0022-538X/92/053118-07$02.00/0
Copyright X 1992, AmericanSocietyforMicrobiology
The
Pathogenesis of Infection with Minute Virus of Mice Depends
on
Expression of the Small Nonstructural
Protein
NS2 and
on
the
Genotype of the Allotropic Determinants VP1
and
VP2
DAVID G. BROWNSTEIN,l* ABIGAIL L. SMITH,' ELIZABETH A.JOHNSON,' DAVIDJ. PINTEL,2 LISAKAYNAEGER,2 ANDPETERTATTERSALL3
Sectionof Comparative Medicine' andDepartment of Laboratory Medicine,3Yale University School of Medicine, New Haven, Connecticut 06510, and Department of Molecular Microbiology and
Immunology, University of Missouri School of Medicine, Columbia, Missouri 652122 Received 20 December1991/Accepted 6 February 1992
Neonatal C3EH/Hemicewereoronasally inoculatedwith similar doses of fourgenotypesofminute virus of
mice(MVM). MVMp,afibroblast-specific variant, causedanasymptomatic infection.MVM(1035),achimera
whichhad the allotropic determinant ofvirulentMVMiinsertedontoanMVMp background, causedalethal infection and renal papillary infarcts, the hallmark of MVMi infection. MVMi(NS2-1990), the virulent lymphocyte-specific variantmutated toeliminateNS2 synthesis,was infectious but caused an asymptomatic
infection. Sequential virus titration, histology, in situhybridizationwith afull-lengthMVMigenomicprobe,
andimmunohistochemistryfor viralcapsid antigenwereusedtocomparethepathogenesisof infection with the four MVM genotypes. Infectious virus was recovered from multiple organs of mice infected with MVMi, MVMp, andMVM(1035) but notfrommice infected withMVMi(NS2-1990). MVMptiterswere lower than MVMititersinallorgansexcepttheintestine. MVM(1035) titerswerehigherthan MVMi titers in allorgans
except the blood. MVMpwas localized to connective tissue elements of the intestine, to cells in mesenteric
lymphnodes,andrarelytocellsinotherorgans.MVM(1035)waslocalizedtomultipleorgansandsharedthe
sametargetcells,endothelium, lymphoid cells,andhematopoietic cells,asMVMi.MVM(1035)alsoreplicated
inexternal germinalcellsofthe cerebellumandsmoothmuscle cellsofthe stomachandcolon,whichwerenot targetsof MVMiorMVMpinfection.MVMi(NS2-1990) replicated toalimiteddegreein someMVMitarget
organs.
Theautonomousmurine parvovirusminutevirus of mice (MVM) contains a5.1-kb, linear, single-stranded DNA ge-nome that encodes only four translation products: two
nonstructural proteins, NS1 and NS2, and twocapsid
pro-teins, VP1 and VP2 (2, 7-9). To replicate, MVM requires cellular factors that are transiently expressed during the S
phaseof thecell cycle(3, 25, 27) and other cellular factors that are expressed during differentiation (3, 19, 24). Two serologically indistinguishable strains ofMVM,termed allo-tropic variants, require different differentiation-dependent factors for theirreplication in vitro. MVMp, the prototype strain, productivelyinfects cells of fibroblastorigin (24, 26). MVMi, avariantstrain,productively infects T cells(4, 17). MVMp and MVMi are 97% homologous at the nucleotide level(2). Thefibrotropic determinantmapsto a 236-nucleo-tidefragment of thegenome ofMVMp in the region of the capsid gene common to both VP1 and VP2 (1, 12). The lymphotropic determinant mapsto an 820-bp region of the overlapping capsid gene of MVMi that encompasses the fibrotropic determinant (12). The 83-kDa NS1 is a stable
nuclear phosphoprotein that is required for viral DNA replicationaswellasfortransactivation of theP38promoter
(7,9-11, 21).The25-kDa NS2 isalabile phosphoprotein that
shares its first 85 amino acids with NS1. Little is known about the function ofNS2. It is required for efficientvirus production and viral DNA replication in cells from the natural hostspecies butnot from heterologous species (16,
*Correspondingauthor.
20). The natural host requirement for NS2 has also been reportedin vivo (16).
MVMi causes a lethal infection in perinatal mice of
susceptible strains(5, 13)whereasMVMpinfection is inap-parent (13). After oronasal inoculation, MVMi causes a
generalized infection primarily of endothelium, lympho-cytes, and hematopoietic cells and produces bilateral in-farcts of the renalpapilli (5).Thecourseof avirulentMVMp
infection in neonatal mice has not been characterized in detail. MVMp has been reported to remain confinedto the oropharynxofneonatal mice afteroronasal inoculation(13). The genetic determinants of MVM pathogenicity have not beeninvestigated.
Thepurposesof thisstudywereto(i)characterize
apatho-genicMVMpinfection inperinatalmice andcompareit with pathogenic MVMi infection, (ii) determine the effect on
perinatalinfection ofreplacingtheallotropicdeterminant of MVMp with that of MVMi using an infectious intertypic
recombinant,and(iii)determinewhether failure of MVMito
express NS2altersits pathogenesisin neonatalmice, using
an NS2mutant.
MATERUILS AND METHODS
Mice. AdultC3H/HeSnJmicewereobtained from Jackson
Laboratory,BarHarbor,Maine. Male and female micewere
pair housed in Micro-isolator cages (Lab Products, May-wood, N.J.),using husbandry practicesthatguardedagainst infection with common murine infections including MVM
andthat have beendescribedpreviously (5).Micewerefree ofinfection with MVM at the time ofinoculation as
deter-3118
on November 9, 2019 by guest
http://jvi.asm.org/
MVM INFECTION OF MICE 3119 mined by routine serological monitoring of the dams.
Be-causeof the stronginfluence ofhost age onMVM
infection,
damswereobserved twicedailyforparturitionandcare was takento ensurethatmicewere infected within 24 h of birth. Virus strains and
infectivity
titrations. A clonal isolate ofMVMi (26), originally recovered as a contaminant of ELA lymphoma cells (4), was passaged twice in S49 ITB2 cells (BALB/c
lymphoma
cells). Aclearedlysate
wasused asthe virus stock.The cloningofMVMpbyterminal dilution in A9ouabrl 1 cells has been described
previously
(25).
A clearedlysate
was used asvirus stock.
MVM(1035)was
produced
fromachimericMVMplasmid,
pPI1035, the construction of which has been described elsewhere (12). Briefly, thefragment ofMVMi between the EcoRI andXbaI restriction sites
(nucleotides
3522 to4342)
was cloned into the parentplasmid,
pMM984, which con-tained an infectious MVMp genome,by
way of severalplasmid intermediates. Infectious virus was
produced by
calcium phosphate transfection of
Hyb 1/11
cells(18).
The identity of the clone was confirmed byrestriction endonu-cleaseandfragment-length
analysis.
Viruswas notpassaged
priorto use.
Construction of
MVMi(NS2-1990)
wasessentially
as de-scribedpreviouslyfor theanalogous
mutantofMVMp
(20).
The MVMi HaeIII fragment
(nucleotides
1854 to2378),
cloned into M13mpl9, was used as mutationtemplate
to-getherwith a
mutagenic
oligonucleotide
of MVMi sequence for site-directedmutagenesis (15).
Theoligonucleotide
intro-duced an A-C transversion at nucleotide1990,
which dis-abled the MVM R2 largesplice
acceptorsite,
thusprevent-ing NS2 production, while
leaving
NS1 unaltered. Aftermutagenesis, the mutantMVMiBstEII-XhoI
fragment
(nu-cleotides 1886 to
2072)
was rebuilt back into the infectious clone ofMVMi,
and the insertedfragment
wascompletely
sequenced to ensure the absence of additional mutations. Themutant clonewas transfected ontoNB324 K
cells,
and infectious virus was releasedby
standardfreeze-thawing.
Viruswas not
passaged prior
to use.BHK-21cellswereused for
primary
isolation and titration ofMVMi, MVMp,MVM(1035),
andMVMi(NS2-1990).
Tis-sue homogenates(1
to10%,
wt/vol)
were screened for infectious virusbyfluorescent focus assay(23).
Anenzymeimmunoassay with
sensitivity
identical to the fluorescent focus assaywasusedtoquantify
virus intissuehomogenates
(23). Titers were
expressed
as fluorescentfocus-forming
units or median infectious doses
(ID50)
per gram of tissue. The limit of detection wasapproximately
100focus-forming
units per g. BHKcellswereequally
permissive
forMVMi,
MVMp, and NS2mutantsofMVMp
asdescribed elsewhere(20,23). Titrations of virus stocks indicated that this
applied
toMVM(1035) and the NS2mutantof MVMi as well. Mouseinoculations and
necropsies.
Mice less than 24 h old were inoculated oronasally withapproximately
103focus-forming units of each virus in S p.1. This dose wasselected because it was the
highest
common dose that could be attainedforthe fourMVMgenotypes. This dosewas infec-tious for the four genotypesasjudged by
the appearance of convalescent serum antibodies in mice infected with aviru-lent strains and bymortality
withtypical
histopathological
changesin mice infected with virulent viruses. The necropsy methods and tissue fixation have been describedpreviously
(5).
Histology. Fixed tissueswere embedded in
paraffin,
sec-tioned at 5 ,um, adhered to
poly-D-lysine-coated slides,
and stained withhematoxylin
and eosin. In situhybridization
and
immunohistochemistry
wereperformed
onadjacent
sections.Probe
preparation
and in situhybridization.
The MVMprobe
wasprepared
asdescribedpreviously
(5).
Briefly,
the entire MVMi genome, cloned intopSP65
(Promega,
Madi-son,Wis.),
was excised from the BamHIsite,
gel
purified
from 0.7%low-melting-temperature
agarose(Boehringer
Mannheim
Biochemicals,
Indianapolis,
Ind.),
and labeledwith 32P
by
using
arandomprimer
extension kit(Bethesda
ResearchLaboratories, Inc.,
Gaithersburg,
Md.).
The meth-ods of sectionpreparation,
hybridization,
washes,
andde-velopment
have been describedpreviously
(5).
Immunohistochemistry.
Rabbitantiserumto MVMcapsid
antigens
wasprepared
asdescribedpreviously
(24).
Sectionswere stained
by
the indirectstreptavidin-biotin
complex
method used
previously
(5).
Semiquantitative
analysis
ofin situhybridization.
Hybrid-izedtissue sectionswereviewedat amagnification
ofx100. The number of nuclei thathybridized
with the MVMprobe
were counted for each field and each organ. The mean
number of
positive
cells per field per organwas calculated forindividualmice. Ascoreof 0to4+wasassigned
for each organby
dividing
thetotal ofmeansby
the number of mice.Ascoreof 0 indicated thatno
positive
cellsweredetected ina
specific
organin anymice infected with avirusgenotype.
Ascoreof 1+indicated thatmorethanzerobut less thanone
cell per fieldwas
positive.
Ascoreof 2+ indicated that fromoneto less thanfive cells per fieldwere
positive.
Ascoreof3+ indicated that from 5 toless than 10 cells per fieldwere
positive.
A score of 4+signified
that 10 or more cells perfieldwere
positive.
Estimation of total MVM DNAin organs. Total DNAwas
extracted and
purified
from organsby
themethodofKrieg
et al.(14).
DNAwasdenatured with 0.5 N sodiumhydroxide,
incubated at65°C
for 10 min,rapidly
cooled onice,
and neutralized with ammonium acetate. Tenmicrograms
of denatured DNAwasblotted ontonylon
membranes (Gene-ScreenPlus;
Dupont,
NEN ResearchProducts, Boston,
Mass.) by
using
adot-blotapparatusandbakedat80°C
for 2 h. Included on the membranes weredoubling
dilutions of MVMiDNAcutfromtheBamHI site of thepSP65
plasmid
andgel
purified.
Membraneswereprehybridized
and hybrid-ized with the32P-labeled
MVMprobe
containing
5 x106
cpm/ml
aspreviously
described(5).
Membraneswereauto-radiographed,
andamountsof viral DNAwereestimatedto the nearest standard dilution.Experimental
design.
Clinical responses to oronasal inoc-ulations withMVMp,
MVM(1035),
andMVMi(NS2-1990)
were studied with one or more litters of
C3H/HeSnJ
mice. Litters were examineddaily,
and deathswere noted. Mice that survived infection were bled at 3 weeks of age, andserum antibodies to MVM were detected
by
afluorescent-antibody
assay(23).
Selected mice that diedwerenecropsied
to determine the cause of death. Mice from 51 litters ofC3H/HeSnJ
mice were infected withMVMi,
MVMp,
orMVM(1035)
and werenecropsied
onpostinoculation
days
(PID) 3, 5,
7,
and 9.The numberofmicesampled
perinterval varied with litter size andmortality.
From 10to21miceweresampled
per virus strain per interval. WithMVMi(NS2-1990),
asingle
sampling
intervalwasselectedbecauseof thelimited
quantity
of infectious virus stock that wasavailablefrom transfected cell cultures. The interval that was
se-lected,
PID5,
wasjudged
to beoptimal
for both infectious virus and viralproducts
in tissue based on results for theother strains of virus. Three litters of mice were infected
with
MVMi(NS2-1990).
Tissues from half the mice were VOL.66, 1992on November 9, 2019 by guest
http://jvi.asm.org/
assayed for infectious virus. Tissues from the remaining
mice were processed for histology, in situ hybridization,
and immunohistochemistry. Total DNAwasextracted from selected tissues offive miceinfected with MVMi and MVM (1035) for comparison of total viral DNA by dot-blot
hybrid-ization. Five uninfected control micewerenecropsied, and tissues were processed for histology, in situ hybridization,
and immunohistochemistry. The clinical responses, histol-ogy,quantitative and qualitative in situhybridization, immu-nohistochemistry, and peak virus titers of themice infected with MVMi have beenpreviously reported (5).
RESULTS
Clinical responses. Perinatal C3H/He mice were infected oronasally with approximately 103 focus-forming units of MVMp,MVM(1035),orMVMi(NS2-1990).MVM(1035)was MVMp with the 820-nucleotide EcoRI-to-XbaI fragment
replaced by that ofMVMi. This fragment encoded overlap-ping capsid sequences which included the allotropic deter-minant.MVMi(NS2-1990)wasMVMi with atransversionat nucleotide 1990 which madenodetectable NS2isoforms. All 22mice that were infected with MVM(1035) and thatwere monitored clinically died (average day of death, 8.9 ± 1.3).
At necropsy, selected mice had bilateral renal papillary
infarcts.There was nomorbidityormortalityamong25 mice thatwereinfected with MVMpor20mice thatwereinfected with MVMi(NS2-1990). Mice that were inoculated with MVMp and MVMi(NS2-1990) developed serum antibodies toMVM.
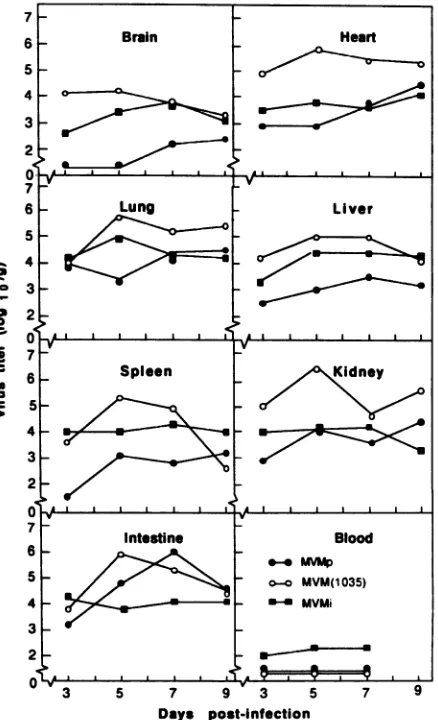
Virus isolation. Infectious virus was recovered from the sevenorgansthatwereexamined in mice thatwereinfected with MVMi, MVMp, and MVM(1035) (Fig. 1). Infectious virus was not detected in the blood of mice infected with MVMp or with MVM(1035). A low-level viremia, which plateaued between PID 5 and 7, was detected in mice infected with MVMi.Noinfectious viruswasdetected in any organsof mice infected with MVMi(NS2-1990).
MVMptiterswerelower than thoseof MVMi in thebrain,
liver, and spleen; similar to those of MVMi in the heart, lung, and kidney; and higher than those of MVMi in the intestine(Fig. 1).Intheintestine, the peak MVMp titer was 100-fold higher than the peak titer for MVMi. MVM(1035)
titers werehigher than MVMi titers in all organs except the blood. The largest difference in infectivity titers (100-fold)
between MVM(1035) and MVMi occurred in the heart,
kidneys, andintestine. Intestinal titers ofMVM(1035)were similarto those of MVMp.
Histology. The histological appearance of organs ofmice infected with MVMp and MVMi(NS2-1990) was
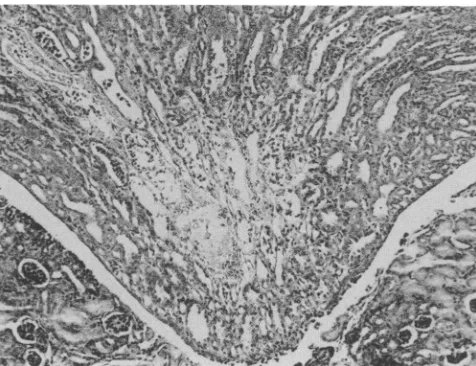
indistin-guishablefrom that of organs of control mice. Mice infected with MVM(1035) had bilateral renal papillary infarcts after PID 5 (Fig. 2). These were identical in appearance and temporal onset to those previously described in mice in-fected with MVMi (5).
In situ hybridization and immunohistochemistry. Tissue sections from infected and control mice were hybridized with a double-stranded DNA probe that was derived from the entire MVMi genome. Adjacent sections were stained for the presence ofMVMcapsid antigens. Tissues from control mice had no detectable MVM nucleotide sequences or
capsid antigens.
Semiquantitative analysis of the density of cells with nuclei that bound the MVM probe on PID 5 was performed on the eight organs listed in Table 1. The intestine of mice infected with MVMp contained more labeled cells than other
5
0-R
a 0
S
S.
3 5 7 9v 3 5 7 9
[image:3.612.332.551.76.436.2]Days post-infection
FIG. 1. Infectious virus titers in mice infected with MVMi, MVMp, andMVM(1035).
organs. The number of labeled cells in the intestine of mice infected withMVMpexceeded the numberof labeled cells in theintestine of miceinfectedwith MVMi.Organsotherthan
intestine andlymphnodes ofmice infected with MVMp had meansof fewer thanonelabeledcell per x100field. Mesen-teric lymph nodes contained two to five labeled cells per
field, whereas other lymph nodes contained few or no labeled cells. Most organs of mice infectedwith MVMi had atleastsix labeled cells per field. Thedensityof celllabeling
wassimilar in organs of mice infected withMVM(1035)and MVMi. MVMi(NS2-1990)DNA was localized to the nuclei ofonlyafew cellsintheintestine,spleen,liver,and heart.A
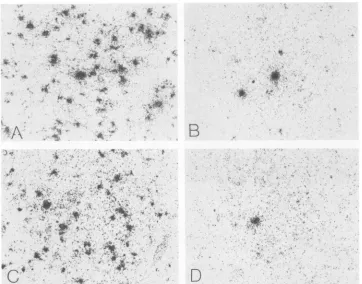
comparison of cell labeling densities in the livers of mice infected with the fourMVMgenotypesis shown in Fig. 3.
Cells in the lamina propria and submucosa of the small
intestine, cecum, and colon of mice infected with MVMp
had nuclei that were labeled by the MVM probe (Fig. 4).
Similar cells in adjacent sections also expressed MVM
capsid antigens. Cells thathybridized withthe MVMprobe
in the intestine appeared to be fibroblasts based on their location and morphology. In the liver and
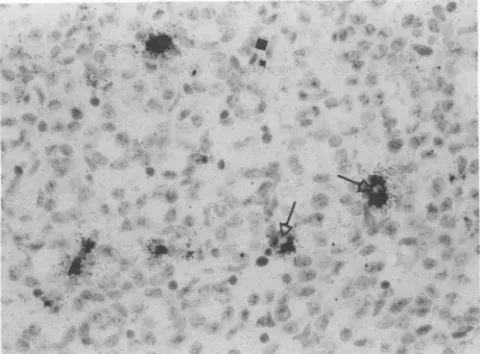
lymph
nodes, however,the few cells thatwerelabeled didnotappeartobe fibroblasts. The rare labeled cells in the liver were in sinusoids either aloneoramonghematopoieticcells(Fig. 5).Labeled cells were scattered throughout the mesenteric
on November 9, 2019 by guest
http://jvi.asm.org/
MVM INFECTION OF MICE 3121
_0gl WL -, _?.t. v40 . -- a .---*-.
FIG. 2. Central infarct of renalpapillaonPID7 in C3Hmouse
infected withMVM(1035). Hematoxylin and eosin. x160.
lymph node (Fig. 6). In no organs of mice infected with MVMpwastherehybridization of the probe with endothelial cells, major sites of MVMi replication (5).
Cell nuclei that hybridizedwith the MVM probe in mice infected with MVM(1035) were similar in distribution and morphologytocells labeled in MVMi-infected mice(5),with severalexceptions. LikeMVMi, MVM(1035) replicated
pri-marily in the capillaryendothelium of the heart andkidneys
(Fig. 7) and in hematopoietic cells in the liver. Nuclei of these target cells bound the MVMprobe and expressed viral capsid antigens. Lymphoid cells in lymph nodes were also labeled and expressed capsid antigens.
Inthe brain andgastrointestinaltractofmice infected with
MVM(1035),cellswerelabeledbythe MVMprobethatwere notlabeled inmice thatwereinfected with MVMiorMVMp. Endothelial cells were the principal sites of MVM(1035)
replication in the brainastheywereinMVMi-infectedmice,
but inaddition, cells in the external germinal celllayerof the cerebellum also contained viral DNA. The latter did not supportdetectablereplicationofMVMporMVMi(5). Inthe
[image:4.612.60.298.76.259.2]gastrointestinal tract, mice infected with MVM(1035) had extensivelabelingof smooth musclenuclei,especiallyin the stomach andcolon, in additionto endothelium and intersti-tial cells of the lamina propria and submucosa. Smooth
TABLE 1. Semiquantitativeanalysisof in situhybridization for MVM DNAin organs from neonatalC3H/Hemice infected
withMVMi, MVMp, MVM(1035),andMVMi(NS2-1990)
Densityof nuclei thathybridizedwith MVMprobea
Organ
MVMi MVMp MVM(1035) MVMi(NS2-1990)
Brain + - ++
Heart +++ + ++ +
Lung ++++ + +++
Liver ++++ + +++ +
Spleen +++ + +++ +
Kidney +++ + ++++
Intestine +++ ++++ ++++ +
Lymph node +++ ++ ++
aNumber ofpositivecellsperx100field: -,0;+,morethan 0 butlessthan
1;++, 1 to less than5; + ++, 5toless than10; + +++,morethan 10.
muscle cells were not targets of infection by MVMp or MVMi (5).
Cells thatwerelabeled inmice infected with MVMi(NS2-1990) were too infrequent to accurately assess their pheno-types. They were located in thesinusoids of the liver, in the interstitium of the heart, liver, and intestine, and in the lymphoid and hematopoietic elements of the spleen.
Estimation of viral DNA in organs of mice infected with MVMiandMVM(1035). Total DNA was extractedonPID S from theintestine, spleen, and kidneys of mice infected with MVMi andMVM(1035), and total MVM DNA was estimated by dot-blot hybridization. There was sevenfold more viral DNA in the intestine of miceinfected with MVM(1035) than in mice infected with MVMi (85 + 22 pg/,ug versus 12 + 8
pg/,ig).
The kidneys of miceinfected with MVM(1035) had three times more viral DNA than the kidneys of mice infected with MVMi(65 ± 34 pg/,ug versus 22 ± 16 pg/,ug). The spleens of mice infected with MVM(1035) and MVMi contained similar amounts of MVM DNA (170 ± 67 pg/,ug and150pg/jug, respectively).DISCUSSION
Thefailure of MVMp toreplicate in capillary endothelium appeared tobe the basis of its avirulence. MVMi, which is uniformly fatalin neonatal C3H/He mice,infects several cell types,butthe lethal effectsprobably result from itstropism for capillary endothelium (5). Specifically, MVMi causes bilateral infarcts of the solitary renal papilli in association withselective virus replication in the capillaryendothelium of the renal medulla. Although it has notbeen established thatMVMi infection ofmedullary capillaries induces infarc-tion or that the infarcts are the cause ofdeath, this scenario seemslikely. MVMp infected some cells in the renal medulla but fewer than were infectedbyMVMi, and they appeared
tobefibroblasts rather than endothelium.
MVMp spread and replicated systemically, which con-trastswith thefindings of Kimsey and coworkers(13).They
reported that MVMp remained confined to the nasal-oral cavity of perinatal BALB/c mice after intranasalinoculation,
asjudgedbywhole-bodyinsituhybridization, and foundno evidenceofvirus replication. Thesecontrasting results may have been due to differences in virus passage histories and mousestrainsorin the relative sensitivities of the detection methods employed. The presence of intranuclear MVM DNA and capsid antigens inmultiple organs and infectious virus in these organs attiters above the level of viremia were proof that MVMp did spread andreplicate systemically.
Organ-specificinfectivitytiters ofMVMpand MVMiwere notwell correlatedwith the densityof cellsin those organs with nuclei that bound the MVM probe. In organs which were equallypermissive for infectious MVMi and MVMp, suchasthe heart,lungs, andkidneys, thereweremanymore labeled cells in MVMi-infected than in MVMp-infected
mice. We previously encountered a similar lackof correla-tion betweeninfectivity titers and thenumber of cells labeled by in situhybridization in neonatal C3H and C57BL/6 mice infected with MVMi (5). In that system, there is parity of organ-specific infectivity titers between mouse strains, but C3Hmice have farmorelabeled cells thanC57BL/6mice
by
in situ hybridization. We have evidence that the lack of correlation in that system stems from host-mediated differ-encesin theproportion of total viral DNAthat is presentin infectious virions (12a). A similar mechanism driven
by
differences in the viral rather than host genomecouldexplain
thedisparity in this study. VOL.66, 1992
on November 9, 2019 by guest
http://jvi.asm.org/
3122 BROWNSTEIN ET AL.
4
-4% .;ru
,ats
*
-.+
**
,t 4
4
(
.e
SA-., M '.
T
1.r .WRe
A
@CSs*
4
..
W'
#t
,*,, . ... i
.*
;*'*'I s
C.
A .. ..WX
B
D
FIG. 3. Insitu hybridization on PID 5 ofrepresentativeliver sections of mice infected with MVMi(A),MVMp(B),MVM(1035) (C),and MVMi(NS2-1990) (D).Reversedimagesofdark-field illumination. x400.
MVMp doesnothavetropismsforlymphocytes(24, 26)or hematopoietic cells (22) in vitro, but infected cells were common in mesenteric lymph nodes and uncommon in
hepatic hematopoieticfoci in vivo. Therewerefewer virus-positive cells in mesentericlymphnodes ofMVMp-infected
mice than in MVMi-infected mice, but the distribution and patternof nuclearlabeling by the MVM probe and of nuclear staining for capsid antigens were indistinguishable. The
possibilitywasconsidered that the presence ofMVMpDNA
andcapsid antigens in lymph nodes represented sequestra-tion of viral products originating from sites ofreplicationin the intestine. The nuclear pattern oflabeling and staining,
however, suggested that this was replicating virus rather than sequestration, which would have been cytoplasmic.
Alternatively, virus may have replicated in stromal cells of the lymph nodes, although morphologically, the cells
ap-peared to belymphocytes. One further possibilityand one thatissupported by the apparent limited presence ofMVMp
Vr
4s
[image:5.612.134.496.74.358.2]FIG. 4. In situhybridizationonPID5 of small intestine of mouse infected with MVMp. Labeled cells are confined to the lamina propria. Hematoxylin and eosin. x400.
FIG. 5. InsituhybridizationonPID 5of liver ofmouseinfected with MVMp. A labeled cell(arrow) is amonghematopoieticcells. Hematoxylin and eosin. x650.
J. VIROL.
.. $
F
I *
;-A ....
*#
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.68.300.515.691.2]VOL.66,1992~~~~~~~~~~~MVMINFECTION OF MICE 3123
44
FIG. 6. In situhybridizationonPID 5 ofmesentericlymphnode of mouse infected with MVMp. Labeled nuclei are scattered throughoutthe node. Hematoxylin and eosin. x400.
in
hepatic hematopoietic
cells is thatspontaneoushost-range
mutants
developed
during
the limitedcycles
ofMVMp
replication
in the gut whichtargeted
cells notnormally
permissive
for theparentvirus. Futurestudies shouldexam-inethe
phenotypes
andgenotypesof virus thatisrecovered fromlymph
nodes and liver ofMVMp-infected
mice.Phenotypically,
MVM(1035)
resembled MVMi. It causeda lethal infection that was associated with renal
papillary
infarcts,it
replicated
incapillary
endothelium,including
that of the renalmedulla, and itreplicated extensively
inhema-topoietic
cells. Thepathogenicity
of MVMi was therefore determinedby
thatsegment of the genome between nucleo-tides 3522 and 4342(820 nucleotides)
which contains 35 nucleotidechanges
relativetoMVMp.
Tenofthesechanges
alter encoded amino acids
(2, 12).
Thisregion
encodesoverlapping
sequences of VPI and VP2 and contains theallotropic
determinant(1,
12).
Therefore, in addition to thelymphotropic
determinant, this segment of the MVMige-.*
4-.41-,.I.mhaik..i '. I
.., j
. 0
Ai
t
I
..lm
4.
4
.4
* "
444j
'V
FIG. 7. InsituhybridizationonPID5 of renal medulla ofmouse
infected with MVM(1035). Capillaryendothelial cells are labeled;
several capillary lumens are shown (arrows). Hematoxylin and eosin. x650.
nome must also determine the
tropisms
forendothelium(5)
and
hematopoietic
cells(5)
or access of the virus to thesetarget cells. It
apparently
does not determine the level ofviremia,
however, becauseMVM(1035)-infected
mice did nothaveadetectableviremia,
whereasMVMi-infected mice did.Contained within the
MVMp background
ofMVM(1035)
were sequences that facilitated the
synthesis
of infectious virus and viral DNA relative to MVMi. This may have resulted fromaugmented
synthesis
within MVMi host cellsor abroader hostrange. Therewasevidence intwoorgans, brain and
intestine,
that the host range ofMVM(1035)
wasbroader than that of MVMi. In the
brain,
in addition toendothelium,
MVM(1035)
replicated
in externalgerminal
cells of the cerebellum. We didnot
previously
seeevidence of MVMireplication
inexternalgerminal
cells(5)
nor werethey
targets ofMVMp.
In theintestine,
smooth muscle nuclei were common sites ofMVM(1035) replication.
Smooth muscle cellswere not
detectably
infectedby
MVMi(5)
orMVMp.
In organs inwhich nochange
in host rangewas seen,
however,,
it seemedlikely
thathigher
titers re-sulted fromaugmented synthesis
in MVMi target cells. Within theMVMp background
ofMVM(1035)
were 128 nucleotide differences withMVMi,
of which 17 encodedchanges
in amino acids(2).
Also contained in theMVMp
background
was a65-bp
directrepeat between nucleotides 4720 and4784,
absent inMVMi,
that contained apolyade.
nylation signal
for alltranscripts
(2).
Thissignal
has been shown not tobe usedby MVMp,,
however(6).
NS2 was
required
for normal MVMireplication
andpathogenicity.
No infectious virus was recovered fromor-gans of mice infected with
MVMi(NS2-1990)
despite
evi-dence of limitedspread
andreplication.
MVMi(NS2-1990)
produces
less infectious virus than doeswild-type
virus when growninmousecells andis deficient in thesynthesis
ofmonomer
replicative
forms and progenysingle
strands(5a).
It isnot known whether the absence of detectable infectious viruswasthe result of detection
limits,
whichwereabout 100ID50/g
oftissue,
or of the failure of the virus to make infectiousparticles despite
dissemination. In either case,infectivity
titers of the NS2 mutant were at least102_
to104-fold lower than those of the parent strain in all organs, andthe number of infectiouscenterswas
greatly
reduced. A similar result has beenreported
for neonatal rats infected withH-i virus mutatedatthe sameR2large
splice
acceptorsite as
MVMi(NS2-1990)
and which was also unable tosynthesize
any NS2 isoforms(16).
In thatstudy,
however,noevidence of virus
spread
orreplication
waspresented,
in contrast toresultspresented
here.The molecular bases of differences in
parvoviral
virulenceare
beginning
to be examinedby
using
the MVM-mouse modelsystem. Thissystemisattractive notonly
becausesomuch is known about the molecular
genetics
of MVM but also because thegenotypeof the hostplays
animportant
role in MVMpathogenesis
(5).
Becausemoreis known aboutthe genome of themousethan ofany other mammalianspecies,
it affordsan
opportunity
tomanipulate
the hostaswellasthe virus genome in ordertodissectparvoviral
virulence mech-anisms.ACKNOWLEDGMENTS
This research was supported by Public Health Service grants
RR00393,
RO1AI21302, K04A100934, andCA29303.We thank Deborah Winograd, Al-Karim Rehemtula, and Ruth
Peeplesfortechnicalsupport. VOL.66, 1992
1
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.59.298.74.255.2] [image:6.612.57.297.508.685.2]REFERENCES
1. Antonietti, J. P., R.Sahli, P. Beard, and B. Hirt.1988. Charac-terization of the celltype-specificdeterminant in the genomeof minute virus of mice. J. Virol. 62:552-557.
2. Astell, C. R., E. M. Gardiner, and P. Tattersall. 1986. DNA sequence ofthelymphotropicvariantof minute virus ofmice, MVM(i), andcomparison with the DNA sequence of the fibro-tropic prototype strain. J. Virol. 57:656-669.
3. Berns, K. I. 1990. Parvovirus replication. Microbiol. Rev. 54:316-329.
4. Bonnard, G. D., E. K. Manders, D. A. Campbell, R. B. Herber-man, and M. J. Collins. 1976.Immunosuppressive activity of a subline of themouseEL4lymphoma.Evidence for minute virus of mice causing theinhibition.J. Exp. Med. 143:187-205. 5. Brownstein, D. G., A. L. Smith, R.0.Jacoby, E. A.Johnson,G.
Hansen, and P. Tattersall.1991.Pathogenesis of infection with a virulent allotropic variant of minute virus of mice andregulation byhostgenotype. Lab. Invest. 65:357-364.
5a.Cater, J. E., and D. J. Pintel.Submitted forpublication. 6. Clemens, K., and D. J. Pintel. 1987. MVM mRNAs
predomi-nantly polyadenylate at asinglesite.Virology 160:511-514. 7. Cotmore, S. F., L. J. Sturzenbecker, and P. Tattersall. 1983. The
autonomous parvovirusMVM encodes two nonstructural pro-teins in addition to its capsid polypeptides. Virology 129:333-343.
8. Cotmore, S. F., and P. Tattersall. 1986. Organization of the nonstructural genesof the autonomous parvovirus minute virus of mice. J. Virol. 58:724-732.
9. Cotmore, S. F., and P. Tattersall. 1987. The autonomously replicatingparvoviruses of vertebrates. Adv. Virus Res. 33:91-174.
10. Doerig, C., H. Bernhard, J. P. Antonietti, and P. Beard. 1990. Nonstructural protein of parvovirus B19 and minute virus of mice controlstranscription.J.Virol. 64:387-396.
11. Doerig, C., B. Hirt, P. Beard, and J. P. Antonietti.1988. Minute virus of mice nonstructural protein NS-1 is necessary and sufficient for trans-activation of the viral P39 promoter. J. Gen. Virol.69:2563-2573.
12. Gardiner, E. M., and P. Tattersall. 1988. Mapping of the fibrotropic and lymphotropic host range determinants of the parvovirusminute virus of mice. J. Virol. 62:2605-2613. 12a.Kapil, S., and D. Brownstein. Unpublished data.
13. Kimsey, P. B., H. D. Engers, B. Hirt, and C. V. Jongeneel. 1986. Pathogenicity of fibroblast- andlymphocyte-specificvariants of minute virusofmice. J. Virol. 59:8-13.
14. Krieg, P., E. Amtmann, and G. Sauer.1983. The simultaneous extraction ofhigh-molecular-weight DNA and RNA from solid
tumors.Anal. Biochem. 134:288-294.
15. Kunkel, T. A. 1985.Rapid and efficient site-directed mutagene-sis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
16. Li, X., andS. L. Rhode III. 1991.NonstructuralproteinNS2 of parvovirus H-1 is required for efficient viral protein synthesis and virus production in rat cells in vivo and in vitro. Virology 184:117-130.
17. McMaster, G. K., P. Beard, H. D. Engers, and B. Hirt. 1981. Characterization of an immunosuppressive parvovirus related tothe minutevirusofmice.J. Virol. 38:317-326.
18. Merchlinsky, M. J., P. J. Tattersall, J. J.Leary,S. F.Cotmore, E. M. Gardiner, and D. C. Ward. 1983. Construction of an infectious molecular clone of the autonomous parvovirus minute virus of mice. J.Virol.47:227-232.
19. Miller, R. A., D.C. Ward, and F. H. Ruddle. 1977.Embryonal carcinoma cells(andtheir somatic cellhybrids)areresistantto
infection by the murine parvovirus MVM, which does infect other teratocarcinoma-derived cell lines. J. Cell. Physiol. 91: 393-402.
20. Naeger, L. K., J. Cater, and D. J. Pintel. 1990. The small nonstructural protein (NS2) ofthe parvovirus minute virus of mice is required for efficient DNA replication and infectious virus production in a cell-type-specific manner. J. Virol. 64: 6166-6175.
21. Rhode, S. L., III. 1985. trans-Activation of parvovirus P38 promoter by the 76K noncapsid protein. J.Virol. 55:886-889. 22. Segovia, J. C., A. Real, J. A. Bueren, and J. M. Almendral. 1991.
Invitromyelosuppressive effects of the parvovirus minute virus of mice(MVM) onhematopoieticstemand committed progen-itor cells. Blood 77:980-988.
23. Smith, A. L. 1983. Response of weanling random-bred mice to
inoculation with minute virus of mice. Lab. Anim. Sci. 33:37-39.
24. Spalholz, B.A., and P. Tattersall. 1983. Interactionof minute virus of mice with differentiated cells: strain-dependent target cell specificity is mediated by intracellular factors. J. Virol. 46:937-943.
25. Tattersall, P. 1972. Replication ofparvovirus minute virus of mice. Dependence of virusmultiplication and plaque formation
oncellgrowth.J. Virol. 10:586-590.
26. Tattersall, P., and J. Bratton. 1983. Reciprocal productive and restrictive virus-cell interactions of immunosuppressive and prototype strains of minute virus of mice. J. Virol.46:944-955. 27. Wolter, S., R. Richards, and R. W. Armentrout. 1980. Cell cycle-dependent replicationof the DNAof minute virus ofmice,
aparvovirus. Biochim.Biophys. Acta 607:420-431.