0022-538X/07/$08.00⫹0 doi:10.1128/JVI.01785-07
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
A Novel Zinc-Binding Domain Is Essential for Formation of the
Functional Junı´n Virus Envelope Glycoprotein Complex
䌤
Joanne York and Jack H. Nunberg*
Montana Biotechnology Center, The University of Montana, Missoula, Montana 59812
Received 14 August 2007/Accepted 25 September 2007
The envelope glycoprotein of the Junı´n arenavirus (GP-C) mediates entry into target cells through a pH-dependent membrane fusion mechanism. Unlike other class I viral fusion proteins, the mature GP-C complex retains a cleaved, 58-amino-acid signal peptide (SSP) as an essential subunit, required both for trafficking of GP-C to the cell surface and for the activation of membrane fusion. SSP has been shown to associate noncovalently in GP-C via the cytoplasmic domain (CTD) of the transmembrane fusion subunit G2. In this report we investigate the molecular basis for this intersubunit interaction. We identify an invariant series of six cysteine and histidine residues in the CTD of G2 that is essential for incorporation of SSP in the GP-C complex. Moreover, we show that a CTD peptide fragment containing His-447, His-449, and Cys-455 specifically binds Zn2ⴙ at subnanomolar
concentrations. Together, these results suggest a zinc finger-like domain structure in the CTD of G2. We propose that the remaining residues in the series (His-459, Cys-467, and Cys-469) form an intersubunit zinc-binding center that incorporates Cys-57 of SSP. This unusual motif may act to retain SSP in the GP-C complex and position the ectodomain loop of SSP for its role in modulating membrane fusion activity. The unique tripartite organization of GP-C could provide novel molecular targets for therapeutic intervention in arenaviral disease.
Arenaviruses are endemic to rodent populations, and zoo-notic infection of humans can result in severe acute hemor-rhagic fevers (43, 49). Up to 300,000 infections by Lassa fever virus occur annually in Africa (44), and cases of Argentine, Bolivian, and Venezuelan hemorrhagic fevers are regularly reported in South America (49). Genetic diversification and worldwide dissemination of arenaviruses with their rodent hosts (51) provide opportunities for the continued emergence of new pathogenic strains (10). Vaccines to prevent arenavirus infection are not available, and treatment options for arenavi-ral hemorrhagic fevers are extremely limited. Ribavirin, a non-specific antiviral agent with an incompletely understood mech-anism of action, is currently used in severe and life-threatening cases with mixed results (60). In the case of Argentine hemor-rhagic fever caused by infection with Junı´n virus (JUNV), early administration of human convalescent antiserum is the treat-ment of choice (17). There is an urgent need for effective vaccines and therapeutic agents to combat endemic arenavirus infections and to address biodefense concerns.
The arenaviruses are enveloped RNA viruses that encode ambisense expression of four viral proteins (8). The nucleo-protein (N) and RNA-dependent RNA polymerase (L) asso-ciate with the two single-strand genomic RNA segments to form the ribonucleoprotein core. Virion assembly occurs at the plasma membrane (45) and is promoted by the matrix protein (Z) (48, 59). Virus entry into the target cell is initiated by binding of the envelope glycoprotein (GP-C) to a cell surface receptor:␣-dystroglycan in the Old World viruses (9, 34, 58) and transferrin receptor 1 in the New World hemorrhagic fever
viruses (50). Upon binding, the virion particle is endocytosed (7), and fusion with cellular membrane is subsequently acti-vated by acidic pH in the maturing endosome (11). As with other class I viral fusion proteins (12, 13, 28, 29, 56), this membrane fusion reaction is mediated by a series of confor-mational changes in GP-C that ultimately lead to formation of a stable six-helix bundle structure (18, 21, 63). Intervention strategies that target the steps in virus entry offer promise for effective antiviral therapeutics. Indeed, small-molecule arenavirus-specific entry inhibitors have recently been discov-ered (5).
GP-C is unique among class I envelope glycoproteins in that the mature complex retains its cleaved signal peptide as an essential element, together with the receptor-binding (G1) and transmembrane fusion (G2) subunits. The stable, 58-residue signal peptide (SSP) was initially identified as a component of the Old World arenavirus GP-C complex (16, 19, 35) and found to be essential for proteolytic maturation of the glyco-protein precursor in the Golgi body (1, 15, 35). Without SSP, the G1-G2 precursor is specifically retained in the endoplasmic reticulum (ER), in part through dibasic ER retrieval signals in the cytoplasmic domain (CTD) of G2 (1). This acts as a quality control mechanism to ensure that only the mature tripartite GP-C complex is transported to the cell surface for virion assembly. Unexpectedly, SSP incorporation in the GP-C com-plex also plays a crucial role in the activation of pH-dependent membrane fusion activity (65). Furthermore, the unusual in-terplay between the ectodomains of SSP and G2 in membrane fusion appears vulnerable to interference by the newly discov-ered small-molecule entry inhibitors (reference 5 and unpub-lished results). In this report, we provide biochemical and genetic evidence for a high-affinity zinc-binding domain (ZBD) in the CTD of G2 and discuss the implications of this novel structure for SSP association in GP-C and its membrane fusion activity.
* Corresponding author. Mailing address: Montana Biotechnology Center, The University of Montana, Science Complex, Room 221, Mis-soula, MT 59812. Phone: (406) 243-6421. Fax: (406) 243-6425. E-mail: [email protected].
䌤Published ahead of print on 10 October 2007.
13385
on November 8, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
GP-C expression and MAbs.GP-C from the pathogenic JUNV strain MC2 (24) was expressed in Vero cells by cotransfection of pcDNA3.1 (Invitrogen) plasmids encoding CD4sp-GPC (in which SSP is replaced by the conventional signal peptide of CD4) and SSP-term (in which a stop codon is introduced following the C-terminal SSP amino acid T58) (65). These components associate intransand reconstitute the native GP-C complex (15). Transient expression utilized a recombinant vaccinia virus expressing T7 RNA polymerase (vTF7-3 [20]) and the T7 promoter of pcDNA3.1. The vaccinia virus-based -galactosi-dase fusion reporter assay (46) was used to characterize the ability of GP-C to mediate pH-dependent cell-cell fusion (65, 66). Mutations in GP-C were intro-duced by QuikChange mutagenesis (Stratagene), and all constructs were verified by DNA sequencing. The murine monoclonal antibody (MAb) BF11 (53), di-rected to the G1 subunit of GP-C, was contributed by Tom Ksiasek and Tony Sanchez (Special Pathogens Branch, CDC, Atlanta, GA) and obtained through the NIH Biodefense and Emerging Infections Research Resources Repository. GP-C expression was detected by immunoprecipitation of metabolically la-beled complexes (65, 66). Briefly, cells were metabolically lala-beled using 50
Ci/ml of35S-ProMix (Amersham Pharmacia Biotech) for 12 to 16 h. Cultures were washed in physiological buffered saline and lysed using cold Tris-saline buffer (50 mM Tris-HCl and 150 mM NaCl at pH 7.5) containing 1% Triton X-100 nonionic detergent and protease inhibitors (1g/ml each of aprotinin, leupeptin, and pepstatin). The expressed glycoproteins were immunoprecipitated from cleared lysates using MAb BF11 and protein A-Sepharose (Sigma). Pro-teins were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophore-sis (SDS-PAGE) using NuPAGE 4 to 12% Bis-Tris gels (Invitrogen) and the recommended sample buffer containing reducing agent. In some cases, the gly-coproteins are deglycosylated by treatment with peptide–N-glycosidase F (New England Biolabs) to facilitate resolution of the G1 and G2 subunits. Molecular size markers included14C-methylated Rainbow proteins (Amersham Pharmacia Biotech). Radiolabeled proteins were visualized by phosphorimaging (Fuji FLA-3000G) and analyzed using ImageGauge software (Fuji).
Purification of chimeric MBP.The sequence corresponding to the entire CTD of G2 (encoding P445 to H485) was adapted by PCR and appended in frame at the C terminus of maltose binding protein (MBP) using the KpnI polylinker site in the plasmid pMAL-c2E (New England Biolabs), to generate pMBP-ZBD. Expression of the chimeric MBP-ZBD protein inEscherichia coliK-12 strain TB1 (New England Biolabs) was induced by the addition of isopropyl- -thioga-lactopyranoside (IPTG) as recommended by New England Biolabs. Cell pellets were frozen overnight and subsequently lysed using Bugbuster protein extraction reagent (primary amine free; Novagen) in the presence of protease inhibitors (1
g/ml each of aprotinin, leupeptin, and pepstatin). Cleared extracts were incu-bated with amylose resin (New England Biolabs) in buffer containing 20 mM HEPES (pH 6.8) and 200 mM NaCl for 2 h at 4°C with rocking. The resin was then packed into a column and washed extensively with this buffer. Bound MBP chimera was eluted by the addition of 10 mM maltose. The yield of purified protein was 2 to 3 mg per g of wet cells.
Protein characterization.Proteins were analyzed by SDS-PAGE and visual-ized using SYPRO Red protein stain (Molecular Probes). Molecular size mark-ers included Mark12 standards (Invitrogen). Protein concentrations were deter-mined by the Coomassie Plus assay (Pierce) and corresponded to those measured by optical density at 280 nm in 6 M GuHCl (14). Zinc content was determined using the 4-(2-pyridylaxo)resorcinol (Aldrich) reagent in 4 M GuHCl (30, 42).
Equilibrium binding studies.Prior to use for zinc binding, the purified pro-teins were dialyzed into buffer containing 50 mM HEPES (pH 6.8) and 20 mM NaCl (HEPES-NaCl buffer) and incubated with an excess of tris(2-carboxyethyl)-phosphine (TCEP; Pierce) for 20 min at room temperature in order to reduce any disulfide bonds that form on exposure to air and in the absence of zinc. TCEP does not bind metal ions and is thus uniquely suited for these studies (23, 32). We used 1 mM TCEP for 10M protein and 0.1 mM for 200 nM protein. These concentrations of TCEP were maintained throughout the subsequent binding experiment. Radioactive zinc (65
Zn) was obtained from Brookhaven National Laboratory (U.S. Department of Energy) as a 2 mM solution of ZnCl2 in 0.1 N HCl. The specific activity is 2 Ci/mmol (batch Zn65-012506). Equilib-rium dialysis studies were performed using a modified method in which unbound ligand is determined by centrifugal ultrafiltration (26, 37). In a typical study, 200 nM of MBP-ZBD protein is incubated at room temperature with 10 nM of neat 65Zn2⫹in 5.0 ml of HEPES-NaCl buffer containing 0.1 mM TCEP. Equilibrium binding is attained within 60 min (not shown). At this time, the reaction mixture is sampled to determine the total amount of65
Zn2⫹, and the free65 Zn2⫹is determined by ultrafiltration (Amicon Ultra-4; 10,000-molecular-weight cutoff).
Based on the specific radioactivity of the reaction mixture (cpm/ml for the known total concentration of Zn2⫹), the concentration of free and bound zinc is deter-mined. Samples were counted using a Captus multichannel analyzer (Capintec) and a single-well detector (2-ml capacity). The high-energy gamma emission of 65
Zn (950 to 1,200 keV) was detected at⬃2.5% efficiency. Using this method-ology, we were effectively limited to detecting zinc binding at concentrations of ⱖ10 nM Zn2⫹(i.e.,ⱖ2,000 cpm per 2-ml sample).
Competitive metal ion binding studies.The specificity and selectivity of metal ion binding were determined in a 5-ml equilibrium binding reaction mixture containing 200 nM MBP-ZBD, 10 nM65Zn2⫹, and a 10- to 10,000-fold excess of CdCl2, CoCl2, CuCl2, MgCl2, or cold ZnCl2. A 50% inhibitory concentration (IC50) for each metal ion was defined as that which competes for 50% of65
Zn2⫹ binding. Because of the excess of MBP-ZBD over65Zn2⫹in the reaction, the relative IC50is reported as a ratio of the competitor IC50compared to that of cold Zn2⫹.
RESULTS
Conserved cysteine and histidine residues in the CTD of G2.
Previous studies have demonstrated that the CTD of G2 is essential for SSP incorporation in the mature GP-C complex (1). GP-C mutants bearing deletions in this region are unable to bind SSP, while chimeric proteins containing only the trans-membrane domain and CTD of G2 associate with and require SSP for transit through the Golgi body (1). To examine the role of the G2 CTD in the assembly of the tripartite GP-C complex, we compared amino acid sequences in the New World and Old World arenaviruses (Fig. 1). Inspection of the CTD revealed a well-conserved N-terminal region (P445 through G470) and a variable C-terminal region (K471 through H485) that includes dibasic ER retrieval motifs (1). Strikingly, the N-terminal re-gion contains a series of six invariantly conserved cysteine and histidine residues (Fig. 1). This array (H447, H449, C455, H459, C467, and C469) was suggestive of the clustered cysteine and histidine sequences found in many zinc finger proteins (27, 31, 33, 38).
Zinc fingers are compact protein domains that coordinate one or more zinc ions (31). Tetrahedral ligation of Zn2⫹serves
to stabilize protein folds and, in some cases, interprotein in-teractions (22, 27, 40). Structurally diverse, zinc fingers are found in cellular proteins that function in DNA replication and repair, transcriptional regulation, protein modification and degradation, metabolism and signaling, vesicular trafficking, and apoptosis (22, 31, 33, 38). It has been estimated that zinc finger proteins represent 2% of the human genome coding capacity (41). Zinc fingers are classified according to the amino acids involved in zinc coordination (predominantly cysteines and histidines) and by the size, topology, and folding of the intervening loops (27, 33). The array of cysteine and histidine residues in G2 generally conforms to the sequence motifs in known zinc fingers, although the identical pattern is not rep-resented in existing databases (InterPro database, European Bioinformatics Institute). These considerations led us to inves-tigate the zinc-binding properties of the CTD.
Zinc-binding activity of the G2 CTD. To characterize zinc binding, we appended the entire CTD sequence (P445 to H485) (Fig. 1) to the C terminus of MBP (New England Bio-labs). The chimeric MBP-ZBD protein was expressed inE. coli
and purified using amylose resin affinity chromatography (Fig. 2). No Zn2⫹was added during the purification procedure, and
the isolated protein was devoid of zinc (⬍0.03 mol per mol of protein), as determined by reaction with
on November 8, 2019 by guest
http://jvi.asm.org/
orcinol (42). We also generated a mutant chimera (MBP-noZBD) in which the conserved CTD cysteine and histidine residues were all replaced by amino acids that do not coordi-nate zinc ion (27). The three cysteines were changed to serine, a similarly sized side chain that differs only at the hydroxyl oxygen, whereas the three histidines were replaced by aspara-gine, another nitrogen-containing side chain. The MBP-noZBD protein was used to assess nonspecific zinc binding and the potential role of the cysteine-histidine sequence motif.
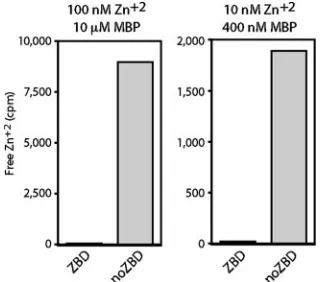
Binding of radioactive65Zn2⫹was measured by a modified
equilibrium dialysis method in which centrifugal ultrafiltration is used to determine the concentration of free ligand (26, 37). In initial experiments, we incubated 10M of MBP-ZBD with
100 nM 65ZnCl
2 (Brookhaven National Laboratory; 2 Ci/
mmol) in 5.0 ml of HEPES-NaCl buffer containing 1 mM TCEP, a nonthiol reducing agent that does not chelate metal ions (23, 32). After 60 min of incubation with MBP-ZBD, less than 1% of the input zinc radioactivity was found as free Zn2⫹
in the ultrafiltrate (Fig. 3, left), indicating complete binding. In contrast, greater than 90% of the input Zn2⫹from the
[image:3.585.103.482.69.279.2] [image:3.585.341.502.508.649.2]MBP-noZBD binding reaction was free. Our results indicate that the
FIG. 1. The tripartite GP-C complex. A representation of the JUNV GP-C open reading frame is shown on top. Amino acids are numbered from the initiating methionine, and cysteine residues () and potential glycosylation sites ( ) are marked. The signal peptidase and SKI-1/S1P (4, 35, 39) cleavage sites, and the resulting SSP, G1, and G2 subunits, are indicated. Within G2, the C-terminal transmembrane domain (TM) and CTD (cyto) are marked, as are the N- and C-terminal heptad-repeat regions in the ectodomain (light gray shading). Below, we show a comparison of arenavirus amino acid sequences in SSP and the CTD of G2. Sequences are from New World isolates Junı´n (D10072), Tacaribe (M20304), Pichinde´ (U77601), Machupo (AY129248), and Sabia´ (YP_089665) and Old World isolates Lassa-Nigeria (X52400), Mopeia (M33879), and LCMV-Armstrong (M20869). The array of cysteine and histidine residues is highlighted in gray and numbered, and the two hydrophobic, membrane-spanning regions in SSP (h1 and h2) are marked. A schematic drawing of the tripartite GP-C complex is shown at the lower right. The model depicts the bitopic membrane topology of SSP in relation to the CTD of G2 (2). The myristoylation site (66) and fusion-critical Lys-33 residue (65) in SSP are indicated, as are the heptad-repeat regions in the G2 ectodomain (thicker lines). The N or C termini of the subunits are labeled. The drawing is not to scale.
FIG. 2. Purified MBP chimeras containing the CTD of G2. The purity of the chimeric proteins (500 ng each) was assessed using SDS-PAGE. Wild-type MBP-ZBD and the individual mutants are indi-cated. C467Stop contains a stop codon at position 467, and MBP-noZBD includes the three cysteine and three histidine mutations. The isoforms seen in the samples vary between analyses and likely reflect different conformational states of the protein. Numbers at right are molecular masses in kilodaltons.
FIG. 3. Specific binding of zinc ion to MBP-ZBD. MBP-ZBD and MBP-noZBD (10M, left, and 400 nM, right) were incubated with 100 ⌴and 10 nM65Zn2⫹, respectively, as described in the text. Free 65Zn2⫹was determined by ultrafiltration. The counts per minute (cpm)
of free65Zn2⫹in the ultrafiltrate (1-ml or 2-ml sample, respectively)
are plotted. The total radioactivity in the binding reaction is used as the full-scale value in each graph (10,000 and 2,000 cpm, respectively) to demonstrate the relative proportion of unbound zinc.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.83.240.594.650.2]CTD of G2 can bind Zn2⫹and that binding is dependent on
the intact array of cysteine and histidine residues. Further-more, all the available Zn2⫹was bound at a zinc concentration
of 100 nM. In other experiments using higher concentrations of Zn2⫹, binding reached a plateau of⬃0.3 mol zinc per mol of
protein (not shown). The substoichiometric amount of zinc binding may reflect oxidative damage to the cysteines.
Efforts to attain subsaturating concentrations of Zn2⫹and
determine an apparent dissociation constant (Kd) were
ulti-mately limited by the relatively low specific activity of the zinc isotope and the high affinity of binding (see Materials and Methods). At the lowest practical zinc concentration (10 nM 65Zn2⫹), binding by the MBP-ZBD protein remained
quanti-tative even at low (200 to 400 nM) protein concentrations (Fig. 3, right). Based on the limits of the assay, we can conclude that theKdfor Zn
2⫹binding to MBP-ZBD is likely to be
signifi-cantly less than 1 nM (see below). The high-affinity interaction of MBP-ZBD with Zn2⫹is consistent with zinc finger
bind-ing (3).
The ability of MBP-ZBD to bind other divalent transition-metal ions was determined by equilibrium dialysis using 10 nM 65Zn2⫹and an excess of Cd2⫹, Co2⫹, or Cu2⫹. The
concen-tration of competitor ion required to inhibit Zn2⫹binding by
50% (IC50) was used to assess the relative binding affinity. The selectivity of binding to MBP-ZBD was determined as: Zn2⫹⬎
Cu2⫹⬎Cd2⫹⬎⬎Co2⫹. This order of preference conforms to
that of well-characterized zinc finger proteins (3, 36). The respective IC50s for Cd
2⫹, Co2⫹, and Cu2⫹were 10-, 450-, and
5-fold that of Zn2⫹. Mg2⫹ did not compete for binding at
concentrations up to 100M, demonstrating the requirement for a transition-metal ion and the specificity of zinc binding.
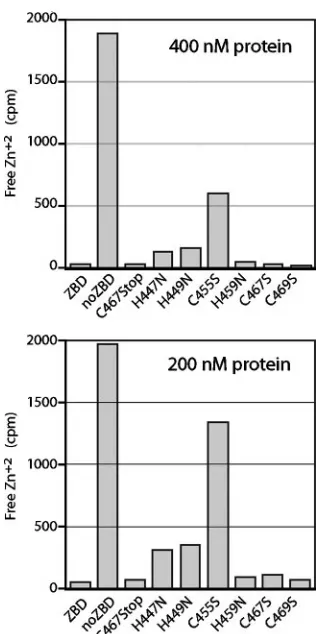
Amino acid requirements for zinc-binding activity. To de-termine the role of specific cysteine and histidine side chains in zinc binding, we introduced the respective serine and aspara-gine mutations individually into MBP-ZBD. The mutant pro-teins were affinity purified (Fig. 2), and 400 nM of each was used to determine binding with 10 nM65Zn2⫹(Fig. 4, top). To
enhance the distinctions among the mutants, the assay was also performed using 200 nM protein (Fig. 4, bottom). Under these conditions, zinc binding was reduced markedly by the C455S mutation and to a lesser extent by H447N and H449N (Fig. 4). These results extrapolate to dissociation constants of 12 nM for C455S and 1 to 2 nM for H447N and H449. We infer that these three membrane-proximal residues (H447, H449, and C455) are required for high-affinity binding of Zn2⫹by MBP-ZBD.
Each is capable of coordinating zinc ion or may indirectly contribute to formation of the properly folded zinc-binding center. Interestingly, mutations in the three distal residues in the array (H459N, C467S, and C469S) had little or no detect-able effect on zinc binding in this assay.
To further dissect the sequence requirements for zinc bind-ing, we truncated MBP-ZBD at position 467 to delete the two C-terminal cysteines. On equilibrium dialysis, this C467Stop mutant was indistinguishable from the wild-type protein in its zinc binding (Fig. 4). Taken together with the wild-type binding by the H459N mutant (above), we conclude that the mem-brane-proximal region of the array including H447, H449, and C455 can fold autonomously to bind zinc ion with high affinity.
Conserved cysteines and histidines are required for SSP incorporation in GP-C. The functional significance of zinc
binding and the conserved residues in GP-C was determined by individually replacing each cysteine and histidine residue in the CTD with serine or asparagine, respectively. These mutations were introduced into a GP-C construct (CD4sp-GPC) that utilizes the conventional signal peptide of human CD4 for expression of the G1-G2 precursor protein (1). Coexpression of wild-type CD4sp-GPC intranswith an SSP polypeptide has been shown to reconstitute the native GP-C complex (1, 15, 64). Thistrans-complementation method was chosen to obviate experimental concerns regarding possible effects of the muta-tions on signal peptidase cleavage of full-length GP-C. The
GP-C complexes were metabolically labeled with35S-ProMix
[image:4.585.341.499.65.382.2](Amersham Pharmacia Biotech) and immunoprecipitated us-ing G1-specific MAb BF11 (53). SDS-PAGE analysis of the wild-type GP-C complex reveals the association with SSP (Fig. 5, top) and the proteolytic maturation of the G1-G2 precursor (Fig. 5, bottom). In contrast to the wild-type CD4sp-GPC, none of the single cysteine and histidine mutants was able to associate with SSP intrans(Fig. 5, top). Proteolytic maturation of the G1-G2 precursor was also absent in the mutants (Fig. 5, bottom), presumably reflecting the inability of the precursor to transit through the Golgi body in the absence of SSP (1). As anticipated, the mutants were unable to support membrane
FIG. 4. Zinc binding by MBP-ZBD mutants. Wild-type and mutant MBP-ZBD proteins (400 nM in top panel and 200 nM in bottom panel) were incubated with 10 nM65Zn2⫹as described in the text. The
cpm of free Zn2⫹are plotted, and the full scale (2,000 cpm) represents
10 nM Zn2⫹. The results shown here are representative of six inde-pendent studies.
on November 8, 2019 by guest
http://jvi.asm.org/
fusion (Fig. 6). Thus, CTD residues that are dispensable for high-affinity zinc binding (H459, C467, and C469) as well as those that are required (H447, H449, and C455) are each essential for SSP incorporation in the GP-C complex.
Taken together, our studies provide strong biochemical ev-idence for a cysteine- and histidine-containing zinc-finger-like
motif in the CTD of G2 and for its essential role in SSP incorporation in the tripartite GP-C complex. Although many viruses have adopted zinc fingers in their structural, regulatory, and catalytic proteins (25, 47, 57, 62, 67), the motif has not been described in a virus envelope glycoprotein. In addition to its involvement in SSP association, the unusual ZBD in G2 protein may participate in functions typically served by the CTD of viral envelope glycoproteins, including roles in the intracellular trafficking and membrane fusion activity of GP-C and in virion assembly and budding.
DISCUSSION
We have identified a novel zinc-binding activity in the CTD of the arenavirus G2 fusion protein. An array of six cysteine and histidine residues in the CTD is uniformly conserved in New World and Old World arenaviruses, and peptide frag-ments containing the three membrane-proximal residues (H447, H449, and C455) specifically bind Zn2⫹at
subnanomo-lar concentrations. Our results indicate that this region of the cysteine-histidine array in the CTD of G2 can fold to form a zinc-finger-like structural domain.
The three distal residues in the array (H459, C467, and C469) are not required for high-affinity zinc binding. Nonethe-less, all six residues in the array are essential for SSP incorpo-ration in the mature GP-C complex; mutations at any of these positions prevent association and render the envelope glyco-protein defective. This shared phenotype may of course arise through a variety of mutational effects. Interestingly, however, SSP association in GP-C is also abolished by mutation at an absolutely conserved cysteine residue (C57) in the C-terminal CTD of SSP (54, 64). Based on the bitopic membrane topology of SSP in which both N and C termini reside in the cytosol (2) (Fig. 1), this C-terminal region must be translocated from the ER lumen following signal peptidase cleavage. With the ex-ception of the inviolable C57, other residues in this short re-gion (R55SCT58in JUNV) are not conserved among the arena-viruses, and each can be mutated without effect on SSP association (64). In light of the shared phenotype in mutants at C57 in SSP and within the cysteine-histidine array of G2, we speculate that H459, C467, and C469 may participate together with C57 in forming an intersubunit zinc-binding center, pos-sibly in conjunction with the membrane-proximal zinc center. Although translocation of C57 into the cytosol is independent of G2 (2), formation of a zinc finger motif bridging SSP and G2 may be needed to stabilize SSP incorporation in the GP-C complex.
We have shown that peptide fragments including the mem-brane-proximal H447, H449, and C455 residues can bind Zn2⫹
[image:5.585.66.261.70.270.2]with extremely high affinity and that single-amino-acid substi-tutions at these positions substantially reduce affinity to a range that is consistent with the effects of single coordination-site mutations in known zinc fingers (6, 55). The residual nanomo-lar affinity observed in these triply ligated mutants, however, might suggest that zinc binding by the three distal cysteine and histidine residues would also be detectable. The lack of binding in our studies may simply reflect the inability of these residues to coordinate zinc or the oxidative lability of the cysteines in the recombinant peptide model in MBP-ZBD. Alternatively, SSP may be important for zinc binding and formation of the
FIG. 5. SSP association in GP-C complexes bearing mutations in the G2 cysteine-histidine array. Vero cells were transfected to express wild-type and mutant CD4sp-GPC intranswith wild-type SSP (⫹SSP) (65). Metabolically labeled complexes were immunoprecipitated using G1-specific MAb BF11 (53) and separated on NuPAGE (Invitrogen) 4 to 12% Bis-Tris gels under denaturing and reducing conditions (top panel). The image has been darkened in order to visualize SSP. The mature G1 and G2 subunits are not as well resolved as the glycopro-teins and are rendered distinct on deglycosylation by peptide–N -gly-cosidase F (lower panel). The G1-G2 precursor and GP-C subunits are labeled, as well as the14C-labeled protein markers (in kilodaltons)
(Amersham Biosciences).
FIG. 6. Membrane fusion activity of mutant GP-C complexes. pH-dependent cell-cell fusion by thetrans-complemented GP-C complex was initiated by a pulse of medium adjusted to pH 5.0 and detected using the recombinant vaccinia virus-based-galactosidase reporter assay (46) as previously described (65, 66).-Galactosidase expression was quantitated using the chemiluminescent substrate GalactoLite Plus (Tropix), and the percentage of fusion relative to the wild-type CD4sp-GPC complex (CD4sp⫹SSP) is indicated. Error bars repre-sent⫾1 standard deviation.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.86.242.492.641.2]proposed intersubunit zinc center. Because the C-terminal re-gion of SSP is short and otherwise permissive to amino acid changes (64), it appears that proper placement of the coordi-nating C57 side chain is promoted by additional sequence elements that lie in the N-terminal region of SSP (54) or the membrane-spanning domains of SSP and G2. Indeed, muta-tions in the second transmembrane domain of SSP that prevent SSP association (2) might do so by perturbing the positioning of C57 and thereby preventing formation of the intersubunit zinc center. Structural studies are needed in order to further assess the potential for zinc binding by H459, C467, and C469 and the proposed participation of C57 in SSP.
We should emphasize that specific interactions between SSP and G2 are required for the membrane fusion activity of GP-C. Recent studies have shown that mutations that diminish posi-tive charge at K33 in the ectodomain loop of SSP increase the H⫹concentration needed for the activation of membrane fu-sion (65). These results suggest that that K33 interacts with titratable residues in the ectodomain of G2 to modulate fusion activation at acidic pH (65). Formation of a zinc-finger-like structure on the cytosolic face of GP-C may be important in positioning K33 and the ectodomain loop of SSP for this role. In this regard, the localized interaction between SSP and the G2 ectodomain in membrane fusion may provide a tractable model for studying pH-induced activation of viral fusion and the structural determinants on either side of the membrane that control this fundamental function of virus envelope gly-coproteins.
In proposing an intersubunit ZBD, we suggest a molecular basis for the retention and positioning of SSP in the GP-C complex for its role in pH-dependent membrane fusion. With this report, the three structural proteins of the arenaviruses are now known to contain a ZBD (52, 61). Biophysical and struc-tural studies of the unusual ZBD in GP-C are needed to define the zinc centers and characterize the requirements for folding. A detailed understanding of the structure and function of the unique SSP subunit in GP-C may suggest novel opportunities for therapeutic intervention in arenavirus infection and hemor-rhagic fevers.
ACKNOWLEDGMENTS
We are grateful to Michele McGuirl and Michael Machczynski (The University of Montana) for their guidance in bioinorganic protein chemistry and to Min Lu (Weill Medical College of Cornell Univer-sity) and Meg Trahey (The University of Montana) for critical review of the manuscript. We thank James R. Thomas and the St. Patrick Hospital and Health Sciences Center (Missoula, MT) for generous access to a multichannel gamma counter and the NIH Biodefense and Emerging Infections Research Resources Repository for providing MAbs.
This work was supported by NIH research grant AI059355 and a subaward from the Rocky Mountain Regional Center of Excellence for Biodefense and Emerging Infectious Diseases (NIH grant U54 AI065357).
REFERENCES
1.Agnihothram, S. S., J. York, and J. H. Nunberg.2006. Role of the stable signal peptide and cytoplasmic domain of G2 in regulating intracellular transport of the Junin virus envelope glycoprotein complex. J. Virol.80:
5189–5198.
2.Agnihothram, S. S., J. York, M. Trahey, and J. H. Nunberg.2007. Bitopic membrane topology of the stable signal peptide in the tripartite Junı´n virus GP-C envelope glycoprotein complex. J. Virol.81:4331–4337.
3.Berg, J. M., and D. L. Merkle.1989. On the metal specificity of “zinc finger” proteins. J. Am. Chem. Soc.111:3759–3761.
4.Beyer, W. R., D. Popplau, W. Garten, D. von Laer, and O. Lenz.2003. Endoproteolytic processing of the lymphocytic choriomeningitis virus glyco-protein by the subtilase SKI-1/S1P. J. Virol.77:2866–2872.
5.Bolken, T. C., S. Laquerre, Y. Zhang, T. R. Bailey, D. C. Pevear, S. S. Kickner, L. E. Sperzel, K. F. Jones, T. K. Warren, S. A. Lund, D. L. Kirkwood-Watts, D. S. King, A. C. Shurtleff, M. C. Guttieri, Y. Deng, M. Bleam, and D. E. Hruby.2006. Identification and characterization of potent small molecule inhibitor of hemorrhagic fever New World arenaviruses. Antivir. Res.69:86–89.
6.Bombarda, E., H. Cherradi, N. Morellet, B. P. Roques, and Y. Mely.2002. Zn2⫹binding properties of single-point mutants of the C-terminal zinc finger of the HIV-1 nucleocapsid protein: evidence of a critical role of cysteine 49 in Zn2⫹dissociation. Biochemistry41:4312–4320.
7.Borrow, P., and M. B. A. Oldstone.1994. Mechanism of lymphocytic chorio-meningitis virus entry into cells. Virology198:1–9.
8.Buchmeier, M. J., M. D. Bowen, and C. J. Peters.2001. Arenaviruses and their replication, p. 1635–1668.InD. M. Knipe and P. M. Howley (ed.), Fields virology, vol. 2. Lippincott Williams & Wilkins, Philadelphia, PA. 9.Cao, W., M. D. Henry, P. Borrow, H. Yamada, J. H. Elder, E. V. Ravkov, S. T.
Nichol, R. W. Compans, K. P. Campbell, and M. B. A. Oldstone.1998. Identification of alpha-dystroglycan as a receptor for lymphocytic chorio-meningitis virus and Lassa fever virus. Science282:2079–2081.
10.Centers for Disease Control and Prevention.2000. Fatal illnesses associated with a new world arenavirus—California, 1999–2000. Morb. Mortal. Wkly. Rep.49:709–711.
11.Di Simone, C., M. A. Zandonatti, and M. J. Buchmeier.1994. Acidic pH triggers LCMV membrane fusion activity and conformational change in the glycoprotein spike. Virology198:455–465.
12.Earp, L. J., S. E. Delos, H. E. Park, and J. M. White.2005. The many mechanisms of viral membrane fusion proteins. Curr. Top. Microbiol. Im-munol.285:25–66.
13.Eckert, D. M., and P. S. Kim.2001. Mechanisms of viral membrane fusion and its inhibition. Annu. Rev. Biochem.70:777–810.
14.Edelhoch, H.1967. Spectroscopic determination of tryptophan and tyrosine in proteins. Biochemistry6:1948–1954.
15.Eichler, R., O. Lenz, T. Strecker, M. Eickmann, H. D. Klenk, and W. Garten.
2003. Identification of Lassa virus glycoprotein signal peptide as a trans-acting maturation factor. EMBO Rep.4:1084–1088.
16.Eichler, R., O. Lenz, T. Strecker, and W. Garten.2003. Signal peptide of Lassa virus glycoprotein GP-C exhibits an unusual length. FEBS Lett.538:
203–206.
17.Enria, D. A., A. M. Briggiler, N. J. Fernandez, S. C. Levis, and J. I. Maiztegui.
1984. Importance of dose of neutralising antibodies in treatment of Argentine haemorrhagic fever with immune plasma. Lancetii:255–256.
18.Eschli, B., K. Quirin, A. Wepf, J. Weber, R. Zinkernagel, and H. Hengartner.
2006. Identification of an N-terminal trimeric coiled-coil core within arena-virus glycoprotein 2 permits assignment to class I viral fusion proteins. J. Virol.80:5897–5907.
19.Froeschke, M., M. Basler, M. Groettrup, and B. Dobberstein.2003. Long-lived signal peptide of lymphocytic choriomeningitis virus glycoprotein pGP-C. J. Biol. Chem.278:41914–41920.
20.Fuerst, T. R., E. G. Niles, F. W. Studier, and B. Moss.1986. Eukaryotic transient-expression system based on recombinant vaccinia virus that syn-thesizes bacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. USA
83:8122–8126.
21.Gallaher, W. R., C. DiSimone, and M. J. Buchmeier.2001. The viral trans-membrane superfamily: possible divergence of Arenavirus and Filovirus glycoproteins from a common RNA virus ancestor. BMC Microbiol.1:1. 22.Gamsjaeger, R., C. K. Liew, F. E. Loughlin, M. Crossley, and J. P. Mackay.
2007. Sticky fingers: zinc-fingers as protein-recognition motifs. Trends Bio-chem. Sci.32:63–70.
23.Getz, E. B., M. Xiao, T. Chakrabarty, R. Cooke, and P. R. Selvin.1999. A comparison between the sulfhydryl reductants tris(2-carboxyethyl)phosphine and dithiothreitol for use in protein biochemistry. Anal. Biochem.273:73–80. 24.Ghiringhelli, P. D., R. V. Rivera-Pomar, M. E. Lozano, O. Grau, and V. Romanowski.1991. Molecular organization of Junin virus S RNA: complete nucleotide sequence, relationship with other members of the Arenaviridae and unusual secondary structures. J. Gen. Virol.72:2129–2141.
25.Green, L. M., and J. M. Berg.1989. A retroviral Cys-Xaa2-Cys-Xaa4-His-Xaa4-Cys peptide binds metal ions: spectroscopic studies and a proposed three-dimensional structure. Proc. Natl. Acad. Sci. USA86:4047–4051. 26.Ha, N. C., B. C. Oh, S. Shin, H. J. Kim, T. K. Oh, Y. O. Kim, K. Y. Choi, and
B. H. Oh.2000. Crystal structures of a novel, thermostable phytase in par-tially and fully calcium-loaded states. Nat. Struct. Biol.7:147–153. 27.Harding, M. M.2004. The architecture of metal coordination groups in
proteins. Acta Crystallogr. D Biol. Crystallogr.60:849–859.
28.Harrison, S. C.2005. Mechanism of membrane fusion by viral envelope proteins. Adv. Virus Res.64:231–261.
29.Hughson, F. M.1997. Enveloped viruses: a common mode of membrane fusion? Curr. Biol.7:R565–R569.
30.Hunt, J. B., S. H. Neece, and A. Ginsburg.1985. The use of
on November 8, 2019 by guest
http://jvi.asm.org/
resorcinol in studies of zinc release from Escherichia coli aspartate transcarbamoylase. Anal. Biochem.146:150–157.
31.Klug, A., and J. W. Schwabe.1995. Protein motifs 5. Zinc fingers. FASEB J.
9:597–604.
32.Krezel, A., R. Latajka, G. D. Bujacz, and W. Bal.2003. Coordination prop-erties of tris(2-carboxyethyl)phosphine, a newly introduced thiol reductant, and its oxide. Inorg. Chem.42:1994–2003.
33.Krishna, S. S., I. Majumdar, and N. V. Grishin.2003. Structural classifica-tion of zinc fingers: survey and summary. Nucleic Acids Res.31:532–550. 34.Kunz, S., P. Borrow, and M. B. A. Oldstone.2002. Receptor structure,
binding, and cell entry of arenaviruses. Curr. Top. Microbiol. Immunol.
262:111–137.
35.Kunz, S., K. H. Edelmann, J.-C. de la Torre, R. Gorney, and M. B. A. Oldstone.2003. Mechanisms for lymphocytic choriomeningitis virus glyco-protein cleavage, transport, and incorporation into virions. Virology314:
168–178.
36.Lachenmann, M. J., J. E. Ladbury, J. Dong, K. Huang, P. Carey, and M. A. Weiss.2004. Why zinc fingers prefer zinc: ligand-field symmetry and the hidden thermodynamics of metal ion selectivity. Biochemistry43:13910– 13925.
37.Ladant, D.1995. Calcium and membrane binding properties of bovine neu-rocalcin delta expressed in Escherichia coli. J. Biol. Chem.270:3179–3185. 38.Laity, J. H., B. M. Lee, and P. E. Wright.2001. Zinc finger proteins: new insights into structural and functional diversity. Curr. Opin. Struct. Biol.
11:39–44.
39.Lenz, O., J. ter Meulen, H.-D. Klenk, N. G. Seidah, and W. Garten.2001. The Lassa virus glycoprotein precursor GP-C is proteolytically processed by subtilase SKI-1/S1P. Proc. Natl. Acad. Sci. USA98:12701–12705. 40.Mackay, J. P., and M. Crossley.1998. Zinc fingers are sticking together.
Trends Biochem. Sci.23:1–4.
41.Matthews, J. M., and M. Sunde.2002. Zinc fingers—folds for many occa-sions. IUBMB Life54:351–355.
42.McCall, K. A., and C. A. Fierke.2000. Colorimetric and fluorimetric assays to quantitate micromolar concentrations of transition metals. Anal. Bio-chem.284:307–315.
43.McCormick, J. B., and S. P. Fisher-Hoch.2002. Lassa fever. Curr. Top. Microbiol. Immunol.262:75–109.
44.McCormick, J. B., P. A. Webb, J. W. Krebs, K. M. Johnson, and E. S. Smith.
1987. A prospective study of the epidemiology and ecology of Lassa fever. J. Infect. Dis.155:437–444.
45.Murphy, F. A., P. A. Webb, K. M. Johnson, S. G. Whitfield, and W. A. Chappell.1970. Arenoviruses in Vero cells: ultrastructural studies. J. Virol.
6:507–518.
46.Nussbaum, O., C. C. Broder, and E. A. Berger.1994. Fusogenic mechanisms of enveloped-virus glycoproteins analyzed by a novel recombinant vaccinia virus-based assay quantitating cell fusion-dependent reporter gene activa-tion. J. Virol.68:5411–5422.
47.Paterson, R. G., G. P. Leser, M. A. Shaughnessy, and R. A. Lamb.1995. The paramyxovirus SV5 V protein binds two atoms of zinc and is a structural component of virions. Virology208:121–131.
48.Perez, M., R. C. Craven, and J. C. de la Torre.2003. The small RING finger protein Z drives arenavirus budding: implications for antiviral strategies. Proc. Natl. Acad. Sci. USA100:12978–12983.
49.Peters, C. J.2002. Human infection with arenaviruses in the Americas. Curr. Top. Microbiol. Immunol.262:65–74.
50.Radoshitzky, S. R., J. Abraham, C. F. Spiropoulou, J. H. Kuhn, D. Nguyen, W. Li, J. Nagel, P. J. Schmidt, J. H. Nunberg, N. C. Andrews, M. Farzan, and
H. Choe.2007. Transferrin receptor 1 is a cellular receptor for New World haemorrhagic fever arenaviruses. Nature446:92–96.
51.Salazar-Bravo, J., L. A. Ruedas, and T. L. Yates.2002. Mammalian reser-voirs of arenaviruses. Curr. Top. Microbiol. Immunol.262:25–63. 52.Salvato, M. S., K. J. Schweighofer, J. Burns, and E. M. Shimomaye.1992.
Biochemical and immunological evidence that the 11 kDa zinc-binding pro-tein of lymphocytic choriomeningitis virus is a structural component of the virus. Virus Res.22:185–198.
53.Sanchez, A., D. Y. Pifat, R. H. Kenyon, C. J. Peters, J. B. McCormick, and M. P. Kiley.1989. Junin virus monoclonal antibodies: characterization and cross-reactivity with other arenaviruses. J. Gen. Virol.70:1125–1132. 54.Saunders, A. A., J. P. Ting, J. Meisner, B. W. Neuman, M. Perez, J. C. de la
Torre, and M. J. Buchmeier.2007. Mapping the landscape of the lympho-cytic choriomeningitis virus stable signal peptide reveals novel functional domains. J. Virol.81:5649–5657.
55.Shi, Y., R. D. Beger, and J. M. Berg.1993. Metal binding properties of single amino acid deletion mutants of zinc finger peptides: studies using cobalt(II) as a spectroscopic probe. Biophys. J.64:749–753.
56.Skehel, J. J., and D. C. Wiley.2000. Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annu. Rev. Biochem.69:531– 569.
57.South, T. L., P. R. Blake, R. C. Sowder III, L. O. Arthur, L. E. Henderson, and M. F. Summers.1990. The nucleocapsid protein isolated from HIV-1 particles binds zinc and forms retroviral-type zinc fingers. Biochemistry29:
7786–7789.
58.Spiropoulou, C. F., S. Kunz, P. E. Rollin, K. P. Campbell, and M. B. A. Oldstone.2002. New World arenavirus clade C, but not clade A and B viruses, utilizes alpha-dystroglycan as its major receptor. J. Virol.76:5140– 5146.
59.Strecker, T., R. Eichler, J. ter Meulen, W. Weissenhorn, H. D. Klenk, W. Garten, and O. Lenz.2003. Lassa virus Z protein is a matrix protein and sufficient for the release of virus-like particle. J. Virol.77:10700–10705. 60.Tam, R. C., J. Y. Lau, and Z. Hong.2001. Mechanisms of action of ribavirin
in antiviral therapies. Antivir. Chem. Chemother.12:261–272.
61.Tortorici, M. A., P. D. Ghiringhelli, M. E. Lozano, C. G. Albarino, and V. Romanowski.2001. Zinc-binding properties of Junin virus nucleocapsid pro-tein. J. Gen. Virol.82:121–128.
62.Tucker, P. A., D. Tsernoglou, A. D. Tucker, F. E. Coenjaerts, H. Leenders, and P. C. van der Vliet.1994. Crystal structure of the adenovirus DNA binding protein reveals a hook-on model for cooperative DNA binding. EMBO J.13:2994–3002.
63.York, J., S. S. Agnihothram, V. Romanowski, and J. H. Nunberg.2005. Genetic analysis of heptad-repeat regions in the G2 fusion subunit of the Junin arenavirus envelope glycoprotein. Virology343:267–279.
64.York, J., and J. H. Nunberg.2007. Distinct requirements for signal peptidase processing and function of the stable signal peptide (SSP) subunit in the Junin virus envelope glycoprotein. Virology359:72–81.
65.York, J., and J. H. Nunberg.2006. Role of the stable signal peptide of the Junı´n arenavirus envelope glycoprotein in pH-dependent membrane fusion. J. Virol.80:7775–7780.
66.York, J., V. Romanowski, M. Lu, and J. H. Nunberg.2004. The signal peptide of the Junı´n arenavirus envelope glycoprotein is myristoylated and forms an essential subunit of the mature G1-G2 complex. J. Virol.78:10783–10792. 67.Zheng, R., T. M. Jenkins, and R. Craigie.1996. Zinc folds the N-terminal
domain of HIV-1 integrase, promotes multimerization, and enhances cata-lytic activity. Proc. Natl. Acad. Sci. USA93:13659–13664.