0022-538X/94/$04.00+0
Copyright X 1994, American Society forMicrobiology
Platelet-Activating
Factor:
a
Candidate Human
Immunodeficiency
Virus
Type
1-Induced Neurotoxin
HARRIS A. GELBARD,l.2* HANS S. L. M. NOTTET,3SUSANSWINDELLS,4 MARTI
JETT,5
KIRK A. DZENKO,1 PETERGENIS,3 ROBERT WHITE,1 LEO WANG,1 YUN-BEOMCHOI,6 DONGXIAN ZHANG,6STUART A. LIPTON,6WALLACE W.TOURTELLOTTE,7
LEON G. EPSTEIN,1'2'8 AND HOWARD E.GENDELMAN3'4'9
ChildNeurology Division, Department of
Neurology,'
and Departments ofPediatrics,2 and Microbiology and Immunology,8 UniversityofRochesterMedicalCenter, Rochester,New York14642; Laboratory ofViralPathogenesis,Departmentsof Medicine4 and PathologyandMicrobiology3 andEppleyInstituteforResearch in Cancer and Allied
Diseases,9
UniversityofNebraska MedicalCenter, Omaha, Nebraska 68198;DivisionofMolecularPathology, Walter Reed Army InstituteforResearch, Washington, D.C.s;Departmentsof Neurology, Children's BethIsrael, BrighamandWomen's, and Massachusetts GeneralHospitals, Harvard MedicalSchool, Boston, Massachusetts021156; and NationalNeurological Research Specimen
Bank,
Wadsworth and BrentwoodDivisions, VA MedicalCenter,LosAngeles, Califomia 900737 Received 8February1994/Accepted 12April 1994
Thepathogenesis of centralnervoussystem diseaseduringhumanimmunodeficiencyvirus type1 (HIV-1) infection revolves aroundproductiveviralinfection ofbrain macrophages and microglia. Neuronallosses in thecortexandsubcortical gray matteraccompanymacrophageinfection. Thequestionof how viral infection of brainmacrophages
ultimately
leadstocentral nervous system (CNS) pathologyremains unanswered. Our previousworkdemonstratedhigh-level production oftumornecrosis factoralpha, interleukin 1 ,arachidonic acid metabolites, and platelet-activating factor (PAF) from HIV-infected monocytes and astroglia (H. E. Gendelman,P. Genis,M.Jett,and H. S.L. M.Nottet,in E.Major, ed., Technical Advances in AIDS Research in the Nervous System, in press; P. Genis, M. Jett, E. W. Bernton, H.A. Gelbard, K. Dzenko, R. Keane, L. Resnick, D. J.Volsky,L.G. Epstein, and H. E.Gendelman, J. Exp. Med. 176:1703-1718, 1992).Thesefactors, together, were neurotoxic. The relative role(s) of eachof these candidate neurotoxins in HIV-1-related CNS dysfunction was not unraveled by these initial experiments. We now report that PAF is produced during HIV-1-infected monocyte-astroglia interactions. PAFwas detected at high levels in CSF of HIV-1-infected patients withimmunosuppression and signs of CNS dysfunction.Thebiologic significanceof the results for neurological disease wasdetermined by addition of PAF to cultures ofprimary human fetal cortical or rat postnatalretinalganglion neurons. Here, PAFatconcentrations of -300pg/ml producedneuronal death. The N-methyl-D-aspartate receptor antagonist MK-801 or memantine partially blocked theneurotoxic effects of PAF.Theidentification of PAFas anHIV-1-induced neurotoxin providesnewinsights into how HIV-1causes neurological impairmentand howit mayultimately be ameliorated.Human immunodeficiency virus type 1(HIV-1) infection of the central nervous system (CNS) results in cognitive and motor abnormalities in a majority of infected individuals. HIV-1-associated dementia or AIDS dementia complex (ADC), adevastating complication of direct viral infection of the brain, arguably represents one of the most severe and significant clinical manifestations of HIV infection (6, 13, 16, 26,43). The pathological hallmarks coincident with cognitive and motor dysfunctions include HIV-1 infection of brain macrophages, microglia, and multinucleated giant cells; astro-cyte proliferation; and neuronal loss in discrete areas of the retina,neocortex,and subcortical brain (14, 18, 29, 40, 49, 50, 53, 59,60).Interestingly, this disease complex is most prevalent invirus-infected children (6).
The molecular mechanisms governing neurologic dysfunc-tionduringHIV-1infection of the brain remain an enigma. No laboratory has convincingly demonstrated HIV-1 mRNA or p24 antigens in neurons or oligodendrocytes within virus-infected brain tissue. How does HIV-1 cause neuronal loss
*Corresponding author. Mailing address: Division of Pediatric Neurology, Box 631, University of Rochester Medical Center, 601
Elmwood Ave., Rochester, NY 14642. Phone: (716) 275-4784. Fax: (716)275-3683.
when neurons are not infected and perhaps only a small number ofmacrophages support productive viral replication (13, 17,40)? Indeed, secretory productsfrom HIV-1-infected macrophages are the likely source of neurotoxic activities. Neuronal deathcanoccur soonafter exposuretoculturefluids from HIV-1-infected monocytes. Monocyte production of neu-rotoxic factors appears to require HIV-1 infection (21, 47), monocyte activation, and astroglia and/or neuronal monocyte interactions(2,12, 17, 20,52).HIV-1 gene productsalso affect neuronalviability. Picomolar amountsofHIV-1 gpl20added tomixedmicroglial-neuronal cells in thepresence of sublethal concentrations ofglutamate are toxic to rat retinal ganglion cells (10, 11,33-35). Thistoxicity is associated with increased neuronal
Ca2'
and is reversed by calcium channel or N-methyl-D-aspartate (NMDA) antagonists (33-35).Ourrecent workhas demonstrated that coculture of HIV-1-infected monocytes and human astrocytoma cell lines in-duces cytokines, arachidonic acid metabolites, and platelet-activating factor (PAF) from virus-infected monocytes (20). Astroglia cells serve to activate macrophages for their produc-tion of these neurotoxic factors (17). Our initial studies, however, didnotprovide evidence that anyorall of the factors areneurotoxinsordetermine whetherthey playabiologically important role in neurological disease. The purpose of the 4628
on November 9, 2019 by guest
http://jvi.asm.org/
HIV-MEDIATED NEUROTOXICITY 4629
present study was to identify PAF as a candidate HIV-1
neurotoxin and to provide a conceptual framework for how
"putative"HIV-1 neurotoxinsare identified.PAFis produced
within cocultures of HIV-1-infected monocytes and astroglia and ispresentinthe cerebrospinal fluid (CSF)of HIV-infected patients during neurologic dysfunction. PAFaddedtoprimary human orrat neurons at levelsapproximating those found in the CSF of HIV-1-infected patientsproduces dose-dependent neurotoxicity. NMDA receptor uncompetitive antagonists MK-801 and memantine (4, 22, 35, 39) block PAF-mediated neurotoxicity. These findings, taken together, provide insights into the pathobiology of ADC and newideas for therapeutic
interventions for neurological impairments associated with HIV-1 infection.
MATERIALS ANDMETHODS
HIV-1 infection ofmonocytes. Monocytes were recovered
fromperipheral blood mononuclear cells ofHIV-and hepatitis B-seronegative donors after leukapheresis and purified by
countercurrent centrifugal elutriation. Cell suspensions were
>98% monocytes by criteria of cell morphology on
Wright-stainedcytosmears,by granular peroxidase,andby nonspecific
esterase (15, 19). Cellswere cultured asadherentmonolayers
(106/ml in 24-mm-diameterplastic culture wells) in Dulbecco modified Eagle medium (Sigma, St. Louis, Mo.) with
recom-binant humanmacrophage colony-stimulatory factor (Genetics Institute, Cambridge, Mass.). The human brain astroglia
tu-mor-derived cell line U251 MG, a generous gift from D.
Bigner,wasutilized in cocultivationassayswith HIV-1-infected
monocytes as described previously (20). These cells were
grown as adherent monolayers in Dulbecco modified Eagle medium (Sigma)-10% heat-inactivated fetal calf serum
(Sig-ma)-50 ,ugofgentamicin perml.
Monocytes were exposed to HIV-1ADA (accession number M60472) (42) at a multiplicity ofinfection of 0.01 infectious
virions pertarget cell (19, 57). The viral inoculawere free of
mycoplasma contamination (Mycoplasma Detection Kit III;
Geneprobe, San Diego, Calif.). Human macrophage colony-stimulating factor-treated monocyteswere cultured as
adher-entmonolayers 7 to 10 days prior to use asviral targetcells. Under these conditions, 10 to 20% of the monocytes are
productivelyinfected 7days after HIV-1 inoculation (27).All cultures were refed with fresh medium every 2 to 3 days. Reversetranscriptase activitywasdetermined in culture fluids
added to a reaction mixture of Nonidet P-40 (Sigma), poly(rA)-oligo(dT) (Pharmacia, Piscataway, N.J.), dithiothrei-tol(Pharmacia), MgCl2,and [oa-32P]dTTP(400 Ci/mmol; Am-ersham Corp., Arlington Heights, Ill.) for 24 h at 37°C. The mixture wasapplied to chromatographypaper, airdried, and washed five times in 0.3 M NaCl-0.03 M sodium citrate (pH 7.4) and twice in 95% ethanol. The paperwasdried and cut,
and the radioactivity wascountedby liquid scintillation
spec-troscopy (27). Five to 7 days after infection and during the peak ofreverse transcriptase activity (107 cpm/ml)in
HIV-1-infected monocytes, equal numbers of astroglial cells, U251 MG, were added and cell lysates were recovered for PAF
determinations.
PAFassay.Cultured cellsorCSFsampleswerefrozenindry ice-ethanol mixtures. CSF samples were frozen immediately after procurement frompatients. Cellsampleswere extracted
within 1 week offreezingtopreservetheintegrityof thelipids, whichare generallyunstableinaqueoussolutions.The
extrac-tionprocedurewasperformedwithaC18disposable cartridge (Waters, Milford, Mass.) as previously described (18). PAF
was eluted as a separate fraction, with approximately 70%
TABLE 1. PAFlevels in CSF of HIV-1-infectedpatients with or
without neurological impairment
Patient ADC No. of CD4+ Mean PAF concn no. Age stage' T cells (pg/ml)± SEM
1 7mo 1 480 22
2 10 yr lb 182 32
3 7yr ob 20 61
4 47yr 1 530 124 17
S Syr 3b 256 152 6
6 61 yr 0 355 157 13
7 37yr 0 116 192 27
8 35yr 0 446 195 11
9 48 yr 2 4 197 73
10 41yr 2 12 198 20
11 34yr 1 374 232 62
12 42yr 4 4 241 22
13 31yr 2 258 245+32
14 23yr 0 320 260 2
15 31yr 1 157 287 35
16 17mo 3b 85 299 24
17 25 mo 3b 354 322 51
18 37 yr 3 6 335 20
19 34 yr 3 20 343 56
20 45yr 2 58 353±151
21 42yr 3 12 398 17
22 35yr 3 45 624 45
23 34yr 1 50 853 248
aNeurologic dysfunction (ADCstage) (46a) ratingwasdonein accordance
with theguidelinesofthe AmericanAcademy of Neurology Task Force on AIDS
(26).
bPediatric patientswereassigned"best-approximation" ADC stages to con-form with the adult data. All of the subjects were male.
recovery,based on inclusion of internal standards of
[3H]PAF
during extraction. The material was dissolved in 1.2 ml of methanol and aliquoted into 0.1-, 0.4-, and 0.6-ml fractions. These were dried by vacuum centrifugation (Savant Instru-ments, Inc.,Farmingdale, N.Y.), and PAF was quantitated by radioimmunoassay (E.I.Du Pontde Nemours & Co., Boston, Mass.).The detectable levels ofPAFin these assayswere30to 1,200pg/ml (20).
Patient material. Forty-four patients were clinically evalu-atedattheUniversityofNebraska, theUniversity of Medicine and Dentistry of New Jersey, the Wadsworth VA Medical Center (UniversityofCalifornia Los Angeles), and the Mas-sachusetts General Hospital. Patients were retrospectively evaluated. Nine HIV-1-seropositive patients had complete postmortem examinations. The HIV-positive patients ranged in age from7monthsto61years(Table1).Twenty-one control patients were between 13 and 78 years old (Table 2). The HIV-seropositive patients were classified as to severity of neurologic dysfunction in accordance with ADC guidelines (46a) and those established by the American Academy of Neurology TaskForce onAIDS (26).
Humanneuronal cellcultures. Humanfetalbrain tissuewas obtained from elective therapeutic abortions under thestrict ethicalguidelinesof theNational Institutes of Health and the University of Rochester Medical Center. Neurons were ob-tainedfrom thetelencephalonwith both cortical and ventric-ularsurfaces ofsecond-trimester
(13
to 16weeksofgestation)
humanfetal brain tissuebyamodification of the
procedure
of Banker andCowan (1).Briefly,brain tissuewascollected and washedin30 ml of coldHanks balanced salt solution(contain-ing
Ca2+, Mg2+,
HEPES[N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid],
and 50 ,ug ofgentamicin
perml).
Brain tissue was separated from adherentmeninges
and blood andVOL. 68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
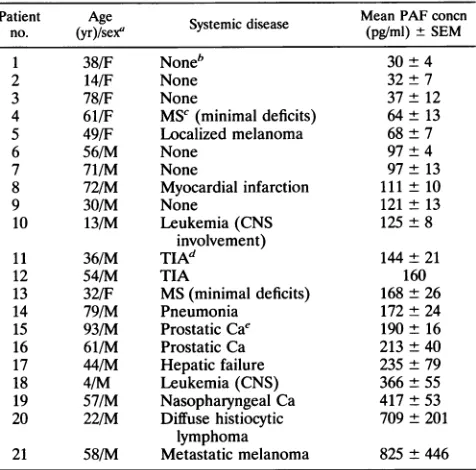
TABLE 2. PAFlevelsin CSF of non-HIV-1 disease and controlsubjects
Patient Age . Mean PAFconcn
no. (yr)/sex' Systemic disease (pg/ml) ±SEM
1 38/F Noneb 30 ± 4
2 14/F None 32+7
3 78/F None 37 ± 12
4 61/F MSC(minimal deficits) 64 ± 13
5 49/F Localized melanoma 68 ± 7
6 56/M None 97+ 4
7 71/M None 97 + 13
8 72/M Myocardial infarction 111 ± 10
9 30/M None 121 ± 13
10 13/M Leukemia(CNS 125 ± 8
involvement)
11 36/M TIAd 144 ± 21
12 54/M TIA 160
13 32/F MS(minimal deficits) 168 ± 26
14 79/M Pneumonia 172 ± 24
15 93/M ProstaticCae 190 ± 16
16 61/M Prostatic Ca 213±40
17 44/M Hepatic failure 235±79
18 4/M Leukemia(CNS) 366±55
19 57/M Nasopharyngeal Ca 417 ±53
20 22/M Diffusehistiocytic 709± 201
lymphoma
21 58/M Metastatic melanoma 825 + 446
a F, female; M,male.
bForsubjectswithoutsystemicdisease, lumbarpunctures wereperformedto
excludemeningitis. Patientspresented withheadache, fever, and/orweakness.
CSFexaminations showed normalproteinandglucoselevels andcellcounts.
c MS,multiple sclerosis.
dTIA,transient ischemicattack.
'Ca, carcinoma.
cut into 2-mm3 pieces. Dissociated tissue was transferred to andthenforcedthrougha230-jimNitexbag.Thedissociated tissuewasgentlytriturated and thencentrifugedat100xgfor 5 min at 4°C. The pellet was resuspended in 5 to 10 ml of MEM-hipp (2 mM D-glucose, 10 mM HEPES, 1 mM Na pyruvate,20 mMKCI) containingNicomponents(insulinat5 mg/liter,transferrinatS mg/liter,seleniteat5 ,ug/liter, proges-teroneat 20nM, andputrescine at 100 ,uM),aswell as 10% fetal calf serum, PSN antibiotic mix (penicillinat 50mg/liter, streptomycinat50mg/liter,andneomycinat100mg/liter),and amphotericin B(Fungizone;2.5 mg/liter). The cellcountand viability were determined by diluting Hanks balanced salt solutionwith0.4%trypanblue (1:1, vol/vol)andcountingwith ahemocytometer.Cellsweregentlytriturated fivetimes witha 10-ml pipet, plated at a densityof 105/12-mm-diameter glass coverslipprecoated withpoly-L-lysine (70K-1SOKMW;Sigma), andplacedin 24-wellculture dishes. Cells were cultured for 10 to28daysat37°Cin a humidified atmosphere of 5%C02-95% air,andthe medium was changed every 3 days (7).
Under these conditions, neuronal cultures were >70% homogeneousas determined by anti-human PGP 9.5 staining (Ultraclone, Ltd., Wellow, Isle of Wight, England) (28) and anti-MAP-2 staining. Glial fibrillary astrocyte protein staining-positiveastrocytes made up25% of the total cell population. Microglia-macrophages made up less than 5% of the popula-tionas determined by RCA-1 lectin and CD68 staining.
Neuronal cultures maintained for >28 days under the conditions described above expressed NMDA receptors. Neu-rons exposed to 300
jiM
NMDA for 10min were reduced in number to20% of that of untreated neurons. This neurotox-icitywasblocked by coincubation with 10jiM
MK-801 during exposure to NMDA (data not shown). Evaluation ofPAF-induced humanneurotoxicitywasperformedwith
1-O-hexade-cyl-2-O-acetyl-sn-glycero-3-phosphorylcholine
PAF(Biomol
Research
Laboratories,
Inc.,Plymouth Meeting, Pa.).
Coverslips
with human neurons were fixed with 4% paraformaldehydeat48 h after exposuretoculture fluids from HIV-iADA-infected monocytes and/or astroglial cells. Cells were incubated with Neurotag Red(Boehringer Mannheim,
Indianapolis, Ind.)
tostainsomasand neuritic processes. Cell morphologyofneurons wasexaminedby
fluorescence micros-copy. Inreplicate
experiments,
neurons were evaluated as described aboveat48 h after exposuretoPAFwithorwithout MK-801. Cellswereimmunocytochemically
stained with PGP 9.5 antiserum. To assayneurotoxicity,
examiners(K.A.D.,
H.A.G.,and
L.G.E.)
wereblindedtothe neuronaltreatments. All assayswereperformed
intriplicate
determinations.Digi-tized
images
of PGP 9.5-stained neurons in .15microscopic
fieldswere
analyzed
for numbers of intact neuronalsomasper 5OX field with a densitometer(Imaging
ResearchInc.,
St.Catharines,
Ontario,
Canada).
Datawereexpressed
as meanneuronal cell counts ± the standard errors of the means
(SEM).
Tests of statisticalsignificance
of the differences between control andexperimental
treatments were deter-minedby paired
ttests.Ratneuronal cell cultures. Ratretinal
ganglion
cells from 7-to10-day-old
postnatal Long-Evans
rats werelabeled insituby
retrograde
transport of the fluorescentdye granular
blue. Sinceganglion
cellsaretheonly
cellstoproject
from the retina to deeperbrain structures, thislabeling
wasaccomplished by
injectionofgranular
blue(as
a1% solution insaline)
into thesuperior colliculus;
maximumlabeling
of retinalganglion
cells occurred within 2days
of theinjections,
so theretinas were harvestedatthis time after therats weresacrificedby
cervical dislocation(36).
With theretrograde
labeling technique,
theganglion
cell neurons could bespecifically
identifiedby
the presence of the bluefluorescentlabel,
evenfollowing
dissoci-ation from the retina andplacement
inculture,
aspreviously
described(31, 36).
Theresulting
retinal cultures contained mixed neuronal andglial
elements. These cells wereplated
ontoglass
coverslips
coated withpoly-L-lysine
and culturedin Eagle's minimumessential mediumsupplemented
with 0.7%(wt/vol)
methylcellulose, 2 mMglutamine,
1 mgofgentamicin
per
ml,
16 mMglucose,
and 5%(vol/vol)
rat serum. This culturemediumcontained 1.8mMCaC12
butnoMgCl2,
since atnegative
membranepotentials magnesium
is knowntoblock NMDAreceptor-operated
channels andthereforemight
have partially masked NMDAreceptor-operated
channels and NMDAreceptor-mediated neurotoxicity
(4,
22,39,
44).
Inparallel experiments, however, qualitatively
similar results wereobtained with aphysiologic
concentration ofMg2+
(0.8mM). Replicate
cultures received PAF with or without the NMDAantagonist
MK-801(dizocilpine)
ormemantine. Ret-inalcultures treated in thismanner were incubatedovernight at37°C
in ahumidifiedatmosphereof5%C02-95%
air,and thenviability
was assessed as described below. For theseexperiments,
carbamyl-PAF(c-PAF;
Biomol ResearchLabo-ratories, Inc.)
wasusedto avoidrapidmetabolism ofPAFby tissuehydrolases (45).
The existence of PAF degradation products might have obfuscated the results of the neuronal assays.Inthis respect,wewouldnothave known ifPAFitself or ametabolitewas aneurotoxic factor in this system. c-PAF hasapharmacologic profilesimilartothat ofPAFbut is 5-to 10-fold less potent in binding and physiological studies of membrane surface receptors for PAF (45). Previous studiesusing high-pressure liquid chromatographic analysis
showed that these retinal cultures containapproximately
25jiM
glu-tamate(an
NMDA receptoragonist)
and 15jiM glycine (an
on November 9, 2019 by guest
http://jvi.asm.org/
HIV-MEDIATED NEUROTOXICITY 4631
NMDA receptor coagonist), but by themselves these levels of the amino acids are not neurotoxic in this system (35).
The ability of retinal ganglion cells to take up and cleave fluorescein diacetate to fluorescein was used as an index of their viability and lack of injury, as described previously (17, 31). Under a combination of phase-contrast microscopy and UV epifluorescence, retinal ganglion cells are the only cells that appear blue with a standard 4',6-diamidino-2-phenylin-dole filter set because of the presence of retrogradely trans-porteddye; viablecells, visualized with a fluorescein filter set, appearyellow-green. Thus, viable retinal ganglion cell neurons appear blue with one fluorescence filter set and yellow-green withasecond, facilitating their rapid scoring. This test appears tobe a more sensitive indicatorof subtle forms of cell injury than dye exclusion studies, which have been used to monitor cell death inmanystudies.Results werereplicated in triplicate dishes on at least four separate occasions. Viable rat retinal ganglion cellswerescored ineach culture dish over an area of 75 um
2.
RESULTS
PAFdetected in HIV-infected monocyte-astroglial mixtures. Our previous work demonstrated that tumor necrosis factor alpha (TNF-oz), interleukin 1B, arachidonic acid metabolites, and PAF are produced during interactions between HIV-infected monocytes and astroglia tumor cell lines. These factors predict neurotoxicity (20). The contribution that each of these factors made toneurotoxicitywas not demonstrated. Therefore,weperformedaseries of experimentsto determine whetherPAF isaneurotoxin associated with HIV-1 infection andneurological disease.
PAF levelswere examined by radioimmunoassay in HIV-1-infected cell cultures. Monocytes were infected with HIV-lADAat amultiplicity of infection of0.01 for7days then added in a 1:1 cell-to-astroglia ratio (U251 MG cells). Methanol extracts of cells were prepared during monocyte-astroglial cocultureat 1,3, 6, 12, 20, and 30min.Asshown inFig. 1, high levels of PAF were selectively observed (3 to 20 min) in cocultures of HIV-infected monocytes and astroglia. PAF concentrations were significantly lower (i) in HIV-1-infected monocytes or (ii) in uninfected monocytesin the presence or absence ofastroglia. PAFcould notbe detected reproducibly atlonger time intervals(hourstodays) (datanotshown).This mayhave beenduetothe temporal regulationofPAFandits instability in culture solutions for longer time intervals(3, 8,9, 20). Alternatively, PAF may be synthesized only after the initial cell-to-cell contact between HIV-1-infected monocytes and astroglia.
Measurement of PAF in CSF of HIV-1-infected control subjects. CSF samples from 23 patients with documented HIV-1 infection and 21 controlswere collected for
measure-mentofPAF levels (Tables 1 and2). Neurologic dysfunction, defined by the clinicalparameters oftheADC,wasscoredon
a scale of 0 to 4. Immunosuppression was measured by determining numbers ofCD4+ Tcells inblood. Six of the 23 patientswerechildren,with agesrangingfrom7monthsto10 years. Inthepediatricgroup, threeofsixchildren had elevated PAF levels (152 to 322 pg/ml),
correlating
withneurologic
dysfunction and immunosuppression
(Fig.
2 and Table1).
In adultpatients, however,elevatedPAFlevels(124
to853pg/ml)
correlatedin 17of23 patientswith theonsetbutnot progres-sion ofneurologic
dysfunction (ADC
stages 1to4).
PAF levels inCSF from this groupofpatientsweresignificantly
different fromthose inCSFfrom control patients (P<0.0002).
Five of 23patientswithnoneurologic dysfunction
(ADC
stage0)
had260
195
I..
1-1
, 1 30
0c
65
-0
t'
I
,~~~~~%
.';
,~~~~~%
v~~~~~%
v~~~~~~~~%
8 16 24 32
Time (minutes)
FIG. 1. Monocytes cultured for 7 days as adherent monolayers wereexposed to
HIV-lAI1A
at a multiplicity ofinfection of 0.1. At 7 days after infection, HIV-infected or control uninfected monocytes werecocultured 1:1 (monocytes-astroglial cells). At various times (in minutes) after coculture, HIV-infected monocytes, uninfected mono-cytes, ormonocyte cocultures werefrozen on dry ice in ethanol and then celllysateswerecombined with methanol washes of broken cells and extracted. PAF was eluted withapproximately 70% recovery andquantitated by radioimmunoassay. The detectable levels ofPAF in these assayswere30to1,200pg/mI.Levels ofPAFareshownasclosed diamonds (HIV-1-infected monocytes and astroglial cells), closed circles(uninfectedmonocytes andastroglial cells),open boxes
(HIV-infected monocytes), and open circles (uninfected monocytes). The
errorbarsrepresent standard deviations.
low levels of PAFinCSF (61 to 195 pg/ml)(Fig. 3 and Table 1). Elevation of PAF in CSF was not specific for HIV-1-infected individuals. PAF was detected in patients with a variety of other medical conditions (multiple sclerosis, cere-brovascular disease, leukemia, disseminated cancer, and he-patic failure) in whom CNS symptoms (changes in cognitive and/ormotorfunctions) werepresent. Patientswith paraneo-plastic syndromes that included CNS dysfunction (Table 2, patients 18 to 21), interestingly, had the highest PAF levels (366 to 825 pg/ml) ofthe non-HIV patients. This includes a
single leukemic patientwith CNS involvement. PAF levels in the CSF of this group of patients were also significantly differentfrom those in the CSF of controlpatients (P<0.006; Fig. 3).
Neurotoxicactivityof PAF.Couldtheobservedelevation of PAF in CSFmean that it plays an important role(s) in CNS dysfunction?Totestthistheory,we
exposed primary
neuronal fetal cultures to PAF at concentrations equal to andgreater thanthose present in coculture fluids. PAF(50to6,000 pg/ml) appliedtoprimaryfetalcultureswastoxictohumanneuronsina dose-dependent manner within 48 h
(Fig. 4).
At PAF concentrations of300 and1,250 pg/ml,
neuronal cell countswere reduced to 54 and
28%,
respectively,
of the control (untreated neurons) value. The PAF-inducedneurotoxicity
VOL.68, 1994
0t
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.324.559.81.338.2]300-1
0
0
I T
0
0
normal ADC stage 0 ADC stage 1-4
FIG. 2. Box plots of PAF levels in CSF obtained from control
(HIV-1-seronegative) patients without systemic disease (n = 6),
HIV-1-seropositive patientswithout evidence ofneurologic dysfunc-tion (ADC stage 0; n = 5), and HIV-1-seropositive patients with
neurologic dysfunction (ADCstages 1 to4;n = 18). Comparisons by
t test: controlversusADC stage0group,P< 0.025; control versus
ADCstage1to4group,P<0.0002;ADC stage 0groupversusADC
stage1to4group,P<0.05. Because threecomparisonsweremade, the level ofsignificancewasset ata=0.05/3 =0.017.Therefore, only
the difference between control patients and the ADC stage 1 to 4
group wassignificant.
4Ji
03
0 0
z
250
-200
150
100
50
o0
*
_~~~
0 50 300
PAF (pg/mi)
1250
FIG. 4. Dose response of human fetal neuronal cell cultures to
PAF. Culturesweregrownoncoverslipsasdescribedin Materialsand
Methods.Human cortical neuronalcell cultureswereincubated for48
hwith thevehicle aloneorwithincreasingdosesof PAF(50, 300,and
1,250pg/ml [97 pM,557pM,and 2.3nM, respectively]).The cultures
were washed, fixed in paraformaldehyde, and subjected to PGP 9.5
immunocytochemical labeling.Neuronal cellcountingwasperformed asdescribedinMaterials and Methods. Observerswereblinded tothe
treatment groups. Each neuronal cell count shown represents the
meanof .15microscopicfields selected from fourreplicates± SEM.
Symbols: *,P < 0.0001, control versusPAF at 300 pg/mI; +,P <
0.0001,controlversusPAFat1,250 pg/ml.
was partially blocked by coincubation with 10 ,uM MK-801 (Fig.5A).The numbers ofneuronstreated with both PAF and MK-801 were 80% of the numbers of untreated control neurons.
Similar results were obtained with rat postnatal retinal cultures. Retinalganglioncellswerespecifically depletedfrom cultures containing c-PAF. Assay ofrat retinal ganglion cell viabilitywasbasedupontheability of healthyneuronstotake
up and cleave fluorescein diacetate to fluorescein. Retinal ganglioncellswerespecificallylabeledby retrogradetransport
ofafluorescentdye injectedinto thesuperiorcolliculus. Viable
retinalganglion cellsweredual labeledwithgranular blueand
fluoresceinyellow.A 35% reduction ofneuronalviabilitywas
1000
8001
.
600-
U-<
400-200
u
0
[image:5.612.321.552.77.219.2]systemic diseases normal
FIG. 3. Boxplot of PAF levels in CSF ofcontrol (HIV-1-seroneg-ative) patients without systemic disease (n = 6)and patients (HIV-1
seronegative) witha variety of systemic diseases (metabolic, cancer, andothers;n = 15). Comparisonbyttestof control patientsversus
patientswithsystemic disease, P<0.006.
observed following addition of c-PAF and was largely
pre-ventedby the specificNMDAantagonist MK-801 or
meman-tine (each at6 ,uM) (Fig. 5B). Thus, PAF-induced neurotox-icity was seen in both human and rat neuronal cell culture
systems.
DISCUSSION
Productive HIV infection of the brain occurs in brain
macrophagesandmicroglia. Cognitive andmotordysfunction
occurs, as well as neuronal loss in infected brain tissue.
HIV-1-infectedbrain macrophagesandmicrogliamayleadto HIV-1-associated dementia, but neurologic deficits may not directly correlate with productive viral infection. A discor-dance exists between the level ofhistopathologic changes and the small numbers of detectable HIV-infected cells. This
supports the notion that diffusible virus-induced neurotoxins
areresponsible, inpart,for theneurological manifestations of HIV-1infection.Moreover,thishypothesis remains likelyeven
if farmoremicroglialcells andastrocytes(48)areinfected with
HIV-1 and low-level or restricted infection results in local
neurotoxic activities. Herewedemonstrate thatPAFisoneof
possibly many HIV-induced neurotoxins. This was shown by
severalassaysystems.First,wedetected PAF inan
experimen-tal model systemforHIVCNS disease. The levels ofPAFin
ourlaboratorysystemwerehigh butnotsustained, supporting thenotion of itscyclical production. Second,weconfirmed the
biological relevance ofour in vitro observations by assay of
PAF in the CSF of HIV-infected patients. Although PAF elevations were not specific for HIV infection and CNS disease, they did correlate with the onset of neurological impairment. Finally, we inoculated PAF, in concentrations
similartothose found inCSF,intoprimaryneurons.Here PAF
was a potent neurotoxin. These findings, taken together, stronglysuggestthat PAFplaysarole inADC. Moreover,the
dataprovidetheidentityofasolubleneurotoxin, distinct from
aviralgeneproduct, produced by HIV-infected cells.
PAF is alipid mediatorthat initiates a large repertoire of
1000
-800
-E
600--
400-0.
200
-
0-..:.."
71
L-1 .. t...'iiiiLl
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.60.295.77.238.2] [image:5.612.65.295.520.681.2]HIV-MEDIATED NEUROTOXICITY 4633
A
200-c
0
-a
c
0
0
z
150
-100
-50
0
+
Control
II
c
0
coCD=
0
c 4-. -C
0
0
4-0 .0
B
120- 100- 80- 60- 40- 20-
o0-*
PAF PAF/MK-801
77 --F~1
[image:6.612.57.299.65.354.2]PAF PAF/MK-801 PAF/memantine
FIG. 5. Neurotoxic effects of PAF onhuman fetal neuronal cell
cultures(A)andratpostnatalretinalganglioncells inculture(B)with
partialblockade ofneurotoxicity byNMDAantagonists.Cultureswere grown on coverslips as described in Materials and Methods. (A)
Humancorticalneuronal cultureswereincubated with PAF for48 h.
The cultures were then washed, fixed in paraformaldehyde, and
subjected to PGP 9.5 immunocytochemical labeling. Neuronal cell
counts were determined as described in Materials and Methods.
Observerswere blinded tothe treatmentgroups. Each neuronal cell
count shown representsthemeanof .15 microscopicfields selected
from fourreplicates±SEM. PAFwasusedat6,000pg/ml (10.9 nM);
MK-801 was usedat 10,uM. Symbols: *,P< 0.0001, controlversus
PAF; +,P< 0.0001, PAFversusPAF-MK-801. (B)Rat retinal cell
cultures were incubated in c-PAF overnight. Retinal ganglion cell
neurons werespecificallyidentified with thefluorescentdye granular
blue andthen scored forviabilityinamaskedfashiononthe basis of
theabilitytotakeupand cleavefluorescein diacetatetofluorescein.
Thus,viable retinalganglioncellsweredual labeled withgranularblue
andfluoresceinyellow.c-PAF(54 ng/mlor100nM) injured32% of the
retinalganglioncells(68%remainedviable). MK-801 ormemantine
(eachat6 p,M) largely preventedthec-PAF-inducedneuronalinjury.
In the experiments whose results are shown, all of the cultures contained 25p,M glutamate.Control dishes containedapproximately
150 viable retinalganglioncellneuronsandwereusedtonormalize the data obtained from each of fourseparate experiments.
pathophysiologicresponses,including bronchoconstriction, hy-potension, neutropenia, thrombocytopenia,and increased
vas-cularpermeability leadingtocardiac and renalimpairment,as
well aspulmonary edema, anaphylaxis,and death(8, 9). PAF has diverseeffects at the cellularlevel, including phagocytosis, exocytosis, aggregation, chemotaxis, calcium mobilization, ei-cosanoid production, and superoxide production (24, 25). Theseresponsesaremediatedby G-protein-coupled receptors (24, 25). PAFhasbeen shown to induce transient increases in
intracellular calcium in cultured neuronal cells (3, 30). PAF
can facilitate excitatory neurotransmission andglutamate
re-lease in mammalian brains(3, 5, 37, 51, 58). Thus,
experimen-tal evidence suggests that PAFplays important roles in neu-ronal function. In contrast to its beneficial effects, it remains possiblethat PAF contributes to brain injury. This could occur by different mechanisms, including superoxide production, increased calcium mobilization, or by glutamate elevation. Theseeventswouldcollectively injure susceptible neurons.
Thepathophysiologic relevance of PAF in HIV-1 infection of the CNS was first demonstrated by analyses of HIV-1-infectedmonocyte-astroglia mixtures. Cytokines, most notably TNF,are detected in braintissues of patients with advanced ADC(54). Reports demonstrate that TNF-a and interleukin 113 are regulated by PAF and arachidonic acid metabolites (23). Indeed, inhibitors of arachidonic acid metabolites, such as dexamethasone and nordihydroguaiaretic acid, decrease TNF-aproduction in HIV-1-infected cultures (20) and dimin-ish neurotoxic activities (14a). Furthermore, PAF and arachi-donic acid metabolitesare potentstimuli for TNF-a produc-tion. The rise in cytokines, PAF, and arachidonic acid metabolites in these HIV-1-infected cell systems correlates with neurotoxicity, suggesting that all of these factors play importantroles inprogressiveHIV-1 CNS disease.
PAFproduction in HIV-1-infectedmonocytes was ephem-eral.Thiswasreported previously for arachidonic acid metab-olites induced in gp120-stimulated monocytes (56). Indeed, detection of PAF was difficult at later time points (hours and/or days). There are several explanations for this result, including the following: PAF biosynthesis may occur only during initial cell-to-cell contact between HIV-1-infected monocytes and astroglial cells; uninfectedor infected mono-cytes maysecrete afactor that inhibitsastroglialcell-mediated biosynthesisofPAF;and enzymesimportant inthe generation of PAF may be expressed at low levels in HIV-1-infected monocytesandastrogliabut mayundergocell-to-cell transfer sothat PAFbiosynthesisis increasedtransiently(38).
Despitetheevanescent natureof PAFmetabolisminthese experimental conditions,themarked increasein PAF biosyn-thesis inthe CSF of HIV-1-infectedpatientsand the ability of PAFtostimulate TNFproductionsuggestthatPAFis biolog-ically relevant for ADC. One of the advantages of the CSF assaysis thatspecimenswereobtainedwith minimalrisk to the patientsandlikely reflected ongoingbrainpathology. The level ofPAFintheCSF represents the average ofdifferentratesof
production
and degradationofPAF in all brain areas.Thus, levels of PAF intheCSFmaybe different from those observed inspecific
neocorticalareasinvolved inADC.Notwithstanding thislimitation,
severalpotentially importantobservationswere made.First, in pediatric patients, elevatedPAFlevels in CSF correlated with severe encephalopathy. Second, in adultpa-tients,
elevated PAF levels inCSF correlatedwith bothimmu-nosuppression
and onset ofneurological impairment
(ADC stages 1 to4).
PAFwas notspecific
forneurological
manifes-tations of HIV infection. PAF in CSFwaselevated inavariety of non-HIVdiseases with CNSmanifestations,
demonstrating that it is not aspecific
marker for HIV-1-associated CNSdysfunction.
PAF
produced dose-dependent neurotoxicity
(Fig. 4).
Neu-rotoxicity
associated with PAFwasblocked,
in part,by
coin-cubation with the NMDAantagonist
MK-801 ormemantine,demonstrating
that PAFcandirectly
activate NMDA receptors withsubsequent
excitotoxic neuronaldamage.
PAF canalso increase neuronalCa2"
and lead to enhancedexcitatory
neurotransmission
through
increasedglutamate
release(3,
5, 37,51,
58).
Inaddition,
recentreportshave demonstrated that arachidonic acidcaninhibithigh-affinity glutamate uptake
in synaptosomes and astrocytes(55)
andpotentiate
NMDA receptorcurrentsby increasing open-channel probability
(41). VOL.68, 1994T-r
I I~~~~~~~~~~~~~~~~~~~~~~
on November 9, 2019 by guest
http://jvi.asm.org/
These mechanisms might be responsible for an increase in glutamateinthesynapticcleft and subsequentNMDA
recep-tor-mediated excitotoxic damage to neurons in HIV-infected
brains. These findings, in toto, support a role of PAF in HIV-1-mediated neurotoxicity.This doesnotmeanthat PAF is
anexclusive HIV neurotoxin. It isgenerallybelievedthatmany
cellular and viral neurotoxins contribute to CNS disease. Indeed, arachidonic acid metabolitesmayplay equally impor-tant roles in HIV-1-induced brain disorders.
The abilityof NMDAantagoniststoblockneurotoxicityhas been a central feature in other models of HIV-1-mediated
neuronal damage (21, 35). The consequences of productive HIV-1infection, includingviralgeneproducts,e.g.,gpl20,may
stimulate arachidonic acid metabolite andcytokine production
in macrophages (56). These alterations in macrophage
secre-toryfunctionmay,inturn,result inneurotoxicityandsupport, for the first time, commonalityamong mechanismsfor HIV-1 neuropathogenesis.
ACKNOWLEDGMENTS
We thankKarenSpiegelforoutstandingadministrative support. We thank Michael P. McDermott for advice on statistical analysis and Bradford Navia and the National Neurological Research Specimen
Bankfor contributionofCSFsamples.
This workwas funded in partby PAF/AmFARgrants
500258-12-PG, 500278-14-PGR (H.A.G.), and 001433-11-R&R (S.A.L.), the
Strong Children's Research Fund (H.A.G.), Universityof Nebraska
Biotechnology start-up funds, and NIH grants PO1 NS31492-01 (H.A.G., L.G.E., and H.E.G.), P01 HD29587-01, ROI EY09024-03
(S.A.L.),and P01 HL43628-05(H.E.G.). Howard E. Gendelman isa
Carter Wallace Fellow of theDepartmentofPathologyand
Microbi-ologyof theUniversityof Nebraska Medical Center. Hans Nottet isa
Nicholas B. Badami Scholar of the Laboratoryof ViralPathogenesis, Department of Pathology and Microbiology, Universityof Nebraska
Medical Center.
REFERENCES
1. Banker, G. A., andW.M.Cowan.1977. Rathippocampalneurons
indispersed cell culture.Brain Res. 126:397-425.
2. Bernton, E., H. Bryant, M.Decoster, J.M.Orenstein, J. Ribas,
M.S.Meltzer, andH. E. Gendelman. 1992. No direct
neurono-toxicity byHIV-1virionsorculture fluids from HIV-1 infected T cellsormonocytes.AIDS Res. Hum. Retroviruses 8:495-503.
3. Bito, H., M. Nakamura, Z. Honda, T. Isumi, T. Iwatsubo, T.
Seyama, A. Segura, Y. Kido, and T. Shimizu. 1992.
Platelet-activating factor (PAF) receptor in rat brain: PAF mobilizes intracellularCa2+inhippocampal neurons.Neuron 9:285-294.
4. Chen, H.-S. V., J. W. Pellegrini, S. K. Aggarwal, S. Z. Lei, S.
Warach,F. E.Jensen, and S. A.Lipton. 1992. Open-channel block
of NMDAresponsesbymemantine:therapeutic advantage against
NMDA receptor-mediated neurotoxicity. J. Neurosci.
12:4427-4436.
5. Clark,G.D.,L. T.Happel, G. F. Zorumski, and N. G. Bazan. 1992.
Enhancement ofhippocampal excitatory synaptic transmission by platelet-activating factor. Neuron 9:1211-1216.
6. Cohen,S.E.,T.Mundy, B. Karassik, L. Lieb, D. D. Ludwig,and
J.Ward. 1991.Neuropsychological functioning in human immu-nodeficiency virus type 1 seropositive children infected through neonatalbloodtransfusion. Pediatrics 88:58-68.
7. Cole, R., and J. de Vellis. 1989. A dissection and tissue culture manual of the nervous system, p. 133. Alan R. Liss, Inc., New York.
8. Criscuoli, M.,andA.Subissi. 1988.Catecholamine released from
e adrenal medulla exert a compensatory, protective effect at
b2-adrenoreceptors against PAF-induced death in mice. Br. J.
Pharmacol. 93:132.
9. Darius, H., D. J. Lefer, J. B. Smith,and A. M. Lefer.1986.Roleof platelet-activating factor-acether in mediating guinea pig
anaphy-laxis.Science232:58.
10. Dawson, V. L., T. M. Dawson, G.R.Uhl,and S. H. Snyder.1992.
Humanimmunodeficiencyvirus-1coatprotein neurotoxicity
me-diated by nitric oxide in primary cortical cultures. Proc. Natl. Acad. Sci. USA90:3256-3259.
11. Dreyer, E. B., P. K.Kaiser,J.T. Offermann, and S. A. Lipton.
1990. HIV-1 coat protein neurotoxicity prevented by calcium channelantagonists.Science248:364-367.
12. Dzenko,K.A.,R.J. White,P.Genis,H.Nottet,L. G.Epstein,H. E.
Gendelman, and H. A. Gelbard. 1993. Increased secretion of
cytokines and neurotoxinsbycocultures of HIV-1 infected
mac-rophages andhumanfetal astrocytes. Soc.Neurosci. 19(3):1893. 13. Epstein,L.G.,andH. E.Gendelman.1993. Human
immunodefi-ciency virustypeIinfection of the nervoussystem:pathogenetic mechanisms.Ann.Neurol. 33:429-436.
14. Everall,I.P.,P.J. Luthbert,and P. L. Lantos. 1991. Neuronal loss in thefrontalcortexin HIV infection. Lancet337:1119-1121. 14a.Gendelman,H. E.Unpublisheddata.
15. Gendelman, H.E.,L.Baca,H. Husayni, J.M. Orenstein,D. C.
Kalter, J. A. Turpin, D. Skillman, D. L. Hoover, and M. S. Meltzer. 1990.Macrophage-humanimmunodeficiencyvirus inter-action: viral isolationand targetcelltropism.AIDS4:221-228. 16. Gendelman,H.E.,and S.Gendelman. 1992.Neurologicalaspects
of human immunodeficiency virus infection, p. 229-254. In S.
Specter, M. Bendinelli, and H. Friedman (ed.), Neurotropic viruses andimmunity.PlenumPress,NewYork.
17. Gendelman, H. E., P. Genis, M.Jett, and H. S. L. M. Nottet.
Experimental modelsystemsfor studiesofHIV-encephalitis.In E. Major (ed.),Technicaladvancesin AIDS researchin thenervous
system,in press. PlenumPress,NewYork.
18. Gendelman,H.E.,and P. M. Morahan. 1992.Themacrophagein viralinfections,p.156-232.In C. E.Lewisand J.0.McGee(ed.), The natural immune system series: the macrophage. Oxford UniversityPress,London.
19. Gendelman,H.E., J.Orenstein,M. A.Martin,C.Ferrua,G. H.
Pezeshkpour, T. Phipps, L. Wahl, A. S. Fauci, D. Burke, D.
Skillman,and M. S. Meltzer.1988. Efficient isolation and
propa-gation of human immunodeficiency virus on CSF-1 stimulated
macrophages.J.Exp. Med. 167:1428-1441.
20. Genis, P.,M.Jett,E. W.Bernton,H. A.Gelbard,K.Dzenko,R.
Keane, L. Resnick, D. J. Voisky, L. G. Epstein, and H. E. Gendelman. 1992. Cytokines and arachidonic acid metabolites
produced during HIV-infected macrophage-astroglial interac-tions: implicationsforthe neuropathogenesis ofHIV disease.J.
Exp. Med. 176:1703-1718.
21. Giulian, D., K. Vaca, and C. A. Noonan. 1991. Secretion of neurotoxins by mononuclear phagocytes infected with HIV-1.
Science250:1593-1596.
22. Hahn, J. S., E. Aizenman, and S. A. Lipton. 1988. Central mammalian neurons resistant to glutamate toxicity are made
sensitivebyelevatedextracellular calcium:toxicityblockedbythe
N-methyl-D-aspartate antagonist MK-801. Proc. Natl. Acad. Sci.
USA 85:6556-6560.
23. Hogan, M. M., and S. N. Vogen. 1988. Production of TNF by rIFN-g-primed CC3HH/HeJ
(Lps")
macrophagesrequiresthepres-enceoflipid-A-associated proteins.J.Immunol.141:4196-4199. 24. Honda, Z., M.Nakamura,I. Miki,M.Minami,T.Watanabe,Y.
Seyama,H.Okado, 0. Toh,K.Ito,T.Miyamoto,and T.Shimizu. 1991.Cloning byfunctionalexpression of platelet-activatingfactor receptorfromguinea pig lung.Nature(London)349:342. 25. Hwang,S. B. 1990.Specificreceptorsofplatelet-activating factor,
receptor heterogeneity, and signal transduction mechanisms. J.
Lipid Mediators2:123.
26. Janssen,R. S.,D.R.Cornblath,L. G.Epstein,R. P. Foa, J. C.
McArthur,R. W. Price,A. K.Asbury,A.Beckett, D. F. Benson, T.P.Bridge,C. M.Leventhal,P.Satz,A.J.Saykin,J. J. Sidtis, and S. Tross. 1991. Nomenclature and researchcasedefinitions
forneurologicalmanifestations ofhumanimmunodeficiencyvirus
type-1 (HIV-1) infection. Report of a Working Group of the American AcademyofNeurology AIDS Task Force.Neurology 41:778-785.
27. Kalter, D. C., M. Nakamura, J. A. Turpin, L. M. Baca, C.
Dieffenbach,P.Ralph,H. E.Gendelman,and M.S. Meltzer. 1991. Enhanced HIVreplicationin MCSF-treated monocytes. J. Immu-nol. 146:298-306.
on November 9, 2019 by guest
http://jvi.asm.org/
HIV-MEDIATED NEUROTOXICITY 4635 28. Kent, C., and P. J. Clarke. 1991. The immunolocalisationof the
neuroendocrine specific protein PGP9.5 during neurogenesis in therat. Dev. Brain Res. 58:147-150.
29. Ketzler, S., S. Weis, H. Haug, and H. Budka. 1990. Loss ofneurons
inthe frontalcortexinAIDS brains. Acta Neuropathol. 80:92-94. 30. Kornecki, M., V. Hogan, and Y. H. Ehrlich. 1993. Long-term potentiation in the hippocampus induced by platelet-activating factor. Neuron 10:553-557.
31. Leifer, D., S. A. Lipton, C. J. Barnstable, and R. H. Masland. 1984. Monoclonal antibodytoThy-1 enhances regeneration ofprocesses byrat retinal ganglion cells in culture. Science 224:303-306. 32. Lindgren, J. A., T. Hokfelt, S.-E. Dahlen, C. Patrono, and B.
Samuelsson. 1984. Leukotrienes in theratcentralnervoussystem.
Proc.Natl. Acad. Sci. USA81:6212-6216.
33. Lipton, S. A. 1993. Humanimmunodeficiencyvirus-infected
mac-rophages, gp120, and N-methyl-D-aspartate receptor-mediated
neurotoxicity. Ann. Neurol. 33:227-228.
34. Lipton, S. A., and F. E. Jensen. 1992. Memantine, a clinically-tolerated NMDAopen-channel blocker,prevents HIVcoat pro-tein-induced neuronal injury in vitro and invivo. Soc. Neurosci. Abstr.18:757.
35. Lipton, S. A., N. J. Sucher, P. K. Kaiser, and E. B. Dreyer. 1991. Synergistic effects of HIV coat protein and NMDA
receptor-mediated neurotoxicity. Neuron 7:111-118.
36. Lipton, S. A., and D. L.Tauck. 1987. Voltage-dependent
conduc-tancesofsolitaryganglion cells dissociated from therat retina. J. Physiol. (London) 385:361-391.
37. Marcheselli, V. L., and N. G. Bazan. 1993. Platelet activating factor(PAF) enhancesglutamic acid release in the retina through apresynapticreceptor. Invest. Ophthalmol. Vis. Sci. 34:1048. 38. Marcus, A. J., L. B. Safier, H. L. Ullman, N. Islam, M. J.
Broekman, J. R. Falck, S. Fischer, and C. von Schacky. 1988. Platelet-neutrophil interactions:
(12S)-hydroxyeicosatetraen-1,20-dioic acid:a neweicosanoid synthesized by unstimulated
neutro-phils from(12S)-20-dihydroxyeicosatetraenoicacid.J. Biol.Chem. 263:2223-2229.
39. Mayer, M. L., G. L. Westbrook, and P. B. Guthrie. 1984. Voltage-dependent block by Mg2+ ofNMDA responses in spinal cord neurons.Nature (London) 309:261-263.
40. Michaels, J., L. R. Sharer, and L. G. Epstein. 1988. Human immunodeficiency virus type 1 (HIV-1) infection of the nervous system:a review. Immunodef. Rev. 1:71-104.
41. Miller, B., M. Sarantis, S. F. Traynelis, and D. Attwell. 1992. Potentiation of NMDA receptor currents by arachidonic acid. Nature (London) 355:722-725.
42. Myers, G., J. A. Berzofsky, B. Korber, andR.F.Smith(ed.).1991.
Human retroviruses and AIDS, p. I A 95. Los Alamos National
Laboratory, Los Alamos, N.Mex.
43. Navia, B. A., B. D. Jordan, and R. W. Price. 1991. The AIDS
dementiacomplex: I.Clinical features. Ann. Neurol. 19:517-524. 44. Nowak, L.,P.Bregestovski,P.Ascher,A.Herbet,and A. Prochi-antz. 1984. Magnesium gates glutamate-activated channels in
mousecentralneurons. Nature(London) 307:462-465.
45. O'Flaherty, J. T., J.R.Redman,Jr., J.D.Schmitt, J.M.Ellis, J.R.
Surles, M. H. Marx, C. Piantadosi, and R. L. Wykle. 1987.
1-O-Alkyl-2-N-methylcarbamyl-glycerophosphocholine: a biologi-cally potent, non-metabolizable analog ofplatelet-activation fac-tor.Biochem. Biophys. Res. Commun. 147:18-24.
46. Poubelle,P.E.,D.Gingras,C.Demers,C.Dubois,D.Harbour, J.
Grassi,and J. Rola-Pleszczynski. 1991. Platelet-activatingfactor
(PAF-acether) enhances the concomitant production of tumor
necrosis factor-alpha and interleukin-1 by subsets of human monocytes. Immunology 72:181-187.
46a.Price,R. W., and B. J. Brew. 1988. TheAIDSdementiacomplex.
J. Infect.Dis. 158:1079-1083.
47. Pulliam, L., B. G. Herndier, N. M. Tang, and M. S. McGrath.
1991. Humanimmunodeficiency virus-infectedmacrophages
pro-duce soluble factors that cause histological and neurochemical alterations in cultured human brains. J. Clin. Invest. 87:503-512.
48. Saito,Y., L.R. Sharer, L. G. Epstein, J. Michaels, M. Mintz, M. Louder, K.Golding,T. A.Cvetkovich,and B. M.Blumberg. 1994.
Overexpression of nefas amarkerfor restrictedHIV-1infection of
astrocytes inpostmortem pediatric central nervoustissues.
Neu-rology44:474-480.
49. Sharer, L. R. 1992. Pathology of HIV-1 infection of the central
nervoussystem(review).J. Neuropathol. Exp.Neurol. 51:3-1 1.
50. Sharer, L. R., L. G. Epstein, E.-S. Cho, V. V. Joshi, M. F. Meyenhofer, L. F. Rankin, and C. K. Petito. 1986. Pathologic features of AIDSencephalopathy in children: evidence for LAV/
HTLV-III infectionof brain. Hum. Pathol. 17:271-284.
51. Shukla, S. D. 1992. Platelet-activatingfactor receptor and signal
transduction mechanisms. FASEBJ. 6:2296-2301.
52. Tardieu,M.,C.Hery, S. Peudenier,0.Boespflug, and L. Montag-nier.1992.Humanimmunodeficiency virustype1-infected
mono-cytic cellscandestroy human neural cells after cell-to-cell adhe-sion.Ann.Neurol. 32:11-17.
53. Tenhula, W. N., S. Z. Xu, M. C. Madigan, K. Heller, W. F.
Freeman,and A. A. Sadun. 1992. Morphometriccomparisonsof optic nerve axon loss in acquired immunodeficiency syndrome. Am.J.Ophthalmol. 15:14-20.
54. Tyor, W. R., J. D. Glass, J. W. Griffin, P. S. Becker, J. C. McArthur, L. Bezman, and D. E. Griffin. 1992. Cytokine
expres-sion in the brainduring the acquired immunodeficiency syndrome.
Ann. Neurol.31:349-355.
55. Volterra, A.,D.Trotti, P.Cassutti, C. Tromba, A.Salvaggio,R.C. Melcangi, and R. Racagni. 1992. High sensitivity of glutamate uptaketoextracellular free arachidonicacid levels inrat cortical
synaptosomes andastrocytes.J.Neurochem. 59:600-606. 56. Wahl, L. M., M. L. Corcoran, S. W. Pyle, L. 0. Arthur, A.
Harel-Bellan,andW. L. Farrar.1989. Human immunodeficiency
virusglycoprotein(gp120)inductionofmonocytearachidonic acid metabolites and interleukin 1.Proc.Natl. Acad. Sci. USA 86:621-625.
57. Westervelt,P., H. E.Gendelman,and L. Ratner. 1991. Identifica-tionofadeterminant within the humanimmunodeficiencyvirus 1 surfaceenvelope glycoproteincritical for productiveinfection of primary monocytes. Proc. Natl. Acad. Sci. USA 88:3097-3101. 58. Wieraszko, A.,G.Li,E.Kornecki,M.V.Hogan,and Y. H. Ehrlich.
1993. Long-term potentiation in the hippocampus induced by platelet-activatingfactor. Neuron 10:553-557.
59. Wiley, C. A., E. Masliah,M.Morey,C. Lemere,R.DeTeresa, M.
Grafe,L.Hansen,and R.Terry. 1991. Neocorticaldamageduring
HIVinfection.Ann.Neurol. 29:651-657.
60. Wiley, C. A., R. D. Schrier, J. A. Nelson, P. W. Lampert, and M. B. A.Oldstone. 1986.Cellular localization of human
immuno-deficiency virus infection within the brains ofacquired immune deficiencysyndromepatients.Proc.Natl. Acad. Sci. USA 83:7089-7093.
VOL. 68? 1994