0022-538X/92/105975-13$02.00/0
Copyright© 1992,American Society forMicrobiology
A
Novel
Oncogene, v-ryk, Encoding a Truncated Receptor Tyrosine
Kinase
Is
Transduced
into the RPL30 Virus
without
Loss
of
Viral Sequences
RANJIA, BRUCE J. MAYER, TERUKOHANAFUSA,ANDHIDESABURO HANAFUSA* The Rockefeller University, 1230 York Avenue, New
York,
New York10021-6399Received12 May 1992/Accepted 17 July 1992
The RPL viruses are acute oncogenic avian retroviruses isolated from chicken tumors. We carried out a genetic analysis of three of theviruses, RPL25, RPL28, and RPL3O. While RPL25 and RPL28 were shown to contain the erbB oncogene, RPL3O appeared to contain a novel protein tyrosine kinase oncogene. This gene,
v-ryk,wascloned and sequenced. The v-ryk oncogene contains a 1.39-kb nonretroviral sequence that includes
atyrosine kinasedomain which was inserted into the viral envelope protein gp37-coding region and fused in frame with upstream gp37 to generate a
p69m,37-y
fusion oncoprotein. Unlike that of otheracutely
transforming retroviruses, transduction of the v-ryk gene into RPL30 did not result in deletion of viral sequences. Sequence
analysis
suggested that v-Ryk is more homologous to receptor-type tyrosine kinases thantononreceptor-typekinases. By reconstitution ofavirus fromits cDNA, the v-ryk oncogene has been shown to be
fully
responsible for the transforming activity of theRPL30virus.Antibodies specific to v-Ryk immuno-precipitated the v-Ryk oncoprotein from cells transformed by theRPL30 virus. The v-Ryk protein was shownto be first synthesized as a 150-kDa precursor and then cleaved into the mature 69-kDa gp37-Ryk fusion protein, both parts of which were found to be localized to the membrane fraction. As expected from the sequence of v-Ryk, immunoprecipitates of v-Ryk from RPL3O-transformed cells were found to display a protein tyrosine kinase activity in vitro, and the levels oftyrosine-phosphorylated proteins are elevated in v-yk-transformed cells.
Acute oncogenic retroviruses have proven to be a rich
source of various categories of oncogenes. More than 25 viral oncogenes have beenmolecularly cloned and studied
(8). Sinceoncogenic retroviruses are the naturally selected
geneticsystemin whichimportantgenesinvolved in regula-tion ofproliferationanddifferentiation in normal cells were targeted, analysis of previously uncharacterized viral onco-geneswillbe valuable inidentifying novel cellular genes in these pathways that otherwise would be difficult to find.
Furthermore, identification ofnewoncogenesinretroviruses
notonly providesanimmediate indication that the genecan
be oncogenic, but it may also provide insights into the
mechanism of activation of the oncogenic potential of its
cellularhomolog.
The RPL25, RPL28, and RPL30 viruses are three acute
oncogenic avian retroviruses that were previously
unchar-acterizedgenetically. These viruseswerefirst isolated from chickentumors (21). Although theycause tumors in
chick-ens (21), the transforming genes present in these viruses havenotbeenidentified.Here,wereport thegenetic analy-sis of these three RPL viruses and themolecularcloningand
characterization of RPL30, which was found to contain a
novel viral oncogene. This viral oncogene isa newmember
of the protein tyrosine kinase gene family and appears to
encode a truncated receptor-type tyrosine kinase. It is
interestingthat unlike all oncogenicretroviruses studied so far, a cellular gene was transduced by the RPL30 virus without loss of any viral sequence.
*Correspondingauthor.
MATERIALS AND METHODS
Viruses, cells, and virus infection.TheRPL25,RPL28, and RPL30 viruses (21) were kindly provided by L. B. Crit-tenden of the Avian Disease andOncologyLaboratory,East
Lansing, Mich. The three viruseswere injectedinto newly
hatched chickens, and acutely transforming virus stocks
wererecovered from thesarcomasinduced in the chickens.
Preparation and maintenance of chicken embryo fibroblast
(CEF)
cultures and infection andpropagationof viruses have already been described (23). Briefly, CEF were preparedfrom 11-day-old embryos and secondary cultures were
in-fected with the virus stock.
Oncogene blotscreenofoncogenicviruses.Probes
specific
for 19 known viral oncogeneswere preparedfrom
molecu-larly cloned DNA, and they contained no retroviral
se-quences. The viral oncogene probes usedwere as follows: felinefms, 1.4-kb PstI fragment from pSM3 (14); chicken ros,0.85-kbEcoRI-PvuII fragmentfrompROS (32);chicken
erbB, 0.56-kb BamHI fragment from pERB5
(51);
murineabl, 1.6-kb SacI-HindIII fragment from pAB3sub3
(35);
feline fgr, 0.93-kb
SmaI-KpnI
fragment from pGR-FeSV (31);chicken src, 0.87-kbPvuII fragmentfrompTI107(44);chicken
fps,
0.41-kb BamHIfragment frompBR-FO4 (20);chicken yes, 0.43-kb BamHI
fragment
frompMR-YA(42);
murine mos, 0.92-kb AvaI-HindIII fragment from pCMOS
(46); chicken mil, 0.58-kb BamHI-SphI fragment from
pMBS28(27);ratH-ras, 0.46-kb EcoRIfragmentfrompBS9
(17); rat K-ras, 0.38-kb SacII-XbaI fragment from
pHiHi3
(18); woollymonkey sis, 0.92-kbPstI-XbaI
fragment
frompSSV (13); chicken
erbA-erbB,
1.2-kb SalI-BamHIfrag-mentfrompERBA(11); turkeyrel,0.97-kbEcoRIfragment
from
pEcoRI-rel
(6); chicken ski, 1.2-kb SacIfragment
from
pSRski-1
(29); murinefos,
1.0-kb PstIfragment
frompFOS1
(10);chicken myc, 0.8-kbClaI-EcoRIfragment
from5975
on November 9, 2019 by guest
http://jvi.asm.org/
5976 JIA ET AL.
pCMYC1 (2); chicken myb, 0.35-kb EcoRI-SalI fragment
frompVM2 (28).A3.6-kb EcoRIfragmentof Roussarcoma
virus (RSV) (non-src region) from pSRA2was used as an
avian retrovirus positivecontrol(12).
To prepare the oncogene blot, 50 ng of each oncogene
fragment was denatured and blotted onto a nitrocellulose
filter by using a slot blot manifold. This blot was then
hybridizedwith 32P-labeled RPL virus cDNA generatedby
reversetranscription of viralgenomic RNAharvestedfrom infected cell culture media. Medium was collected at 4-h
intervals,prespunto removecells and
debris,
andthenspun for90 minat40,000rpmtopelletthevirus. Theviralpelletsweresuspendedin 10 mMTris-HCl
(pH
7.4)-5mMEDTA,0.5%sodium
dodecyl
sulfate(SDS)-1
mgofproteinaseK per ml(1
ml of extraction buffer per25 mloforiginal
mediumcollected)andincubatedat37°Cfor 1 h. From 5to10 ,ug of viral RNA was usually recovered from 100 ml of culture
fluid.Theviral RNAswere thensubjectedtoreverse
tran-scription
in the presence of[a-32P]dCTP
to generate32P-labeled retroviruscDNAs(37).
cDNAlibraryandcloningof the viral oncogene.Total RNA
from RPL30 virus-transformed nonproducer cells (49) was
isolated, mRNAwas purified, andcDNAwas synthesized
by
standard procedures(37).
Internal EcoRI sites of thecDNA were methylated. The ends of the cDNA were
blunted with T4 DNApolymerase and then ligated to syn-thetic EcoRI linkers. The cDNAwas digested withEcoRI andrun on a0.8% agarosegel, andcDNAin thesizerange of 1to15 kbwasrecovered. This cDNAwasligatedtoAgtlO
vector arms to construct an RPL30 cDNA library. The
recombinant bacteriophage were packaged with Gigapack
Gold
Packaging
Extract(Stratagene,
La Jolla, Calif.) in accordance with the manufacturer's protocol and used to infectEscherichia
coliC600.Three DNA probes derived from the gag,pol, and env
regions
of the genome of cloned helpervirus UR2AVwereused to screen
105 plaques
from the RPL30cDNAlibrary(see Fig. 2A).
Positive clones weresubjected
to Southern blotanalysis
withprobes derived from otherregions
of the UR2AV genome. The sizes of the cDNA inserts from the clones and the distancebetween thehybridizing
probes onthe viral genome were compared. Those clones containing
cDNAinsertsthatwerelargerthan thedistancebetween the
hybridizing
probes were subcloned into the pBluescript phagemid(Stratagene)
by usingstandardtechniques(37)andsequenced.
DNA sequencing. Complete DNA sequences from both
strands of the clones identified above were determined by
the
dideoxy-chain
termination method(38)
by using theSequenase sequencing
system(United
StatesBiochemical,
Cleveland,
Ohio). Overlapping fragments
forsequencing
were generated
by
restrictiondigestion
andsubcloning.
DNA sequences and deduced amino acid sequences were used to search the GenBank data base by the FASTA program
(33).
Viral plasmid construction. Clones 8-1 and 3-1 (see Fig.
2B)
weredigestedandligatedinto anonoverlappingRPL30 clone(gp85-5'gp37-ryk-3'gp37-3'
long terminal repeat[LTR]).
Thisconstruct was thendigestedwithSail
(withingp85)
andEagI (within
the 3'polylinker)
andligatedtoSalI-and EagI-digested pSR-REP (9), which is an RSV-derived
retroviralvectorcontaining5'
LTR-gag-pol-gp85
(to the Sallsite).
Theresulting
plasmid construct, pRV-ryk,represents themolecularly
cloned RPL30virus. Helper virus plasmidpUR2AVhas been describedpreviously (49).
Transfection. Transfection of CEFwasperformed bythe
calcium phosphate method. Briefly, 4 to 10 ,ug of
plasmid
DNA per 60-mm-diameter dish was digested with EagI
(pRV-ryk)orSacI(pUR2AV), phenol-chloroformextracted,
precipitated with ethanol, and dissolved in 0.5 ml of 1 mM Tris-HCl(pH 8.0)-0.1 mM EDTA. A mixture of 10 ,ug of calf
thymus DNA and 55
p,l
of 2.5 M CaCl2was added to theplasmid DNA. A mixture of 0.5 ml of 50 mM HEPES (N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid), 280 mM NaCl (pH 7.1), and 5 ,ul of 0.15 M Na2HPO4wasthen added dropwise with mild vortexing. This solution was
allowed to stand at roomtemperature for 30min, andthen thefineprecipitatewasaddedto7 x
105
secondary CEF per 60-mm-diameter dish and left for 8 hat37°C. After thattime,the mediumwasreplaced with fresh medium.
Transformation, colony formation, and animal
tumorige-nicity
assays.Transfected, infected, and mock-treated(sameprocedure except that no plasmid was added) cells were
maintained in medium containing 0.375% agar at 37°C. Transformationwasassessedby focus formation. To assay anchorage-independent growth, cells were trypsinized and replated in 10-cm-diameter dishes in 10 ml of medium containing 0.4% agaron alayer of 15 ml of the same medium containing 0.7% agar. Theplates were incubated at 40°C.
Tumorigenicity was assayed by injecting 0.1 ml of virus intothewing webs of newborn (7 days posthatching) White Leghorn chickens (SPAFAS, Inc.).
Generation of specific antiserum. The C-terminal unique region of v-Ryk was expressed in E. coli BL21 (DE3)pLysS
by using the T7 RNA polymerase expression system (36,41).
An 897-bp Sau3AI fragment containing the region from amino acid 463tothe C terminuswascloned intoa compat-ible BamHI site inthe linker region of the pET-3c vector. Plasmid-bearing bacteria were induced with
isopropyl-P-D-thiogalactopyranoside for 2 h, spun out, and lysed on ice by sonication in RIPA buffercontaining 1 mM phenylmethyl-sulfonyl fluoride. Proteins in the lysate were then separated by SDS-12% polyacrylamide gel electrophoresis (PAGE). The v-Ryk protein band was excised from the Coomassie blue-stained gel, washed with 10% methanol, dried, and homogenized in phosphate-buffered saline (PBS). Approxi-mately 100p,gofv-Ryk proteinwasemulsifiedwith Freund's complete adjuvant and usedtoimmunizerabbits,and 50 ,ug ofproteinwasused for boosterinjections. Preimmune serum
wascollected for control experiments.
Thespecificity of the antiserum was tested by an antigen
competition assay.v-ryk-transformed CEFwere labeled in
vivo with
32Pi
for 8 h(19) and thenlysed in RIPA buffer (seebelow). Preimmune or immune serum(20 ,ul)was
preincu-bated in 120
pl
of RIPA buffer withorwithout 144p,gof the immune antigenorcontrolantigen(vector-bearingbacteriallysates) for 3 hat 4°C and thenused to immunoprecipitate 14.4 p,g of32P-labeled cell lysate (see below).
Immunopre-cipitated proteinswere analyzed bySDS-7.5% PAGE.
Immunoprecipitation and in vitro kinase assay. Cell
cul-tures werewashed with ice-coldTris-glubuffer(25mM Tris, 137 mMNaCl,5 mMKCl, 0.7 mMNa2HPO4, 0.1%glucose, pH 7.4), and the cells were scraped off the plate, quickly
centrifuged, and frozen in a dry ice-ethanol bath. Frozen
pellets were suspended in RIPA buffer (10 mM Tris-HCl,
(pH 7.4), 5 mM EDTA, 150 mM NaCl, 10% glycerol, 1%
TritonX-100, 1%sodiumdeoxycholate, 1%Trasylol, 1 mM
phenylmethylsulfonyl fluoride,1 mMsodiumorthovanadate, 0.1 mMsodiummolybdate),incubatedonice for 20 min with
periodicshaking, and spun inamicrocentrifugeat4°C, and the supernatants were recovered. Protein concentrations
wereassayed witha ProteinAssay Kit(Bio-Rad Laborato-J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
THE v-ryk KINASE ONCOGENE
ries, Richmond, Calif.), and equal amounts of proteins were used in the subsequent assays. For immunoprecipitation, antibodies were added to the above-described cell lysates and incubated on a rotating wheel at 4°C for 1 h and then protein A-Sepharose (Pharmacia, Piscataway, N.J.) was added and the incubation was continued for another 30min. Theimmunoprecipitates were washed three times with RIPA buffercontaining 300 mM NaCl and twice with RIPA buffer containing 10 mM NaCl and then used directly either for SDS-PAGE analysis or for in vitro kinase assays.
For invitro kinase assays, the immunoprecipitates were washed twice with the kinase reaction buffer (see below) instead of RIPA buffer containing 10 mM NaCl, and then each was suspended in 30 ,ul of kinase reaction buffer (50 mMHEPESbuffer[pH7.5],10 mMMnCl2).Atotal of 5
pCi
of [-y-32P]ATPwas added to each sample, and it was
incu-batedat30°C for 10 min. Kinase reactions were terminated
by washingthe samplestwice with RIPA buffer containing
10mMNaCl and then subjected to SDS-10% PAGE. Rain-bow markers(14.3 to 200-kDa range; Amersham, Arlington
Heights,Ill.) were used as molecular weight standards in all protein gels. The gels were dried and exposed to X-ray film
at -70°Cfor 24 h withan intensifyingscreen.
Westernblotting(immunoblotting).Equalamountsof pro-tein from cell lysates (see above) were subjected to
SDS-10% PAGE and then electrophoretically transferred to a
polyvinylidene difluoride nylon filter (Millipore Corp.,
Bed-ford, Mass.)withtransfer buffer(2.5mMTris-HCl[pH 8.3],
9.2 mM glycine, 10% methanol). The filters were then probed withanti-Ryk serum orantiphosphotyrosine (PTyr) polyclonal antibodies, followed by incubation with
1"I-labeledproteinA(Amersham)asdescribedpreviously(22).
Autoradiographywasperformedasdescribed above. Partial protease mapping with Staphylococcus aureus V8 protease. 32P-labeled protein bands were excised from in vitro kinase assaygel andsubjectedtopartial digestion with S. aureusV8 protease asdescribedpreviously (7).
Pulse-chaseanalysis.CEFtransformed by the clonedv-ryk gene were labeled with
[35S]methionine
(110 ,uCi;Amer-sham)for 1 h(pulse)in MEM(without methionine)medium
(GIBCO, Gaithersburg, Md.) after starvation in the same
medium for 2 h. The cellswerethenwashed,incubated with
completemedium,andchased for0, 15,or30 minor1, 2, 4, or 8 h. At those timepoints, cellswere lysed as described
above,
immunoprecipitated
withanti-Ryk antibodies,
andanalyzed by SDS-10% PAGE.
Cell fractionation. Cells were swollen in hypotonic lysis
buffer (20 mM borate, 0.2 mM EDTA [pH
10.1]) (0.45
ml/10-cm-diameterdish) for 10min,collected,and
homoge-nized in a Dounce homogenizer (15 strokes), and then 8 volumes ofhypertonicsolution(0.5Mborate[pH
10.21)
wasadded. Thelysatewas spunat500 xg and4°Cfor 10min,
the supernatant was collected and spun at
20,000
x gand4°Cfor 30 min, and the supernatant
(soluble fraction)
wascollected. The
pellets
were dissolved in PBS andlayered
over sucrosegradients
(40
and20%[wt/vol]
sucroseinPBS)
and then spun at 100,000 xg and4°C for 1 h. The
plasma
membrane-enriched fractionwas
collected,
diluted inPBS,
and spunat 100,000 xgand4°Cfor 30min, and the
pellets
werecollected asthe membrane fraction. Thetwofractions
werethensubjectedtobothWestern
blotting
and the in vitro kinase assay.Nucleotide sequence accession number. The sequences
re-portedhere have been
deposited
in theGenBank data baseunderaccessionno. M92847.
RESULTS
Genetic analysis of threeRPLoncogenicretroviruses.Three ofthe RPL series ofacuteoncogenic retroviruses, RPL25,
RPL28,and RPL30,werefirst isolated from chickentumors
in farmflocksbyBurmester's groupattheRegionalPoultry Research Laboratory (now named the Avian Disease and
OncologyLaboratory)in EastLansing, Mich.,in the 1950's and 1960's(21).These RPL viruseswereshowntobe ableto
induce chickenerythroblastosis, fibrosarcomas, endothelio-mas, viscerallymphomatosis, nephroblastoma,
osteopetro-sis, and hemorrhages. The nature of the lesionswas influ-encedbythe virus dose inoculated, the ageat inoculation,
and therouteof inoculation(21).Since these viruses hadnot
previously been genetically characterized, we decided to
analyzethe transforminggenes presentin these three RPL
viruses.Injectionof the viruses into newly hatched chickens resulted in induction ofsarcomasfrom whichacutely
trans-forming avianviruseswere recovered. These RPL viruses
werealso showntobeabletotransform CEF in culture and
wereused for furtheranalysis. First,wedetermined whether
theyencode oncogenes that havealreadybeen identifiedin other oncogenic retroviruses. A screening method which allowsrapid comparison ofnewretroviral genes toknown viral oncogenes was developed. Oncogene-specific DNA
fragmentswereisolated fromapanelof 19plasmids contain-ingknownviral oncogenes. Each DNAprobewascarefully
chosensothatit containednovirus-derived sequences that could hybridize nonspecifically with the retroviruses to be tested. These probes were bound to nitrocellulose
filters,
and this oncogeneblotwasthen
hybridized
under conditionsof moderate
stringency
with 32P-labeled cDNA madeby
reversetranscriptionof different RPL virus
genomic
RNAs. These viral RNAs were obtained from virionparticles
re-leased from CEF transformed with the RPL viruses. If any of these RPL viruses contained a known oncogene, its cDNAwould beexpectedtohybridize
tothecorresponding
probeonthe oncogeneblot. The results of the
screening
are shown inFig.
1. Allthree RPLvirus cDNAs,asexpected,
hybridized to thepositive control, a DNAfragment of the
avian retrovirus structural gene. RPL25 and RPL28 also
hybridizedtothe slotscorrespondingtotheerbBand
erbA-erbB probes. Southern
blotting
of therestriction-digested
original oncogene-containing
plasmids
with the RPL25 andRPL28 cDNAs demonstrated that the
hybridization
wasspecific
toerbB sequences(data
notshown).
This kind of Southernblotting analysis
also indicated that the low-levelhybridization to the src and
myb
probes observed withRPL25 and RPL30was due tocontamination of the probe
fragmentwith non-oncogene-derived viral sequences
(data
not
shown).
Furthermore, Northern(RNA)
blots of RNAs from cells infected with RPL25 and RPL28hybridizedwithan erbB probe,
showing
specific hybridization
bands notpresent in RNA from uninfected CEF
(data
notshown).
These results demonstrated that the RPL25 and RPL28 viruses contain the erbB oncogene,similartoavian
erythro-blastosis virus
(AEV),
whose cellular counterpart is theepidermal growth factor receptor
(15, 51).
Moreinterest-ingly,
nospecific hybridization
of RPL30 to any of the oncogeneprobes
wasdetected,
suggesting
that the RPL30virus containsanovel oncogene.Likealmost all of theacute
oncogenic retroviruses,RPL30 is alsoa
replication-defective
virus and needs a helper virus to produce infectious viral progeny
(data
notshown).
The size of the RPL30 viral genome, 9
kb,
isunusually
largeamongaviansarcomaviruses and is exceeded
only by
on November 9, 2019 by guest
http://jvi.asm.org/
5978 JIA ET AL.
LTR gag pol env LTR
I a I I I UR2AV
gag pol env
B
LTR gag pol
clones:
RPL30
*: i
FIG. 1. Hybridization of three RPL virus cDNAs to oncogene
probes.DNAprobeswereisolatedfrom 19 known viraloncogenes
and boundtonitrocellulosefiltersasdiagrammedin the lowerright panel (seethetextforprecise probeidentities).32P-labeled cDNAs preparedfromRPL25, RPL28,andRPL30virionswerehybridized
tofiltersasdescribed in thetext. ALV,avian leukosis virus.
that of RSV(9.31 kb),which contains thesrc oncogene in additionto all of the viralgenes(48). To determine what,if
any, viral genetic information had been deleted from the
RPL30 virusduringtheprocessofoncogenetransduction,as seen in almost all of the oncogenic retroviruses character-ized so far, nine probes spanning the whole retroviral
genomewerepreparedfrom cloned UR2AVDNA and then hybridizedtothe 9-kbRPL30genome. The resultsshowed, surprisingly, that all nine probes hybridized to the RPL30
genome(datanotshown).Thissuggestedthat eithernoviral sequencedeletion occurredduringtransduction of the
onco-geneoronlyasmall deletion occurred.Inotherexperiments, immunoprecipitation of lysates from RPL30-infected CEF with tumor-bearing rabbit serum showed elevated invitro
tyrosine kinase activity relative tolysates from uninfected CEF (data not shown). Tumor-bearing rabbit serum was
raised in rabbits bearing RSV-induced tumors and
recog-nizes avian retroviral structural proteins in addition to
pp60-s'c (4).Thispreliminarycharacterization of theRPL30
virus strongly suggested that RPL30 contains a novel ty-rosine kinaseoncogene.Therefore,wedecidedtoclone this
oncogenemolecularly.
CloningofthetransforminggeneintheRPL3O virus.Since
wedidnothaveaspecific probefor thetransforminggenein theRPL30virusand didnotknow thepreciselocation of the
geneinthe RPL30genome,wedecidedtousethreeprobes
derived from UR2AVgag,pol, andenvtoscreen acDNA
libraryconstructed from RPL30-infected nonproducer CEF (Fig. 2A).The rationale of this strategywastoisolateRPL30
env env LTR
iI
-+onc
1.7kb
2.2 kb
2.5kb
RPL30
3-1
8-1
[image:4.612.62.299.72.356.2]2-4
FIG. 2. Cloningof thetransforminggenein theRPL30virus.(A) Diagram of the UR2AVgenome,ahelpervirusrequiredforRPL30 replication.gag, pol, and env denote the viral structural genes.
Three DNA probes usedintheinitialscreeningfor theoncogenein RPL30arerepresented bythick bars beneath the positionsin the viral genome fromwhich theywere derived. (B) Cloning of the oncogenein the RPL30virus. The three thick bars represent the threeoncogene-containingclones thatwereisolated from thelibrary andsequenced.Their sizesareindicated beneath thebars,and their
names areindicatedto theright of thebars. Above the bars isa
diagram showingthe location of thetransforminggenein theRPL30 viralgenome; this informationwas deduced from thecloningand sequencinganalyses.
viralgene-oncogenefusionfragments. The fusionfragments
could then be identified asclones containinginserts larger
than thoseexpectedfromprototypic retrovirusse uences.
Six positive clones were isolated from 3 x 10 plaques
from the RPL30 cDNA library with the env probe, eight
were isolated with thepol probe, and none were isolated
from 4 x
104
plaques with thegag probe. The inserts inpositive clones were excised from the phage vectors and
subjectedtoSouthern blotanalysis. TheSouthern blotswere
firsthybridizedwiththeprobesused in thelibrary screening toconfirm theauthenticityof theclones and then
rehybrid-ized withprobesfromregionsin theUR2AVgenome
adja-cent tothose from which thescreening probeswerederived.
The insert sizewascompared withthe distance betweenthe
hybridizing probes on the UR2AV genome to determine whether the size was consistent with or larger than that
distance. Three clones originally detected with the 3' env
probe caught our attention. These three clones were later
foundtobe identical to each other and arerepresented by
clone 3-1 inFig.2B.The inserts fromthese cloneswere 1.7
kblong,andtheyalsohybridizedwithanLTRprobe butnot
witha5'envprobe(datanotshown). However, the distance
between the5'end ofthe3'envprobe (0.8 kb) and the 3' end
of the LTRprobe (0.6 kb)isonly1.4kb. Thenegative results
obtained with the 5' env probe suggested that these 1.7-kb
clones containsequencesin additiontotheviralgenes;the
extrasequences werepotentiallythose of the RPL30
onco-gene.Theremainingcloneswerenotfoundtocontainextra sequences. By subcloning and sequencing, clone 3-1 was
foundtocontaina1,665-bp insert. Atthe3' end of thisinsert
is the viral3'envgp37gene,followedby the LTRsequence.
At the5'end,however,theinsert contains 818 bp ofnonviral
sequenceswhicharehomologous tothecatalyticdomain of protein tyrosine kinases(Fig. 2B). To isolatethefull-length
RPL25 RPL28 A
probes:
J.VIROL.
.1. i. ;..
;I-.f:`
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.321.560.75.240.2]cDNA for this putative transforminggene, the kinase frag-ment from clone 3-1 was used as a probe to rescreen the
RPL30 cDNA library. Numerous positive clones were
iso-lated, and twoof them (clones 8-1 and 2-4), containing the
longest inserts (2.2 and 2.5 kb, respectively), were chosen
forsubcloning and sequencing (Fig. 2B). Clone8-1contains 234 bp of the viralenvgp85 gene atthe 5' end, followed by 538 bpofthegp37gene,1,390 bp of nonviralsequences,and
10 bp of the gp37gene. Clone2-4 started with 378bp of the envgp37geneatits5' end, followed by1,390 bp of nonviral
sequencesand77 bp of 3' gp37, andended with 298 bp of the
LTRsequence.Therefore, both clones 8-1 and 2-4appeared
to contain the full-length putative oncogene. Since this
1.39-kb nonviral sequence has a protein tyrosine kinase
domain,wenamed thisputative viraloncogenev-ryk, for the
RPL30virustyrosine kinase.
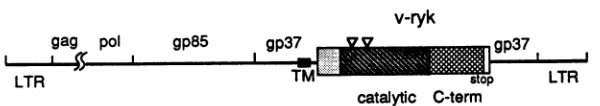
Sequenceanalysisofv-ryk.Thesequenceofthe3'region of theRPL30 virus, including the v-ryk oncogene, is shown in
Fig. 3A. The v-rykgenewasinserted into the viral envelope
protein gp37 coding region within theenvgene. Itcontained an open reading frame of 442 amino acids, followed by a
62-bp3' untranslated region containinganAATAAA
polya-denylation signal. This open reading frame was fused in
framewith theupstreamviral gp37gene, which contains a
transmembrane domain. Thus, the putativeoncogenein the
RPL30 virus encodes a 621-amino-acid gp37-Ryk fusion
proteinwith a predicted molecular mass of 69 kDa termed
P69gp37-,yk. Interestingly, in contrast to almost all acute oncogenic retroviruses, v-rykwasinserted into the 3' region
of theviral envelope protein gp37 genewithout deletion of
anyviralsequence. The DNA sequences flanking the v-ryk oncogene in the RPL30 viral genome match perfectly the
continuous sequenceintheenvgene of other similar avian retroviruses, likeRSV(39) and UR2 (32).
InthesequenceC terminaltothetransmembrane domain ofgp37,p69WI37-ik has four amino acids fromgp37, followed
by thev-Ryk sequence, whichconsists of a42-amino-acid
N-terminal domain,a277-amino-acid protein tyrosinekinase
catalytic domain, anda 123-amino-acid C-terminal domain. The structure of v-Ryk is diagrammed in Fig. 3B. The boundaries ofthe kinase catalytic domain were defined as
describedby Hanks etal. (25). Itcontains the ATP-binding
siteconsensus sequenceG-X-G-X-X-G andalysine residue
atposition 334 (Fig. 3A), which is invariantamongall protein
kinases (24). v-Ryk contains all 11 of the subdomains
con-served in all of the known protein kinases (24). Further analysis revealed that the sequenceofv-Ryk more closely
resemblesthatoftyrosine kinases rather thanthat of serine-threoninekinases.Namely, 38 of40 highly conserved amino acidsoverthewholecatalytic domainof 42 tyrosine kinases
(24) were present in v-Ryk. In the regions thought to be
importantindetermining the substratespecificity ofakinase
(20, 25), D-L-A-A-R-N in subdomainVIb ofreceptor-type tyrosinekinaseswasconserved in v-Ryk and
P-I/V-KIR-W-T/M-A-P-Einsubdomain VIII ofmosttyrosine kinaseswas
P-V-K-W-i-A-i-Einv-Ryk; both regionswerequitedifferent
from those characteristic of serine-threonine kinases. This
stronglysuggested that v-Ryk isa tyrosinekinase. Tyr-469
corresponds to the conserved autophosphorylation site in otherkinases.Furthermore, intheposition correspondingto
the kinase insert of the platelet-derived growth factor and
colony-stimulating factor 1receptors (25),v-Ryk contains a
seven-amino-acid insert withthesequenceLEMAPQF. The
functional significance ofthis insert is not known. Beside this short kinase insert, v-Ryk contains an additional
six-amino-acidinsert,LSSQQI, betweenkinasedomainsIVand
V. The full-length v-Ryk sequence was used to search for
protein sequence homology in GenBank. The region of
v-Rykhomologous to otherproteins is limited to the kinase catalytic domain(Fig. 4). The highest homologies found in the kinase domain were to theaxl-UFO oncogene product (67.3% amino acididentity), the ark gene product (67.3%),
thev-sea oncogene product (47.4%), the humanmet
onco-geneproduct (43.2%), the human insulin receptor (41.4%),
the bovineinsulin-like growthfactor 1 receptor(41.1%),and the humanretoncogeneproduct(40.0%).axl(32a)andUFO (26a) are the same gene, isolated from human myeloid
leukemia cells, which encodes a putative tyrosine kinase receptor. ark isa murine putative tyrosinekinase receptor gene(35a). The v-sea oncogene is the transforminggene of avian erythroblastosis retrovirus S13, and its cellular ho-molog is a receptor-type tyrosine kinase (40). Since these
highlyhomologous kinasesarereceptor-typemolecules and
v-Ryk is only 36.1% homologous to c-Src in the kinase
catalyticregion, c-Rykismorelikelyareceptor-typethana
non-receptor-type tyrosine kinase. Another interesting
fea-tureofv-Ryk is that the 123 amino acids C terminal to the kinase domain show no significant homology to any other
protein in GenBank. Since this domain contains three
ty-rosine residues, theymaybepotential autophosphorylation
sites and thus may havesomeregulatoryrole(s),asproposed
fortyrosine residues in theC-terminal region ofthe
epider-malgrowth factor receptor (45)orthe Neuoncoprotein (1).
Biologicalactivityofv-ryk.The RPL30 virus caused trans-formation of CEF 10 to 14days after infection. The
trans-formed CEFexhibited an elongated and fusiform
morphol-ogyand grewto ahighercell density than normal CEF(Fig.
5D).
These transformed CEF formed colonies, but control cells did not (Fig. 6D), reflecting anchorage-independentgrowth. The RPL30 virus induced chicken sarcomas 5 to 6 weeks after injection into the wing webs of 1-week-old chickens(datanotshown).Todetermine whether thecloned v-ryk gene is fully responsible for thetransformingability of the RPL30 virus, we tested the biological activity of this
putative oncogene.To dothis, v-rykwascloned into
retro-virus vector pSR-REP to construct retroretro-virus pRV-ryk, which contains the 5' LTR, gag, pol, and gp85 from RSV
andgp37-rykand the 3' LTR from RPL30. When pRV-ryk
wascotransfected into CEF with pUR2AV, the CEF became transformed and formed focimorphologically indistinguish-able from those ofRPL30-transformed cells after 10 to 14
days (Fig.
SC),
while neither pUR2AV itself nor mocktransfection caused any transformation (Fig. 5A and B). Whenthesetransformed cellswereplatedon agar,colonies verysimilar to those induced by the original RPL30 virus formed (Fig. 6C). Virus collected from these pRV-ryk-transformed cells induced sarcomas in chickens within 6 weeks, and histological examination of the sarcomas re-vealed that theywere fibrosarcomas and
rhabdomyosarco-mas(datanotshown).Therefore, the cloned v-ryk gene can fully reproduce the transforming capacity of the RPL30 virus.
Biochemical analysis of the v-Ryk oncoprotein. To avoid cross-reaction with othertyrosine kinases, the unique region C terminal to the kinase catalytic domain of v-Ryk was
chosen as an immunogen to generate specific anti-v-Ryk antibodies in rabbits (see Materials and Methods). The
specificityof theserum wastested inanantigen competition assay. Figure 7 shows that in lanes 3, 5, and 7, cold
immunogencompeted successfullyto remove aband witha
molecularmassof 70kDaimmunoprecipitated from in vivo
phosphate-labeled RPL30-transformed cells by sera from
VOL. 66,1992
on November 9, 2019 by guest
http://jvi.asm.org/
5980 JIA ET AL.
A
TSCCCO AnCSCCOGCGA TOI OTC GAC LC ACACMaAARTTTCAIC AMA CCA AAM 60
phe pro11l loU proglyva1 trpv-1*ap a.rthrginglymenph. thrlyeprolye 20
gps
O00 CTA CCI CCCOCA ATTTTCCTC ATrTOC OATCOC WATOGC> BOAATTCCTAOT120
&laleu pro pro ciaIleph. ileeye giy*aprg &IatrpginglyIlepro*sr 40
COT CCA 0TA 0C GMOC CCCTOC TATCTA WOA AAACT? ACCATO TTAOA CCC AAC CATACA 160
argproval glyglyprocya tyrloU giylye leo thrmetleu*lapro&&n his thr60
CATAT? CAC AAM ATACTTOCTAT TCC TCO CAA MCA OC AMA CAM TT CIUA COT? 240
aspilehis lys ileleu atlasnner serg1n thrglyva1 erghi. ph. ergserv-1 60
qP37->
TCA CATCT? GAT CA? ACA TOC TCA GAC GAA CAGCT? TG OCTCCTACA BCA AA ATC 300
*-S hi. loU asp aspthreye *apglU va1 ginloU tapglyprothraia argile 100 TT? BATCTATC TTACC CCA GOO OTAWCAOCCCC CAAOC CTA AX GM ATCCM AMA 360
ph.&lamer ileleoU al proglyva1 &ia*- &lagin &laloU erggln Ileglo erg 120
CTACC TOTTOOTCTC?AAA CAM OCTAAC TT?ACA MATCACTC CT?Go CGMTTATT 420
loU clacys trpserv-1 lys gin cla can loU thr thr merleo leo gly asploU loU 140
CAT CGMCC MA ACT AT? CGA CAT0CC OT? CTC CAM AAC CCA C00 AT? GMTT? TT? 480
cap cap vclthr serile crghis lac vcl loU gin canarg &ia ciaile*apph. leu 160 CT? CTA OCTCAM CM? TOTAACCACT? GC AT? TOTTOOC TTC AAM CT? AMT540
leu leou hi. glyhis glycye lys capile&lagSlyntCy. eyeph. can lou *er180
GAM CACAOCGMCT ATA CAM AAC TTC?C CAM CTAAT? AMU GMCAM OTC AAM AMGATC .00
caphi. oar glU c&ailegin lys lys ph. gin loU netlye glu his vl can lysllo 200
0GC GMCAGCCAC CCA ATCCG ACTTOO CTCQGAGMA TTA TC CB WAATS C0T GM660
giyval cap ser capproileglysertrp1iU argglyleo ph. glyglyil* glyglU 220
TOGGC ATA CAC TTACTAAAMCGA CT CTT TTS CT? OTACT? ATC TTATT CTA 0TA 720
tap ilehisloU ioU lyeglyloU leo iougiyleo va1 vai ile lou loU loU vcl240 GTC TOC CT? CCT TOC CTT TT? CAM TTS OTATCT A WCCGA AAC ATCacTACT ACA760
vaicys ioU proCye ioU leo ginph. vai serser aerthrSrg lye netthr thrthr260
...A... Iv-ryk-2,
OTe OTAAAM TacACA CCM AA AAM TCCTAC TOC COOAMA 0CCOTCGMCT?ACATTO 640 vcl val&an tyrthr cialys lye eertyr eye crgarg*a1 v-1 giu loU thr leo gly 280
AMCC? GGACTC AGC AGC GAM CTC CAM CAM AAC CT? C GM SCTOT AT? AMA AAM 00
earlou glyvcl aermerglU loU gin gin lys loU gin capv.1 va1 ileaeparg cen 300
kinasoI -->
GCCCT?AGO CT? AAC GTC CT?CGA GAM CGM TTC TCA ATC GAM GA C0T960
alaloU aer loUglylye vcllouglygluglygluph. glyser va1met glo giyarg320 CTCACC CMGCOCAA GGC ACCCCA CaM AAC GT OCTC? AAC ACCAT? AAM TTG OATAAC 1020 loU ner gin proglu glythrprogin lys v.1 &lav.1 lys thrsetlys lOU cap can 341
TT? TCCCAT AMA GAM ATACMCM TTC CTC AMTGMWcA BATOC ATAAAC Gac TT GM1060
ph.*erhi. argglo ileglugluph.ahleo glo cilac&a eys1il lya capph.*ap360
Cac COC AATCST ATC AAM CTC CTAGOT GT? TOC ATCCM CT?AOCTCTCAM CAM ATCCCC 1140
his pro &anva1 ilelye loU ioU glyvc& cys ileglu loU aer aergin ginilepro360
AAM CCCAT?CT CTT CTCCCATTC AT? AAM TATCT CMCTC CACACCT CT? CT? COC1200 lys pro netvclvcl loU proph. met lys tyrgly cap1lU hi. aerph. loU loU crg400
TCCCOGCTCCA0 AT? oCC CCC CAM TTC OT CCCCT? CAM ATC CT? CT? AM TTCAT? GT? 1260
aararg loUglU met*iaproginph. vclproloU gin matleu l-u lyeph. met vcl420 GAM AT? 0CCCT? Oa ATC CGM TAC CTC AOC AM CO0 CA TmCTT CAC AO OAT TT? GOG 1320
cap 1ie&aloU glynet glu tyr loU serser crggin ph. loU hi. crg caploU &la440
OCTC00 AAC TOC AT? TTACC0 GA? CACAT? ACG GTGTOT GT? WCAGM TT? c0 CT? TCC1360 ilaarg cancys met leo crg capaspmetthr vcl eye va1&ca capph. glylou ser460
AM AM ATC TA AOC GO GMATM TACTCOT CM CO ATACCAA AT CCAOT? AAM 1440 lys lye il tyr aergly captyr tyr crg gingly arg1ie cal lysmetprovcl lye 460
TOO ATT ATAGMTCCCTC CT MAC COTGT TAC accACCAM ATOATT TOO ¢cA1500
trp ile cal ili glu serlou cla cspargv1ltyrthrthrlyeser cspvaltrp cla 500
TT 00c an ACCAT? TOOGMa ATA 0CO ACC AMA GO AT? ACTCCC TMAC cCA 0o OTCCAM 1560 ph. glyvalthrmettapglu iliea thr erggly met thrprotyrproglyvalgin 620
AACCACGMAT? TATGMTATCTATTC Cac GM CAM COOCTCAAA AACCCT AmAACTOC 1620
canhi. glu iletyrglutyr leu ph. hi. glygln cap leulye lyeproglu can eye 540
T?ACATGAM CT? TAC OATATCAT? TCC TCCTOC TOOAo OGCTGM OCT OC GACCOAC00 1660
lou capglu l-o tyr capi1e met aer eer cye trparg cal glupro ila cap crgpro560 ACC TT TCA CAGCT? aAAOT CATCT? GMA AacCT? TT? GA aCc CTTCCT 0CCC00 AG 1740
thrph. ,erginleu lye vcl his leu glu lysil- lou glunor leuproalaproarg560
IC-tere ->
TCCAAC CACG? ATc TA GTC AACACC MCCCTOCeaGM GA aocCCC GACTCCAce 1600
gly merlye capval iletyrvcl canthr*ar leuproglu gluser pro cap eorthr 600 CMCGMTT? 0C0 TT? CAT TCOG GT ATCCCC CAA GMOG TCTGMTT? CGCCCCGMG GM 1660
gln cap l-ugly louasp serval11. pro gln*1- capsor caplou*approgly cap620
ATT OCTGMCCCTOCTOC ?CC CAC acM AAM GCAGCGCT? OT?COAOT? GMAT? CACGAM 1920
1lo cal gluproeys cye serhisthr lys c,laal leuv.1 cal v.l aspilohis cap640
T MGTAC GT? CTTCAAAI GMA 0CaO CCCaAGMCAT CTTACVT? COA1660
glyglyser crgtyrvcl leugluserglu glyser prothrglu cap cal tyrvclpro660
CAGCT? CCC CAC GM 0O0 TCO TooACC GM 0CCaMC ACC TTO CCTOT? Aac TCG2040
gln leuprohisglugly ser cal trp thrglu cia aerthr leuprovcl gly*erser680
CTTocA OCT Ca CTC CCATOT CT CAT TOC CTC cMcM TCC GA CTGCTOTOA 2100 lou cia cal gln leupro eys cla cspglycysleuglu cap*-r glu alaleu lou '" 699 AT? CM CCT C CAM aoCAOC GT? TOAATC AMC ACA CM aA TTTTATTT AA AAaCAM 2160
TCT TATTA TTCACTCG CTA TCO COAGOA ATACAM GMATT Aea AGA TAM TAM GCA2220
^3'env-->
CCC TAAAM aMC acW ATAGOCCBO TTCCTCTAM TCO TOTCAT ACCTO TT? CGM C0 TAM 2280
TCO A?CGOTTAM CATACM GCMTST AAM AGOTCO CT? AWT AWTOCCGACA AMTT? AM2340
CTACAM TAM GCC AAC CT TGACCO OCA T?S GOA TCA AQ AAM CT? CT? MG T?T AMCC0 2400
TT? TT?.CT? COC OAT CTA C GOTAGo TTTACM COT AAC TOG 0o CCaTQA TAT2460
OTATAM CCA ACMG 0o CT? CO TTSS TAC CC ATMaGCeC CTC A" AMTAM TOG2520
AM COc TTT TOC ATA AGO AATOT AWT CTT An CAM aM TA AMTOT ATA2580
^ U3 -->
TTaCOA MAT AAM COAATC 0CC TOATOCACC aAA AA COTATTATATOATCC CATTooTOO 2640 TOAAGOACC CACCT? AMG OCA TAT 00c CCT TMA TMA AACTCT CTG TCCTSGC0TCAT TCC2700
T?C CTCAT?C BATCA TCT acM COG CAM AMT ATCAT? OGATa CAM GA BOC aMc ATT CAT2760
COT 0GCCOATOC TCA TTG CT? TCA CTAACG aWT TAT 0TA ACC CAT GMTOTACT TaA OCT 2620
TOT AW TOC TAM CAA TAA AWI 0CCATT TTA CCT CCC ACC ACaTTC OT? TOCACC TOO OTT 2660
R--> A US-->
CAT 0O CG0 aCCOTC OATTCCTCAC0G CTACQAOACCT GA TOA aoCaMA aGo CTT CAC 2940
COa 0C0 CAT?CT AT?ACW CCA TOATCAAAA CT? AAA acc TACACTAT TT CCCAACCOO3000 cell->
CMOCAAT?
B
v-ryk
gag pol gp85
I l M
LTR
gpif r
~~~~~~-
xx 0 #4TMso,
17
LTR
catalytic
C-termFIG. 3. cDNAsequenceandstructureof the 3' portionof theRPL30virus. (A)cDNAsequence and translatedprotein sequence ofthe
3'portionofRPL30,includingthe v-tykoncogene. The starting site foreachgene regionisindicated byasinglecaretunderneath thefirst
nucleotidefollowedby the name anddirection of thatgene. Aseries ofcaretsunderlinesthetransmembrane domainof gp37, an asterisk
marks theATP-bindingsiteatLys-334in thekinase catalytic domain of v-Ryk, andc-termdenotes theC-terminaldomain of v-Ryk (seepanel
B).Aseries of threeasterisks indicatesthestopcodon for theproteinencoded bythisgene,andthe AATAAA polyadenylation signalsite isunderlined.The numbersfor nucleotidesand amino acidsaretothe right of the corresponding lines. (B) Genomicstructureof theRPL30
virus. Theviralgene names are indicatedabove their locations, and the coding regions for thetransmembrane domain, kinase catalytic domain,and C-terminal domain(C-term)of v-Ryk,aswellasthestopcodon,arelabeledunderneath. Thetrianglesdenote the kinaseinserts. Thisdiagramis drawn approximatelytoscale.
J.VIROL.
04r%27 I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.167.467.576.629.2]v-RYK NALSLGKVLGEGEFGSVMUGRLSQPEG--TPQKVAVETMKLDNFSHRIIEEFLSEAACIKDrDHPN
AXL/UFO HKVALGKTLGEGEFGAVKEGQLNQDDS---ILKVAVTMKIAICTRSELEDFLSEAVCMKEFDBPN
ARK HKVALGKTLGEGEFGAVKEGQLNQDDS---ILKVAVKTKIVIC;TRSELEDFLSEAVCMXEFDHPN
v-SEA LITHRSRVIGRGHFGSVYHGTYMDPLL--GNLHCAVESLHRIT-DLEEVEEFLREGILMKGFHHPQ
HIR EKITLLRELGQGSFGMVYZGNARDIIKGEAETRVAVXTVNESA-SLRBRIEFLNEASVMKGFTCHH
MET LIVHFNEVIGRGHFGCVYHGTLLDNDG--KRIHCAVKSLNRIT-DIGBVSQFLTEGIIIMKDFSHPN
IGF1R EKITMSRELGQGSFGMVYEGVAKGVVKDEPETRVAIKTVNEAA-SMRERIEFLNEASVMKEFNCHH
RET KNLVLGKTLGZGZFGKVVEATAFHLKGRAGYTTVAVKMLKENA-SPSELRDLLSEFNVLKQVNHPH
c-SRC ESLRLEVKLGQGCFQEVWMGTWNG---TTRVAIETLEPGTM--SPEAFLQEAQVMEKLRHEK
v-RYK VIELLGVCIELSSQQI-PKPMVVLPFXKYGDLHSFLLRSRLEMAPQF---VP AXL/UFOVMRLIGVCFQGSERESFPAPVVILPFKHGDLEESFLLYSRLGDQPVYL---PT ARE VMRLIGVCFQGSDREGFPEPVVILPFKKHGDLSLHLYSRLGDQPVFL---PT
v-SEA VLSLLGVCLPRH----GLPLVVLPYNRHGDLRHFVRAQERS---PT HIR VVRLGVVSEG---QPTLVVMELKAHGDLKSYLRSLRPEAENNPGRPP---PT MET VLSLLGICLRSE----GSPLVVLPYKHODLRNFIRNETHN---PT IGF1R VVRLLGWSQG---QPTLVIMELKTRGDLKSYLRSLRPEMENNPVLAP---PS RET VIELYGACSQD---GPLLLIVEYAKYGSLRGFLRESRKVGPGYLGSGGSRNSSSLDHPDERALT
c-SRC LVQLYAVVSE---EPIYIVTEYKSKGSLLDFLKGETGKY---LR
v-RYE LQMLLKFKVDIALQEYLSSRQFLHRDLAARNCHLRDDTVCVADFGLSKIKYSGDYYRQG--RIA
AXL/UFO -QILVEFKADIASQIZYLSTKRFIHRDLEAARNCLNENKSVCVADFGLSEKIYNGDYYRQG--RIA
ARE -QMLVKFADIASQTLSTKRFITR tDLAA.RNC)LNENMSVCVADFGLSEEIYNGDYYAKG--RIA
v-SEA VKELIGFGLQVALGKEYLAQKKFVERDLAARNCNLDETLTVKVADFGLARDVFGKEYYSIRQHRHA
HIR LQEMIQMAAEIADQIAYLNAEKFVHRDLAARNCXVAHDFTVKIGDFGMTRDIYETDYYRKG--GKG
MET VKDLIGFGLQVAKANKYLASKKFVHRDLAARNCMLDEKFTVKVADFGLARDMYDKEYYSVHNKTGA
IGFlR LSEMIQMAGEIAD(MAYLNANKFVSREAARNCNVAEDFTVKIGDFGMTRDIYETDYYRKG--GKG
RET MGDLISFAWQISQQ4QYLAEMKLVHRDLAAREILVAEGRKMRISDFGLSRDVYEEDSYVKR--SQG
c-SRC LPQLVDMAAQIASGKArVERMNYVHRDLRAANILVGENLVCKVADFGLARLIEDNEYTAR ---QGA v-RYK IKPVENlARIESLADRVYTTKSDWVAFGVTf NATRGMTPYPGVQNHEITEYLFHGQRLKKPENCL
AXL/UFOIOIVEWIAIESLADRVTTSKSDVSFGVTRNNIATRGQTP!PGVENSEIYDYLRQGNRLKQPADCL
ARK IKPVKWIAIESLADRVTTSKSDVNLFGVTD4NKIATRGQPYPGVENSEIYDYLRKGNRLKQPVDCL
v-SEA KLPVRNMALESLQTQEFTTKSDNSFGVIJNILLTRGASPYPEVDPYDMARYLLRGRRLPQPQPCP
HIR LLPVRHMAPESLEDGVFTTSSDWSFGVVLWITSLAEQPYQGLSNEQVLKFVMDGGYLDQPDNCP
MET KLPVEWMALZSLQTQEFTTSDVWSFGVVLNZEMTRGAPPYPDVNTFDITVYLLQGRRLLQPEYCP
IGF1R LLPVRHMSPZSLKDGVFTTYSDVSFGVVLWEIATLAEQPYQGLSEEQVLRFVMEGGLLDKPDNCP
RET RIPVEWMAIUSLFDHIY!TQSDVWSFGVLLWKIVTLGGNPYPGIPPERLFNLLKTGHRMERPDNCS
c-SRC KFPIEWTAPKAALYGRFTIKSDWSFGILLTULTTEGRVPYPGMVERBVLDQVERGYRMPCPPECP
v-RYE DELTDIMSSCNRAEPADRPTFSQELVHLUELLESL
AXL/UFO DGLYALSRCNELNPQDRPSFTELREDLZNTLKAL
ARE DGLYALKSRCNELNPRDRPSFAELREDLENTLKLC
v-SEA DTLYGVKLSCWAPTPEERPSFSGLVCELERVLASL
HIR ERVTDLRRMCNQFNPNMRPTFLEIVNLLEDDLHPS
MET DPL!EVNLKCWHPEAEMRPSFSELVSRISAIFSTF
IGF1R DMLFELXIMCNQYNPEMRPSFLEIISSIKEEMEPG
RET EEMYRLMLQCWKQEPDERPVFADISEDLEEMMVER
[image:7.612.58.299.75.335.2]c-SRC ESLHDLNCQCVREEPEERPTFEYLQAFI.DYFTST
FIG. 4. Homology of v-Ryktoother tyrosine kinases.The v-Ryk protein sequence is aligned with those of other tyrosine kinases (sequencesarefromGenBank). Thenameof each protein isonthe left, dashes denotegaps in thesequences introduced for maximal
alignment,and theresidues inv-Ryk thatarealsopresentinatleast twoother kinasesarein boldletters.
Control
three immunized rabbits. This competition was specific,
becausethe controlantigen (vector withnoinsert) couldnot compete successfully for binding of the antibodies to the
70-kDa band (lane 1). In addition, preimmune serumcould not precipitate the 70-kDa protein from 32P-labeled cells
(lanes 8and9).
This70-kDaphosphoproteinwasalsoimmunoprecipitated
frompRV-ryk-transformed CEF (Fig. 8, lane 4) butnotfrom
pUR2AV-transfected cells (lane 2). The size of this phos-phoprotein,70kDa, is consistent with thepredicted molec-ular mass of v-Ryk, as determined from its sequence.
Therefore, this 70-kDa protein is believed to be thev-Ryk oncoprotein.
Since the v-ryk geneencodes aputative tyrosine kinase, weperformedin vitrokinaseassays onimmunoprecipitates
ofv-Ryk. Following incubation of anti-v-Ryk immunopre-cipitates fromv-ryk-transformed CEF and nontransformed
CEF with
[.y-32PjATP,
two proteinswithmolecularmassesof70 and 150 kDa becamephosphorylatedinboth pRV-ryk (Fig. 9, lanes 2 and 4)- and RPL30 (data not
shown)-transformed cells. These two phosphoproteins were not
detected in mock-transfected (data notshown) or
UR2AV-transfected(lane 6)CEF.Phosphoaminoacidanalysisof the
two proteins showed that the phosphorylation was on
ty-rosineresidues(datanotshown).We concluded from these experiments that v-Ryk is indeed a tyrosine kinase. The
70-kDaproteinis believedtobeautophosphorylated v-Ryk,
and the 150-kDa band is either a protein substrate which coimmunoprecipitatedwithv-Rykor av-Ryk-related protein
whichcanalso berecognized
by
theserum weused.v-Ryk antibodies were also used in Western blotting analysis. These antibodies detected a prominent 150-kDa
UR2AV
-c Ak
..:5leBS
pRV-ryk+UR2AV RPL30+UR2AV
FIG. 5. Transformation of CEFby
v-iyk.
CEFweremock transfected(A),transfected with thehelpervirusplasmid pUR2AV (B)orthe clonedv-ryk
constructpRV-ryk plus
pUR2AV(C),
orinfected with theRPL30virusplus helpervirus UR2AV(D).Cellmorphologyat20days
aftertransfectionorinfection is shown.Magnification,
x100.on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.137.474.418.694.2]5982 JIA ET AL.
Control UR2AV
[image:8.612.142.482.77.354.2]pRV-ryk+UR2AV RPL30+UR2AV
FIG. 6. Colonyformation ofv-ryk-transformedcells. As forFig. 5,cellswereputonagar 20days after transfectionorinfection, theplates wereculturedat40°C, coloniesappeared2weekslater,and thesepicturesweretaken 3 weeks later. Magnification, x30.
ConstructContrut UR2AVU2AV+UR2AVpRV-ryk
RPL30
UR2AV+SeruLm111 303
Ao ;-- +
304 305 Prc-lmm1ni
+
± - ± - ± KD Serum P I P I P I
PP1P
aWWSk4WI'm
t g #_ ~~200
6974
TX4xEWXW:
~
69- 46
1 2 3 4 5 6 7 8 9
FIG. 7. Antigen (Ag)competitionassay.v-ryk-transformedCEF
werelabeled in vivo with32Pi,lysed,immunoprecipitatedwith either preimmune(Pre-Imm)serum(lanes8and9)or serum303(lanes 1, 2,and3),304(lanes4and5),or305(lanes6 and7)intheabsence ofantigen(lanes 2, 4, 6,and8)orin thepresenceofimmuneantigen (lanes 3, 5, 7, and9) orcontrol antigen (lane 1), and analyzedon
SDS-7.5% PAGE. The arrowhead shows the bandthatwas
specif-icallyeliminatedbycompetitionwith the immuneantigen.
1 2 3 4 5 6
FIG. 8. Immunoprecipitation of the v-Ryk oncoprotein. CEF transfected withpRV-ryk plus pUR2AV (lanes3 and4)orpUR2AV alone(lanes1and2)orinfectedwith theoriginal RPL30virusmixed withUR2AV(lanes5 and6)werelabeled in vivo with32pi,lysed, immunoprecipitated with either preimmune serum (P) or immune
serum (I), and analyzed by SDS-10% PAGE. The arrowhead indicates the protein that was present only in v-ryk-transformed cells.
- 200
KD
97,4
- 69
- 46
- 30
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.344.536.409.650.2] [image:8.612.87.275.423.648.2]THEv-ryk
-- +
KD 200 200
_ 97.4 97.4
_ 69 69
_ 46
46
- 30
30
1 2 3 4 5 6
FIG. 9. In vitro kinaseassay.CelllysatesfromCEFtransfected witheitherpRV-ryk plus pUR2AV (lanes1to4)orpUR2AV alone (lanes 5 and 6) were immunoprecipitated with either preimmune
serum(P)orimmuneserum(I), subjectedtoanin vitrokinaseassay
in thekinasereactionbuffer, and analyzed bySDS-10% PAGE. The arrowheads indicate the two proteins that were present only in v-ryk-transformedcells.
protein in v-ryk-transformed CEF (Fig. 10). Upon careful
examinationof lanes3 and4 inFig. 10, another specific band
withamolecularmass of 70kDacanbeseen (indicated by
thearrowatthe lowerposition); thisbecameobvious when the filmwasexposed longer (datanotshown). These results
were obtained with serum 303; the same results were
ob-tained withtwootherantisera(datanotshown).These data showed thatpRV-ryk-transformed cells have the same
bio-chemical properties as RPL30-transformed cells, further
provingthattheclonedv-rykgenecanfully recapitulatethe
transforming activityof theRPL30virus.
Biosynthesis of thev-Rykoncoprotein. On Westernblots (Fig. 10),theanti-Rykantibodiespredominately recognized
a150-kDaprotein, suggesting thatthe150-kDa protein is the
precursor ofthe 70-kDa protein. To studythe relationship
betweenthesetwoproteins, the 70-and150-kDa bandswere
excised fromthe invitro kinaseassaygelandsubjectedtoS.
aureus V8 protease mapping. The V8 protease digestion
patternsofthe twoproteinswere similar(Fig. 11), suggest-ing that the 70-kDa protein is derived from the 150-kDa protein.Toruleoutthepossibilitythatthe150-kDaprotein
arosefromdimerization of the 70-kDaprotein,
v-ryk-trans-formed cells were pulse-labeled with [35S]methionine and thenchasedfor various periodswith cold methionine. The results (Fig. 12) demonstrate that the 70-kDa protein is
derived from the 150-kDa protein. The 150-kDa protein
peaked 15 min after the pulse-labeling step and then was
1 2 3 4
FIG. 10. Western blot of thev-Ryk oncoprotein. Cell lysates
from CEF transfected with pRV-ryk plus pUR2AV (lane 3) or
pUR2AValone(lane 2),mocktransfected(lane 1),orinfected with the original RPI30 virus (lane 4) were run on SDS-10% PAGE, transferred to a polyvinylidene difluoride nylon filter, andprobed witha 1:200dilution ofv-Ryk-specificantiserum. Thearrowheads indicate the two proteins that were present only in v-ryk-trans-formed cells.
gradually chasedintothe 70-kDaprotein,whichpeakedat2 h. Because anti-Rykantibodiesweregenerated against the
mature form of v-Ryk, other proteins derived from the
precursorwouldnotberecognized bythis antiserum.
Enve-lope proteins ofretroviruses are known to be first
synthe-sized as theenv polyprotein precursorPr95, which is then cleaved intotwo matureenv proteins, gp85 andgp37, by a
cellular protease(5).Webelieve thatanalogoustothis,inthe
RPL30 virus, an Env-Ryk precursorfusion protein is first
synthesizedand then cellular protease cleaves thisprecursor
intogp85andP69W-37ryk(Fig. 13).Theglycoprotein complex
of RSV lacking the cytoplasmic domain ofgp37 has been
showntobestably expressedonthe cell surface inamanner
similar to thatofwild-type RSV (34). Therefore, the cyto-plasmic tail of gp37 is not required for assembly of the
envelopeglycoproteins intovirions. In Fig. 12,the labeling
of the150-kDaproteinwasstrongerthan that of the70-kDa
protein,consistent with resultsobtained from Western
blot-ting experiments (Fig. 10), which show that the 150-kDa
protein is more abundant than the 70-kDa protein. The in vitro kinaseassaydata(Fig. 9)show that the70-kDamature oncoproteinseemstoexhibithigherkinaseactivitythan the 150-kDaproteinorthatthe150-kDaprotein
autophosphory-lates poorly. V8 mapping revealed that all of the phos-phopeptideswerederived from the 70-kDaprotein (Fig. 11), asexpectedifthe otherprotein (gp85) isextracellular.
Cellular localizationof thev-Ryk oncoprotein. The
recep-torlikestructureof
P69337ryk
suggeststhatthisoncoproteinis localizedonthe cell membrane. Cellfractionation
exper-iments with sucrose gradients were performed to test this pRV-ryk p
+UR2AV 4
P I
_t
_4r,
pRV-ryk UR2AV
PUR2AV
p I p I
KD
pRV-ryk RPL30 UR2AV VOL.66,1992
.":...4.11'.1
"'Wooole,
qmpxmmwlon November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.91.269.76.372.2] [image:9.612.350.525.76.333.2]5984 JIA ET AL.
pRV-ryk
RPL30 KDI
p
-200
pol gp85 gp37
-
97.4
-69
-
46
v-ryk gp37
stop LTR
*
150 kD Envprecursor-Ryk
*
CellularproteaseAGP37
v-RykMKSN=~~~
69kD [image:10.612.318.556.72.251.2]GP85 GP37-Ryk
FIG. 13. Biosynthesisof the v-Ryk oncoprotein. This diagram shows thattheRPL30 virus (top line)first synthesizes a 150-kDa Env-Ryk precursorprotein (middle line)and then has it cleaved into twomatureproteins,gp85and
p69j37-yk
(bottom line). TM,trans-membrane domain.
150kD70kD 150kD70kD
FIG. 11. V8 mapping of two proteins fromv-ryk-transformed cells. The 150- and 70-kDa proteins from either pRV-ryk- or
RPL30-transformedcellswereexcisedfromthe in vitro kinaseassay
gel, subjected toV8mappinganalysisasdescribedinthe text, and
run onSDS-12.5% PAGE.
prediction.Theresults(Fig. 14)showed thatonthebasis of the data from both the in vitro kinase assay and Western
blotting analysis, v-Ryk is indeed located primarily in the
membrane-enrichedfraction rather than in thecytosol frac-tion. This is consistent with the idea that P69gP37-ryk is a
membrane-boundreceptorlikemolecule.
Phosphotyrosine-containingproteinsareincreased in
v-ryk-transformed cells. Since thev-rykgene is atyrosine kinase
oncogeneonthe basis ofsequenceanalysis and the in vitro kinaseassay, weexamined whether therewas anincrease in
phosphotyrosinelevels inv-ryk-transformedcells. Antiphos-photyrosine antibodieswereusedinaWesternblotanalysis
0'
15'
30'
1h
2h
of whole-cell lysates. The results in Fig. 15 show that in v-iyk-transformed CEF, there were multiple
phosphoty-rosine-containing proteins which were not detected in
un-transformed cells; among them were the 150- and 70-kDa
proteins, whichpresumably correspond tov-Ryk
oncopro-teins.Thissuggested that v-Ryk tyrosine kinase activity is required for the transforming phenotype. The identities of those multiple tyrosine-phosphorylated proteins are not known, but they are presumably substrates for the v-Ryk
kinase.
DISCUSSION
Inthis study,we analyzed three previously
uncharacter-ized RPLoncogenic retroviruses; twoof them, RPL25and RPL28,were shown to contain the erbB oncogene in their
genomes. Wethenfurtheridentified,and subsequent
molec-S M S M
4h
8h
--
150kD
.... ..:
.-m
4-
70
kD
1
2
3
4
5
6
7
FIG. 12. Pulse-chase analysisof thev-Ryk oncoproteins. CEF transfected withpRV-ryk plus pUR2AVweremetabolically labeled invivo with[35S]methioninefor 2h,chased for the times indicated at the tops of the lanes, immunoprecipitated by v-Ryk-specific antiserum,andanalyzed by SDS-10% PAGE.
MW
-
200
kD- 97.4
.4
-
69
-
46
1 2 3 4
FIG. 14. The v-Rykoncoprotein is localized onthe cell
mem-brane.CEF transfected withpRV-ryk plus pUR2AVwere fraction-atedintosoluble (S) and membrane(M) fractions as described in
Materialsand Methods. Thesetwofractionswerethensubjectedto either Western blotting analysis with v-Ryk-specific antiserum (lanes 1 and 2) or an in vitro kinase assay (lanes 3 and 4) and analyzed bySDS-10% PAGE. Thearrowsindicatethetwoformsof thev-Rykprotein. MW,molecularmass.
RPL30
-
30
- 21.5
- 14.3
I I . r..
J. VIROL.
i
.,
on November 9, 2019 by guest
http://jvi.asm.org/
[image:10.612.104.261.77.330.2] [image:10.612.353.523.503.648.2] [image:10.612.67.304.539.677.2]pRV-ryk RPL30
UR2AV - + + +
4r .1:
.,F 7t .-*,: :,- o
200
- 97.4
-- 69
- 46
[image:11.612.99.266.77.334.2]1 2 3 4
FIG. 15. Phosphotyrosine-containing proteins are increased in
v-ryk-transformedcells.AWesternfilterwaspreparedasdescribed
inthe legendtoFig. 10 and blotted with polyclonal antiphosphoty-rosine antibodies.The arrowheads indicate the positionsof thetwo
v-Rykproteins.
ularlycloned, anoveltransforminggenefrom thegenomeof
the third virus, RPL30. This viral oncogene, v-ryk, is a member of the protein tyrosine kinase gene family. It
in-serted itself into the RPL30 virus envelope gene without
deletionofanyof the viralsequencesand is fused inframe
with thegp37gene upstream toform a69-kDa gp37-v-Ryk
transmembrane protein, P69gp37-yk. v-ryk can transform CEF, form colonies, and induce tumors in chickens, fully recapitulating the transformingactivity of theoriginalRPL30
virus. The v-Ryk oncoprotein is first synthesized as a
150-kDaprecursorproteinand then cleavedinto themature
p69gp37-ryk protein. v-Ryk has a tyrosine kinase activity in
vitro and is phosphorylated in vivo in v-ryk-transformed cells. Furthermore, elevated phosphotyrosine-containing protein levelsarefound in thesev-ryk-transformed cells.
Viral oncogenes are transduced from cellular genes by nononcogenic retroviruses. The v-erbB oncogene was first identified in the AEVgenome (strainsAEV-ES4 and AEV-R), together with another oncogene, erbA (47). The erbB
gene,however,isnotalways cotransduced into retroviruses
with theerbA gene, since erbB transducingviruses lacking
erbA have also been isolated (3, 30, 50). The finding that RPL25 and RPL28both contain only the erbB oncogene is
consistent withthe notion that the transforming activity of
erbBis stronger.
All acute oncogenic retroviruses lose part of their viral
genes during the oncogene transduction process, thus be-comingreplication deficientand relyingonhelpervirusesto
replicate and assemble infectious viral particles (43). Al-though the Schmidt-Ruppin and Prague strains of RSV
contain thev-srcgeneyetpreserveallof the viral structural
genes andarereplication competent, there isevidence that
the original RSV was a replication-defective transforming virus (16). Thecurrent replication-competent strains of RSV wereprobably produced after a second recombination event. TheRPL30virus is the first example of a virus that did not lose any of the viral sequences in transducing a proto-oncogene, although the envelope protein gp37 coding region was disrupted by insertion of the v-ryk gene, rendering the virusdeficient in replication. The reason that no viral genes were deleted in RPL30 is probably that these are homolo-goussequences between retroviral sequences and the c-ryk
gene at the region of the insertion. This will become clear when the sequence for the c-rykproto-oncogene is known.
Some viral tyrosine kinase oncogenes were transduced from cellular receptors for peptide growth factors. For example, v-erbB in AEV was transduced from the epidermal growth factor receptor (15), and v-sea was presumably transduced from a cellular receptor (8, 26). Since v-Ryk more closelyresembles receptor-type tyrosine kinases than nonreceptor tyrosine kinases, c-rykmay encode another cell surfacereceptor. We are currently cloning and characteriz-ing thec-ryk gene, and results from preliminary experiments have confirmed thatc-ryk isindeed a receptor-type tyrosine kinase (26b). Therefore, it is very likely that transduction of the c-ryk gene by the RPL30 virus rendered this kinase active in the absence of its ligand. The two oncogenes erbB and sea are particularly interesting because of their pro-cesses of viral transduction and the structures of their final oncogene products. These two oncogenic proteins are simi-lar to v-Ryk in that their basic mechanism of activation is truncation ofextracellular domains and the final products still maintaintransmembrane domains, albeit the one of v-ErbB is derived from theepidermal growth factor receptor and those of v-Sea and v-Ryk came from viral env product gp37. Althoughrelatively high amino acid identities with Axl-UFO and Ark(67.3%) in the kinasecatalytic domains were found, thepreliminary data on the cloning ofc-rykindicate that they areencoded by different genes (26b). In addition, the catalytic domain of v-Ryk contains sequences similar to polymerase chain reaction fragments JTKll (32b), tyro-12, tyro-7, and tyro-3 (28a). Particularly, tyro-12 has 53 of the total of 54 amino acids identical to those of v-Ryk. However, the rela-tionships between v-Ryk and thesepolymerase chain reaction fragments are difficult to judge because these polymerase chain reaction fragments are only 50 to 60 amino acids long and are derived from the most conserved region within the tyrosine kinase catalytic domain.
The results of Western blot analysis (Fig. 10) and the pulse-chase experiment (Fig. 12) indicate that the 150-kDa precursor protein is moreabundant thanp69w37-ryk in
v-ryk-transformed cells. This raises the interesting question of whether the Env-Ryk precursor protein has transforming ability or whether cleavage is required for transformation. Other unanswered questions about the v-ryk oncogene in-clude the function of the C-terminal sequences ofv-rykand the small inserts in the kinase domain. We are currently addressing these questions by mutagenesis of thev-ryk onco-gene.
ACKNOWLEDGMENTS
We are particularly indebted to L. B. Crittenden of the Avian Disease andOncologyLaboratory,EastLansing, Mich., for gener-ouslyproviding us with theRPL30virus. Wesincerely thank K. A. Sokol of the Laboratory Animal Research Center for excellent assistance inpathology studies and many members of our laboratory fortechnical help. We are grateful to G. Scholz for critical reading ofthemanuscript.
VOL.66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
5986 JIA ET AL.
Thiswork was supported by grant CA44356. REFERENCES
1. Akiyama, T., S. Matsuda, Y. Namba, T. Saito, K. Toyoshima,
and T. Yamamoto. 1991. The transforming potential of the
c-erbB-2 protein is regulated by its autophosphorylation at the
carboxyl-terminal domain.Mol. Cell. Biol. 11:833-842.
2. Alitalo, K., J. M. Bishop, D. H. Smith, E. Y. Chen, W. W.
Colby, andA. D. Levinson. 1983. Nucleotide sequence ofthe
v-myc oncogene ofavian retrovirus MC29. Proc. Natl. Acad. Sci. USA80:100-104.
3. Beug, H., M. J. Hayman, M. B. Raines, H. J. Kung, and B.
Vennstrom. 1986. Rous-associated virus 1-induced
erythroleu-kemic cells exhibitaweaklytransformed phenotype in vitro and
release c-erbB-containing retroviruses unable to transform
fi-broblasts.J. Virol.57:1127-1138.
4. Brugge, J. S., and R. L. Erikson. 1977. Identification of a
transformation-specific antigen induced by an avian sarcoma virus. Nature(London)269:346-348.
5. Buchhagen, D. L., and H. Hanafusa. 1978. Intracellular precur-sors tothe major glycoprotein of avianoncoviruses in chicken
embryo fibroblasts. J. Virol.25:845-851.
6. Chen,I.S.Y., T. W. Mak, J. J. O'Rear, and H. Temin. 1981. Characterization of areticuloendotheliosis virus strain T DNA and isolation of a novel variant of reticuloendotheliosis virus strain Tbymolecularcloning. J.Virol. 40:800-811.
7. Cleveland, D.W., S. G. Fischer, M. W. Kirschner, and U. K. Laemmli. 1977. Peptide mapping by limited proteolysis in so-dium dodecyl sulfate and analysis by gel electrophoresis. J. Biol. Chem. 252:1102-1106.
8. Cooper, G. M. 1990. Oncogenes. Jones & Bartlett Publishers, Boston.
9. Cross,F.R.,and H.Hanafusa. 1983.Local mutagenesisof Rous sarcomavirus: themajorsites oftyrosine and serine
phosphor-ylation ofp6Orcaredispensablefortransformation.Cell 34:597-607.
10. Curran,T., G.Peters,C. Van Beveren, N. M. Teich, and I. M. Verma.1982. FBJmurineosteosarcomavirus:identificationand molecularcloningofbiologically active proviralDNA. J. Virol.
44:674-682.
11. Debuire, B., C. Henry, M. Benaissa, G. Biserte, S. Saule, P. Martin, and D. Stehelin. 1984. Sequencing the erbA gene of avian erythroblastosis virus reveals a new type of oncogene. Science224:1456-1459.
12. DeLorbe, W. J., P. A. Luciw, H. M. Goodman, H. E. Varmus, andJ.M.Bishop. 1980.Molecularcloning and characterization of avian sarcoma virus circular DNA molecules. J. Virol.
36:50-61.
13. Devare,S.G., E. P. Reddy, K. C. Robbins, P. R. Anderson, S. R.
Tronick,andS. A. Aaronson.1982.Nucleotidesequence of the
transforminggene ofsimian sarcoma virus. Proc. Natl. Acad. Sci. USA 79:3179-3182.
14. Donner,L., L. A. Fedele, C. F. Garon, S. J. Anderson, and C. J. Sherr.1982.McDonough felinesarcomavirus:characterization of themolecularlyclonedprovirusand its feline counterpart. J. Virol. 41:489-500.
15. Downward, J., Y. Yarden, E. Mayes, G. Scrace, N. Totty, P.
Stockwell, A. Ullrich, J. Schlessinger, and M. D. Waterfield.
1984.Closesimilarityofepidermal growthfactor receptor and v-erb-B oncogeneproteinsequences. Nature(London) 307:521-527.
16. Dutta, A., L.-H. Wang, T.Hanafusa, and H. Hanafusa. 1985. Partial nucleotide sequence of Rous sarcoma virus-29 provides evidence that the original Rous sarcoma virus was replication defective. J. Virol. 55:728-735.
17. Ellis,R.W.,D.DeFeo, J. M.Maryak,H. A. Young, T. Y. Shih, E. H. Chang, D. R. Lowy, and E. M. Scolnick. 1980. Dual
evolutionary origin for the rat genetic sequences of Harvey murinesarcomavirus. J. Virol. 38:408-420.
18. Ellis,R.W., D. DeFeo, T. Y. Shih, M. A. Gonda, H. A. Young, N.Tsuchida,D. R.Lowy, and E. M.Scolnick 1981.Thep21ras genes ofHarvey and Kirsten sarcoma viruses originate from
divergent members of a family of normal vertebrate genes.
Nature (London) 292:506-511.
19. Feldman, R. A., T. Hanafusa, and H. Hanafusa. 1980.
Charac-terization ofprotein kinase activity associated with the
trans-forminggene product ofFujinami sarcoma virus. Cell
22:757-765.
20. Foster, D. A., J. B. Levy, G. Q. Daley, M. C. Simon, and H. Hanafusa. 1986. Isolation of chicken cellular DNA sequences with homology tothe region of viral oncogenes that encodes the tyrosine kinase domain. Mol. Cell. Biol. 6:325-331.
21. Fredrickson, T. N., H. G. Purchase, and B. R. Burmester. 1964. Transmission of virus from field cases of avian lymphomatosis. III. Variation in the oncogenic spectra of passaged virus
iso-lates. Natl. Cancer Inst. Monogr. 17:1-29.
22. Hamaguchi, M., C. Grandori, and H. Hanafusa. 1988. Phosphor-ylation of cellular proteins in Rous sarcoma virus-infected cells: analysis by use of anti-phosphotyrosine antibodies. Mol. Cell. Biol. 8:3035-3042.
23. Hanafusa, H. 1969. Rapid transformation of cells by Rous sarcomavirus. Proc. Natl. Acad. Sci. USA 63:318-325. 24. Hanks, S. K., and A. M. Quinn. 1991. Protein kinase catalytic
domain sequencedatabase: identification of conserved features of primary structure and classification of family members. Methods Enzymol. 200:38-62.
25. Hanks, S. K., A.M. Quinn, and T. Hunter. 1988. The protein
kinasefamily:conservedfeatures and deduced phylogeny of the
catalytic domains.Science 241:42-52.
26. Hayman, M. J., G.Kitchener, P. K. Vogt, and H. Beug. 1985.
Theputativetransforming protein of S13 avian erythroblastosis
virus is a transmembrane glycoprotein with an associated pro-tein kinase activity. Proc.Natl. Acad. Sci. USA 82:8237-8241. 26a.Janssen, J. W. G., A. S. Schulz, A. C. M. Steenvoorden, M.
Schmidberger, S. Strehl, P. F. Ambros, and C. R. Bartram.
1991. A novelputative tyrosinekinasereceptor withoncogenic
potential. Oncogene 6:2113-2120.
26b.Jia,R., and H.Hanafusa.Unpublisheddata.
27. Kan, N. C., C. S. Flordellis, G. E. Mark, P. H.Duesberg, and
T. S.Papas.1984.Nucleotide sequence of avian sarcoma virus MH2: two potential onc genes, one related to aviansarcoma
virus MC29 and the other related to murine sarcoma virus 3611. Proc.Natl. Acad. Sci. USA81:3000-3004.
28. Klempnauer, K.-H., T. J. Gonda, and J. M. Bishop. 1982. Nucleotide sequence of the retroviral leukemia gene v-myb and its cellular progenitor c-myb: the architecture of a transduced oncogene.Cell 31:453-463.
28a.Lai, C., and G. Lemke. 1991. An extended family of protein-tyrosine kinase genes differently expressed in the vertebrate nervous system. Neuron 6:691-704.
29. Li, Y., C. M. Turck, J. K. Teumer, and E. Stavnezer. 1986. Unique sequence, ski, in Sloan-Kettering avian retroviruses with properties of a new cell-derived oncogene. J. Virol. 57:
1065-1072.
30. Miles, B. D., and H. L. Robinson. 1985. High-frequency trans-duction ofc-erbB in avian leukosis virus-induced erythroblas-tosis. J. Virol. 54:295-303.
31. Naharro, G., K. C. Robbins, and E. P. Reddy. 1984. Gene product of v-fgr onc: hybrid protein containing a portion of actin and a tyrosine-specific protein kinase. Science 223:63-66. 32. Neckameyer, W. S., and L.-H. Wang. 1985. Nucleotide
se-quence of avian sarcoma virus UR2 and comparison of its transforming gene with other members of the tyrosine protein kinaseoncogene family. J. Virol. 53:879-884.
32a.O'Bryan,J. P., R. A. Frye, P. C. Cogswell, A. Neubauer, B. Kitch, C. Prokop, R.Espinosa III, M. M. Le Beau, H. S. Earp, and E. T. Liu. 1991. axl, a transforming gene isolated from primary human myeloid leukemia cells, encodes a novel recep-tortyrosine kinase. Mol. Cell. Biol.11:5016-5031.
32b.Partanen, J., T. P. Makela, R. Alitalo, H. Lehvaslaiho, and K. Alitalo. 1990. Putative tyrosine kinases expressed in K-562 human leukemia cells. Proc. Natl. Acad. Sci. USA 87:8913-8917.
33. Pearson, W. R., and D. J. Lipman. 1988. Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85:2444-2448.
J. VIROL.