Copyright© 1994, AmericanSociety forMicrobiology
A
Simian
Immunodeficiency Virus Envelope V3 Cytotoxic
T-Lymphocyte
Epitope in Rhesus
Monkeys
and
Its
Restricting
Major
Histocompatibility Complex
Class I
Molecule
Mamu-A*02
NORIAKI
WATANABE,'
STEPHEN N.McADAM,2
JONATHANE.BOYSON,2
MARIAN S. PIEKARCZYK,2YASUHIRO YASUTOMI,1 DAVID I.WATKINS,2 ANDNORMAN L.
LETVIN1*
HarvardMedicalSchool, Beth IsraelHospital, Boston, Massachusetts 02215,1and WisconsinRegional PrimateResearch Center and Departmentof Pathology, University ofWisconsin
MedicalSchool, Madison, Wisconsin 537152
Received 25 January 1994/Accepted 18July1994
The use ofthe simianimmunodeficiency virus(SIV) macaquemodel forassessinghumanimmunodeficiency
virusvaccine strategieswill befacilitatedbythecharacterizationofpredominantSIV
cytotoxic
T-lymphocyte(CTL) epitopes and their restricting majorhistocompatibilitycomplex (MHC) class I molecules inmacaque species.We now define a rhesusmonkey SIVmacCTLepitopein the thirdhypervariable regionof theenvelope glycoprotein of the virus. Thisepitope,YNLTMKCR,contains the first twoamino acidsofacysteine-cysteine loop which is the
SIVmac
analog ofthe human immunodeficiency virus type 1 V3 loop. We also employed one-dimensional isoelectricfocusingtocharacterize the MHC class I molecule of the rhesusmonkeythatbinds thisSIVmaC
envelopepeptide fragment.Cloningandsequencingthe cDNAencodingthis rhesusmonkeyMHC class I moleculedemonstratesthat it is anewly described HLA-Ahomolog, Mamu-A*02.This viral CTLepitopeand itsrestricting MHCclass I molecule willfacilitatethe useofthe
SIVmac
rhesusmonkeymodel for studies ofenvelope-basedvaccinestrategiesand forexploring AIDSimmunopathogenesis.Aneffective human immunodeficiency virus (HIV) vaccine should be capable of stimulating HIV-specific cytotoxic T
lymphocytes (CTL) (20, 34). Since HIV is spread at least in partthrough exposuretocell-associatedvirus,vaccine-elicited neutralizing antibody responses alone maynotprevent infec-tion with this virus. Considerable experimental evidence has implicated virus-specific CTL in the control of HIV infection.
Inanimals and inhumans, the early containment of AIDS virus replication appears to coincide with the development of
virus-specificCTL responses (27,28, 44). Moreover, the replication ofHIV inCD4+ lymphocytescanbe inhibitedby CD8+ cells withphenotypic characteristics of CTL (33, 35). It is, therefore, crucial that the evaluation ofHIVvaccine strategies includes assessing the elicitation of virus-specific CTL.
A number of HIV vaccine strategies currently being
ex-plored employ the envelope glycoprotein of the virus as the soleimmunogen(4).Theseapproaches include the use of live
vectors carrying the HIV env gene and recombinant HIV
envelope glycoprotein formulated with adjuvant. A model system in which to explore such envelope-based AIDS virus vaccines for the ability to elicit envelope-specific CTL would be extremely valuable.
The simianimmunodeficiency virus (SIV) macaque model has provenuseful in assessing vaccine strategies for the prevention ofHIVinfection (19). Our previous definition of a
predomi-nant
SIVmac
Gag CTL epitope and its restricting major histo-compatibility complex (MHC) class I allele Mamu-A*01 has facilitated the use of this model system for exploring vaccine approaches to the induction of AIDS virus-specific CTL re-sponses (23, 39). A detailed understanding of anSIVmac
*Corresponding author.Mailing address:
Harvard
Medical School,BethIsraelHospital, 330 Brookline Ave., Boston, MA 02215. Phone: (617)735-2766. Fax:(617)278-8210.
envelope epitopewould be of considerable value in devising strategiesforevaluatingtheefficacyof variousenvelope-based
vaccines. Moreover, the definition of further SIV CTL epitopes beyond thesingle definedGagpeptideand its restrict-ing MHC class I allele would greatly expand the ability to performAIDS immunopathogenesis studies with this
nonhu-manprimatemodel. Wenowdefinean
SIVmac
envelopeCTLepitopeand itsrestrictingMHC classImolecule in the rhesus
monkey. Interestingly,thisepitopeincludesaportionofthe V3
loop analog in the third hypervariable region of the
SIVmac
envelope.
MATERUILSANDMETHODS
Animals.Heparinized bloodwasobtainedfrom rhesus
mon-key 381 after infection with
SIVmac
isolate 251. This animalwasmaintained inaccordance with guidelines of the
Commit-tee onAnimalsforthe Harvard Medical School and the Guide
forthe Care and Useof Laboratory Animals (33a).
Generation of CTL effector cells. Peripheralblood lympho-cytes (PBL) isolated by Ficoll-diatrizoate density gradient
centrifugationwerecultured for 2 to 3 days at 1 x 106cells per mlwith 5
[Lg
ofconcanavalinAper ml in RPMI 1640(GIBCO, GrandIsland, N.Y.) supplemented with L-glutamine (2 mM),penicillin (50 U/ml), streptomycin (40 ,ug/ml), and 10% fetal calfserum (ICN Biomedicals, Costa Mesa, Calif.). This me-diumwasthen supplemented with human recombinant inter-leukin-2 at a concentration of 20 U/ml, and cultures were maintained another 3 days. For studies to assess the fine
specificityof the CTL epitope recognition, PBL cultures were established initially with either a pool of 4
SIVmac
Envsynthetic peptides, eachat a5-,ug/ml concentration, orwith 5 ,ugofasingle peptide alone with no concanavalinAadded.
Isolation ofCD8+ lymphocytes. CD8+ cells were isolated by incubating the cultured cells with CD8 monoclonal
anti-6690
on November 9, 2019 by guest
http://jvi.asm.org/
41
V)
.' 3
U)
20
af)
41U1)
a-0 5 10 15 20 25 30 35 40 45 50 >C
SlVmac Env peptide number
FIG. 1. SIVmacenvelope peptide recognition byPBLeffector cells from rhesus monkey 381. B-LCL were pulsed with a panel of 54
peptides representing sequential overlapping fragments of the entire
SIVmaCenvelope protein and assessedastargetpopulations ina5'Cr
releaseassaywith effector PBL from rhesus monkey381. Asacontrol
target, autologous B-LCLwereinfected witharecombinant vaccinia
virus-SIVmac Envconstruct (V-Env). An EST ratio of 80:1 was
em-ployed in this study.
body (7Pt3F9; 1:500 dilution of ascites; S. Schlossman,
Dana-Farber Cancer Institute, Boston, Mass.) and thensubjectedto
immunomagnetic bead separation (Dynal, Oslo, Norway). After furtherculture, the magnetic beadswereremoved from
thepurified CD8+ cells by magnet. The live cellswere then
isolated by Ficoll-diatrizoate density centrifugation and
as-sessedfor effector function.
Cytotoxicity assay. Target cellswere autologous or
alloge-neic B-lymphoblastoid cell lines(B-LCL), generated through herpesvirus papio immortalization of PBL,ortransfected ClR
cells (23). These cells were incubated at 37°C in a 5% CO2
atmosphere with S ,ug of peptide per ml or recombinant
vaccinia virus expressing SIVmac env or the irrelevant equine
herpesvirus gHgene(40) and 0.5 ,uCi of Na2CrO4perml for
16 h and then washed twice. Cultured PBL were used as
effector cells. 51Cr-labeledtarget cellswereincubated for 5 h
with effector cells at EST ratios of 80:1, 40:1, 20:1, and 10:1. Spontaneous release varied from 10to 20%. Specific release
was calculated as [(experimental release - spontaneous
re-lease)/(100% release - spontaneousrelease)] x 100.
SIVmaC
envelope peptides. Overlapping25-amino-acid pep-tidesnumbered 1through54weresynthesized bythe methodofHoughtonetal.(10), usingtheSIVmacenvelopesequenceof Franchini et al. (5). Each consecutive peptide overlaps the preceding oneby 8 amino acids.
Radioimmunoprecipitation. Aliquots (5 x 106 cells) from
the B-LCL were cultured in methionine-free medium for 30
min. [35S]methionine (200 ,Ci; ICNRadiochemicals, Irvine, Calif.; specific activity, 1,000 Ci/mmol)wasthen addedtoeach culture. At the end ofa5-h incubationperiod, the cellswere
pelleted and frozenat -70°C. Cellswerelysed for 30minon
ice in 500,lIoflysisbuffer(20mMTris-HCl[pH 8.0],0.15M
NaCl, 1 mM MgCl2, 0.1 mM phenylmethylsulfonyl fluoride, 5% TritonX-114). The Triton X-114 used in the lysis buffer had been precondensed three times by adding cold distilled
H20, heatingat37°Cfor 5min, separatingintodetergentand
aqueousphases by centrifugation at300 Xgatroom
temper-ature, anddiscarding theaqueous phase. Following lysis,the
sampleswere separatedintodetergentandaqueousphasesas
described above, the aqueous phase was discarded, and the lower detergent phase was brought up to 1 ml with NET-N
A
u) 30
~20 -|
>0
O
4U)10-none 17 18 19
Peptide number 20
B
V
30-
20-U)
10
none 18 19 cont
Peptide number
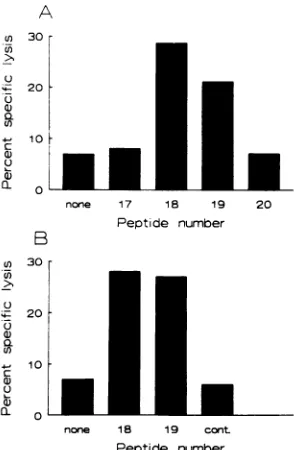
FIG. 2. Peptides 18 and 19arerecognized by CD8+ SIVmac
enve-lope-specific effector cells from rhesus monkey 381. (A) PBL of monkey 381werestimulated in vitro withSIVmacEnv peptides 17, 18,
19, and 20 and assessed for lysis of B-LCL pulsed with each individual peptide. (B)PBLofmonkey 381werestimulated in vitro withSIVmac Env 18, positively selected for CD8+ cells, and assessed for lysis of B-LCL pulsed with SIVmac Env peptides 18 and 19 and a control (cont.) peptide. 51Cr releasewasassessed in both studies at anE/T
ratio of 10:1.
(150mMNaCl, 5 mM EDTA, 50 mM Tris-HCl [pH 8.0], 0.5%
NonidetP-40;Sigma). Sampleswereprecleared bytwo
succes-siveincubationson30,ul (50%
[vol/voll)
of proteinA-Sepha-rose CL-4B beads(Sigma) previously saturated with an
irrel-evant ascites and then precleared on protein A-Sepharose
beads alone. Allpreclearing stepswerecarried outfor 1 h at
4°Cwith continuousshaking. Forimmunoprecipitation,50 ,ul
ofprotein-A Sepharosebeadswasincubated for 4 h with5 ,ul
ofW6/32ascites and washed twice with NET-N. Immunopre-cipitation was performed overnight, and the beads were
washed once with NET-N. Each immunoprecipitate was
treated with 20
[lI
of neuraminidase (10 U/ml; type VIII; Sigma)in0.1 M sodiumacetatebuffer(pH 5.5)-i mMCaCl2on ashaking platform at37°C for 3 h and then washed three times with NET-N.
1-D IEF. One-dimensional isoelectric focusing (1-D IEF)
wascarriedoutaspreviouslydescribed(37). Briefly,a0.75-mm
polyacrylamide gel was made with 5.4% ampholines (LKB,
Gaithersburg, Md.) comprising4%pH5to7,1%pH3.5to10,
and0.4%pH7to9.Immunoprecipitateswereresuspendedin IEFsamplebuffer(9.5Murea,4%[vol/vol] ampholine [pH3.5 to 10],5%2-mercaptoethanol, 2% Nonidet P-40,0.01%
bro-mophenol blue), and the gelwas run on a DNAsequencing
apparatus (SQU-2000; American Bionetics, Hayward, Calif.)
foratotal of 13 h, with thevoltage risingfrom 400to 1,500V ataconstantpowerof 10 W. Afterelectrophoresis,thegelwas
treated with IEF fixative (0.13 M sulfosalicylic acid, 0.7 M trichloroacetic acid, 30% methanol), fixed in 10% methanol and 7.5% acetic acid, treated with En3Hance (New England Nuclear, Boston, Mass.) for 60 min, washed for 15 min in distilled water, dried, and visualized by autoradiographywith XAR-5 film(Eastman Kodak, Rochester, N.Y.).
0
10
0-I I M
WLJJLJLLd
mlal
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.364.511.81.306.2] [image:2.612.67.294.81.229.2]TABLE 1. TheSIVmacenvelope CTL epitope (residues 306to313) iswell conserved among macaque andmangabeySIV isolatesa
Isolate Sequenceb Species
SIVMM251 YNLTMKCR Macaque(rhesus)
SIVMM32H --- Macaque (rhesus)
SIVMM142 --- Macaque (rhesus)
SIVMM239 --- Macaque (rhesus)
SIVMNE --- Macaque (pigtailed)
SIVMMlA1 ----I--- Macaque(rhesus)
SIVSTM ---s-- Macaque(stump tailed)
SIVSMMH4 ---R-- Sooty mangabey
SIVSMMPBJ ---R-- Sooty mangabey
SIVAGMTYO ---SVT-- African greenmonkey
HIV2ROD ---SLH-K Human
SIVMNDGB1 -G-KIE-H Mandrill
SIVSYK LR-QII-E Sykes' monkey
HIVlHXB2R TSVEIN-T Human
HIVMN ESVZIN-T Human
HIVSF2 ESVAIN-T Human
a Sequences (withaccession numbers shown inparentheses) wereobtained from the Human RetrovirusandAIDS databaseattheLos Alamos National Laboratory: SIVMM251 (M19499), SIVMM32A (D01065), SIVMM142
(M16403), SIVMM239(M33252),SIVMNE(M32742), SIVMMlAl(M76764),
SIVSTM(M83293),SIVSMMH4(X14307),SIVSMMPBJ(M31325), SIVAG-MTYO(X07805), HIV2ROD (M15390), SIVMNDGB1 (M27270), SIVSYK
L06042), HIVlHXB2R(K03455),HIVMN(M17449),and HIVSF2(K02007).
bDashes indicateidentitywith the SIVMM251 sequence.
RNA extraction, cDNA synthesis, and PCR. mRNA was
extracted from PBL of monkey 458 by using oligo(dT)25
Dynabeads (Dynal, Great Neck, N.Y.) accordingto the
man-ufacturer's instructions. PBL(3 x 106)werewashed with cold
phosphate-buffered saline andresuspended in 100
RI
oflysisbuffer(10mMTris [pH 7.5], 140 mM NaCl, 5 mM KCl, 1% NonidetP-40,3 ,u of RNasin[Promega, Madison, Wis.]).The
lysates were centrifuged, and the supernatants were trans-ferredto 1.2 mg of washedoligo(dT)25 beads. Aftera 4-min
incubation at room temperature, samples were magnetically
isolatedandwashed twice. Beadswereresuspendedin20 pIof
elution buffer (2 mM EDTA, pH 7.5) containing 1 pul of
RNasin,and thesuspensionwasheatedto65°Cfor 2minand
placed inthe magnet,after which the eluatecontaining poly(A)
RNA was withdrawn to a new tube on ice. cDNA synthesis
wasperformedunder thefollowingconditions. RNasin(3 pI),
0.5 mM (each) deoxynucleoside triphosphates (dNTP), 100 mMdithiothreitol, 30 pmol of 3' primer (3' PRH3; 5'-GCAA
GCTFIAGTCCCACACAAGGCAGCTG-3'), and 12U of
Su-perscript M-MLV (Gibco/BRL, Gaithersburg, Md.) were
30 05
._.
- 20 Li
10
o
303-1 303-2 303-3
transfected C 1 R targets
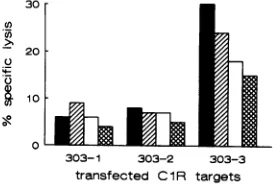
FIG. 3. Effector cells from rhesus monkey 381 lyse SIVmac
env-expressingClR cells transfected with the MHC class I cDNA Mamu-A*02 (303-3). Peptide 18-stimulated PBL from rhesus monkey 381
were assessed for the ability to lyseSIVmac env-expressing ClR cells transfected with threedifferent rhesus monkey MHC class I cDNAs. The
[image:3.612.61.301.102.267.2]assaywasdoneatEST ratios of 80:1 (R), 40:1 (0), 20:1 (C]), and 10:1 (X).
FIG. 4. ClR cellstransfected with Mamu-A*02(303-3)expressan
MHCclass I molecule that hasanisoelectricpointthat is identical to that ofan MHC class I molecule expressed by a B-LCL of rhesus monkey 458.
present.Sampleswereincubated for 1 hat37°Cand heatedto
94°Cfor 3 min.Ten microliters ofsampleswasemployedina PCRmixture that contained 50 mMKCl,10 mMTris-HCl,1.5 mMMgCl2, 0.01%gelatin,and 0.25 mM(each)dNTP. Primers
wereaddedasfollows: 60pmolof the 5'primerLPXI(5'-GC
CTCGAGATGACGGTCATGGCTCCCCGAACC-3') and 30
pmolof 3'primer (3'PRH3). Sampleswereheatedto95°Cfor 2 minand thencooledto 37°Cfor 5min, after which 5 U of
Taq polymerasewas added. Cycleconditions usedwere94°C
for 1 min20s;37°Cfor3min20s; and72°Cfor 10min. Subcloning, sequencing, and transfection. After amplifica-tion,thePCRproductwasligatedintopKG5,andthe first 200 bases encoding the leader peptide and part of the alpha-1 domain of plasmids with full-length inserts were sequenced
(29). Three distinct clones were isolated from the rhesus
monkey cDNA, and at least three copies of each of these
cDNAsweresequenced.ClRcellsweretransfected with 20 ,ug
oflinearizedplasmid by electroporation (0.2kV, 960 ,uF) and
selected in mediumcontaining 1.8mgof G418 (GIBCO,Life
Technologies, Inc.)per ,ul. Expressed MHC classI molecules wereprecipitated withW6/32andfocused by1-D IEF.
RESULTS
Definition ofSIVmacenvelope CTL epitopeinvirus-infected rhesus monkey. Since CD8+ CTL recognize fragments of endogenously synthesized viral protein expressed in
associa-tion with MHC classImoleculesonthe surface of autologous
infectedcells,theprecise regionsofaviralproteinrecognized
byaspecific CTL population canbe determinedby assessing
CTLrecognition oftarget cellspulsedwith syntheticpeptide fragments representing regions of viral protein (30, 32). PBL
-J
uoo I
t ro)
__.r3C
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.113.250.581.673.2]LeaderPeptide Alpha 1
C CTCCTCCTG GTG CTC TCA GGG GCC CTGGCC CTG ACT CAG ACCCGGGCA GGC TCT CACTCC ATG AGG TATTTCTAC ACC TCC L L L V L S G A L A L T Q T R A G S H S M R Y F Y T S ATG TCC CGG CCCGGC CGC TGG GAG CCC CGCTTC ATC GCCGTG GGC TAC GTG GAC GACACG CAGTTC GTG CGGTTCGAC AGC
M S R P G R W E P R F I A V G Y V D D T Q F V R F D S GAC GCC GCG AGC CAGAGGAGGAGCCG CGG GCG CCG TGGGTGGAG CAG GAGGGTCCAGAG TATTCG GACCGGGAG ACACGG
D A A S Q R H E P R A P W V E Q E G P E Y W D R E T R
Alpha 2 AACATGAAG GCCGAG ACA CAG AAT GCC CCAGTGAACCTG CGG AAC CTG CGC GGC TACTAC AAC CAG AGC GAG GCC GGG TCT N M K A E T Q N A P V N L R N L R G Y Y N Q S E A G S CAC ACC ATC CAGAGG AT TAC GGC TGCGACCTGGGGCCG GACGGG CGC CTC CTC CGCGGG TAT CAC CAGTCC GCC TAC GAC
H T I Q R M Y G C D L G P D G R L L R G Y H Q S A Y D GGCAAGGATTAC ATC GCCCTG AAC GAG GACCT CGCTCCTGG ACC GCC GCG GACAM GCG GCT CAGAAC ACC CAG CGCAAG G K D Y I A L N E D L R S W T A A D M A A Q N T Q R K TGGGAG GCG GCGGGT GAG GCG GAG CAG CAC AGA ACC TAC CTG GAG GGC GAGTGCCTGGAG TGGCTC CGC AGA TAC CTGGAG
W E A A G E A E Q H R T Y L E G E C L E W L R R Y L E
Alpha3
AACGGGAAG GAGACG CTGCAG CGCGCG GAC CCCCCC AAG ACA CATGTGACC CAC CACCCC GTC TCT GACCAAGAG GCC ACC N G K E T L Q R A D P P K T H V T H H P V S D Q E A T CTGAGGTGCTGG GCC CTG GGC TTCTAC CCT GCG GAG ATC ACACTGACC TGG CAG CGGGATGGAGAG GACCAAACT CAG GAC
L R C W A L G F Y P A E I T L T W Q R D G E D Q T Q D ACG GAG CTCGTGGAG ACC AGG CCTGCAGGG GACGGAACCTTCCAG AAGTGGGCG GCTGTG GTG GTGCCTTCTGGAAAGGAG
T E L V E T R P A G D G T F Q K W A A V V V P S G K E
Tranamembran.
CAG AGATAC ACC TGT CATGT CAG CAT GAG GGTCTG CGTGAG CCCCTC ACCCTG AGATGG GAG CCG TCTTCCCAG TCCACC Q R Y T C H V Q H E C L R E P L T L R W E P S S Q S T ATCCTC ATCGTGGGC ATCATT'GCTGGCCTl GTTCTCCTT'GGAATT' GIG GTC ATT GGA GCTGIG ATT GCTGCTG1GATATOG
I L I V G I I A G L V L L G I V V I G A V I A A V I W Cytoplasmic Dosainj
AGG AGG AAG AGCTCAGATAGA AMA GGA GGG AGC TAC TCT CAG GCT GCA AGCAGTGAC AGT GCC CAG GGC TCT GATGIG TCT R R K S S D R K G G S Y S Q A A S S D S A Q G S D V S CTC ACGGCTTGT AAAGO MTA
L T A C K V
FIG. 5. Nucleotide and predicted amino acid sequences of Mamu-A*02. The predicted amino acid sequence is shown below the nucleotide sequence for clone303-3.
fromSIVmac-infectedmonkey 381 were assessed for the ability
to lyse B-LCL pulsed withapanel of25-amino-acid peptides representing sequential, overlapping fragments of the entire
SIVmac
Envprotein. The B-LCLemployedastargetsincluded both autologous and allogeneic cells. The effector cells frommonkey 381recognized the adjacent Env peptidesp18andp19
pulsed ontoboth autologous B-LCL and one of the allogeneic B-LCL, cell line 458 (Fig. 1 anddata not shown). p37-pulsed targets were also lysed by these effector cells, although less
efficiently (Fig. 1).
Toconfirm the fine specificityof the Envpeptide 18 and19 recognition, PBLofmonkey381 werestimulated in vitro with the four adjacent, overlapping
SIVmac
Env peptides, p17 to p20, andassessed for theability tolyse458 B-LCLpulsed with each of these individualpeptides (Fig. 2A). Significant lysisof B-LCL pulsed with either p18 or p19 was observed.p18-stimulated PBLofmonkey 381, positively selected forCD8+
cells by using immunomagnetic beads, lysed B-LCL pulsed
withp18 orp19 (Fig. 2B). Therefore, effectorcellrecognition
of these peptidesis mediated by CD8+ cells. These observa-tions suggested that the
SIVmac
Env epitope lies in theoverlapping 8 amino acids shared
by peptides
p18
andp19,
YNLTMKCR. This sequence includes a portion of the third hypervariableregionof
SIVmac
Env.Itis well conserved among macaqueandmangabeySIVisolates (Table 1).Cloningandsequencingof therhesusmonkeycDNA
encod-ing the MHC class I molecule that presents
SIV.c
Envpeptide 18 toCD8+ CTL. To characterize further the interac-tion leading to CD8+ CTL recognition of
SIVmac
Envin the infected monkey381, the MHC class I molecule that binds p18 and p19 and presents these peptides to CD8+ CTL wascharacterized. Three distinct MHC classIcDNAswerecloned from the 458B-LCL, the allogeneic rhesus monkey B-LCL that binds these peptides and presents them to the CTL of monkey 381. These cDNAs were transfected into ClR cells. p18-stimulated PBL from
SIVmac-infected
rhesusmonkey381werethen assessed for theabilitytolyse each of thetransfected ClR cells lines following infection with the vacciniavirus-SIVmac
envelope. The effector cellslysed the cell line transfected with the 303-3 clone butnotthose transfected with the other rhesus monkey MHC class IcDNAs (Fig. 3).
The moleculeencoded by this cDNAwascharacterized by
1-DIEF (Fig. 4),and theMHCclass IcDNA wassequenced (Fig. 5 and6).Comparisonof the nucleotide sequence ofthis cDNA to otherprimateMHC class Isequencesrevealed that the 303-3 cDNA was most closelyrelated to alleles of theA
locus.Accordingly, this allelewasrenamedMamu-A*02,
con-sistent with the 1990 nomenclature proposal of Klein et al. (15).ThesimilarityofMamu-A*02tootherA-locus alleleswas especially evident in the exons
encoding
the transmembrane andcytoplasmic domains. Alleles of theB locus are 9 nucle-otides shorter than alleles of the A locus, and many alleles ofMamu-A*02 EPSSQSTILIVGIIAGLVLLGIVVIGAVIAAVIWRRKSS DRKGGSYSQAASSDSAQGSDVSLTACKV*
Mamu-A*01 --F---PM---A--T---V---M--- ---*
Mamu-B*01 ---P----V---AV-AV--T---V---M--- GG---*
L
Patr-C*01 G----P--P----A---AV-AVA-L---V-V-MC--- GG----C---N---E--I---A*
FIG. 6. Mamu-A*02 isanHLA-Ahomolog.The Mamu-A*02 sequence iscomparedtoseveralprimate HLA-A,-B,and-C
homologs,
including Mamu-A*01 (23), Mamu-B*01 (ourunpublished observations), and Patr-C*01 (16). The sequencesshown representthe transmembrane and cytoplasmicdomains of nonhumanprimateMHC class I molecules. Dashesindicateidentitywith Mamu-A*02.on November 9, 2019 by guest
http://jvi.asm.org/
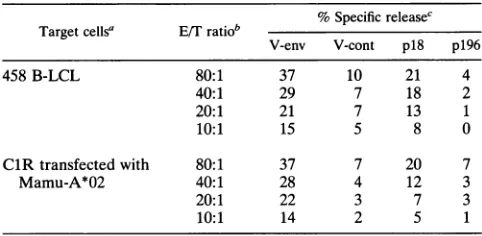
TABLE 2. SIVmacEnvp18-pulsedClR cellstransfectedwith Mamu-A*02 arelysed by CTL fromrhesus monkey381
NoSpecific releasec
Targetcellsa E/Tratiob
V-env V-cont p18 p196
458 B-LCL 80:1 37 10 21 4
40:1 29 7 18 2
20:1 21 7 13 1
10:1 15 5 8 0
ClR transfected with 80:1 37 7 20 7
Mamu-A*02 40:1 28 4 12 3
20:1 22 3 7 3
10:1 14 2 5 1
aTarget cells were B-LCL thatexpressedthe MHC class Imolecule
Mamu-A*02(458B-LCL)orClR cells transfected withMamu-A*02.
bEffector cells generatedfrom PBL of theSIVmac-infectedrhesusmonkey381 were stimulatedwithconcanavalin A andexpandedininterleukin-2-containing
medium.
c Target cells were infectedwith vacciniavirus-SIVmacEnv(V-env)orvaccinia
viruscarryingthe controlequineherpesvirus gHgene(V-cont)or werepulsed
with theSIVmacenvelopep18 peptideor acontrolHIV-1 reversetranscriptase peptide (IAKEPVHGV)(p196).
the C locus have a 3-nucleotide insertion in exon 5. As
expected,thepredictedaminoacid sequence of the
transmem-brane andcytoplasmicdomains of Mamu-A*02 is also similar
tothepredictedamino acid sequences of allelesof theAlocus
(Fig. 6).
Finally, we demonstrated that Mamu-A*02 is therestricting
MHCclass Imolecule for thisdefined
SIVmac
Envepitope in the rhesus monkey. Concanavalin A-stimulated PBL from monkey 381wereassessed inparallelfor theabilitytolyse458 B-LCL andthe Mamu-A*02-transfected ClR cell line follow-ingp18pulsing(Table 2). Equivalentp18-specific lysisof these target cells was observed. Thus, the 8-amino-acid peptidefragment spanning residues 306to313 of
SIVmac
Env canbe bound by the rhesus monkey MHC class I moleculeMamu-A*02 and presented toCD8+ CTL.
DISCUSSION
The immune response to HIV type 1 (HIV-1) gpl20 is of central importance in containing anAIDS virus infection (3, 6). Moreover, accumulating data are pointing to the impor-tance of the cell-mediated immune response in blocking the spread of the AIDS virus(28, 44).Nevertheless, little is known about the cell-mediated immune response to the AIDS virus
envelope glycoprotein. Surprisingly few HIV-1 gpl20 CTL
epitopes have been confirmed employing cloned effector T
lymphocytes(13, 14, 15a). This might be attributable, at least in part, to technical difficulties that arise as a result of extreme sequence variation in large portions of the HIV-1 envelope that occurs through mutation (7). Because of envelope se-quence variation among HIV isolates, it is difficult to match virus-elicited CTL responses with synthetic peptides employed
todefine CTL recognition epitopes. Our ability to study CTL
epitoperecognition in rhesus monkeys by employing an AIDS virus with a known envelope sequence and synthetic peptides generatedto match that sequence provides a powerful model forcircumventing thistechnical problem.
The CTL epitope suggested by the results of this study is composed of amino acid residues 306 to 313, which includes the first two amino acids of the V3-loop analog in the third hypervariable region of the
SIVmac
envelope glycoprotein. Theearly, type-specificHIV-1-neutralizing activity in the serum of
infected humans appears to be mediated by antibodies that
recognizetheloopstructureinthe third
hypervariable
domain of the HIVenvelope.For this reason, tremendous interest has focused upon this region of the envelope glycoprotein as atarget forvaccine-mediatedprotective immunity. Thisregion
of HIV-1 also has been implicatedin the cell tropismof the virus
(1, 25).
Accumulating data, however,
suggest that theanalogous region of
SIVmac
is not the principal neutralizingdomainof
SIVmae (11).
Moreover,thisregion
of theenvelope
does not appear to dictate the celltropism
ofSIVmac
(24).
Interestingly, recent studies suggest that this
portion
of theSIVmac
envelope doesplay
animportant
role indetermining
the pathogenicity of the virus in macaques (9). Our demon-stration of a predominantSIVmac
Env CTLepitope
in thisregionof theenvelope
glycoprotein
inMamu-A*02+monkeys
should further facilitate theuseof theSIVmac
rhesusmonkey
model for studies ofenvelope-dependent
issues of AIDSimmunopathogenesis.
Many of the MHC class I loci are conserved in macaques and
humans,
making
SIV infection of rhesusmonkeys
arelevant model for
investigating
the cell-mediated immune responsetothe AIDS virus in humans(38).
Mamu-A*02 is the second example ofanA-locus allelecloned,
sequenced,
andexpressed from Old World
primates.
A-locus alleles havepreviouslybeenclonedand
sequenced
fromchimpanzees
(16,
21),
gorillas(17,
36),
orangutans,gibbons
(2),
and rhesusmonkeys
(23).
The transmembrane andcytoplasmic
domainsarewell conserved among A-locus alleles in these nonhuman
primate
species (Fig.
6) (38).
HLA-B-like lociarealso present in rhesusmonkeys
(data
notshown),
and alleles of HLA-E-like(data
notshown)
and HLA-F-like(26)
loci from macaques have also beencloned. Evidence forhomologs
of HLA-C andHLA-G,
however,
has been difficultto obtainby
PCRampli-fication of both
genomic
DNAand cDNAderived from RNA from avariety
of tissues.Only in-depth
analysis
ofagenomic
library will determine whether these two loci are indeed present intherhesus
monkey.
The definition of this CTLepitopeand its
restricting
MHC class I molecule willsignificantly
facilitatefurther use of theSIVmac
model in AIDS research. We havepreviously
employed
the single defined Gag CTL epitope in anSIVmac-infected
monkey to explore live-vector- andpeptide-based
vaccinestrategies
forinducing
AIDSvirus-specific
CTL(22, 31, 42,
43).
Wehave also madeuseof thisdefinedepitope
inexploring
theimmunopathogenesis
of AIDS virus infections(27,
41).
Thedefinition ofan
SIVmac
envelopeepitope
inMamu-A*02+ rhesusmonkeys
will facilitate the use of theSIVmac
rhesusmonkey model for studies ofenvelope-based vaccine strate-gies.
ACKNOWLEDGMENTS
WethankShelleyKotlikoff forpreparingthemanuscript.
This work was supported by NIH grants AI-20729, AI-35351, AI-32426, CA-50139, DK-43351, RR-00167,and RR-00168.
REFERENCES
1. Chang-Mayer, C., M. Quiroga, J.W. Tung, D. Dina, andJ.A. Levy.1990. Viraldeterminantsof humanimmunodeficiencyvirus type 1T-cellormacrophage tropism,cytopathogenicity,andCD4
antigenmodulation.J. Virol. 64:4390-4398.
2. Chen, Z.W., S. N.McAdam, A. L.Hughes, A.L. Dogan,N. L. Letvin,and D.I.Watkins.1992. Molecularcloningof orangutan andgibbonMHCclass I cDNAs: the HLA-A and -B locidiverged
over30 million years ago. J. Immunol. 148:2547-2554.
3. Emini,E.A.,W. A.Schleif, J.H.Nunberg,etal.1992. Prevention of HIV-1 infection inchimpanzees by gpl20V3domain-specific
monoclonalantibody. Nature(London)355:728-730.
on November 9, 2019 by guest
http://jvi.asm.org/
4. Fast,P.E.,and M. C. Walker.1993. Human trials of experimental AIDS vaccines. AIDS1992/1993(Suppl.):S147-S159.
5. Franchini, G.,G.Gurgo, H.-G. Guo, R C. Gallo, E.Collalti,K. A. Fargnoli, L. F. Hall, F. Wong-Staal, and M. S. Reitz, Jr. 1987.
Sequenceofsimianimmunodeficiencyvirus from macaque and its
relationshiptoother humanand simianimmunodeficiency viruses. Nature(London) 328:539-543.
6. Girard, M.,M. P.Kieny,A.Pinter, et al. 1991. Immunization of
chimpanzees confers protection against challenge with human
immunodeficiencyvirus. Proc. Natl. Acad. Sci.USA 88:542-546. 7. Goodenow, M.,T. Nuet,W. Saurin, S. Kwok, J. Sinisky, and S.
Wain-Hobson. 1989. HIV-1isolatesarerapidlyevolving quasispe-cies: evidence for viral mixturesand preferrednucleotide substi-tutions. J.AcquiredImmuneDefic.Syndr. 2:344-352.
8. Goudsmit,J.,C. A. B.Boucher,RH.Meloen,etal.1988. Human immunodeficiency virus type 1 neutralization epitope with con-servedarchitectureelicitsearly type-specificantibodies in experi-mentally infected chimpanzees. Proc. Natl. Acad. Sci. USA 85: 4478-4482.
9. Hirsch,V. M., J. E. Martin, G. Dapolito, W. R. Elkins, W. T. London, S. Goldstein, and P. R Johnson. 1994. Spontaneous substitutions inthevicinityof theV3analog affect cell tropismand
pathogenicityofsimianimmunodeficiencyvirus.J.Virol. 68:2649-2661.
10. Houghton,R.A.,S. T.DeGraw,M. K.Bray,S. RHoffmann,and N. D.Frizell. 1986. Simultaneousmultiple peptide synthesis: the rapid preparation of a large numbers of discrete peptides for
biological, immunological, and methodologicalstudies.
BioTech-niques4:522.
11. Javaherian, K.,A.Langlois,S.Schmidt,M.Kaufmann,N.Cates, J. Langediik, R Meloen, R Desrosiers, D. Burns, and D. Bolognesi. 1992. The principal neutralization determinant of simianvirus differs from thathumanimmunodeficiencyvirustype 1. Proc. Natl.Acad. Sci. USA 89:1418-1422.
12. Javaherian, K.,A.J. Langlois,C.McDanal,etal.1989.Principal
neutralizingdomain of the humanimmunodeficiencyvirustype 1
envelope protein.Proc.Natl.Acad.Sci. USA 86:6768-6772. 13. Johnson, R P.,S. A.Hammond,A.Trocha,R F.Siliciano,and
B. D. Walker. 1994. Induction of a major histocompatibility complex class I-restricted cytotoxic T-lymphocyte response to a
highlyconservedregionof humanimmunodeficiencyvirustype 1
(HIV-1)gpl20inseronegativehumansimmunized witha candi-dateHIV-1vaccine. J.Virol.68:3145-3153.
14. Johnson, R P., A.Trocha,T. M. Buchanan,and B. D. Walker. 1993.Recognitionofahighlyconservedregion of human
immu-nodeficiencyvirustype 1 gp120 byanHLA-Cw4-restricted cyto-toxicT-lymphocyteclone.J.Virol. 67:438 445.
15. Klein, J., R E. Bontrop, R L. Dawkins, H. A. Erlich, U. V.
Gyllensten,E.RHeise,P. P.Jones,P.Parham,E. K.Wakeland, and D. I. Watkins. 1990. Nomenclature for the major
histocom-patibility complexes ofdifferent species: aproposal.
Immunoge-netics 31:217-219.
15a.Koup,R Personalcommunication.
16. Lawlor,D.A.,F. E.Ward,P. D.Ennis,A.P.Jackson,and P. Parham. 1988.HI-A-Aand Bpolymorphism predatethe divergenceof
hu-mansandchimpanzees.Nature(London)335:268-271.
17. Lawlor,D.A.,E.Warren,P.Taylor,and P. Parham. 1991.Gorilla class I major histocompatibility complex alleles: comparison to
human andchimpanzeeclassI. J.Exp.Med. 174:1491-1509. 18. Leonard,C. K.,M.W. Spellman,C.Riddle, R. V.Harris,J.N.
Thomas, and T. J. Gregory. 1990. Assignment of intrachain disulfide bonds and characterization ofpotential glycosylation sites of the type 1 recombinanthuman immunodeficiencyvirus
envelopeglycoprotein(gpl20) expressedinChinesehamster ovary cells. J. Biol. Chem. 265:10373-10382.
19. Letvin, N. L. 1990. Animal models for AIDS. Immunol. Today 11:322-326.
20. Letvin,N.L.1993.Vaccinesagainstthehuman
immunodeficiency
viruses:progressand prospects. N.Engl.J.Med.329:1300-1405. 21. Mayer,W.E.,M.Jonker,D.Klein,P.Ivanyi,G.vanSeventer,and J.Klein.1988.NucleotidesequencesofchimpanzeeMHCclass I alleles: evidence for trans-species mode of evolution. EMBO J. 7:2765-2774.
22. Miller, M. D., S. Gould-Fogerite, L. Shen, R M. Woods, S. Koenig, R J. Mannino, and N. L. Letvin. 1992. Vaccination of rhesus monkeys with synthetic peptide in a fusogenic proteoliposome elicits simian immunodeficiency virus-specific CD8+ cytotoxicT lymphocytes.J. Exp. Med. 176:1739-1744.
23. Miller, M. D., H. Yamamoto, A. L. Hughes, D. I. Watkins, and N.L.Letvin.1991.Definition of an epitope and an MHC class I molecule recognized by gag-specific CTL in SIVmac infected rhesusmonkeys. J. Immunol. 147:320-329.
24. Morrison, H. G., F. Kirchoff, and R C. Desrosiers. 1993. Evidence for cooperation of gpl20 amino acids 322 and 448 in SIVmac entry.Virology 195:167-174.
25. O'Brien, W., Y. Koyanagi, A. Namazie, J. Zhao, A. Diagne, K. Idler, J. Zack, and I. Chen. 1990. HIV-1 tropism for mononuclear phagocytes can be determined by regions ofgpl20 outside the CD4-binding domain.Nature(London) 348:69-73.
26. Otting, N., and R E. Bontrop. 1993. Characterization of the rhesus macaque(Macaca mulatta)equivalentof HLA-F.Immunogenetics 38:141-145.
27. Reimann, K. A., K. Tenner-Racz, P. Racz, D. C. Montefiori, Y. Yasutomi, W. Lin, B. J. Ransil, and N. L.Letvin. 1994. Immuno-pathogenic events in acute infection of rhesus monkeys with simian immunodeficiency virus ofmacaques. J. Virol. 68:2362-2370.
28. Saffrit, J. T., Y. Cao, C. A. Andrews, D. D. Ho, and R. A. Koup. 1993. Role ofcytotoxic T lymphocytes in acute HIV-1 infections, p.31,abstr.WS-A15-1. IXth Int. Conf. on AIDS.
29. Sanger,F., S. Nicklen, and A. R. Coulson. 1977. DNAsequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
30. Schumacher,T. N.M., M.-T. Heemels, J. J.Neefjes,W. M. Kast, C.J. M. Melief, and H. L. Ploegh. 1990.Direct binding of peptide toemptyMHC classImolecules onintact cells and in vitro. Cell 62:563.
31. Shen, L., Z. W. Chen, M. D. Miller, V. Stallard, G. P. Mazzara, D. L.Panicali,and N. L.Letvin.Induction of simian immunode-ficiency virus-specific CD8+ cytotoxic T lymphocytes following vaccination with a recombinant virus vaccine. Science 252:440-443.
32. Townsend, A. R. M., J. Rothbard, F. M. Gotch, G. Bahadur, D. Wraith, and A. J. McMichael. 1986. The epitopes of influenza nucleoprotein recognized by cytotoxic T lymphocytes can be definedwith shortsynthetic peptides. Cell44:959-968.
33. Tsubota, H., C.L. Lord, D. I. Watkins, C. Morimoto, and N. L. Letvin. 1989. A cytotoxic T lymphocyte inhibits AIDS virus replication in peripheral blood lymphocytes. J. Exp. Med. 169: 1421-1434.
33a.U.S. Department of Health and Human Services. 1985. Guide for thecareanduseoflaboratory animals, publicationno.85-23.U.S. DepartmentofHealthand HumanServices, Washington, D.C. 34. Venet, A., and B. D. Walker. 1993. Cytotoxic T-cell epitopesin
HIV/SIV infection. AIDS 7:S117-S126.
35. Walker,C. M.,D.J. Moody, D. P.Stites, andJ.A.Levy. 1986. CD8+lymphocytescancontrolHIVinfectionin vitroby suppres-singvirusreplication. Science 234:1563-1566.
36. Watkins,D.I.,Z. W.Chen,T.L.Garber,A. L.Hughes,and N.L. Letvin.1991. Segmental exchange between MHC classIgenesin higher primate: recombinationinthegorillabetweentheancestor ofahumannon-functionalgeneandanAlocus gene. Immuno-genetics 34:185-191.
37. Watkins,D.I.,M.Kannagi,M. E.Stone,andN.L.Letvin. 1988. Majorhistocompatibility complexclass Imoleculesof nonhuman primates.Eur. J.Immunol. 18:1425-1432.
38. Watkins, D. I.,J. Zemmour,and P. Parham. 1993.Non-human primateMHC class I sequences, 1992. Immunogenetics 37:317-330.
39. Yamamoto,H., M. D.Miller, H. Tsubota, D. I.Watkins, G. P. Mazzara,V.Stallard,D. L.Panicali,A.Aldovini,R.A.Young,and N. L.Letvin. 1990. Studies of cloned simian immunodeficiency
virus-specificTlymphocytes:Gag-specific cytotoxicTlymphocytes
exhibitarestrictedepitopespecificity.J.Immunol. 144:3385-3391. 40. Yamamoto, H.,M. D.Miller,D. I.Watkins, G. B.Snyder,N. E. Chase,G.P.Mazzara,V.Stallard,D.Panicali,andN. L.Itvin.
on November 9, 2019 by guest
http://jvi.asm.org/
1990. Two distinct lymphocyte populations mediate simian immu-nodeficiency virus envelope-specifictarget cell lysis. J. Immunol. 145:3740-3746.
41. Yamamoto, H., D. J. Ringler, M. D. Miller, Y. Yasutomi, T. Hasunuma, and N. L. Letvin. 1992. SIV-specific cytotoxic T lymphocytes are present in the AIDS-associated skin rash in rhesus monkeys. J. Immunol. 149:728-734.
42. Yasutomi,Y.,S. Koenig, S. S. Haun, K. Stover, R. K. Jackson, P. Conrad,A.J.Conley,E. A. Emini, T. R. Fuerst, and N. L. Letvin. 1993. Immunization with recombinant BCG-SIV elicits
SIV-spe-cific cytotoxic T lymphocytes in rhesus monkeys. J. Immunol. 150:3101-3107.
43. Yasutomi, Y., T. J. Palker, M. B. Gardner, B. F. Haynes, and N. L. Letvin. 1993. Synthetic peptide in mineral oil adjuvant elicits simian immunodeficiency virus-specific CD8+ cytotoxic T lym-phocytes in rhesus monkeys. J. Immunol. 151:5096-5105. 44. Yasutomi, Y., K A. Reimann, C. L.Lord, M. D. Miller, and N. L.
Letvin. 1993.Simian immunodeficiency virus-specific CD8+
lym-phocyte response in acutely infected rhesus monkeys. J. Virol.
67:1707-1711.