Copyi-ight j1972 AmericanSociety forMicrobiology Priittedin U.S.A.
Isolation
and Properties of
rex-
Mutants of
Bacteriophage Lambda
GARY N. GUSSIN AND VICTORIA PETERSON
Departmentt of Zoology, University ofIowa, IowaCity,Iowa52240
Received for publication26 May 1972
Twenty-fiverex- mutantsofphagelambda have beenisolated.Three of the
mu-tants, including one amber mutant, map at three distinct sites within the rex
re-gion of the lambdageneticmap.Theexistence of the ambermutantprovides further evidencethatrexandcIaredistinctgenes,sinceit seemstobeidenticaltowild-type lambdainitsabilityto establish ormaintainlysogeny.
Lysogenization of Escherichia coli
by
thetem-perate bacteriophage lambda requires the
pres-ence ofarepressor, the product of thephage cI
gene,whichdirectly (18) orindirectly (8)prevents expression ofalmost all lambdagenes.Twogenes that are expressed are the cI gene itself, since
the repressor must be present continuously to
maintain
thelysogenic
state(8,
20), and therex gene, which is defined by its effect on the
multiplication of rII mutantsofphageT4. T4rII mutants cannot
multiply
inalysogen
foraXrex±
prophage (3),
butrex- mutantscanbe selected onthebasis of their inability to prevent multiplica-tion ofT4rIIphage (14;and R.Friedman,Ph.D.
thesis, Cambridge Univ., 1967).
Since mutantsselected in this way donot seem tobedefective in
anyX
function,
the role ofrexin thelifecycle
ofthe
phage
may beincidental. On the otherhand,
rexand cI are
adjacent
in the Ximmunity
region
(see Fig.
1)
andarecoordinately
expressed
during
infection
(2)
and in thelysogenic
state(9,
10).
These facts suggest that rex may play an im-portant, ifnot always essential, rolein
lysogeny.
In this paper, wedescribe
theisolation of
25 new rex- mutants. Mapping data show that atleast three
of
the mutants arelocatedat distinct siteswithin the rexgene.One of themutantsisanonsense mutant, defined by sensitivity of its Rex7phenotype tothe action of the
SuII+
amber suppressor(13, 19).
The existenceof thismutantprovides further evidence that the rex and
cI
genes are in fact distinct and confirms the
pre-sumption
thatthe rex genespecifies
aprotein.
In the suppressed bacterial host, the nonsense mutation is temperature-sensitive. It is pheno-typically rex+ at temperatures between 25 and
30C, but rex- attemperatures above 35 C.
MATERIALS AND METHODS
Bacteria and bacteriophage. Table 1 lists the
bac-teriophage and bacterialstrains used in thesestudies.
The derivation ofadditional strains is described where necessary.
Media.Tbrothcontains per liter of distilled water: tryptone (Difco), 10 g.; NaCl, 2.5 g.;KCl,2.5 g. For phage assays, bottom agar contains 10 g of B agar (Difco) and top agar contains 6 g of agar per liter of
Tbroth.
Assays. Phage andinfectious centers were assayed byplaque formation using the agar layer method (1). Mutagenesis and isolation of mutants. Cultures of WNQ8 (seeTable 1) were grown at 32 C to a density of about 108 cells/ml in T broth. Nitrosoguanidine (N-methyl-N'-nitro-N-nitrosoguanidine) was added to
aconcentration of 10
,tg/ml
withcontinued aeration for7min. Thecells were then diluted 1:1,000 infreshT broth, and incubation was continued for 3 hr at
32C.
Mutagenized cultures were spread on agar plates together with 105 T4rII1589 phage and incubated overnightat 32C.Nibbledcolonies (initiated by cells sensitive toT4rII infection) were screened by spot-testing for theabilitytosupportT4rII multiplication
and for immunityto superinfection by
XcI6Ob2.
All mutantsreported inthis paper are phenotypicallycI+rex- on thebasis of these tests.
Recovery ofrex-mutations fromWNQ8. The rex-lysogens, WNQ8 (XNsus7 Nsus53 c1857 Od58 rex-),
were superinfected with Ximm434 atacell density of
2 X 108cells/mlandamultiplicity offive phage per cell. Phagewere allowedto adsorb in 0.01 M MgSO4
for 20minatroomtemperature, after which the cells
were diluted 1:10 in T broth and incubated for 60 min at 37 C.N+O+ recombinants having lambda
im-munity (abletoplateon imm434lysogens) werethen screened for their rex- andc1857 phenotypes. Phage with thegenotype cI+rex-wereobtained bycrossing
cIsusl4andrex-c1857 phage.
Infectionof
lysogens
withT4rlI 1589. Strain594anditslysogens ofrex- mutants weregrownto a density of2 X 108cells/mlin T broth.To one ml ofculture
were added: NaN3, 0.002 M; tryptophan, 40
jAg/ml;
and T4rII1589 or wild-type T4 to a multiplicity of two tofive phage per cell. After 10 min at 32 C for
phageadsorption, T4 antiserumwasaddedto afinal K value of
0.5/'min
(1); thecultures were incubated760
on November 10, 2019 by guest
http://jvi.asm.org/
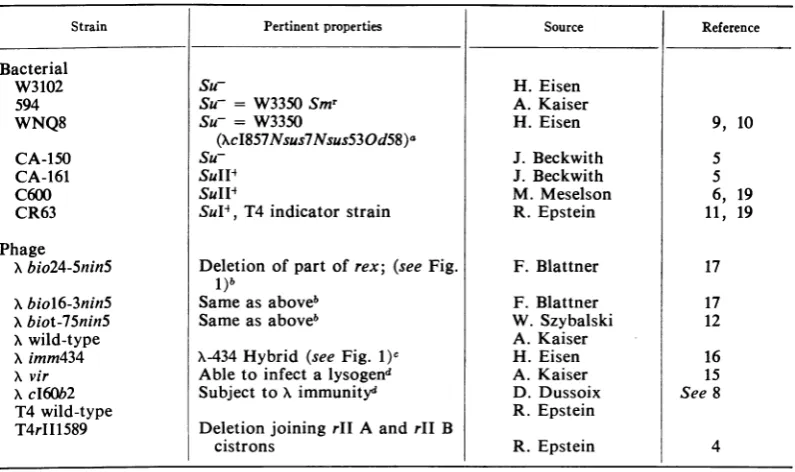
TABLE 1. Bacteria and phage strains
Strain Pertinent properties Source Reference
Bacterial
W3102 Su- H. Eisen
594 Su- = W3350Smr A. Kaiser
WNQ8 Su- = W3350 H. Eisen 9, 10
(XcI857Nsus7Nsus53Od58)a
CA-150 Su- J. Beckwith 5
CA-161 SuIIT J. Beckwith 5
C600 SuII4 M. Meselson 6, 19
CR63 SulP, T4 indicator strain R.Epstein 11, 19
Phage
Xbio24-Snin5 Deletion ofpart of rex; (see Fig. F. Blattner 17 1)b
Xbiol6-3nin5 Sameas aboveb F. Blattner 17
Xbiot-75nin5 Sameas aboveb W. Szybalski 12
Xwild-type A. Kaiser
Ximm434 X-434Hybrid (see Fig. 1)c H. Eisen 16
Xvir Ableto infectalysogend A. Kaiser 15
XcI60b2 Subject toXimmunity" D. Dussoix See 8
T4wild-type R. Epstein
T4rII1589 Deletionjoining rIIA and rII B
cistrons R. Epstein 4
a
c1857
isatemperature-sensitive
mutation incI (20);thetermsusis used to indicateis suppressor-sensitive (6).
bnin5 is amutation that enables bio
transducing
phage toformplaques (7).cThe
hybrid
wasisolated by Tomizawa.dUsed to test
presumptive lysogens.
for anadditional 10 min, diluted into T brothby a
factor of 105, and assayedimmediatelyfor infectious
centersand survivingbacteria. After 90 min of incu-bation with aerationat32C, the diluted cultureswere
assayed for progenyphageonCR63.
Phageadsorptionunder theseconditions was very poor. On the average, only about 75% of the cells
were actually infected, and in most cases less than half theinfectingphagewereadsorbed.Wecouldnot
enhancethese percentages byconcentrating the cells before adsorption, allowinggreater time for
adsorp-tion, changing the growth medium, or by omitting NaN3. With bacterial strains other than 594, there
was even less adsorption. We were unable to select
phage strains that adsorbedmoreefficiently.
Mapping rex- mutations. Recombinants were
selected incrossesbetween Xrex-cI857 and Xbio ninS
deletion mutants. Phage were added to W3102 in 0.01 MMgSO4at acell densityof about2 X 108/ml
andaphagemultiplicityof five of eachparent. After
20minat roomtemperature for adsorption, infected cells were diluted 1:10into T broth at 37 C. After
60min, progenyphagewereplatedonW3102 at 42C. Turbid plaques, formed by recombinants arising
from an exchange betweenc1857 andtheendof the biodeletion,wereidentified inthebackgroundofclear plaques. (The bio nin5 parents formclearplaques
be-cause of the deletion of the cIII gene.) Lysogens
formedby each recombinant were isolated and spot-tested for their rex genotype to determine the fre-quencyof rex+phageamong theturbidrecombinants.
that amutation
Heatinducibilityof rex-c1857 lysogens.Theeffect
ofrex- mutations on the heat inducibility ofc1857
lysogens was tested by incubating W3102 lysogens for
15 min at the indicated temperatures. After incuba-tion, a 0.1-ml portion was added directly to W3102 plating bacteria and plated without preadsorption. Plates were incubated at 32 C, and the number of plaques was taken to be the number ofcellsinduced during the 15-mn incubation period at the indicated temperature.
Materials. T4 antiserum having a K value of 600/min was purchased from Gateway Immunosera Co., Cahokia, Ill. (In some experiments, antiserum provided by F.
Eiserling
wasused.) Nitrosoguanidinewaspurchased from Aldrich Chemical Co., Milwaukee,
Wis.
RESULTS
Properties
ofmutants.Table
2 lists the burst sizesobtained
in one-stepgrowth
ofT4rII1589
and
wild-type
phage
after infectionof lysogens ofthree of the rex- mutants isolated according to
the
procedure
described above. Ineachcase, theT4rIIphageyield on infection of a rex- lysogen is
comparable
to theyield
on the correspondingnonlysogenic
strain and is in marked contrast totheT4rII
phage
yield
oninfection of thewild-type(rex+)
lysogen.
The percentage of cells thatscored
as infectious centers,indicating
theon November 10, 2019 by guest
http://jvi.asm.org/
TABLE 2. Inifection of rex lysogenis by T4rIHJ589
Percent
infectious Phageyield/cella
centersafter after infection by Infectedstrain infection by
T4 T4rI1589 T4 T4rII1589
594 Nonlysogenic 87 62 134 122
594 (X rexQ) 92 59 109 134
594 (X rex5a) 95 75 110 173
594 (X rex30a) 79 50 100| 107
594 (X rex-) 75 0.5 104 2.6 X 10-'
a Phageyield per cell to which phage was added. The phage yield perinfectious center can be ob-tained by dividing by the corresponding value for the percentage ofinfectious centers given in the firsttwo columns.
I In other experiments with 594 (X rex30a), this percentage was as high as 94; the low value observed in this experiment is therefore a
re-flection of poor phage adsorption rather than of restriction of T4rlI1589 multiplication. Multi-plicities ofinfection: T4, 4.0;T4rI1589,2.3.
duction ofatleast oneprogeny
phage particle
in the initiallyinfected
cell, was in each casecom-parable
to the percentage obtained oninfection
of the
corresponding nonlysogen.
Thus,by
these two criteria, there is no residual rex functionexpressed
inlysogens
of thethreemutantstested.Spottests
of
T4rII1589onC600lysogens
of the25 rex- mutants isolated in these studies indi-cated that
only
oneof
them, rexQ, was suppres-sible by the Sull amber suppressor contained in C600. Quantitative estimation ofT4 burst sizesin
SuII+
lysogens of
rexQ prophage is difficultbecause
noneof
theSuII+
bacterial strains wehave tested
adsorbs
T4efficiently.
Nevertheless, the results ofsingle-burst
experiments in the SuII+strain
CA-161 and in theclosely related
(but
notprecisely isogenic)
Su- strain CA-150clearly demonstrate
that rexQ is an ambermuta-tion
(Table
3).
At 26 C in strain CA-161, it isphenotypically
rex4,
effectively preventing T4rII multiplication; butatthe same temperature, it is rex- intheunsuppressed strain CA-150,
in which T4rII multiplication is normal.In addition to its suppressor-sensitivity, the rexQ mutation is temperature sensitive when suppressed
(Table
3).
At 41C,
itis rex- even in CA-161. These resultsareconfirmed by assays of relative efficiencies of T4rII plaque formation at 30, 35, and 40 C in rex lysogens (Table 4). NotethatrexQis phenotypically rex+ only at 30 C in
the
SuII+
strain CA-161. Since temperaturesensitivity
isobserved in both C600 and CA-161lysogens, it is most likely due to some property of
the amber suppressor Sull and not to physio-logical properties of the strains used. One possi-bility is that the suppressed rexQ protein,
con-taining glutamineat the site of the amber mutation
(19), is itself temperature sensitive. Alternatively, T4rIImultiplication may be differentiallyaffected at high and low temperatures by the reduced
levels of rex protein synthesized in suppressed strains, since the
frequency
ofsuppression
by SuIIis less than30%- (13, 19).TABLE 3. Phageyields of T4rIlI589oniilnfectionz of
SulI+ and Sut- lysogens of X rexQ at 26anid41C
Phageyield/celF
SuIII+ after infectionat Bacterial S strain allele
26C 41C
CA-161 SulIt 24 17
CA-150 Su'- 23 20
CA-161 (X) SulI+ 0.02 0.1
CA-S50 (X) Si- 0.004 0.01
CA-161 (X rexQ) SuIlI 0.1 12
CA-150 (X rexQ) Su-w 12 21
aPhage yield per cell to which phage were
added. Experimentswereperformedasdescribed,
except that cells were grown and infected at the indicated temperatures and that cells infected at 26C were incubated for 135 min after infection before assaying. Phage yields are low because of poorphageadsorptionandaconsequent low per-centage of infectious centers. For CA-161, the frequencies of infectious centers were
31%'/^
(26 C) and 39% (41 C);for CA-150,thefrequencies were26% (26 C) and
29%
(41 C).TABLE 4. Relative efficiencies of platintg of T4rII589 on SuII+ andSu lysogelis of
X rexQ at various temperatures
Tempb(C) Bacterial strain allele
30 35 40
CA-161 SuII+ 1i.a 1.2 1.2
CA-150 Su- 0.7 1.1 1.1
CA-161 (X rexQ) SuII+ 0.003 1.0 1.2
CA-150 (X rexQ) Su- 0.95 1.2 1.2
a Relative efficiencies of plating are normalized
to a value of 1.0 for the number ofplaques ob-tained on plating T4rII1589 on CA-161 at 30 C. Efficiencies of plating of wild-type phage, nor-malized inthesameway, ranged from 0.95 to 1.2. bPlating bacteria were grown at the indicated temperature.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.495.270.461.467.589.2]rex- MUTANTS OF BACTERIOPHAGE LAMBDA
Mapping of rex- mutations. The three
rex-mutants used inthese experiments were mapped with Xbio transducing phage in which the bio
"deletions" had been shown to extend into the
rex region, but not beyond it into cI (17; W.
Szybalski, rersonal communication). Turbid
re-combinantsfromcrossesofthetype Xbio ninS X
Xrex-cI857 (seeTable 1andFig. 1) werescreened
for their rex genotype. If no rex+ turbid
re-combinants could be recovered from a cross, it
was concludedthatthe bio deletion included the
particular rex- site. The three deletion mutants
used define fourregions of therex gene (Fig. 1)
and the results of nine crosses (Table 5) define the following map order: bio24-5 end point,
rex5a,biol6-3endpoint, rexQ, biot-75endpoint,
rex30a.
In asimilar cross, the rex- mutant (go293) of
Friedmanwasconcludedtobe in thesameregion (region II, Fig. 1) asrex5a (17).
We have placed rex30a to the left ofcIsusl4
because the evidence is convincing that cI and rex are distinct genes. In a three-factor cross, XcIsusl4 X Xrex3OacI857, 13 of 14 cI+
recombi-nants were rex-.These data are consistent with, but do not prove,the assumptionthat rex30a is
tothe left ofcIsus14.
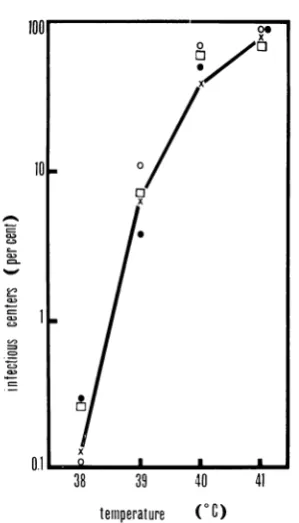
Effects ofrex-mutationsonlysogeny.The
pos-sibility that rex- mutants are affected in their
ability to maintain lysogeny was investigated by
examining
theefficiency of
heatinduction
ofrex-c857
lysogens during a 15-min incubation atvarioustemperatures. The results of these experi-ments (Fig. 2) demonstrate that the freouencies of heat
induction
are similar, if not identical, in lysogens ofrex+cI857
and rex-cI857 phage. The small differences observed are no greater than thoseobserved with separate isolates of the same rex+cI857 lysogen.Thus,within the limits of sensitivity of this kind
of
assay, therexlocus
doesnotaffect the thermal stability of lysogens of a temperature-sensitive prophage.Studies offreouencies of lysogenization yieldsimilar results; that is, no difference betweenTABLE 5.
Fractionz
ofX rex+plhage
amontg
tuirbid
recombinianits
in crosses ofthe typeX bio1,1,15 X X rex-c1857
rex-Parent bioParent
rex5acI857 XrexQcI857 rex3OacI857
X
bio24-51in5
3/19a 8/38 19/26Xbio16-3nin5 0/30 6/25 14/32
X
biot-75nin5
0/25 0/27 12/26aFor each entry, the denominator indicates
the number of turbid recombinants examined (see Materials and Methods) and the numerator indicates the number of recombinants that were
rex+.
i434
att int xis exo1
la
1Ciii
I N Isex V21V
rexIXCl
IV3I|xXI
y Ci' O I P~ I
I'III
- IN sex V2 reX5* rexQ rex,3a CISUS14
C1857
CISUS34I 1
1-5
III
Vi V3 x
t7> bio 16-3l
2?
bio
t75FIG. 1. Maplocationof rexregionz oflambda. Thlerexgenecani be subdivided inttofour regionls (I-IV) based oltthepositiollsoftheriglht-hanid en2dpointsofthebiodeletions,allofwhich haveastheirleftenldpointttheceniter ofatt. Thebounldariesbetweeniv2anidrex,betweenirexandcI, and betweent vlv3and cIarentotpreciselydefined.
The v2mutation is in the operator towhich repressorbinds to block transcripti9ni leftwardfrom sex; vlv3are
mutationsin the operatortowhich repressorbindstoblock transcriptionrighltward fromx. Theimmuniity
regiont
isdefined by theregioniofnzonhomologybetweenXanid 434 (indicatedby j434). Tlhe cIsusmutantts wereobtainied
from M. Lieb. Herunpublisheddata andoursindicate thatcIsusl4isfarthest to theleftintcl, anid thatcisus34
isfarthesttotheright.Att istheregioninwhich recombinationbetweentthebacterial chromosome anid thelambda
deoxyribonucleic acidmoleculeoccursduringinitegrationoftheprophage. ciandl cIIIarerequiired forthe estab-lishment oflysogeny,butntot foritsmainteniance. (See reference 8.)
I*I
_-b10 24
763
VOL. 10, 1972
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.495.247.440.236.332.2] [image:4.495.50.438.393.563.2]GUSSIN AND PETERSON
38 39 40 41
temiperature
(°C)
FIG. 2. Heat-inducibility ofrex-cI857 lysogens at various temperatures. Cells heat-induced during a
15-min incubationperiodatthe indicatedtemperatures
were assayedasinfectiouscentersby plaque formation
at 32 C on C600. Symbols: X, iniduction of W3102
(XcI857); 0, induction of W3102 (XrexQcI857); 0, inductioni of W3102
(Xrex5acl857);
Z, induction of W3102 (Xrex30acI857). The lihle connects the pointsfor the induction ofthe XcI857 lysogen.Sam-plinig errorat39, 40, and 41 C isapproximately
indi-cated by the sizeofthe symbols used;at 38C, error is greater because the nlumber of plaques was small.
efficiency of lysogenization by rex+ and rex-phageis observed.
DISCUSSION
The fact that the amber mutant rexQ is
phenotypically cI clarifies several points about
the relation between cI and rex. First, together
withthefindingthatatleastonecIambermutant
retains normalrex function (2), the existence of
rexQ demonstrates unambiguously that rex and cI are distinct genes. These findings support
Kayajanian's similar conclusion based on the existence of bio deletionmutantsthatarerex-c±+.
Second, rex cannot be essential to
lysogeniza-tion underconditionsnormally used in studiesof lambda. Because rex- mutants selected for their
inability to block T4rII multiplication need not
be defective in the lambda-specific function of rex, onlyanonsense mutant canbe presumedto
be truly rex- in terms of the role ofrex in the lambdalifecycle.
Finally, if an interaction between rex protein
and therepressor occurs, it cannotbeobligatory either for repression or for the prevention of
T4rIImultiplication.
The possibilitythat the rexand cI genes were
not distinct, or that they specified two poly-peptides that interacted in some way, was first
inferred from experiments of Tomizawa and
Ogawa (21). In these experiments, incubation of
cultures at 47.5 C simultaneously inactivated a
temperature-sensitive (cIts1-1) repressor and
abolished normal rex function. Recently, how-ever,K-K. Mark (Szybalski, personal
communica-tion; Mark and Szybalski, manuscriptin
prepara-tion) has found that the effect on rex function
depends on the abnormally high temperature usedandnotonthe inactivation of therepressor.
According to Mark's results, inactivation of the
cltsl-1 repressor at 42 C does not affect rex
function.
None of theseresultsprovidesacluetotheway
inwhich rex protein blocks themultiplicationof
T4rII phage. Thefact that rexQ is temperature-sensitive in the permissive host CA-161 offers a
wayofinvestigating this question inmore detail.
ACKNOWLEDGMENTS
This investigation was supported by Public Health Service research grant l-ROI-AI-10019 from the National Institute of Allergy and Infectious Diseases and by Public Health Service Biomedical Sciencessupport grant FR-07035 from the Division ofResearch Facilities and Resources.
Wearegrateful to H.Eisen,A. Kaiser, F. Blattner,andW. Szybalski for providingsomeofthephageandbacterialstrains used in thesestudies, andto L. Astrachan and J. F. Miller for communication of some of their results prior to publication.
LITERATURE CITED
1.Adams,M. H.1959.Bacteriophages. Interscience Publishers Inc.,NewYork.
2. Astrachan, L.,andJ. F. Miller. 1972. Regulation ofXre.
expressionafter infection of EscherichiacoliKby lambda bacteriophage. J. Virol.9:510-518.
3. Benzer, S. 1955. Finestructureofagenetic regionin bac-teriophage.Proc. Nat. Acad. Sci. U.S.A. 41:344-354. 4. Benzer, S., and S.P. Champe. 1962. A change from
non-senseto sensein thegeneticcode. Proc. Nat. Acad.Sci. U.S.A.48:1114-1121.
5. Brenner, S., and J.R.Beckwith. 1965.Oclhremutants,a new
classofsuppressiblenonsensemutants.J. Mol. Biol. 13:629--637.
6.Campbell, A. 1961. Sensitive mutants of bacteriophage X. Virology 14:22-32.
7.Court, D., and K.Sato. 1969.Studiesof novel transducing variantsoflambda: dispensability of genes N and Q. Virology39:348-352.
8. Echols, H. 1971.Lysogeny: viralrepression and site-specific recombination. Annu. Rev.Biochem. 40:827-854. 9. Eisen, H., L. Pereira daSilva,and F. Jacob. 1968. Sur la
regulation precoce du bacteriophage X. C.R. Acad. Sci. 266:1176-1178.
10. Eisen, H.,L.PereiradaSilva,and F.Jacob.1968.The
regu-lation andmechanismof DNAsynthesisinbacteriophage X.ColdSpringHarborSymp. Quant.Biol. 33:755-764. 11. Epstein, R. H., A.Bolle, C. M.Steinberg, E. Kellenberger,
764
J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.495.85.235.71.333.2]
rex-E.Boy de la Tour, R. Chevalley, R. S. Edgar, M.Susman,
G. H. Denhardt, and A. Lielausis. 1963. Physiological studies of conditional lethal mutants of bacteriophage T4D.Cold Spring Harbor Symp. Quant. Biol. 28:375-394. 12. Fuerst, C. R. 1966. Defective biotin-transducingmutantsof
bacteriophage lambda. Virology 30:581-583.
13. Garen, A. 1968. Sense and nonsense in the genetic code.
Science169:149-159.
14. Howard, B. D. 1967. Phage lambda mutants deficient in
rllexclusion.Science 158:1588-1589.
15. Jacob, F., and E. L. Wollman. 1954. Etude genetique d'un bacteriophage tempere d'E;cherichia coli. I. Le syst6me
genetique du bacteriophage X. Ann.Inst. Pasteur 87:653-673.
16. Kaiser, A. D., and F. Jacob. 1957. Recombination between relatedtemperate bacteriophages andthe genetic control
ofimmunity and prophage localization. Virology 4:509-521.
17. Kayajanian, G.1970. Deletion mapping of theclIl-cIl region
ofbacteriophageX.Virology 41:170-174.
18. Ptashne, M., and N. Hopkins. 1968. The operators
con-trolled by the X phage repressor. Proc. Nat. Acad. Sci. U.S.A.69:1282-1287.
19. Stretton, A.0.W., S.Kaplan, and S. Brenner. 1966.
Non-sense Codons. Cold Spring Harbor Symp. Quant. Biol. 31:173-179.
20. Sussman, R., and F. Jacob. 1962. Sur un systeme de
re-pression thermosensible chez lebacteriophaged'Escherichia coli.C. R. Acad. Sci. 254:1517-1519.
21. Tomizawa, J., and T. Ogawa. 1967. Inhibition of growth of rlImutants ofbacteriophage T4 by immunity substance ofbacteriophage lambda. J. Mol. Biol. 23:277-280.