Copyright _©1972 AmericanSociety for Microbiology Printedin U.S.A.
Methylation
Pattern of Lambda
Deoxyribonucleic Acid
CECILIA HIDALGO' AND HOWARD A. NASH
LaboratoryofNeurochemistry, National Institute of Mental Health, Bethesda, Maryland 20014
Receivedforpublication 14July 1972
Deoxyribonucleic acid
(DNA)
extracted from phage X grown on Escherichia coliK-12strain W4032 had 113 i 105-methylcytosine residues and215 i 206-methyl adenine residues per genome, asdetermined
by
threeindependent methods.Thesemethylated nucleotidesweredistributed equallyamong thetwo strandsofX DNA.
Shearing ofdouble-stranded DNA tohalf-length fragments revealed a
slight
defi-ciencyof 5-methyl cytosine in the55%guanineplus cytosine half. ShearingtheDNAtofragments ofsmaller
length
showed that thedistribution ofmethylated nucleotides along the double helixwasuniform with the exception ofanundermethylated frag-ment arising from the center ofthe X DNA molecule. The implication of theseresults for the function ofmethylated nucleotides in the X DNAmolecule is dis-cussed.
Methylated
bases havebeenfoundtobe presentin
deoxyribonucleic
acid(DNA)
frombacteria,
phages,and animal andplantcells. A small
frac-tion ofthe
methyl
groupspresentin bacterial andphage DNA have been
implicated
inhost-con-trolled restriction and modification
(1).
Nobio-logical
role hasyet been found for therestof themethylated
bases present in the DNA molecule. In anattempttoinfer therole of these othermeth-ylated bases, we determined the distribution of
methylated
nucleotides in the X DNAmolecule.DNA from
phage
Xwaschosen because itsgenet-ics have been
extensively
studied,
thephysical
localization
of most of its genes on the DNAmolecule is
known,
andtechniques
exist for thephysical
resolution of differentgenetic regions
as well as theseparation
ofsingle
strands of thephagegenome
(6, 10).
We found that X DNA extracted from
phage
grown in a K-12derivative of Escherichiacolicon-tainsboth
5-methylcytosine
(5MG)
and6-methyl-aminopurine
(6MAP),
in agreementwithprevious
studies(5,
9).
Inaddition,
wehavedemonstrated that both strands have thesamemethyl
content,and, along
the doublehelix,
only
asmallfragment
corresponding
to aregion
in the center of the linear genetic map shows a different degree ofmethylationfrom therest.
MATERIALS AND METHODS
Bacterial and phage strains. E. coli K-12 strain W4032 (HfrC, lac A [complete], pro-, metA, T6R,
1Presentaddress: Facultad de Quimica y Farmacia, Uni-versidaddeChile, Vicuiia Mackenna20,Santiago,Chile.
described by Cook andLederberg [3]),wasobtained from R. Perlman. E. coli K-12 C600 was obtained from M. Gottesman. Thelambdaphage strains used were XcI857andXb2c1857.
Media and reagents. ModifiedTPA medium con-tained, per liter: NaCl, 0.5 g; KCI, 8.0 g; NH4Cl, 1.1 g; tris(hydroxymethyl)aminomethane (Tris), 12.1 g;KH2PO4,0.46 g;sodiumpyruvate,0.8 g; 19 L-amino acids, 150 mg each; L-methionine, 10 mg; 0.16 M Na2SO4,1.0ml;andadenine, 10 mg. Thefinal pH was 7.4, adjusted with HCl. Tris-Mg buffer was 0.015 M Tris-hydrochlorideand 0.01MMgSO4,pH 7.4.
Electrophoretically pure pancreatic deoxyribonu-cleasewasobtainedfromSigmaChemical Co. Snake venom phosphodiesterasewasobtained from Worth-ington Biochemical Corp.['H-methyl]-methionineand
[14C-methyl]-methionine were obtained from New EnglandNuclear Corp., Amersham-Searle, or Schwarz BioResearch, Inc. Specific activities of ['H-methyl]-methionine ranged from 2.6 to 7.6 Ci/mmole. [14C-methyl]-methionine specificactivities ranged from 12.8 to60mCi/mmole.Carrier-free32Pwasobtained from NewEnglandNuclear Corp.
DNApreparation. To obtain XDNA labeled either with [14C-methyl]-methionine or with "2P and [3H-methyl]-methionine,Xphagewasgrown onW4032 in modified TPA medium. Cells wereallowedtogrowin modified TPA mediumat37 C to a concentration of 3 X 108cells/ml, centrifuged,andresuspendedin 0.1 volume ofTris-Mgbuffer. The phage was added at a multiplicityof infection of1 to 3,andadsorption was allowedtoproceedfor 15minat 37 C. At thispoint, theoriginal volumewasrestoredby adding fresh modi-fied TPA medium with the radioactive isotopes, and the culturewas further incubated for 2.5 hr at 37 C withshaking. Chloroform was added to 1% volume, and,after shaking for 10moremin,the bacterialdebris wasremovedbycentrifugation and the clear lysate was
937
on November 10, 2019 by guest
http://jvi.asm.org/
HIDALGO AND NASH
treated with 10 g of polyethyleneglycol/100ml and 10 ml of 5 M NaCl/100 ml to precipitate the phage. After centrifugation for 15 min at 19,000 X g, the phage pellet was resuspended in 10 ml of Tris-Mg buffer, 7.76 gofCsCl was added, and the resulting solution was spunfor12 to 15 hr at 34,000rev/minin a Beck-manno. 40 rotor. The resulting band of phage was banded again in CsCland then dialyzed in Tris-Mg buffer. TheDNAwas extracted three times with redis-tilledphenol, and it was extensively dialyzed against 0.1 MNa2SO4orTris-Mg buffer.
Shearing ofXDNA andmercurycomplexes. DNA solutionswereusuallysheared at aconcentration of 25 ,ug/ml in0.1 MunbufferedNa2SO4.Atotal volumeof 22ml wasalwaysused.
The shearingtechnique described by Skalka,Burgi, and Hershey (10)was followed. To shear the DNA to one-half, one-tenth,andone-fifteenth lengthfragments, aGT-21 Hellermotor withcontroller unit wasused. One-twentieth length fragments were obtained by shearing the DNA for 30 min at 20,000 rev/min in a Virtis45 homogenizer.
The XDNA halveswereobtained byshearing for60 min at 1,500 rev/min. One-tenth and one-fifteenth lengthfragments were produced by shearing at 4,000 rev/minfor4hr or at8,000rev/minfor 1 hr, respec-tively. All DNA solutions were sheared in a Virtis stainless-steelcup at 4 C. Fragmentlengthswere char-acterized bysedimentation in 1 M NaClneutral 5 to 20%sucrosegradients withintact XDNAas marker. TheBurgi-Hershey formula (2) wasused for estima-tion ofmolecularweights.
After shearing oftheDNA, it was banded in Hg-Cs2SO4 density gradients according to Skalka et al. (10). Every timeanewpreparationof DNAwas ob-tained, atrial centrifugationwasmadetodeterminean appropriateRF.AllRFvaluesusedranged from0.20 to 0.30.Thesolutionscontained,perml,5to50,ug of DNA,0.05ml of0.1 Msodiumborate, pH 9.0,43to 45% byweightofCs2SO4 (Gallard-Schlesinger), and the amountof mercuryspecified above. The mercury was always added last. Volumes of 3.2 to5.3 ml of solution were used. The tubes were overlaid with mineraloiland spunfor36 hrat36,000rev/mmnina fixed-anglerotor,at4C. Atthe end of the run, frac-tions werecollecteddropwisefrom the bottomofthe tube.
Separation of strands. The technique described by
HradecnaandSzybalski(6) toseparate the strandsof X DNA was followed, with the use of
polyuridylic-polyguanylic acid[poly (UG)Iobtained from Schwarz BioResearch, Inc. (lot 6801). The purified labeled phage wasdialyzedovernightin 10-3M sodium
ethyl-enediaminetetraaceticacid,pH8.0,at4 C.Thephage
solution was then mixed withpoly (UG)andsarkosyl,
heat-denatured for3 minat95C,chilledonice, and banded in CsClatadensityof 1.72g/cm3inanSW39 rotor at30,000rev/minfor 60 hr. Fractionswere col-lecteddropwisefrom the bottom of the tube.
Hydrolysis of DNAandthin-layerchromatography: bases.ThepurifiedDNA(ca.50ug)washydrolyzedto
free bases with70%perchloricacidat100 Cfor 1 hr. The hydrolysate was neutralized with KOH and
centrifuged to remove the precipitated potassium
perchlorate and charred sugars. The resulting bases were separated by two-dimensional chromatography on commercial cellulose plates (Eastman Organic Chemicals) without fluorescent indicator. Unlabeled markers of 5MC and 6MAP were added. The first sol-vent system used was n-butanol-absolute methanol-concentrated ammonia-water (60:20:20:1); the sec-ond solvent was absolute methanol-concentrated hy-drochloric acid-water (65:17:18). The spots were located under ultraviolet light, scraped into liquid scintillationvials, and counteddirectly in a Beckman liquid scintillation counter.
Mononucleotides. DNAsolutionsin amounts of0.1 to 1ml containing a total of 50 ,ug of DNA in Tris-Mg buffer,pH 7.4, were hydrolyzed to mononucleotides by consecutive treatment with pancreatic deoxyribonu-clease and snake venom phosphodiesterase (8). A reactionmixturecontaining20M&gofpancreatic deoxy-ribonucleaseand DNA wasincubated at 37 C for 30 min. The enzyme was then inactivated by heating to 95 Cfor5min. The digest was mixed with 0.1volume of 1 M glycine buffer, pH 9.2, and 400 ,ug of snake venom phosphodiesterase, and was hydrolyzed to mononucleotides by incubating at 37 C for 30 rnin. Theresultingmixture was adsorbed to activated char-coal, washed three times with distilled water, and eluted bywashingthreetimeswith50%ethanol-0.5% concentrated ammonia. Theelutedsolution was con-centratedin vacuo to a final volumeof0.2ml,and the mononucleotides were separated by two-dimensional chromatography on commercial PEI-cellulose plates (Eastman).
The firstsolventused was 1 MLiCl,and the second was 0.5 Nacetic acid. In betweensolvents, theplates wereimmersedinabsolute methanolandshaken for 15 min to remove theLiCl.The spots were demonstrated bymeansofautoradiography withKodakNo-screen X-rayfilm.Nucleotidecompositionsweredetermined by scraping the radioactive spots and counting in a Beckman liquid scintillationcounter.
RESULTS
Methyl content of whole X DNA.The methyla-tion ofDNAobtained fromXc1857
phage
grown on W4032 was studiedby
several differentpro-cedures. Purified
[14C-methyl]-labeled
DNA washydrolyzed
eithertobasesor tomononucleotides,
and theextentof
methylation
wasdeterminedby
assumingthe
specific activity
ofthemethyl
groups intheDNA to beequaltothespecific activity
of methionineinthegrowth
medium. Table1 showsthe results of severaldeterminations.
Autoradio-grams ofthin-layer chromatograms ofthese hy-drolysates demonstrated that 6MAP and 5MC
aretheonly
methyl-labeled compounds. Labeling
the DNA with32P-orthophosphate
followedby
hydrolysistomononucleotidesalsogave thesame total content of 5MC and 6MAP. The
determina-tion of the
methyl
contentby
directanalysis
of the32P-labeled mononucleotides did not
require
anyassumption
regarding
thespecific activity
ofthe938 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Methylation ofX cl857DNA grownon
W4032
5MC/X, 6MAP/X Ratio, D.NAprepn genomeA genomea 65MAC/
14C-methylbases... 114±t10(8) 213±18(8) 0.5440.05 4C-methyl
mononucle-otides.1107(4)I. 218+7(4) 0.50+0. 04 32P-mononucleotides.... 114+10(6) 215 i20(6)0.53±0.07 aContent of 5-methylcytosine (5MC) or 5-methylamino-purine (5MAP) per genome was calculated by using the molecular weight of X as 32 X 106 and the molar absorb-ancycoefficient(basedonphosphate)as6.4 X 103.
bNumbers in parentheses represent number of determina-tions.
phosphate,since thecountsobtained inthe
meth-ylated mononucleotide spots were directly
com-paredwith thecountsobtained in the four main mononucleotide spots. The fact that the same values for
methyl
content were obtainedby
32Plabeling
and'4C-methyl labeling
validates theassumption made
regarding
thespecific activity
ofthemethylgroups.
The
methyl
contents for X DNA described above areapproximately
twofoldhigher
thanthosereported
by Gough
andLederberg (5)
forX grown onK-12 strain C600. We have examinedthemethylation ofXDNAgrownonC600onlow
phosphate medium
using orthophosphate labeling
and
hydrolysis
to mononucleotides. From theaverageoftwo
experiments,
wefound 5MC con-tentsof56moles/genomeand 6MAP contentsof132 moles/genome, in
good
agreement with thevalues reported
by Gough
andLederberg.
Wehave no
explanation
forthe twofolddifference
in methylation forX grown onW4032 andC600. Thepossibility
ofa differentrestriction-modification
system betweenW4032 and C600 has been ruledout directly
by
comparing plating
efficiencies ofphage X on each strain (data not shown). The
difference in
methylation might
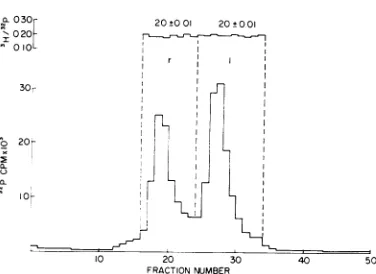
reflect the pres-ence ofthe sexfactorin W4032 orits absencein C600, or both, but we have no evidence on this point.Methyl content of X DNA single strands. The
proportion ofmethylatedto totalnucleotideswas
compared by
using
the separated strands of X DNA doubly labeled with[3H-methyl]-methio-nine and 32p
phosphate.
A typical experiment is illustrated inFig. 1. Theratioof3Hto32pcounts was calculated for each DNA-containingfrac-tion; nosignificantdifferences werefoundbetween
therand 1strands, indicatingthat the total con-tent of methylated bases is the same for each strand (Table2).To examine thecontent ofeach
of the twomethylatedbases in eachstrand,DNA waslabeledbygrowingXc1857onW4032 inTPA
X030r 20+001 20±001 I020
010I
I
do
20-a.I
10 20 30 40 50
FRACTIONNUMBER
FIG. 1. Distribution of methylated base residues between the strands ofX DNA. Phage X was doubly labeled with ['H-methyl]-methionine and32p. The two
strands wereseparatedbyheatdenaturing the DNAin the presence ofpoly (UG) followed by equilibrium density centrifugation in CsCl. Theratiosbetween 3H and 32p counts were determined for each DNA-con-tainingfraction.
medium with
['4C-methyl]-methionine.
Thesepa-rated strands were
hydrolyzed
with perchloric acid and the bases were resolvedby thin-layer
chromatography.
The ratio between 5MC and 6MAP obtained for each of the two separated strands was the same withinexperimental
error(Table 3).Weconclude that thetwo strandsofX
DNAare
equally
methylated
for both 5MC and6MAP.
Methylation along theDNA molecule.
Phage
Xc1857wasgrownonW4032 in TPAmedium
con-taining
['4C-methyl]-methionine.
The DNA wassheared to halves and the
separated fragments
werehydrolyzed
tobases. Table 4shows the aver-agevalues formethylated
base content obtained inthree differentexperiments.
Thefractional con-tentof 6MAP and 5MCpernucleotide (Table 4,columns 2 and
3)
in each halfis similar to thatobtained
for theintact DNA molecule.To
investigate
in further detail any possibleheterogeneity
inmethylation
along the DNAmolecule, smaller fragments were prepared by
shearing doubly labeled
[3H-methyl]-
and32P-DNA to
one-tenth,
one-fifteenth,
andone-twentieth molecular lengths and separating the
fragmentsby bandingthem in
Hg-Cs2SO4
densitygradients. Themethylation of each fragmentwas
inferred from theratios of3Hto
32p
countsin each DNA-containing fraction. Figure 2 illustrates atypical
experiment
ofthis kind. In all threetypes ofgradients,
with any of the three fragment lengths,thesamegeneralpatternfor total nucleo-tides wasobtained:ashouldercorrespondingto aregionof lowguaninepluscytosine (GC) content and twomajor peaks.
Skalkaetal. (10) haveshownthat, forXDNA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.255.443.77.214.2] [image:3.493.47.246.80.178.2]HIDALGO AND NASH
sheared to one-tenth length fragments, the shoulder contains material of
37%
GC contentderived from the b2 region. We have confirmed
these results by shearingtogether X c1857 and X
b2c1857 DNA to one-tenthlengthfragments
fol-lowed by banding the fragments in Hg-Cs2SO4 gradients. The shoulder corresponding to a low
GCcompositionshownin Fig. 2 wasmissingin
thecaseofXb2c1857DNA (datanot shown). By
direct analysis of nucleotide composition of X
DNAsheared toone-tenthlengths,wefound that
the shoulder corresponds to
40%0
GC composi-tion, and the two peaks to 46 and 57%oGC,
respectively.Figure3 shows agradient of X DNA sheared to one-fifteenth fragment length. FromFig. 2 and 3, it can be seen that alongthe two
major
peaks
the 3Hto32Pratiosareindistinguish-TABLE 2. Methylatiotn ofXDNA sinlgle strands
3H to32pratio Expta,
rstrand 1strand
1 0.34 i 0.02 (ll)b 0.34 + 0.01 (5) 2 0.44 + 0.01 (12) 0.44 i 0.02 (8) Since thephageusedtoobtain the strands had different specific activities of "P at the times at
which theexperimentsillustratedinthe tablewere
done,comparisonsofratiosfor1andrstrandare
valid only within each
experiment.
dThe numbers in parentheses indicate the number offractions
averaged
for each strand.TABLE 3. Methylated S-meilhylcytosine (SMC) and 6-methylaminopurinie (6MAP) ineach stranzdof
XDNA
Counts/min
Strand Ratio
5MCa 6MAPa
r 283 +t 12 570 + 25 0.50 i 0.03
1 381 i 15 683 + 18 0.56 ± 0.03
aFive determinations were made for each strand.
able. However, the 3Hto32Pratio decreasedin the shoulder region. Thesameresultwasobtained for
one-twentieth fragment lengths. Table 5 shows a summaryofall resultsobtained by this method. The X DNA one-fifteenth fragments show more
dramatically the presenceofanundermethylated
region of DNA in thecentralpartof themolecule. However,owingtothe large variation indifferent gradients, it isnotpossibletocalculatetheexact
length or the absolute extent of methylation of thisregion.
DISCUSSION
The major portion of the methylated nucleo-tides ofX DNA havenoknownroleineither the
lytic or lysogenic phases of the bacteriophage X
life cycle. The results presented above rule out
several different models concerning the function ofmethylgroupsin X DNA.
After infection of E.coli by phage X, the
paren-talstrandsare foundto haveanasymmetric fate
during replication; the1strand is found in circular
monomersandtherstrandin multigenomelength molecules (7). Asymmetry in the fatesof comple-mentary strands is a predicted feature of the
rolling circle model of DNA replication andmay
be a widespread feature in the physiology of
viruses. Thefinding that both strandsareequally
methylated, both in terms of total content per
strand and composition in each strand, implies that the bulk ofmethylated basescannotbeused
as arecognition elementtosignal theror1strand
in directing asymmetric replication, or in other processes that distinguish between the two strands.
The genes of X are clustered in functional
groups onthephage DNA, and these clustersare
characterized by differing GC content (10). The head and tail genes makeup the left arm of the
phage chromosome, as it is usually written, and haveauniform GC content of57%. Mostof the rightarmofthephagemapistakenupwithgenes
concerned with replication, recombination, and immunity; this region has a distribution ofGC
[image:4.493.55.245.280.352.2]contentthat varies from43 to 48%.Theleftarm
TABLE 4. MethylatedbasesinzXDNA halvesa
DNAhalf 61IAP/1001\LNb 5MC/100MNb 5MC/6MIAP 6MAP/100AC 5-\fC 100CI1
456X70 GC half 0.23 ± 0.02 0.12 i 0.01 0.52 ± 0.06 0.83 + 0.06 0.52 4 0.02
55% GChalf 0.21 + 0.01 0.09 + 0.01 0.43 + 0.05 0.93 + 0.05 0.34 0.02
aTheresultsareaverages ofthreedifferent
experiments.
bValuesare100 X (moles ofmethylated nucleotide per mole ofmononucleotide ineach half). ¢Valuesare100 X (moles of6-methylaminopurine per mole of adenine in eachhalf).
dValuesare100 X (molesof
5-methylcytosine
permole ofcytosine in each half).940 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.56.251.430.541.2] [image:4.493.56.451.573.644.2]METHYLATION PATIERN OF LAMBDA DNA
64±003 .72±0O02 .68±t004
0. 07
06
10 20 30 40
FRACTION NUMBER
FIG. 2. Distribution of methylated base residues
among one-tenth length fragments of differing GC
composition. Phage X DNA doubly labeledwith [3H-methyl]-methionine and 31P was sheared to one-tenth lengthfragments. Thefigure illustrates a typical
Hg-CS2SO4equilibrium centrifugation gradient. The ratios between 3H and 32P counts were calculated for all fractions corresponding to regions I, II, and III. The averageratio valuesaregivenforeach gradientzonte.
0-08
10000 8000 2
6000-2- 4000-2000
.54± 003 87±0 04 78±0 03
~~ ~
10 20 30 40 50 FRACTION NUMBER
FIG. 3. Distributioni of methylated base residues
among one-fifteenth kengthfragmenits of differing GC
composition. Phage X DNA doubly labeled with [3H-methyl]-methionineand 32Pwasshearedtoone-fifteenth length fragmentsandbanded inHg-CS2SO4asdescribed in the text. The ratios between 3H and 32pcounts were
calculatedforall fractions corresponding to regionsI, II,andIII.The averageratio valuesaregivenforeach gradientzone.
genesaretypicallytranscribed and translated late after induction or infection of X; the right arm
genes are expressed early. The data presented in Table 5 andFig. 2and 3show thattheseregions
cannotbedistinguished fromoneanother onthe basis ofmethylcontent,andthus ruleoutthe
pos-sibilitythatearlyversuslatetranscriptionis
regu-lated byrecognitionof sitesidentifiedby amajor
fraction of themethylgroupsin X.
Anextremehypothesis,i.e.,thatmethylationis featureless and thereforeplays norolein
recogni-TABLE 5. Methylation inXDNAfragments
No.of Normalized8Hto32Pratios XDNAshearedto 'deter- ingradientregiona
mina-tions
One-tenth 17' 911 6100 3 102 -4 5
One-fifteenth 7 76 -4- 9106 -4 10 103 ±~5 One-twentieth 6 90 4- 3104 4± 3 100 ±4 2 aRatiosbetween 3H and321 countsinall
DNA-containing fractions on any given gradient were
averaged,
thegradientwasdividedinthreeregionsfrom low to high GC content (the shoulder and the twomainpeaks illustrated onFig. 2 and Fig. 3), and the mean 3H to 32P ratio for each
region
was divided bythetotal gradient average.
tion ofspecific DNA elements in X, mayalso be ruledoutfromanexamination of the fine detail of
methylatedbasedistribution. Ifmethylationwere
not based on the interaction of methylating
en-zymeswith specific DNA sites,onewould expect
that themethylcontent ofaregion would reflect
thedensityofthepotential substratenucleotides,'
adenine and cytosine. The density ofmethylated nucleotides with respect to potential precursor nucleotides may be calculated from the data in Table 4by combining the density of methylated bases per nucleotide and thecompositionof each half. Such acalculation (Table 4, columns 5 and 6) revealsasimilardensityof6MAP/adenine
be-tween thetwohalves butavery different density of5MC/cytosine.Thismeansthatmethylationof
cytosine has proceeded by a mechanism that is
responsive to dinucleotide or longer sequences.
Thisfindingis inagreementwith earlier chemical
studies showing that the distribution of methyl
cytosines amongthe variouspyrimidine tractsof
coli DNA isnonrandom (4).
T'he results obtained by shearing the DNA to
small fragment lengths reveal a more striking asymmetrythanthat discussed for halves. A small
fragment, arisingfromthe centralportionof the
X genome, shows a decrease in methylation as
compared to the totalmethylation of the rest of
the DNAmolecule,which is itselffairlyconstant.
Ifweassumethat themethylationof adenine and
cytosineisonlyafunction of the absoluteamount
of each base present, as done above for half
molecules, we calculate that the ratio of
methyl-atedtototal nucleotides would beexpectedto be
%higherfor a40%0/ GCfragmentthan forthe
entire molecule.Instead,wefind that the40%,- GC
fragment is between 10 and25% lessmethylated
than the average molecule. In the other regions resolvedbythe CS2SO4-Hg gradient, the
nonuni-formityofmethylationis much less dramatic.
VOL. 10, 1972
941
m(
a.
a.I
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.44.232.62.209.2] [image:5.493.245.444.79.175.2] [image:5.493.44.236.314.449.2]HIDALGO AND NASH
Thus, the distribution of methylated nucleo-tidesis not a simple reflection of either total nu-cleotides or the content of the potential acceptor
nucleotides, adenine and cytosine. It is possible
thatthisnonrandomness in methyl group distribu-tion reflects the sum of many sequence-specific methylated sites-a few each for modification, transcriptional starts and stops, and replication signals. The data ofearlier workerson modifica-tion andourdataondistribution of methylgroups simply rule out that one of these functions ac-counts for the majority of methyl groups in X DNA. On the other hand,a yet unknown single function for the bulk of the methylated
nucleo-tidesmay exist.
ACKNOWLEDGMENTS
We thank J. Little for his help in teaching us his shearing techniques. We also thankJ.Little,D.Neville, J.L.Rosner, and M.Shulman fortheir carefulreadingsof the manuscript.
LITERATURE CITED
1. Arber,W., and S. Linn.1969.DNAmodification and restric-tion. Annu. Rev. Biochem.38:467-500.
2. Burgi,E., and A. D.Hershey. 1963.Sedimentationrate as a measure ofmolecularweight of DNA. Biophys. J. 3:309-321.
3.Cook,A., and J. Lederberg. 1962.Recombination studies of lactose nonfermenting mutants of Escherichia coli K 12. Genetics47:1335-1353.
4. Dosko6il, J., and Z. Sormova. 1965. The sequences of 5-methylcytosineinthe DNA of Escherichia coli. Biochem. Biophys.Res.Commun. 20:334-339.
5. Gough,M., and S. Lederberg. 1966. Methylated basesinthe host-modified deoxyribonucleicacid ofEscherichia coli and bacteriophageX. J.Bacteriol.91:1460-1468.
6. Hradecna, Z., and W.Szybalski. 1967.Fractionation ofthe complementary strands of coliphageXDNAbasedon the asymmetric distribution ofthe Poly I, G-binding sites. Virology32:633-643.
7. Ihler,G., andY. Kawai. 1971.Alternate fates of the comple-mentarystrands of lambdaDNAafter infection of Esche-richiacoli. J. Mol.Biol. 61:311-328.
8. Kelly, T. J.,Jr., and H.0.Smith. 1970. A restriction enzyme from Hemophilus influenzae. I. Purification and general properties. J.Mol.Biol. 51:379-391.
9. Ledinko, N. 1964. Occurrenceof5-methyldeoxycytidylate in the DNAof phage lambda.J.Mol. Biol. 9:834-835. 10. Skalka, A., E.Burgi, and A. D. Hershey. 1968.Segmental
distribution of nucleotidesin the DNA ofbacteriophage lambda.J.Mol.Biol. 34:1-16.
942 J. VIROL.

![FIG.2.fractionslengthaveragebetweencomposition.methyl]-methionineCS2SO4amongDistributionof methylatedbaseresiduesone-tenthlengthfragmentsof differingGCPhageX DNA doubly labeled with[3H- and 31Pwas shearedtoone-tenth fragments.The figureillustratesa typical Hg- equilibrium centrifugation gradient.Theratios3Hand 32Pcountswerecalculated forall correspondingtoregionsI,II, and III.Theratio values are given for each gradient zonte.](https://thumb-us.123doks.com/thumbv2/123dok_us/1598774.112718/5.493.44.236.314.449/fractionslengthaveragebetweencomposition-amongdistributionof-methylatedbaseresiduesone-tenthlengthfragmentsof-differinggcphagex-figureillustratesa-pcountswerecalculated-correspondingtoregionsi.webp)