JOURNAL OF VIROLOGY, June 1994, p. 4009-4016 Vol. 68, No. 6 0022-538X/94/$04.00+0
Copyright ©D 1994,American Society for Microbiology
Rotavirus Interaction with
Isolated Membrane Vesicles
MARIE-CHRISTINE RUIZ,-.* SARA R. ALONSO-TORRE,' ANNIE CHARPILIENNE3 MONIQUE
VASSEUR,'t
FABIAN MICHELANGELI,2 JEAN COHEN,3AND FRANCISCOALVARADO1t
Centre de Recherchesur/'Endocrinologie Moleculaire et le Developpement, Centre Nationale de la Recherche Scientifique, 92190Meudon,' and Unitede Virologie et Immunologie Moleculaires, Centre de RechercheJouy, InstitutNational de la Recherche Agronomique, 78350 Jouy en Josas,3 France, and Laboratorio deFisiologia
Gastrointestinal, Instituto Venezolano de Investigaciones Cientificas, Caracas 1020A, Venezuela2
Received 16 September 1993/Accepted 25 February 1994
Togain information about the mechanism of epithelial cell infection by rotavirus, we studied the interaction of bovine rotavirus, RF strain, with isolated membrane vesicles from apical membrane of pig enterocytes. Vesicleswere charged with high (quenching) concentrations of either carboxyfluorescein or calcein, and the rateoffluorophore release (dequenching) was monitored as a function of time after mixing with purified virus particles. Purified single-shelled particles and untrypsinized double-shelled ones had no effect. Trypsinized double-shelled virionsinduced carboxyfluorescein release according to sigmoid curves whose lag period and amplitudewere afunctionof virus concentration and depended on both temperature and pH. The presence of 100mMsalts (Tris Cl,NaCi,or KCI) was required, since there was no reaction in isoosmotic salt-freesorbitol media.Othermembrane vesicle preparations such as apical membranes of piglet enterocyte and rat placenta syncytiotrophoblasts,basolateral membranes ofpigenterocytes, and theundifferentiatedplasma membrane of cultured MA104 cells all gave qualitatively similar responses. Inhibition by a specific monoclonal antibody suggests that the active species causing carboxyfluorescein release is VP5*. Ca2+ (1 mM), but not Mg2+,
inhibited the reaction. In situ solubilization of the outer capsid of trypsinized double-shelled particles changed release kinetics from sigmoidal to hyperbolic and was not inhibited by Ca2 . Our results indicate that membrane destabilization caused bytrypsinized outer capsid proteins of rotavirus leads to fluorophore release. From the data presented here, a hypothetical model of the interaction of the various states of the viral particles with the membrane lipid phase is proposed. Membrane permeabilization induced by rotavirus may be related tothemechanism of entry of the virus into the host cell.
Rotaviruses belong to theReoviridae family, RNA viruses which are the principal cause of acute gastroenteritis in children and in certain animal species (14). These nonenvel-oped viruses have a complex icosahedral structure with two protein capsids and an inner core containing a segmented double-stranded RNA genome (7).
The mechanisms of rotavirusbinding ofandpenetration into cellsare notfullyunderstood. Itisthoughtthatthe firststepin virus infection involves attachment to a plasma membrane receptor, followed by internalization (2). Both proteins ofthe outer capsid, VP7 and VP4, have been implicated in the bindingtothetargetcell(11, 27)andin theinfectiouscapacity of thevirus(21). Upon controlled digestion by trypsin, theVP4 molecule is cleaved into two fragments, VP5* and VP8* (8). This proteolytic cleavage strongly enhances rotavirus infectiv-ity in cell culture and is believed to occur inthe lumen of the intestine prior toinfection of the enterocyte (8, 32).
Attempts to identify rotavirus receptors have been made. Binding studies have revealed the presence of at least two families ofputative receptors: one sialic acid dependent, for animal rotaviruses, and the other sialic acid independent, for human rotaviruses (10, 15, 22, 31, 35, 38). Rhesus rotavirus seems to bind, via VP4, to a large sialic acid-dependent
*Corresponding author. Mailing address: IVIC-CBB, P.O. Box
21827,Caracas 1020A, Venezuela. Phone: (582)501 1396. Fax:(582) 501 1093, (582) 5727446.Electronic mail address: [email protected].
tPresentaddress:DepartementdeMicrobiologie etImmunologie,
FacultedePharmacie, Universitede Paris-XI,92296 Chatenay-Mala-bry,France.
glycoprotein of brush border membranes from suckling-mice enterocytes (3).
Twohypotheses have been advanced toexplainthe mecha-nism of rotavirus penetration into cells. The endocytosis pathway through receptor-mediated entry was firstproposed on the basis ofelectron-microscopic images of rotavirus asso-ciated withcoatedpits and vesicles(19, 25, 26). Alternatively, Suzukietal.(30)haveproposedthatrotavirusespenetrateinto cells directly through the lipid phase of plasma membranes. The biochemical studies of Kaljot et al. (13) showed that trypsinized double-capsid rotaviruses are infectious and are notsensitivetolysosomotropicagents.Rotavirusentrywasfast and concomitant witharelease of intracellularspacemarkers. Kaljot et al. proposed that trypsinized viruses induce an increase ofcell membrane permeability preceding viral
repli-cation (13). Untrypsinized double-shelled particleswould en-terthe cellvia endocytosiswithout leadingtoviral
multiplica-tion.
Morerecently,theinteraction ofpurifiedrotavirusparticles
with liposomes containing the self-quenching fluorophore 6-carboxyfluorescein (CF) has been usedas a modelto study
the mechanismsofrotavirusentryinto cells (27).
Seeking further information onthe mechanism of rotavirus infection of epithelial cells, we have studied the interaction between rotavirus and isolated membrane vesicles derived from avarietyof cells.ByusingvesiclespreloadedwithCFor
calcein, our aim was to understand the interaction between rotavirusesandmembranes. Our results indicate the existence of direct interactions between the virus particles and the membrane vesicles, supporting the hypothesis of a direct rotaviruspenetration acrossthe membranelipid phase.
4009
on November 9, 2019 by guest
http://jvi.asm.org/
(A preliminary account of this work has been presented previously [28].)
MATERMILSAND METHODS
Viruspreparations. The viruses used (bovinestrain RFand
porcinestrainOSU)weremultiplied in African green monkey
kidney cells (MA104 cells). Cells grown as monolayers in Eagle's minimum essential medium were infected either at low multiplicity of infection (0.1 PFU per cell) in the presence of
trypsin (0.44,ug/ml)or at highmultiplicity of infection (5 PFU percell) inthe absence oftrypsin but with aprotinin (1
pLg/ml).
Virus particles were extracted from infected cells with Freon 113 and purified by centrifugation on a cesium chloride gradient. Upper (double-shelled particle) and lower (single-shelled particle) bands were collected and separately
repuri-fied on acesium chloride gradient. Particle suspensions were stocked at -4°C in the cesium chloride solution and desalted before use by centrifugation through a Sephadex G-25 spin column equilibratedwith 10mM
N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid (HEPES; pH 6.9)-50 mM NaCl,
containing 2 ,uM CaCl2. Destabilization of the outer capsid
(decapsidation) wasachieved by treatment with 10 to 15 mM
ethylene glycol-bis(,3-aminoethyl ether)-N,N,N',N'-tetraacetic acid (EGTA) or by heating to 60°C for 30 s. The state of
decapsidation was checked by electrophoresis in a 25 mM Tris-192 mMglycine-0.6% agarose gel (pH 8.3) followed by
stainingwith ethidium bromide (16).
Protein andenzyme assays.Protein and virus concentrations weremeasured with the Bio-Radprotein assay kit, with serum
globulinand bovine serumalbumin, respectively, as the stan-dards. To monitor vesicle purification and yield, we deter-mined the activity of appropriate enzyme markers essentially as describedbyVasseur etal. (36) andTouzani et al. (33).
Preparationofmembranevesicles.Brush border membrane vesicles from pig jejunum or ileum were prepared by the
magnesium-EGTA precipitationmethod of Hauser et al. (12). We used both 14-day-old piglets and young pigs weighing about 100kg.The final vesiclepreparationswereresuspended in asolution of 500 mM sorbitol in a 10 mM HEPES-7 mM
n-butylaminemetal-freebuffer, adjustedtopH 7.4with maleic acid. Vesicles isolated frompig jejunumwereusedformostof the study. Basolateral membrane vesicles from pig jejunun were prepared bythe method of Touzanietal.(33). The final
preparationwasresuspended in the samebuffer used for the brushborder vesicles. Microvillous membrane vesicles from rat
placentawerepreparedby the method of Alonso de la Torre etal.
(1)
andresuspendedina20 mMTris-morpholineethane-sulfonic acid (MES) buffer (pH 7.4) containing 500 mM sorbitol. Plasma membrane vesicles from MA104 cells were
prepared bythe method of Mian(23) modified by Vasseur et al. (36). The plasma membrane-rich band at the 10 to 34%
interphaseofasucrosegradient was separated and washed by two cycles ofcentrifugation at 110,000 x g for 30 min and
resuspended in a small volume of the n-butylamine buffer
supplementedwith 300 mM sorbitol. The final plasma mem-brane fraction exhibited a Na+/K+-ATPase (EC 3.6.1.37) enrichment of15-fold compared with the crude homogenate. Allfinalvesicle preparations were stored in liquid nitrogen in
200-,ulbatchesuntilthe day of the experiment.
Vesicle loading and fluorescence measurements. Aqueous solutions of either CFor calcein (140 mM) were neutralized with Tris base (finalconcentration,400mM) and adjusted to pH7.4with20 mMHEPES. Vesicle loading wasaccomplished
by
mixing 1 volume of concentratedfluorophore solution with 1volume of vesiclesuspension.
Twocyclesoffreezinginliquidnitrogen and thawing were thenperformed by the method of Vasseur et al. (35). To eliminate extravesicular fluorophore,
the suspension waswashed twiceby centrifugation and resus-pension in a standard assaymedium (100 mM sorbitol and 200 mM Tris HCI in 20 mM HEPES-10 mM Tris [pH 7.4]). The finalpellets were assumed to contain a fully quenched, 70 mM concentration of either fluorophore.
Fluorophore-loaded membrane vesicles were kept in the dark over ice during the course ofeach experimental session. Small volumes (10 to 20
[l)
ofpreloaded vesiclesweremixed in the fluorometer cuvette with 1.6 ml of standard assay medium and preincubated for 3 min before the addition of rotavirus at the appropriate temperature (37°C unless stated otherwise) with magnetic stirring. All experiments were re-peated several times and on different days with different membrane and virus preparations. Particular conditions are specifiedin the figure legends. Fluorophore release caused byrotavirus addition was monitored as a fluorescence increase due to dilution dequenching. Dequenching was measured by using a thermostated Jobin Yvon JY3C spectrofluorimeter at excitation wavelengths of 496 nm (CF) or 490 nm(calcein) and an emission wavelength of 520 nm. The fluorometer output was connected directly to a microcomputer for sampling and data analysis.
The results are expressed as percentages of the total fluo-rescence at the end of each experiment after membrane lysis determined by the addition of Triton X-100 (final concentra-tion, 20 mM). Thepercentage of fluorophore dequenching was calculated according to the following formula: percent release
=
[(Ft
-FO)/(FT
-FO)]
X100,
whereFO
is thebackground
fluorescence, F1 is the fluorescence at time t, and FT is the total fluorescence. The dequenching rate constants are defined as thelimiting slope of the release curves during the linear phase. The lag period wasdetermined graphically as the timerequired for the establishment of a clear change of slope aftervirus addition.Antibodies and chemical. Monoclonal antibodies directed against VP4 were kindlyprovided as purifiedimmunoglobulins by M. K. Estes, Baylor College of Medicine, Houston, Tex., and H. B. Greenberg, Stanford University, Stanford, Calif. Calcein and CF were from Sigma, Saint Louis, Mo. All other reagents were of analytical grade.
RESULTS
Time course of CF release from pig brush bordermembrane vesicles. Trypsinized double-shelled particles (TDS) of both bovine RF and porcine OSU rotaviruses both induced CF release from brush border membrane vesicles of pigjejunum. Since fluorophore release was more efficient with RF rotavirus, thewhole of the present study was performed with this strain. CF release exhibited sigmoid kinetics characterized by a lag period followed by a fastincrease influorescence (Fig. 1). The increase apparently follows first-order kinetics. Neither puri-fied single-shelled particles nor untrypsinized double-shelled particles(UDS) had any effect. After a period of incubation of theuntrypsinized virus (UDS) with the vesicles, direct trypsin addition to the fluorometer cuvette resulted in a drastic shortening of the lag period and a higher rate of CF release (Fig. 1). By itself, trypsin did not produce any increase in fluorescence (data not shown). Taken together, such results suggest that thelag period corresponds to a time-dependent interaction (that includes binding) of the double-shelled par-ticles with the membranes, followed by a faster reaction(s) leading to CF release. The two processes are consecutive, and
on November 9, 2019 by guest
http://jvi.asm.org/
ROTAVIRUS-MEMBRANE INTERACTIONS 4011
60, .
TDS
-40
0) ~~~~~~~~~~~~~UDS
IL I
J'20
/
1,,; Trypsin / SS
.
_~~~~~~~~~~~~S
0 100 200
[image:3.612.335.552.72.234.2]Time (seconds) FIG. 1. Time course of CF release frompig jeju vesiclesinduced by the addition of different prepar (RF strain). TDS, UDS, or single-shelledpurified' were added tofluorophore-loaded vesicles preincu assay medium (see Materials and Methods). In I
figures,time zero represents time of virus addition. 1 was added to UDS at thearrow. The membranepro was 34.8pg/mland the virus concentrationwas 6.5
only thesecond one requirespreviousvirus tr equation 1,below).
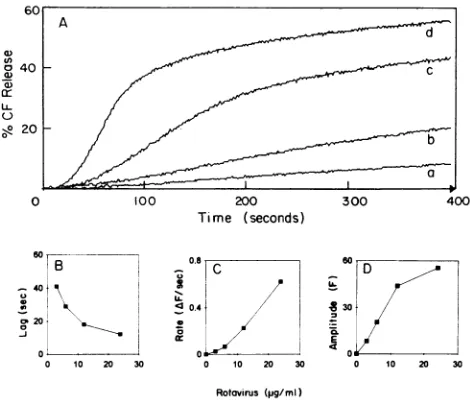
TDS-induced CF releasewas afunctionof tion (Fig. 2A). The lag duration decrease concentration increased (Fig. 2B), whereas tI constant and the amplitude of the effect boti 2C and D). At a constant virus concentrat vesicle protein concentration from 5 to 50
o 40
0)
I2
(-)
20
200
Time (seconds)
B
40>
I20 ,
0 10 20 30 0.6
0 10 20 30
[image:3.612.81.297.74.239.2]Rotavirus(pg/ml)
FIG. 2. (A) Effect ofvirus concentration on Cl
jejunum brush border vesicles at constant proti
Different volumes of TDS were added to standa containingthevesicles(15.1 pLg/mI), yielding final vi of 3 p.g/ml (a),6,ug/ml (b), 12 p.g/ml (c), and 24 ,ug forthe analysis plotsoflagversusconcentration (
concentration(C),andamplitudeversusconcentrati
fromoriginal tracesshown in panel A.
300 400
inum brush border rations of rotavirus viralparticles (SS) lbated in standard this and following [rypsin(8.8,ug/ml) iteinconcentration pLg/mlin all cases.
U-5
O 100 200 300 400
Time (seconds)
FIG. 3. Effect of monoclonal antibodies 3D8 and 2G4 on CF releaseinduced by TDS from pig jejunum brush border vesicles. Viral particleswerepreincubated overnight with antibodies (0.2pugof2G4 per ,ug of virus and0.1 ,ugof3D8perpugof virus) orassay medium at 37°C. Protein and final virus concentrations were 12.4 and 3.0 ,ug/ml, respectively.
change eitherthelag, therate, orthepercentageof CF release (results not illustrated). It may be noted here that in our ypsinization(see experimentswith membranevesicles, the overall reactionyield (percent CF release for a given virus particle concentration) virus concentra- seems to be lower than that observed by using CF-loaded ed as the virus liposomes (24).
he apparent rate Effect of monoclonal antibodies to VP4. As shown above, increased (Fig. rotavirus-induced CF release from brush border membrane ion, varying the vesicles strictly requires virus trypsinization. It is known that ),g/ml did not trypsin cleaves the outer capsid protein, VP4, into its two subunits, VP5* and VP8* (8).Thissuggeststhat oneof these protein moieties might be responsible for the CF release reaction. Toascertain the nature of the active component(s), we have used two monoclonal antibodies, 2G4 and 3D8, directed against the VP5* and VP8* moieties ofVP4, respec-tively(20, 29).These antibodies have beenobservedby immu-nofluorescence torecognize the RFstrain(5a). Preincubation of TDS with antibody 2G4 inhibited CF release by 40%, whereas3D8 had no effect (Fig. 3). These results suggest the involvement of VP5* intheCF release reaction. The possibil-ity that a 2G4-VP5* complex inhibits indirectly through an interaction with another viral protein(s) deserves further in-vestigation.
500 400 Temperature and pH effects onrotavirus-membrane inter-action. The kinetics of CF release induced by TDS changed withtemperature(Fig. 4A).Asthetemperaturedecreased,the D duration of the lag increased and the CF release rate de-creased. An Arrhenius analysis of the dependence of the
o0
apparentCF releaserate constantontemperature (inset, Fig. 4A) gave two straight lines intersecting at about 30°C. From o. here we have calculated activation energies of 4.9 and 34.90 10 20 30 kcal/mol (20.5 and 146kJ/mol),respectively, for the 41 to 310C and the 31 to24°Ctemperature ranges. The existence of such release from pig phase transition
suggests
thatmembranefluidity
plays
arole inFropig rotavirus-membrane interaction.
ein concentration. The effect of pH on CF release was studied by using calcein, ird assay medium a
pH-independent
fluorophore (Fig.
4B).
As thepH
of theirus
(d).nDatinse
incubation mediumdecreased,
thepermeabilizing
effect of B), velocity versus TDS was inhibited; the lag clearly increased, and the rate of ion(D) were taken calcein release decreased. The virus effect appears to reach aplateau in the pH 7.4to7.8 range. VOL. 68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.69.304.446.646.2]4012 RUIZ ET AL.
a)K
// / X29C
3.-3
20- 3] 32 3.337
L FI/TPK103
0 200 400
Time (seconds)
30-~20
cc
pH7 0
10
/V pH6.4
0 50 100
Time(seconds)
FIG. 4. Effect oftemperature (A) and extravesicular pH (B) on
fluorophorerelease frompig jejunumbrush border vesicles inducedby TDS. Forthetemperature experiments (A),CF-loaded vesicleswere
added to standard assay medium preequilibrated at the required
temperature, followed by virus addition 3 min later. The Arrhenius
transformation of these data is shown in the insert. For the pH experiments (B),the standardassaymediumwastakentothe desired pH with HEPES. Membrane protein concentrationswere48.8 ,ug/ml (A) and 17.0 pg/ml (B).The virus concentration was 16.2 p.g/mlfor both series.
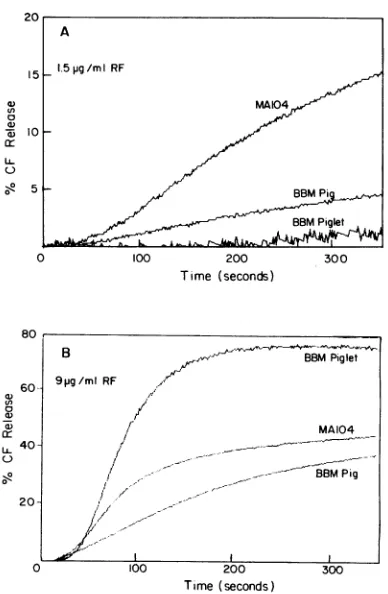
Comparisonof the rotavirus effectonmembranevesiclesof diversebiological origins.Inapreliminary attempttostudythe specificity of rotavirus-membrane interactions, we have
com-pared the effect of the TDS bovine rotavirus strain RF on
membrane vesiclepreparations isolated fromavariety of cells.
Resultsqualitatively identicaltothose inFig. 2wereobtained with all the membrane vesiclesused, which included the apical (brush border) membrane of either pigorpiglet jejunum and ileum; basolateral membranes from pig jejunum; apical
mem-branes from ratplacenta syncytiotrophoblast, and undifferen-tiated plasma membranes fromMA104 cells. The interaction oftheRFrotavirus with allthe membranes mentioned above resulted inCF release whose lag,rate, andamplitudewere a
function ofvirus concentration. However, therewere
quanti-tative differences insensitivity of the different membranes to virus concentration, as illustrated in Fig. 5. At low rotavirus
concentrations (1.5 ,ug/ml), the MA104 vesicles seemedtobe the most sensitive (Fig. 5A), whereas at sixfold-higher virus concentrations, the piglet vesicleswereclearly themost sensi-tive (Fig. 5B). This difference in sensitivity of the various membranes may be related to their respective physical and chemical characteristics, which in turn may determine
virus-membrane interactions.
15
tn
a)
cc LL
a
60-cn
0
cx
a)
-20
20-0
l00 200
Time (seconds)
300
B .r', ; ,BBMPiglet
9pg/ml RF
1'
MA104
--
B8MPigltJ
I , ._ U
loo 200
Time(seconds)
FIG. 5. Comparative effect oflow (1.5 ,ug/ml) (A) and high (9 ,ug/ml) (B)virus concentrationsonCF release in different membrane
types. Experiments were performed as described for Fig. 2 at the
following protein concentrations for each preparation: MA104 cell membranes, 8.2 ,ug/ml; pig jejunum brush border (BBM Pig), 24.1
p.g/ml; andpiglet jejunumbrush border(BBM Piglet),8.1 gg/ml. Effect ofmono-and divalent cationsonrotavirus-membrane
interaction. The CF release induced by TDS strictly requires
the presence of monovalent cations in theassay medium. At
200 mM, the chloride salts of either Na+, K+, or Tris were
about equally effective in supporting rotavirus-membrane in-teraction. Substitution of these salts by an equiosmolar
con-centration of sorbitol rendered the vesicles insensitive to the virus, indicating that a high ionic strength rather than the
presence of specific ions is the key requirement for virus-membrane interaction. The virus-membrane potential, however, doesnot seemtoplay anyrole, because theresponsewasnot modified whena60-mVinside-positive potentialwasimposed
by applying an appropriate K+ gradient in the presence of valinomycin (resultsnotillustrated).
We next studied the effect of divalent cations on the
rotavirus-membrane interaction. When addedto a nominally
calcium-free assaymedium, 1 mM Ca2+ completely inhibited CF release(Fig. 6),whereasupto 10 mMMg2+hadnoeffect (results not illustrated). Addition of 1 mM EGTA to the medium containing 1 mM Ca2+ (which can be calculated to
cause a free Ca2+ concentration drop to 5 ,uM [9]) fully reversedthe inhibition (Fig. 6). It is noteworthy that the CF releasecurvesunder these last conditions exhibitedlonger lag periodsthan didthose for the controls without calcium. These resultssuggest that calcium inhibits virus-membrane interac-tion by either acting onthe membrane vesicle or formingan
inactiveCa2+-viruscomplex. However,seethenextsection for evidenceofalack ofdirect effects ofCa2+ onthemembrane
A
1.5pg/mI RF
MA104
t~~ _ ~
~~~
BBMPiglet . AA~~
~
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.333.527.68.365.2] [image:4.612.70.269.71.385.2]ROTAVIRUS-MEMBRANE INTERACTIONS 4013
0~~~~C2
0
0Co2
/20 _
EGTA
ImMCo2+ I
100 200 300 400
[image:5.612.82.297.77.240.2]Time (seconds)
FIG. 6. Effects of 1 mMCa2+on CF releaseinduced by TDS from pig jejunum brush border vesicles and reversal ofCa2+ inhibition by chelation with 1mMEGTA.Vesicleswerepreincubated for 3 min in the presence(linelmMCa2+)orabsence(line0Ca2+)of 1 mMCa2+ beforetheadditionof TDS.Proteinconcentrations were 34.8 ,ug/ml, andthe virus concentration was 6.5 pg/ml.
itself. The chelating agent EGTA reverses the inhibition by removingcalcium and regenerating the interactiveforms of the virus. It seems worth mentioning here that, at the Ca2+ and EGTA concentrations used in these experiments, TDS re-mained in adouble-capsid conformation, as checked by elec-trophoresis, in contrast tothe situation described in the next section(see Materials and Methods).
Effectofouter capsid proteins released byinsitu rotavirus decapsidation. To study the role of the outer capsid proteins onvirus-membraneinteraction,wehave treatedpurified TDS either with 15 mM EGTA or by heating to 60°C for 30 s immediately prior to mixing with the vesicles. Both of these treatments areknowntodestabilize the external capsid, lead-ing to the release of the outer capsid proteins, VP7 and VP4
(or VP7, VP5* and VP8*, if TDS are used), while the single-shelled virions remain intact (6). We verified by electro-phoresis (results not illustrated) that, after application of either of the above treatments, solubilization of the outer capsid (abbreviated "decapsidation") indeedwas complete in ourexperiments.
Whenthedecapsidated TDS suspensionwasmixed, without further treatment, with the CF-loadedvesicles,bothqualitative
and quantitative changes in the virus-membrane interaction were observed (Fig. 7): (i) the shape of the curves changed from sigmoidal tohyperbolic, the lagperiod practically disap-pearing; (ii) therewas anabrupt increase in thenet rateof CF release; and,finally,(iii) the reaction became totally insensitive toCa2+concentrationsashighas10mM.This result indicates thattheCa2+inhibition observed above with intact TDS is due not to aninteraction with the membrane vesiclebut,rather,to theformation ofaninactive
Ca2+-TDS
complex.Furthermore, although, the rate of CF release was still related to the total virus concentration, the lag period was absent even at the lowest virus concentrations used (Fig. 8).
When theapparentCF release rate constants wereplotted as afunction of the total virusconcentration, both the native and the in situ-decapsidated TDS exhibited sigmoid kinetics, but the maximal velocity evinced by the decapsidated virus was about 1 orderofmagnitude higher (Fig. 2Cand8[insert]).
Since single-shelled particles are inactive (Fig. 1), the dra-matic effects evidenced with the in situ-decapsidated TDS
0(4 40
Iogdv
cr
LL 0
20
aTDS-EGTA
bTDS-EGTA+IOmMCa2O
cTDS-HEAT
10 d TDS
.I II
0 100 200 300 400
[image:5.612.336.556.78.240.2]Time(seconds)
FIG. 7. Effect of the outer capsid proteins on CF release frompig jejunum brush border vesicles. Purified TDS were decapsidatedby pretreatmentwith EGTA(14mM)orheating (60°Cfor 30s). These (TDS-EGTAorTDS-HEAT)orthe untreatedsuspension(TDS)were added to startthe reaction. Thefinalconcentration ofEGTAinthe cuvette was0.1 mM.For curve btheassaymediumcontained 10 mM Ca2+.The vesicleproteinconcentration was36.6p.g/ml,andthevirus proteinconcentrationwas8.1,ug/ml.
rotavirus must be attributed to a direct action of the outer capsid proteins when dissociated from the virion. We have proved that this is the case because, after decapsidation by EGTA, the high-speed supernatantcontaining onlysolubilized outercapsid proteins, without single-shelled particles (which
00,
80
a)
(I)
o 60
a)o
a) 0
l40
20
0 200
Time (seconds)
FIG. 8. Effect ofoutercapsidproteinconcentrationonCFrelease frompig jejunumbrush border vesicles. Different volumes of suspen-sion of decapsidated TDS (pretreatedwith EGTA) were added to standard assay medium containing the vesicles so as to give the followingvirus concentrations: 3.5 ,ug/ml(a),5.2,ug/ml(b), 8.1,ug/ml (c),and 16.2p.g/ml(d).The membraneproteinconcentrationwas36.6 ,ug/mlfor the series.Experimentswereperformed asdetailed inthe legend to Fig. 2. Data used for the analysis plot ofvelocityversus concentration(inset)weretaken from theoriginaltracesshown inthe figure.
VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.337.550.415.634.2]40 TDS-EGTA
UDS-EGTA
20 cr LI.
TRYPSIN
0 100 200
Time (seconds)
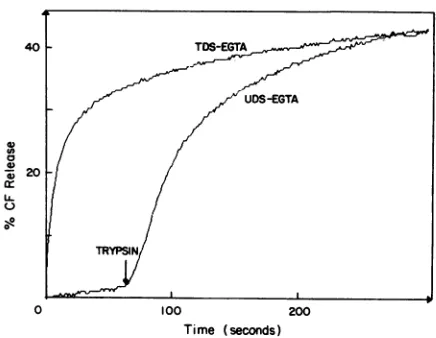
FIG. 9. Timecourse of CF release from pig jejunum brush border
vesicles induced by the addition of different decapsidated rotavirus suspensions containingoutercapsidproteins. Decapsidated TDS (line TDS-EGTA) orUDS (line UDS-EGTA) wasadded at time zero to fluorophore-loaded vesicle membrane suspensions preincubated in standardassaymedium.Trypsin(8.8,ug/ml)wasaddedtoUDS-EGTA during thecourseof the experiment. The membrane protein
concen-trationwas49.3,ug/ml, and the virus concentrationwas6.5 pLg/ml.
went to the pellet, as checked by electrophoresis), produced
the full effect with the same kinetics as that of the
decapsi-dated-particle suspension. On the other hand, the pellet (sin-gle-shelled particles) did not produce any CF release (results
notshown).
It isnoteworthy that suspensions ofuntrypsinized, decapsi-dated rotavirus did not produce any CF release but that addition oftrypsin directly tothe fluorometer cuvetteinduced
afully dequenching effect (Fig. 9). However, the effect inthis case wassomewhatslower,whichmayinprinciplebeexplained in terms ofthe timerequired for the trypsin activity products (VP5*, VP8*) to reach a critical concentration under these conditions.
DISCUSSION
In this paper, we present evidence for an interaction of
rotavirus particles with isolated plasma membrane vesicles, which was monitored in terms of the release of vesicle-entrapped fluorophores.Our results confirmand extendthose ofNandiet al. (24), who used CF-loaded liposomes as their
experimental model. At present, we cannot say whether the
mechanismofCFrelease measuredinthese experiments isat least inpartthesame asthat associated with virus
internaliza-tion undernatural conditions. Nevertheless, thispossibility is suggested bytheexistence of certainkey similarities between thetwo processes: (i)one orbothof the twoprotein
constit-uentsof theexternal capsidof therotaviruses, VP4andVP7,
areimplicated, and single-shelled particles havenoeffect; (ii)
virus trypsinization isanabsolute requirement for CF release
andfacilitates infectious-virus entryinto the cell; and (iii) CF release is inhibited by the neutralizing monoclonal antibody 2G4, directed againstVP5*. This point is strengthenedby the lackof inhibitionby antibody3D8(nonneutralizing), which is directed against VP8 (5).
Theoptimal pH forpermeabilization might be also related to the pathway of virus entry into the cell. The fact that rotaviruspermeabilizes membrane vesicles atslightly alkaline pH values (pH 7.4 to 7.8) suggests that penetration occurs
directly through the plasma membrane rather than by endocy-tosis. If the virion entered the cytosol via the lysosomal membrane, the optimal permeabilization should be expected to occur atacidic pH values, as described for several enveloped andnonenveloped viruses (4, 18). This interpretation agrees with that of Kaljot et al. (13), who have proposed a direct rotavirus penetration through the plasma membrane of the MA104 cell.
The interaction between enveloped virions and biological membranes is believed to involve fusion between the lipid phases of both membranes (18). Such fusion, however, cannot take place with rotaviruses, which lack a lipid membrane. Rather, rotavirus-membrane interactions may be expected to involve only the readily exposed proteins of the outer capsid and either the membrane lipids, membraneproteins, orboth. As weshall see, our results can be best interpreted in terms of directviral protein-membrane lipidinteractions. Such interac-tions have been shown to occur between rotaviruses and membrane glycosphingolipids (37) and between rotaviruses and liposomes (24). The absence of protein in the liposome preparations used by these workers excludes the possibility that a membraneprotein receptorwasrequired for release in theirexperiments. Furthermore, CF release was obtained with all the biological membranes studied by us. However, the sensitivities of the different membranes were not the same, with the liposomes (24) being the most sensitive. This would suggest that no specific membrane protein(s) interacts with outer capsid proteins; rather, the chemical composition and physical characteristics of the membrane (e.g., lipid composi-tion, presence or absence of membraneproteins) may deter-mine the kinetics of virus-membrane interactions. However, data presented here do not exclude the participation of membraneproteins in the release process. At the same time, it may bepossible that all of these membranes possess copies of certainwell-conserved proteins involved in virusbinding.
The rate of dye release from membrane vesicles is strongly affectedby the phase transition temperature. The fact that this rate is lower and more temperature sensitive below the gel-liquid-crystalline phase transition may indicate that a fluid membranefacilitates fluorophore release. The large activation energies obtained below the phase transition temperature may reflect the existence of a large-scale membrane lipid reorgani-zation during the release step (34). Again, such results are essentially identicaltothose obtained by using liposomes(24), furthersuggesting that fluorophorereleasedependson adirect interaction of the rotavirusproteins with the membranelipids. Protein-protein interactions of the ligand-receptor type have indeed been found in both MA104 cells and murine entero-cytes (3, 15, 38). However, the presence of this type of interaction cannot be concluded from our results. This suggests that rotaviruses are capable of bypassing the membrane pro-tein receptor tointeract directlywith membrane lipids in the different typesofmembranes assayed.
Strong evidence thatspecificreceptorscanbebypassed has been presented recently by Mendez et al. (22). They have shown the existenceofat least twositesontheVP4protein of animal rotaviruses: a sialic acid-binding site in the VP8* moiety, and a sialic acid-independent site which (they also suggest among other possibilities) might involve the fusion peptide withinVP5.Thefirstbinding reaction isnotnecessary forinfection,asdemonstrated by the isolation by these workers of sialicacid-independent mutantsinwhich the interaction of VP8* with the membrane receptor can in fact be bypassed. Our resultsarein accordance with thisinterpretation. Thefact that mutations in escape mutants to 2G4 aremappedatamino acid 393 of VP5* from the simian rotavirus RRV (20, 29)
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.60.278.73.242.2]ROTAVIRUS-MEMBRANE INTERACTIONS 4015 suggests that the active site for permeabilization of the
mem-brane vesicles is located close to or inside the putative fuso-genic region,which is homologousto that present in Semliki Forestvirus (17).
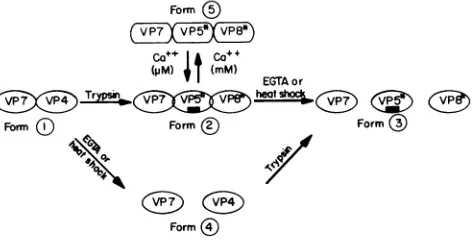
In view of the above considerations, we propose therefore that ourresultscanbe interpreted intermsof two consecutive bindingsteps,leadingtothefinal, irreversiblepermeabilization step:
k,
k,k3
TDS +M =TDS-M =TDS-M* - permeabilization (1)
where M is the membrane, TDS-M is the double-capsid rotavirus-membrane complex, and TDS-M* is an activated formofthis complex which immediatelyprecedesfluorophore release.
Wedefinethefirstbinding reactioninequation 1 as aglobal one that may or may not include an initial virus binding to specific protein receptors; it can use both TDS and UDS as substrates, asindicatedbythe results in Fig. 1.This would be therate-limiting stepforthe overall reaction and accounts for thelag phase. Incontrast,thesecondbinding reaction implies the existence of a second binding site with low affinity for untrypsinized VP4 (and/or VP7). Since this step strictly re-quires rotavirus trypsinization, it can be suggested that it dependsontheuncovering of the fusogenicsequenceinVP5*, which would be cryptic in VP4. This proposal is fully in agreementwith the observation that cleavage ofVP4 enhances virus penetration but notbinding tothe cell (11).
Theinhibition byCa2>,but notby
Mg2+,
of the CF release inducedby TDS canbepostulatedto involve formation ofan inactiveTDS-Ca2+
complex according to a freely reversible equilibrium that can be displaced toward dissociation by EGTA (Fig. 6). The reason why Ca2+ concentrations in the millimolar range maintain the outer capsid in an inactive conformation remainstobeestablished. Itisknownthat, with intact cells, the presence of extracellular Ca2+ does not prevent rotavirus infection, an apparent contradiction that should beexplained. One possible explanation for this anomaly is that, because the TDS-Ca2+ complex involves a freely reversible equilibrium, there will always be some free TDS available for infection to occur.Conclusions. Our results indicate that there are rotavirus-membrane interactions which involve several distinct virion conformations that are schematized in Fig. 10. We will focus attentionontheoutercapsid proteinsVP7andVP4.Theyare depicted simply as"domains" (ovals and rectangles), without any inference as to either stoichiometry or molecular size. Trypsinization of VP4 leads to the appearance of two new domains (VP5* and VP8*). The key trypsinization reaction makes available for interaction with the membrane the fuso-genic region (black rectangle) whichwascrypticin VP4. In the present model we make no inference about the role of each individual capsid protein in the interaction of viral particles with the membrane lipid phase.Thefusogenic region has been depicted as being localized on VP5 on the basis of previous sequenceanalysis (20). However,apeptideofsuch kind could either be present in another protein or result from domains present inseveral proteins.
Form 1 isthe UDS,which is inactive in membranelysisbut does bindtothemembrane,sincepreboundUDS respondsto in situ addition of trypsin by CF release at full rate, with a greatlyshortenedlag period (Fig. 1). Form2 isthe TDS. This is the form that leads to CF releaseaftertheprevioustwo-step consecutivebindingdescribed in equation 1. Form3
schema-Form )
CO++l Ca+
M)t(MM)
EGTAor(3iXG3) Try(X~j~psin< heat shoc C (3 3
Form (C Form Form®
Form
FIG. 10. Schematicrepresentationof thepossible rotavirus forms evinced by our results. Unidirectional arrows represent irreversible reactions. Thefusogenicpeptidesequenceisrepresentedbythe black rectangle.Furtherexplanationsaregiven in thetext.
tizes thedecapsidatedform2. It appearstoleadrapidlytoCF release, because this form can bypass the rate-limiting first
binding reaction in equation 1 and contains the fusogenic
region fully exposed.Form 4isanuntrypsinized, decapsidated virion that is inactivein termsoflysis but would become active as soon astrypsin is addedtoittogiveform3. Finally,form5 represents the TDS existing in the presence ofhigh (1 mM) Ca21- concentrations. The resulting TDS-Ca2+ complex is
representedby squaressignifying its inabilityto catalyze per-meabilization of the membrane. Thefusogenic regionis
prob-ablyhidden, which wouldexplain the absence ofactivity.The reaction, however,can be reversedbyaddition oflow (nonde-capsidating)EGTAconcentrations, capableofbringing [Ca2+]
back to the micromolar range. This form appears to be nonbinding as judged from the lag period observed upon reversalbyEGTA(Fig. 6). Alternatively,thelagmay represent the time requiredfor returningto the active TDS conforma-tion (form 2).
ACKNOWLEDGMENTS
This workwassupportedinpartbygrant489NS1 from the Institut National de la Santeetde la RechercheMedicale (INSERM), France, andgrantsS1-1832 and MPS-RP-IV-140031 fromCONICIT, Venezu-ela. M.-C.R.wasonleaveof absence from IVIC andwassupported by fellowships from the Fondation pour la Recherche Medicale, Paris, France; CONICIT, Venezuela;andthe FrenchEmbassy, Venezuela. S.R.A.-T.was afellow of theMES-MRT,Spain.
We thankDaniele Gerbaud for excellent technical assistance and Duwhya Otero for thedrawings.
REFERENCES
1. Alonso de laTorre, S. R.,M. A.Serrano,F.Alvarado,andJ.M. Medina. 1991. Carrier-mediated L-lactatetransportin brush bor-der membrane vesicles from rat placenta during late gestation. Biochem. J. 278:535-541.
2. Bass, D. M., and H. B. Greenberg. 1992. Strategies for the identification of icosahedral virusreceptors.J.Clin.Invest.89:3-9. 3. Bass,D.M.,E. R.Mackow,and H. B.Greenberg. 1991. Identifi-cation andpartial characterization ofarhesus rotavirus binding glycoproteinon murineenterocytes.Virology 183:602-610. 4. Blumenthal, R., P. Seth,M.C.Willingham,and I. Pastan. 1985.
pH-dependent lysis of liposomes by adenovirus. Biochemistry 25:2231-2237.
5. Burns,J. W.,H. B.Greenberg,R. D.Shaw,and M. K. Estes.1988. Functionalandtopographical analysesofepitopesonthe hemag-glutinin (VP4) ofthe simian rotavirus SAl1. J. Virol. 62:2164-2172.
5a.Cohen, J.Unpublished results. VOL. 68? 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.331.567.77.197.2]6. Cohen, J., J. Laporte, A. Charpilienne, and R. Scherrer. 1979. Activation of rotavirus RNA polymerase by calcium chelation. Arch.Virol. 60:177-186.
7. Estes, M. K., and J. Cohen. 1989. Rotavirus gene structure and function. Microbiol. Rev. 53:410-499.
8. Estes,M.K.,D. Y.Graham, and B. B. Mason. 1981. Proteolytic enhancement of rotavirus infectivity: molecular mechanisms. J. Virol. 39:879-888.
9. Fabiato, A., and F. Fabiato. 1988. Computer programs for calcu-lating total from specified free or free from specified total ionic concentrations in aqueous solutions containing multiple metals and ligands. Methods Enzymol. 157:378-417.
10. Fukudome, K., 0. Yoshie, and T. Konno. 1989. Comparison of human, simian, and bovine rotaviruses for requirement of sialic acid in hemagglutination and cell adsorption. Virology 172:196-205.
11. Fukuhara, N., 0. Yoshie, S. Kitaoka, andT.Konno. 1988.Role of VP3 in human rotavirus internalization after target cell attach-mentvia VP7. J. Virol. 62:2209-2218.
12. Hauser, H.,K.Howell,R. M.C. Dawson,and D. E.Bowyer. 1980. Rabbit small intestinal brush border membrane preparation and lipidcomposition. Biochim.Biophys. Acta 602:567-577. 13. Kaljot,K.T.,R. D.Shaw,D. H.Rubin, and H. B.Greenberg.1988.
Infectious rotavirus enters cells by direct cell membrane penetra-tion, notby endocytosis. J. Virol. 62:1136-1144.
14. Kapikian, A. Z., and R. M. Chanock. 1990. Rotaviruses, p. 1353-1404. In B. N. Fields and D. M.Knipe (ed.), Virology. Raven Press, New York.
15. Keljo,D.J.,andA. K. Smith. 1988.Characterization of binding of simian rotavirus SAl1 to cultured epithelial cells. J. Pediatr. Gastroenterol. Nutr. 7:249-256.
16. Labbe, M.,A.Charpilienne, S.E.Crawford,M. K.Estes, andJ. Cohen. 1991. Expression of rotavirus VP2 produces empty core-likeparticles. J. Virol. 65:2946-2952.
17. Levy-Mintz, P.,and M.Kielian.1991.Mutagenesisof theputative fusion domain of the Semliki Forest virusspike protein. J. Virol. 65:4292-4300.
18. Loyter, A., V. Citovsky, and R. Blumenthal. 1988. The use of fluorescence dequenching measurements to follow viral mem-branefusion events. Methods Biochem. Anal. 33:129-164. 19. Ludert, J. E.,F.Michelangeli,F.Gil,F.Liprandi, and J. Esparza.
1987. Penetration and uncoating of rotaviruses in cultured cells. Intervirology 27:95-101.
20. Mackow,E.R., R. D.Shaw,S. M.Matsui,P. T.Vo,M.-N.Dang, and H. B. Greenberg. 1988. The rhesus rotavirus gene encoding protein VP3: location of amino acids involved in homologous and heterologous rotavirus neutralization and identification of a puta-tivefusionregion.Proc. Natl. Acad.Sci. USA 85:645-649. 21. Matsui, S. M.,P. A.Offit,P. T.Vo,E. R.Mackow,D. A.Benfield,
R. D. Shaw, L. Padilla-Noriega, and H. B. Greenberg. 1989. Passiveprotection against rotavirus-induced diarrhea by monoclo-nal antibodies to the heterotypic neutralization domain of VP7
andthe VP8 fragment of VP4. J. Clin. Microbiol. 27:780-782. 22. Mendez, E., C. F. Barias, and S. Lopez. 1993. Binding to sialic
acids isnot anessential step forthe entry of animal rotaviruses to epithelial cells in culture. J. Virol. 67:5253-5259.
23. Mian,N. 1986. Analysis ofcell-growth-phase-related variations in hyaluronate synthase activity of isolated plasma-membrane frac-tionsof cultured human skin fibroblasts. Biochem. J. 237:333-342. 24. Nandi, P., A. Charpilienne, and J. Cohen. 1992. Interaction of
rotavirusparticles with liposomes. J. Virol. 66:3363-3367. 25. Petrie,B. L., D. Y.Graham, and M. K. Estes. 1981.Identification
ofrotavirus particles types. Intervirology 16:20-28.
26. Quan, C. M., and F. W.Doane. 1983. Ultrastructuralevidence for the cellular uptake of rotavirus by endocytosis. Intervirology 20:223-231.
27. Ruggeri, F. M., and H. B. Greenberg. 1991. Antibodies to the trypsin cleavage peptide VP8* neutralize rotavirus by inhibiting binding of virions to target cells in culture. J. Virol. 65:2211-2219. 28. Ruiz, M. C.,S. R.Alonso-Torre, A. Charpilienne,F.Michelangeli, M.Vasseur, F. Alvarado, and J. Cohen. 1992. Rotavirus interac-tions with isolated cell membrane vesicles. Abstr. Fourth Int. Symp. Double-StrandedRNAViruses.
29. Shaw, R. D., P. T. Vo, P. A. Offit, B. S. Coulson, and H. B. Greenberg. 1986. Antigenic mapping of the surface proteins of rhesus rotavirus. Virology 155:434-451.
30. Suzuki, H., S. Kitaoka,T. Konno,T.Sato, and N. Ishida. 1985. Twomodes of human rotavirus entry into MA 104 cells. Arch. Virol. 85:25-34.
31. Svensson, L. 1992. Group C rotavirus requires sialic acid for erythrocyte and cell receptorbinding.J.Virol. 66:5582-5585. 32. Theil, K.W.,E. H. Bohl, R. F. Cross, E. M. Kohler,andA. G.
Agnes.1978. Pathogenesisof porcine rotaviral infection in exper-imentally inoculated gnotobiotic pigs. Am. J. Vet. Res. 39:213-220.
33. Touzani, K.,M.Vasseur, and F.Alvarado.Chlorideuptakeacross isolated intestinal guinea pig basolateral membrane vesicles: evi-dence for a chloride uniport and a Cl-/Cl- antiport. Am. J. Physiol., in press.
34. Tsao,Y.S.,and L.Huang.1986.Sendai virusinduced leakage of liposomescontaininggangliosides. Biochemistry 25:3971-3976. 35. Vasseur, M., M. Cauzac, R. Frangne, and F. Alvarado. 1992.
Trans-potassium effects on the chloride/proton symporter activity ofguine-pigilealbrush-border membrane vesicles. Biochim. Bio-phys. Acta 1107:150-158.
36. Vasseur, M., M. Cauzac, I. Garcia, and F. Alvarado. 1992. Chloride transport in control and cystic fibrosis human skin fibroblast membrane vesicles. Biochim.Biophys. Acta 1139:41-48. 37. Willoughby, R. E., R. H. Yolken, and R. L. Schnaar. 1990. Rotaviruses specifically bind to the neutral glycosphingolipid. J. Virol.64:4830-4835.
38. Yolken, R. H., R. Willoughby, S.-B. Wee, R. Miskuff, and S. Vonderfecht. 1987.Sialic acid glycoproteins inhibit in vitro and in vivoreplication of rotaviruses. J. Clin. Invest. 79:148-154.