in the absence of adenovirus, the RBS-mediated repression of the p5 promoter and the YBS-mediated stimulation of the p5 promoter appear to be independent. Third, the extent of AAV DNA replication correlates with the levels of p5 transcripts. For example, when the p5 transcripts are high, the level of AAV DNA replication is also high (pXS-70A, pXS-70C, and pXS-48B; compare Fig. 2 and 4). Fourth, the RBS and the YBS appear to selectively affect the levels of p5 transcripts. For example, mutations in the RBS and the YBS result in a signif-icant increase in the ratio of p5 transcripts to p19 and p40 transcripts. Expression of the p5 transcripts also affects the levels of p19 and p40 transcripts because when the level of p5 transcripts is high, the levels of p19 and p40 transcripts are also high. When these experiments were carried out in the presence of adenovirus, the efficiency of transcription from each of the three AAV promoters was increased significantly, requiring autoradiographic exposure of Northern blots only for approx-imately 1 h, but there was an abundant increase in the ratio of p40 transcripts to p19 or p5 transcripts (data not shown), an observation consistent with a recently published report (25). However, the levels of p5 transcripts in adenovirus-infected
293 cells were not significantly different following transfection with plasmid pXS-70A or pXS-70C but less than that with plasmid pXS-70B or pSub201 (data not shown). Thus, the RBS is required for activation of AAV transcription by adenovirus but does not have an effect on viral DNA replication in the presence of adenovirus.
Adenovirus E1A gene products are insufficient to mediate rescue and replication of the AAV genome in HeLa cells.As indicated above, rescue and autonomous replication of the AAV genome from none of the transfected recombinant plasmids could not be detected in HeLa cells even when high-efficiency transfection protocols were used. Since 293 cells constitutively express the adenovirus early-region E1 gene products that are known to transactivate the YBS (6, 51), it was reasonable to assume that adenovirus E1A and/or E1B gene products were responsible for the rescue and autonomous rep-lication of the AAV genome in 293 cells in the absence of adenovirus. This possibility was experimentally tested when each of the recombinant AAV plasmids was transfected into HeLa cells either in the absence or in the presence of an expression plasmid containing the adenovirus E1A gene. The results of these experiments are depicted in Fig. 6. It is evident that the adenovirus E1A gene products alone could not sup-port rescue and replication of the AAV genome in HeLa cells in the absence of adenovirus. Although it remains possible that the adenovirus E1B gene products are required for AAV DNA replication, this was not pursued further in view of the fact that even in 293 cells, an additional factor(s), other than the ade-novirus E1A and E1B gene products, is required for autono-mous replication of AAV DNA since rescue and/or replication also occurred from plasmids pXS-70C and pXS-48B (Fig. 2), which contain a mutation or a deletion in the YBS.
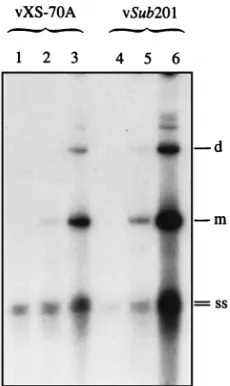
Rescue and autonomous replication of the AAV genome lead to generation of biologically active progeny virions.Since com-plete AAV DNA replication and gene expression occurred in 293 cells, it was next of interest to examine whether progeny AAV particles could also be assembled in the absence of ad-enovirus. Plasmids pXS-70A and pSub201 were transfected separately into 293 cells in the absence of adenovirus as
de-FIG. 4. Northern blot analyses of expression of AAV genes containing var-ious mutations and/or deletions in the RBS and YBS in 293 cells. Total cellular RNA was isolated from mock-transfected cells or cells transfected with 10mg of each indicated plasmid DNA in two separate experiments, and 20mg RNA from each transfectant was analyzed on Northern blots, using an AAV-specific DNA probe. The three viral transcripts initiating from p5, p19, and p40 promoters are indicated. The same blots were stripped of the AAV probe and rehybridized with the GAPDH gene probe to ascertain the equivalence of RNA loads and transfer. Autoradiography was performed at270°C for 72 h.
FIG. 5. DNA slot blot analyses of efficiency of plasmid transfection in 293 cells. Twofold serial dilutions of 2mg each low-MrDNA isolated from
mock-transfected cells or cells mock-transfected with 10mg of each indicated plasmid DNA from the same two separate experiments described in the legend to Fig. 4 were analyzed on quantitative DNA slot blots, using an AAV-specific DNA probe as described in Materials and Methods. Autoradiography was performed at270°C for 2 h.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.65.274.68.337.2]scribed in Materials and Methods. Seventy-two hours post-transfection, cells were harvested and subjected to three rounds of freezing and thawing to release virus particles, which were purified on CsCl equilibrium density gradients followed by exhaustive digestion with DNase I as described in Materials and Methods. These stocks were used to infect HeLa cells in the presence of adenovirus. Low-MrDNA samples isolated at various times postinfection were analyzed on Southern blots, using AAV DNA as a probe. These results are shown in Fig. 7. Time-dependent accumulation of the characteristic mono-meric and dimono-meric replicative AAV DNA intermediates could be readily detected, indicating that biologically active progeny virions were indeed assembled following transfection of plas-mids pXS-70A and pSub201 in 293 cells in the absence of adenovirus. These data, although not quantitative, provide fur-ther evidence that a productive life cycle of AAV can be accomplished in 293 cells in the absence of a helper virus (42, 65).
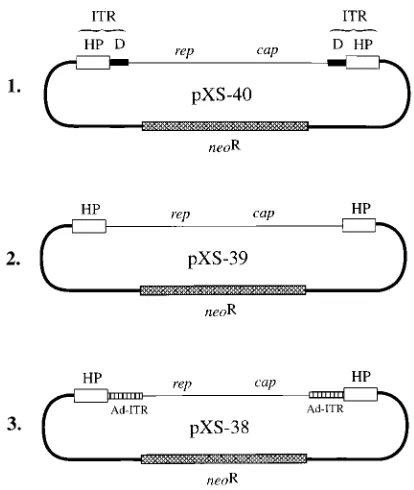
Autonomous replication of AAV affects stable transduction of recombinant plasmids in 293 cells.We next wished to de-termine the consequences of rescue and autonomous replica-tion of the AAV genome from recombinant plasmids contain-ing a selectable marker gene on stable transduction. The three recombinant plasmids pXS-38, pXS-39, and pXS-40, shown schematically in Fig. 8, were constructed as described in Ma-terials and Methods. Plasmid pXS-40 contains the intact wt AAV genome. Plasmid pXS-39 contains an AAV genome from which the D sequences have been deleted, and plasmid pXS-38 contains an AAV genome in which the D sequences have been replaced by the adenovirus ITRs. Each of the plasmids also contains the herpesvirus (TK-neor) gene in the vector back-bone. The efficiency of rescue and replication of the AAV genome from these plasmids in adenovirus-infected cells varies
[image:5.612.100.243.67.313.2]greatly, ranging from very high (pXS-40) to low (pXS-39) due to D-sequence deletions (59–61). Little or no rescue and rep-lication of the AAV genome occur from plasmid pXS-38 (data not shown). These plasmids were transfected into HeLa, KB, and 293 cells separately under identical conditions, and G418-resistant colonies were enumerated as described in Materials and Methods. These results are shown in Table 1. It is evident that the total numbers of G418-resistant colonies obtained with all three plasmids in HeLa or KB cells were not signifi-cantly different for the given cell types, although the transduc-tion efficiency in HeLa cells was approximately threefold higher than that in KB cells. In 293 cells, however, the numbers of G418-resistant colonies obtained with these plasmids varied significantly. We interpret these results as follows. Since no rescue and replication of the AAV genome from any of the plasmids occur in HeLa or KB cells (data not shown), the plasmid integrity is maintained, leading to stable integration. Plasmid pXS-38, from which the AAV genome does not un-dergo rescue and replication in 293 cells, is highly efficient in yielding G418-resistant colonies. With plasmid pXS-39, from which a low-level rescue and replication of the AAV genome occur in 293 cells, there is approximately fourfold reduction in the number of G418-resistant colonies, and with plasmid pXS-40, from which the AAV genome undergoes efficient rescue and replication, the numbers of G418-resistant colonies are further reduced approximately fivefold. These data are consis-tent with the conclusion that rescue and autonomous replica-tion of the AAV genome from recombinant plasmids in 293 cells compromise the structural integrity of the transfected plasmid, leading to reduction in functional DNA molecules capable of stable integration into the host chromosome (33).
FIG. 6. Southern blot analyses of rescue and replication of AAV genomes containing various mutations and/or deletions in the RBS and YBS in HeLa cells in the absence or presence of the adenovirus E1A gene products. Ten micro-grams of each indicated plasmid DNA was transfected separately in HeLa cells, either in the absence (2pAd2-E1A) or in the presence (1pAd2-E1A) of an adenovirus E1A expression plasmid. Low-MrDNA isolated at 72 h
posttrans-fection was digested with DpnI and analyzed as described in the legend to Fig. 2.
FIG. 7. Southern blot analyses of replication of the AAV genome in second-ary infections following autonomous rescue, replication, and encapsidation in 293 cells. Ten-micrograms of each of plasmids pXS-70A and pSub201 DNA was transfected separately in 293 cells, and progeny virions were purified on CsCl equilibrium density gradients as described in Materials and Methods. The viral stocks were used to infect HeLa cells in the presence of adenovirus. Low-Mr
DNA isolated at 24 h (lanes 1 and 4), 48 h (lanes 2 and 5), and 72 h (lanes 3 and 6) postinfection was analyzed on Southern blots, using an AAV-specific DNA probe. The monomeric (m) and dimeric (d) replicative forms and the single strands of the AAV genome (ss) are indicated. Autoradiography was performed at270°C for 2 h.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.372.487.441.634.2]DISCUSSION
More than a decade ago, Yakobson et al. (65) first docu-mented that AAV is capable of undergoing a completely pro-ductive replication in the absence of a helper virus, albeit inefficiently, in certain cell types treated with various genotoxic stress. Since expression from the viral p5 promoter and the interaction of AAV Rep proteins with the viral regulatory elements play a crucial role in a productive infection by AAV, Kyo¨stio¨ et al. (24) and Ni et al. (35) investigated these steps in the virus life cycle, but those studies were carried out in the absence of AAV DNA replication. We undertook the present study to delineate the role of AAV Rep proteins and their interaction with the regulatory elements in the p5 promoter in the context of a complete wt viral genome. Consistent with previous studies, our data indicate that mutations in the RBS relieve Rep protein-mediated suppression of expression of the viral p5 transcripts and that mutations in the YBS augment
Rep protein-mediated suppression of expression from the p5 promoter (24, 37).
More interestingly, our data document that AAV is capable of autonomous replication, even in the absence of any geno-toxic stress, but only in 293 cells. Autonomous replication of AAV in 293 cells was not detected during infection studies when a multiplicity of infection of 20 (approximately 2,000 viral genomes/cell) was used (65). We suspect that we detected a low-level replication of AAV in the absence of adenovirus because substantially larger numbers (.500,000 viral genomes/ cell) were delivered in 293 cells during plasmid-mediated transfections (39, 40). Abundant expression of viral transcripts occurs from transfected recombinant plasmids, even in the absence of adenovirus, which does not lead to efficient repli-cation of viral genomes (25), suggesting suboptimal transport to the cytoplasm and/or inefficient translation of these tran-scripts. In the presence of adenovirus, on the other hand, these processes are carried out more efficiently (46). The fact that the adenovirus early gene products alone did not support res-cue and replication of the AAV genome in HeLa cells in the absence of adenovirus suggests that additional factors other than the adenovirus E1A gene products are required for the autonomous replication of AAV DNA. The observation that even in 293 cells no rescue and/or replication occurred from a recombinant plasmids that contained a mutation or a deletion in the YBS indicates that factors other than E1A and E1B are required for autonomous replication.
The regulation of expression from the AAV promoters in general, and that from the p5 promoter in particular, is com-plex (24, 25, 37, 38). The roles of two control elements in the p5 promoter, the RBS and the YBS, have been studied in some detail (24, 37). Although the RBS, which is localized between the p5 TATA box and the transcription start site (Fig. 1), appears to be crucial in effectively repressing expression from the p5 promoter during a natural infection by AAV, detectable levels of transcripts from each of the AAV promoters could be obtained in 293 cells following plasmid transfections, an ob-servation consistent with previously published reports (23, 25). Although the latter means might not represent a physiologi-cally natural situation, it provides some explanation for why AAV is unable to undergo autonomous replication: presum-ably because a threshold concentration of the viral Rep pro-teins fails to accumulate. This contention is borne out by the fact that there was a strong correlation between the levels of the p5 transcripts and the ability of AAV to replicate autono-mously in 293 cells (Fig. 2 and 4). With reference to the YBS, we focused our studies only on the site which is localized downstream from the p5 TATA box and the RBS because a second YBS, which is present upstream of the p5 TATA box, binds to YY1 and mediates repression of expression from the p5 promoter in the absence of adenovirus, and the adenovirus E1A-YY1 complex formation relieves this repression (6, 50, 51).
In sum, our studies document that the Rep protein interac-tion with the RBS plays a dominant role in down-regulating viral gene expression from the p5 promoter, and perturbation in this interaction is sufficient to confer autonomous replica-tion competence to AAV in 293 cells. Since autonomous rep-lication of the AAV genome does not occur in HeLa cells, even in the presence of adenovirus early gene products, these stud-ies also suggest that additional, hitherto unknown viral and/or cellular factors are required for a productive infection by AAV.
FIG. 8. Schematic structures of recombinant AAV plasmids containing the neorselectable marker gene and deletion or substitution in the D sequence in the
viral ITRs. The hairpin (HP) and D sequences are denoted by the open and closed boxes, respectively. The adenovirus ITRs are denoted by boxes with vertical lines, and the neorgene is represented by cross-hatched boxes. Each of
[image:6.612.65.272.65.310.2]the recombinant plasmids was constructed as described in Materials and Meth-ods.
TABLE 1. Effects of rescue and autonomous replication of the AAV genome from recombinant plasmids on stable
transfection in HeLa, KB, and 293 cellsa
Cell line Total no. of G418-resistant colonies from three dishes pXS-38 pXS-39 pXS-40
HeLa 840 752 932
KB 357 248 220
293 1,272 300 60
aEach of the recombinant AAV plasmids containing the neorgene was
trans-fected separately in triplicate in HeLa, KB, and 293 cells under identical condi-tions, and G418 was added at an active final concentration of 400mg/ml 48 h posttransfection. Total numbers of G418-resistant colonies were enumerated from three dishes each 14 days postselection.
on November 9, 2019 by guest
http://jvi.asm.org/
ACKNOWLEDGMENTS
We thank Richard J. Samulski for providing plasmid pSub201. This research was supported in part by the Public Health Service grants (HL-48342, HL-53586, HL-58881, and DK-49218, Centers of Excellence in Molecular Hematology) from the National Institutes of Health and by a grant from the Phi Beta Psi sorority. A.S. was sup-ported by an Established Investigator Award from the American Heart Association.
REFERENCES
1. Ashktorab, H., and A. Srivastava. 1989. Identification of nuclear proteins that specifically interact with adeno-associated virus type 2 inverted terminal repeat hairpin DNA. J. Virol. 63:3034–3039.
2. Berns, K. I. 1990. Parvovirus replication. Microbiol. Rev. 54:316–329. 3. Berns, K. I., and R. A. Bohenzky. 1987. Adeno-associated viruses: an update.
Adv. Virus Res. 32:243–306.
4. Berns, K. I., R. M. Kotin, and M. A. Labow. 1988. Regulation of adeno-associated virus DNA replication. Biochim. Biophys. Acta 95:425–429. 5. Buller, R. M., J. E. Janik, E. D. Sebring, and J. A. Rose. 1981. Herpes
simplex virus types 1 and 2 completely help adeno-associated virus replica-tion. J. Virol. 40:241–247.
6. Chang, L.-S., Y. Shi, and T. Shenk. 1989. Adeno-associated virus p5 pro-moter contains an adenovirus E1A-inducible element and a binding site for the major late transcription factor. J. Virol. 63:3479–3488.
7. Cheung, A. K. M., M. D. Hoggan, W. W. Hauswirth, and K. I. Berns. 1980. Integration of the adeno-associated virus genome into cellular DNA in latently infected human Detroit 6 cells. J. Virol. 33:739–748.
8. Giraud, C., E. Winocour, and K. I. Berns. 1995. Recombinant junctions formed by site-specific integration of adeno-associated virus into an episome. J. Virol. 69:6917–6924.
9. Hauswirth, W. W., and K. I. Berns. 1977. Origin and termination of adeno-associated virus DNA replication. Virology 78:488–499.
10. Hermonat, P. L., M. A. Labow, R. Wright, K. I. Berns, and N. Muzyczka. 1984. Genetics of adeno-associated virus: isolation and preliminary charac-terization of adeno-associated virus type 2 mutants. J. Virol. 51:329–339. 11. Hirt, B. 1967. Selective extraction of polyoma DNA from infected mouse cell
cultures. J. Mol. Biol. 26:365–367.
12. Hong, G., P. Ward, and K. I. Berns. 1992. In vitro replication of adeno-associated virus DNA. Proc. Natl. Acad. Sci. USA 89:4673–4677. 13. Hong, G., P. Ward, and K. I. Berns. 1994. Intermediates of adeno-associated
virus DNA replication in vitro. J. Virol. 68:2011–2015.
14. Im, D.-S., and N. Muzyczka. 1989. Factors that bind to adeno-associated virus terminal repeats. J. Virol. 63:3095–3104.
15. Im, D.-S., and N. Muzyczka. 1990. The AAV origin binding protein Rep68 is an ATP-dependent site-specific endonuclease with DNA helicase activity. Cell 61:447–457.
16. Im, D.-S., and N. Muzyczka. 1992. Partial purification of adeno-associated virus Rep78, Rep52, and Rep40 proteins and their biochemical character-ization. J. Virol. 66:1119–1128.
17. Kotin, R. M., and K. I. Berns. 1989. Organization of adeno-associated virus DNA in latently infected Detroit 6 cells. Virology 170:460–467.
18. Kotin, R. M., R. M. Linden, and K. I. Berns. 1992. Characterization of a preferred site on human chromosome 19q for integration of adeno-associ-ated virus DNA by nonhomologous recombination. EMBO J. 11:5071–5078. 19. Kotin, R. M., J. C. Menninger, D. C. Ward, and K. I. Berns. 1991. Mapping and direct visualization of a region-specific viral DNA integration site on chromosome 19q13-qter. Genomics 10:831–834.
20. Kotin, R. M., M. Siniscalco, R. J. Samulski, X. Zhu, L. Hunter, C. A.
Laughlin, S. McLaughlin, N. Muzyczka, M. Rocchi, and K. I. Berns.1990.
Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci. USA 87:2211–2215.
21. Kube, D. M., S. Ponnazhagan, and A. Srivastava. 1997. Encapsidation of adeno-associated virus type 2 Rep proteins in wild-type and recombinant progeny virions: Rep-mediated growth inhibition of primary human cells. J. Virol. 71:7361–7371.
22. Kube, D. M., and A. Srivastava. 1997. Quantitative DNA slot blot analysis: inhibition of DNA binding to membranes by magnesium ions. Nucleic Acids Res. 25:3375–3376.
23. Kyo¨stio¨, S. R. M., R. A. Owens, M. D. Weitzman, B. A. Antoni, N.
Che-janovsky, and B. J. Carter.1994. Analysis of the adeno-associated virus
(AAV) wild-type and mutant Rep proteins for their ability to regulate neg-atively AAV p5 and p19 mRNA levels. J. Virol. 68:2947–2957.
24. Kyo¨stio¨, S. R. M., R. S. Wonderling, and R. A. Owens. 1995. Negative regulation of the adeno-associated virus (AAV) p5 promoter involves both the p5 rep binding site and the consensus ATP-binding motif of the AAV Rep68 protein. J. Virol. 69:6787–6796.
25. Li, J., R. J. Samulski, and X. Xiao. 1997. Role of highly regulated rep gene expression in adeno-associated virus vector production. J. Virol. 71:5236– 5243.
26. Lusby, E., K. H. Fife, and K. I. Berns. 1980. Nucleotide sequence of the
inverted terminal repetition in adeno-associated virus DNA. J. Virol. 34: 402–409.
27. McCarty, D. M., D. J. Pereira, I. Zolotukhin, X. Zhou, J. H. Ryan, and N.
Muzyczka.1994. Identification of linear DNA sequences that specifically
bind the adeno-associated virus Rep protein. J. Virol. 68:4988–4997. 28. McCarty, D. M., J. H. Ryan, S. Zolotukhin, X. Zhou, and N. Muzyczka. 1994.
Interaction of the adeno-associated virus Rep protein with a sequence within the palindrome of the viral terminal repeat. J. Virol. 68:4998–5006. 29. Mendelson, E., M. G. Smith, I. L. Miller, and B. J. Carter. 1988. Effect of
viral rep gene on transformation of cells by adeno-associated virus vector. Virology 166:612–615.
30. Muzyczka, N. 1992. Use of adeno-associated virus as a general transduction vector for mammalian cells. Curr. Top. Microbiol. Immunol. 158:97–129. 31. Nahreini, P., and A. Srivastava. 1989. Rescue and replication of the
adeno-associated virus 2 genome in mortal and immortal human cells. Intervirology
30:74–85.
32. Nahreini, P., and A. Srivastava. 1992. Rescue of the adeno-associated virus 2 genome correlates with alterations in DNA-modifying enzymes in human cells. Intervirology 33:109–115.
33. Nahreini, P., S. H. Larsen, and A. Srivastava. 1992. Cloning and integration of DNA fragments in human cells via the inverted terminal repeats of the adeno-associated virus 2 genome. Gene 119:265–272.
34. Nahreini, P., M. J. Woody, S. Z. Zhou, and A. Srivastava. 1993. Versatile adeno-associated virus 2-based vectors for constructing recombinant virions. Gene 124:257–262.
35. Ni, T. H., X. H. Zhou, D. M. McCarty, I. Zolotukhin, and N. Muzyczka. 1994. In vitro replication of adeno-associated virus DNA. J. Virol. 68:1128–1138. 36. Owens, R. A., and B. J. Carter. 1992. In vitro resolution of adeno-associated virus DNA hairpin termini by wild-type Rep protein is inhibited by a dom-inant-negative mutant of Rep. J. Virol. 66:1236–1240.
37. Pereira, D. J., D. M. McCarty, and N. Muzyczka. 1997. The adeno-associated virus (AAV) Rep protein acts as both a repressor and an activator to regulate AAV transcription during a productive infection. J. Virol. 71:1079– 1088.
38. Pereira, D. J., and N. Muzyczka. 1997. The cellular transcription factor SP1 and an unknown cellular protein are required to mediate Rep protein acti-vation of the adeno-associated virus p19 promoter. J. Virol. 71:1747–1756. 39. Ponnazhagan, S., X.-S. Wang, M. J. Woody, F. Luo, L. Y. Kang, M. L.
Nallari, N. C. Munshi, S. Z. Zhou, and A. Srivastava.1996. Differential
expression from p6 promoter of parvovirus B19 in human cells following plasmid transfection and recombinant adeno-associated virus 2 (AAV) in-fection: human megakaryocytic leukaemia cells are non-permissive for AAV infection. J. Gen. Virol. 77:1111–1122.
40. Ponnazhagan, S., M. J. Woody, X.-S. Wang, S. Z. Zhou, and A. Srivastava. 1995. Transcriptional transactivation of parvovirus B19 promoters in non-permissive human cells by adenovirus type 2. J. Virol. 69:8096–8101. 41. Qing, K. Y., B. Khuntirat, C. Mah, D. M. Kube, X.-S. Wang, S.
Ponnazha-gan, S. Z. Zhou, V. J. Dwarki, M. C. Yoder, and A. Srivastava. 1998.
Adeno-associated virus type 2-mediated gene transfer: correlation of ty-rosine phosphorylation of the cellular single-stranded D sequence-binding protein with transgene expression in human cells in vitro and murine tissues in vivo. J. Virol. 72:1593–1599.
42. Qing, K. Y., X.-S. Wang, D. M. Kube, S. Ponnazhagan, A. Bajpai, and A.
Srivastava.1997. Role of tyrosine phosphorylation of a cellular protein in
adeno-associated virus 2-mediated transgene expression. Proc. Natl. Acad. Sci. USA 94:10879–10884.
43. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a laboratory manual, p. 1.53–1.110. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
44. Samulski, R. J., K. I. Berns, M. Tan, and N. Muzyczka. 1982. Cloning of adeno-associated virus into pBR322: rescue of intact virus from the recom-binant plasmid in human cells. Proc. Natl. Acad. Sci. USA 79:2077–2081. 45. Samulski, R. J., L.-S. Chang, and T. Shenk. 1987. A recombinant plasmid
from which an infectious adeno-associated virus genome can be excised in vitro and its use to study viral replication. J. Virol. 61:3096–3101. 46. Samulski, R. J., and T. Shenk. 1988. Adenovirus E1B 55-Mrpolypeptide
facilitates timely cytoplasmic accumulation of adeno-associated virus mRNAs. J. Virol. 62:206–210.
47. Samulski, R. J., A. Srivastava, K. I. Berns, and N. Muzyczka. 1983. Rescue of adeno-associated virus from recombinant plasmids: gene correction within the terminal repeats of AAV. Cell 33:135–143.
48. Samulski, R. J., X. Zhu, X. Xiao, J. Brook, D. E. Houseman, N. Epstein, and
L. A. Hunter.1991. Targeted integration of adeno-associated virus (AAV)
into human chromosome 19. EMBO J. 10:3941–3950.
49. Senapathy, P., J.-D. Tratschin, and B. J. Carter. 1984. Replication of adeno-associated virus DNA: complementation of naturally occurring rep2mutants
by a wild-type genome or an ori2mutant and correction of terminal
palin-drome deletions. J. Mol. Biol. 179:1–20.
50. Seto, Y., Y. Shi, and T. Shenk. 1991. YY1 is an initiator sequence-binding protein that directs and activates transcription in vitro. Nature 354:241–245. 51. Shi, Y., E. Seto, L.-S. Chang, and T. Shenk. 1991. Transcriptional repression by YY1, a human GL1-Kru¨ppel-related protein, and relief of repression by
on November 9, 2019 by guest
http://jvi.asm.org/
adenovirus E1A protein. Cell 67:377–388.
52. Snyder, R. O., D.-S. Im, T.-H. Ni, X. Xiao, R. J. Samulski, and N. Muzyczka. 1993. Features of the adeno-associated virus origin involved in substrate recognition by the viral Rep protein. J. Virol. 67:6096–6104.
53. Snyder, R. O., R. J. Samulski, and N. Muzyczka. 1990. In vitro resolution of covalently joined AAV chromosome ends. Cell 60:105–113.
54. Srivastava, A. 1987. Replication of the adeno-associated virus DNA termini in vitro. Intervirology 27:138–147.
55. Srivastava, A., E. W. Lusby, and K. I. Berns. 1983. Nucleotide sequence and organization of the adeno-associated virus 2 genome. J. Virol. 45:555–564. 56. Tratschin, J.-D., I. L. Miller, and B. J. Carter. 1984. Genetic analysis of
adeno-associated virus: properties of deletion mutants constructed in vitro and evidence for an adeno-associated virus replication function. J. Virol.
51:611–619.
57. Urcelay, E., P. Ward, S. M. Weiner, B. Safer, and R. M. Kotin. 1995. Asymmetric replication in vitro from a human sequence element is depen-dent on adeno-associated virus Rep protein. J. Virol. 69:2038–2046. 58. Usheva, A., and T. Shenk. 1996. YY1 transcription initiator: protein
inter-actions and association with a DNA site containing unpaired strands. Proc. Natl. Acad. Sci. USA 93:13571–13576.
59. Wang, X.-S., S. Ponnazhagan, and A. Srivastava. 1995. Rescue and
replica-tion signals of the adeno-associated virus 2 genome. J. Mol. Biol. 250:573– 580.
60. Wang, X.-S., S. Ponnazhagan, and A. Srivastava. 1996. Rescue and replica-tion of adeno-associated virus type 2 as well as vector DNA sequences from recombinant plasmids containing deletions in the viral inverted terminal repeats: selective encapsidation of viral genomes in progeny virions. J. Virol.
70:1668–1677.
61. Wang, X.-S., K. Y. Qing, S. Ponnazhagan, and A. Srivastava. 1997. Adeno-associated virus type 2 DNA replication in vivo: mutation analyses of the D sequence in viral inverted terminal repeats. J. Virol. 71:3077–3082. 62. Wang, X.-S., and A. Srivastava. 1997. A novel terminal resolution-like site in
the adeno-associated virus type 2 genome. J. Virol. 71:1140–1146. 63. Ward, P., and K. I. Berns. 1991. In vitro rescue of an integrated hybrid
adeno-associated virus/simian virus 40 genome. J. Mol. Biol. 218:791–804. 64. Ward, P., E. Urcelay, R. M. Kotin, B. Safer, and K. I. Berns. 1994.
Adeno-associated virus DNA replication in vitro: activation by a maltose-binding protein/Rep68 fusion protein. J. Virol. 68:6029–6037.
65. Yakobson, B., T. Koch, and E. Winocour. 1987. Replication of adeno-asso-ciated virus in synchronized cells without the addition of a helper virus. J. Virol. 61:972–981.