Hepatitis B Virus Replication in Mice Expressing A2 and
HLA-DR1 Molecules
Sarah Dion,a,bMaryline Bourgine,a,bOphélie Godon,a,bFlorence Levillayer,aMarie-Louise Michela,b Institut Pasteur, Laboratoire de Pathogenèse des Virus de l’Hépatite B, Département de Virologie, Paris, Francea
; INSERM, U845, Paris, Franceb
Hepatitis B virus (HBV) persistence may be due to impaired HBV-specific immune responses being unable to eliminate effi-ciently or cure infected hepatocytes. The immune mechanisms that lead to HBV persistence have not been completely identified, and no appropriate animal model is available for such studies. Therefore, we established a chronic HBV infection model in a mouse strain with human leukocyte antigen A2/DR1 (HLA-A2/DR1) transgenes and an H-2 class I/class II knockout. The liver of these mice was transduced with adeno-associated virus serotype 2/8 (AAV2/8) carrying a replication-competent HBV DNA ge-nome. In all AAV2/8-transduced mice, hepatitis B virus surface antigen, hepatitis B virus e antigen, and HBV DNA persisted in serum for at least 1 year. Viral replication intermediates and transcripts were detected in the livers of the AAV-injected mice. The hepatitis B core antigen was expressed in 60% of hepatocytes. No significant inflammation was observed in the liver. This was linked to a higher number of regulatory T cells in liver than in controls and a defect in HBV-specific functional T-cell responses. Despite the substantial tolerance resulting from expression of HBV antigens in hepatocytes, we succeeded in priming functional HBV-specific T-cell responses in peripheral tissues, which subsequently reached the liver. This AAV2/8-HBV-transduced HLA-A2/DR1 murine model recapitulates virological and immunological characteristics of chronic HBV infection, and it could be useful for the development of new treatments and immune-based therapies or therapeutic vaccines for chronic HBV infections.
H
epatitis B virus (HBV) infection is a major health problem. There are more than 350 million chronic carriers worldwide, and they are at high risk of developing liver cirrhosis and hepato-cellular carcinoma (1). Chronic HBV infection is the result of impaired HBV-specific immune responses such that the infected hepatocytes cannot be eliminated or cured efficiently, but many of the associated issues remain unclear (2,3).Due to the paucity ofin vitro andin vivomodels for HBV infection, HBV-transgenic mice are the most widely used model. These mice have the viral genome integrated into the chromo-some and produce infectious HBV particles or viral antigens in the liver; however, the main limitation of HBV-transgenic mouse models is that they are immunologically tolerant to viral antigens (4,5). Various routes have been exploited to introduce the HBV genome into the hepatocytes of adult mice. One is to introduce a replication-competent HBV genome into the mouse liver by hy-drodynamic injection (HDI) through the tail vein (6); although HBV replicates in the mouse liver, the virus is rapidly cleared by immune responses against HBV proteins (7). Recently, Huang and colleagues used HDI to create a nontransgenic model of per-sistent HBV replication (8). The virus persisted in 40% of mice or was eliminated according to the genetic background. These mice rapidly develop anti-hepatitis B virus core (HBc) antibody, which is the first serological marker of acute HBV infection in humans. An alternative method uses adenoviral vectors to transfer 1.3 cop-ies of the HBV genome into immunocompetent mice (9,10), and acute or chronic HBV infection was obtained depending on the dose of adenoviral vector injected.
Here, we describe an alternative murine model for the study of HBV persistence based on the liver-targeted transduction of ad-eno-associated virus serotype 2/8 (AAV2/8). We produced an AAV2/8 construct carrying a replication-competent HBV DNA genome and by intravenous injection established a model of HBV
persistence in humanized HLA-A2/DR1 immunocompetent mice. Hepatitis B virus surface antigen (HBsAg), hepatitis B virus e antigen (HBeAg), and HBV DNA persisted for at least 1 year in sera of all AAV2/8-injected mice, and viral replication intermedi-ates and transcripts were detected in their livers. HBcAg was ex-pressed in 60% of hepatocytes without significant inflammation in the liver. The persistence of infection was associated with the presence of regulatory T cells (Tregs) in the liver. This mouse model of HBV persistence recapitulates viral and histological characteristics of human chronic HBV infection in the immune-tolerant stage of the disease (11,12).
In HLA-A2/DR1 mice, cellular immune responses were com-pletely restricted to HLA molecules. Antibody, T-helper, and cy-totoxic-T-lymphocyte responses to vaccination with recombinant HBsAg or HBsAg-expressing DNA were similar to those in vacci-nated humans (13, 14) or in HBV-infected individuals (15). Therefore, this AAV2/8-HBV-transduced HLA-A2/DR1 murine model may be useful for the development of therapeutic strategies that require cooperation between HLA-restricted CD4⫹T-helper cells and cytolytic CD8⫹T cells to clear HBV during viral persis-tence.
To our knowledge, this is the first mouse model with a cellular immune response restricted to human major histocompatibility complex (MHC) molecules in the context of chronic HBV
infec-Received9 November 2012Accepted28 February 2013
Published ahead of print6 March 2013
Address correspondence to Marie-Louise Michel, [email protected].
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.03134-12
on November 7, 2019 by guest
http://jvi.asm.org/
tion that allows the evaluation of new treatments and immune-based therapies or therapeutic vaccines.
MATERIALS AND METHODS
AAV2/8-HBV vector.A recombinant AAV2/8 vector carrying 1.2 copies of the HBV genome (genotype D) was constructed. Virus stocks were produced and titrated by the Plateforme de Thérapie Génique in Nantes, France (INSERM U1089), with a protocol derived from one described previously by Ayuso et al. (16). Briefly, a 1.2 copy of the HBV genome was excised by PvuII digestion from the payw 1.2 plasmid (kindly provided by M. Melegari [17]) and ligated between the inverted terminal repeats (ITRs) of AAV2, resulting in pAAV-HBV. The 1.2-full-length HBV ge-nome flanked by AAV2 ITRs was packaged into capsids from AAV8 after cotransfection of HEK 293 cells with pAAV-HBV and an auxiliary plas-mid, leading to the AAV2/8-HBV viral vector. A stock of AAV2/8 without any transgene, referred to as AAV2/8-empty, was also produced and pu-rified under the same conditions. The vector stocks were titrated as virus genomes (vg) per milliliter and stored at⫺80°C until use.
DNA vectors and peptides.Plasmid DNA was endotoxin free and manufactured by PlasmidFactory (Germany). pCMV-S2.S ayw carries the gene encoding the pre-S2 and S domains of HBsAg (genotype D), and their expression is controlled by the cytomegalovirus immediate-early gene promoter (18). pCMV-HBc encodes the HBV capsid carry-ing HBcAg.
A set of synthetic HLA-A2- and HLA-DR1-restricted peptides derived from HBs and HBc antigens was purchased from Polypeptide Group (France) and used at a final concentration of 1g/ml. Peptide purity was ⬎80%.
Animal procedures. Six- to eight-week-old HLA-A*0201/ DRB1*0101-transgenic, H-2 class I/class II knockout (KO) mice (here referred to as HLA-A2/DR1) were used in this study and were previously described (13). All animals were housed in a specific-pathogen-free envi-ronment in the animal facilities of the Institut Pasteur, Paris, France. All protocols have been reviewed and approved by the institutional animal care committee of the Institut Pasteur for compliance with French and European regulations on animal welfare and with Public Health Service recommendations. Mice received a single tail vein injection of 5⫻1010vg of the AAV2/8-HBV vector or AAV2/8-empty vector or an equivalent volume of phosphate-buffered saline (PBS). Serum samples were ob-tained from mice by retro-orbital puncture at various times after injec-tion. Groups (4 to 6 mice/group) were immunized once by bilateral in-tramuscular (i.m.) injection of 100g of DNA into cardiotoxin-treated tibialis anterior muscles, as previously described (18), under anesthesia (100l of 12.5 mg/ml ketamine and 1.25 mg/ml xylazine).
Histological assays and immunohistochemical staining.Liver tis-sues were collected from mice and fixed in 3.7% formaldehyde. Each liver was divided into six parts that were individually embedded in paraffin and sectioned. Liver sections (4m thick) were stained with hematoxylin-eosin or immunostained with a rabbit anti-HBcAg primary antibody (Dako, France). HBcAg-positive hepatocytes were analyzed by using Im-ageJ software (version 1.43; W. S. Rasband, U.S. NIH, Bethesda, MD). The percentage of HBcAg-positive hepatocytes was calculated as (HBcAg-pos-itive hepatocytes/total hepatocytes)⫻100.
Detection of HBV antigens, antibodies, and serum aminotrans-ferases.HBsAg and HBeAg in mouse sera were assayed with commercial enzyme-linked immunosorbent assay (ELISA) kits (Bio-Rad, France). The HBsAg concentration was calculated ing/ml by reference to a stan-dard curve established with known concentrations of HBsAg. Serum HBeAg levels were determined in 10-fold-diluted sera. The threshold value for HBeAg positivity was defined as an optical density of the sample/ cutoff (S/CO) ratio ofⱖ1. Antibodies were quantified by ELISA as previ-ously described (19). Alanine aminotransferase (ALAT) and aspartate aminotransferase (ASAT) activities in sera of AAV2/8-HBV- and PBS-injected mice were determined by the Laboratoire de Biologie Vétérinaire Vebiotel (France). Results are expressed as U/liter.
Quantification of HBV viremia. Virions from mouse sera were immunoprecipitated with a monoclonal anti-HBV pre-S1 antibody (MA18/7; kindly provided by W. H. Gerlich, Giessen, Germany) and quantified by quantitative PCR as previously described (20). Results are expressed as copies/ml. The threshold of detection was 102copies/ml.
Northern and Southern blot analyses.Northern and Southern blot analysis of total liver RNA and total genomic liver DNA were used to detect HBV RNAs and replicative HBV DNA intermediates, respectively, as previously described (21).
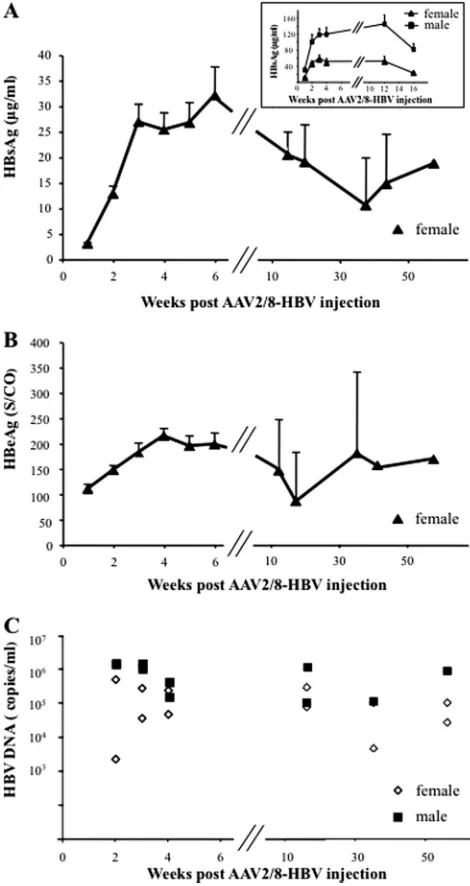
FIG 1Persistence of HBV antigens and HBV DNA in sera of AAV2/8-HBV-injected mice. A total of 5⫻1010vg of AAV2/8-HBV was administered into
female or male HLA-A2/DR1 mice by injection into the tail vein. (A) HBsAg in sera was assayed at the indicated time points, and the results are expressed as g/ml. (Inset) Kinetics of HBsAg levels in sera of male and female mice during the first 16 weeks post-AAV2/8-HBV injection. (B) Serum HBeAg levels are expressed as an S/CO ratio. The values reported are means⫾SEM. (C) HBV DNA titers, expressed as copies/ml, were determined at the indicated times postinjection in sera from individual female or male mice. Results are repre-sentative of three independent experiments.
AAV-Mediated HBV Replication in HLA-Transgenic Mice
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.300.535.68.511.2]Gamma interferon ELISPOT assay.Gamma interferon (IFN-␥ )-pro-ducing T cells among the splenocytes were quantified by an enzyme-linked immunosorbent spot (ELISPOT) assay after peptide stimulation for 24 h, as previously described (15). Wells containing cells (1⫻106) in
culture medium without peptide stimulation were used as negative con-trols to determine background levels. A Bioreader 4000 counter (BioSys, Germany) was used to score the number of spots. Each cell population was titrated in triplicate. The response was considered positive if the median number of spot-forming cells in triplicate wells was at least twice that observed in control wells and at least 10 spot-forming cells per million splenocytes were detected after subtraction of the background.
Flow cytometry.Isolated cells (1⫻106) from spleen and liver were
incubated for 1 h either with medium alone or with peptide pools. Brefel-din A (Sigma-Aldrich, France) was added to a final concentration of 2 g/ml, and the cultures were incubated overnight at 37°C. Cells were harvested, washed, and surface stained with the following monoclonal antibodies: anti-CD3 (145-2C11), anti-CD4 (GK1.5), and anti-CD8 (53-6.7). A viability marker was also used (Live/Dead Fixable Dead Cell Yellow kit; Life Technologies, Frederick, MD). Surface-stained cells were fixed and permeabilized (BD Cytofix/Cytoperm Fixation/Permeabilization kit; BD Biosciences, France) and probed for intracellular cytokines with monoclonal anti-IFN-␥ (XMG1.2), anti-interleukin-2 (IL-2) (JES6-5H4), or anti-tumor necrosis factor alpha (TNF-␣) (MP6-XT22) anti-bodies. The Treg population was evaluated by surface staining with monoclonal anti-CD4 (L3T4) and anti-CD25 (PC61) antibodies and in-tracellular staining with anti-Foxp3 (MF23) antibodies. CD4⫹CD25⫹ IL-10⫹Treg cells in liver leukocyte or splenocyte preparations (2⫻105
cells/well) were enumerated after a 96-h culture in 96-well plates pre-coated with anti-CD3 antibody (145-2C11) in the presence of 1g/ml soluble anti-CD28 antibody (37-51) and 100 IU/ml human IL-2 (Roche, France). Cultured cells were stained with antibodies to CD4, CD25, and IL-10 (JES5-16E3). All antibodies were purchased from BD Biosciences (France). Stained cells were analyzed on a CyAn ADP analyzer (Beckman
Coulter, Miami, FL). Data were analyzed by using FlowJo software (Tree-Star, San Carlos, CA).
Statistical analysis.Data are expressed as means⫾standard errors of the means (SEM). The Mann-Whitney U test was used for nonparametric unpaired comparisons. Statistical analysis was carried out by using Graphpad Prism 5 software (Graphpad, San Diego, CA).P values of ⬍0.05 were considered significant.
RESULTS
Liver-targeted expression of HBV and persistence of HBV DNA
and antigens in sera of AAV2/8-HBV-injected mice.To ensure
the targeting of the HBV genome to the liver, we used an AAV serotype 8 vector that displays liver-specific transduction (22). We cloned 1.2 copies of the HBV genome between the two ITRs of AAV serotype 2 in a carrier plasmid, which was then used to pro-duce the hybrid AAV2/8-HBV vector. Six- to eight-week-old
FIG 2HBV replication and expression in the liver of AAV2/8-HBV-injected mice. Liver tissues were collected from HLA-A2/DR1 mice after PBS (lane 1) or AAV2/8-empty (lane 2) injection or 6 weeks, 12 weeks, and 1 year after AAV2/8-HBV injection (lanes 3, 4, and 5, respectively). (A) Intermediates of HBV DNA replication were detected by Southern blotting. The relaxed-circu-lar (RC), double-stranded linear (DL), and single-stranded (SS) DNAs were revealed. (B) Northern blotting to detect 3.5-kb pregenomic and 2.4- and 2.1-kb envelope mRNAs.
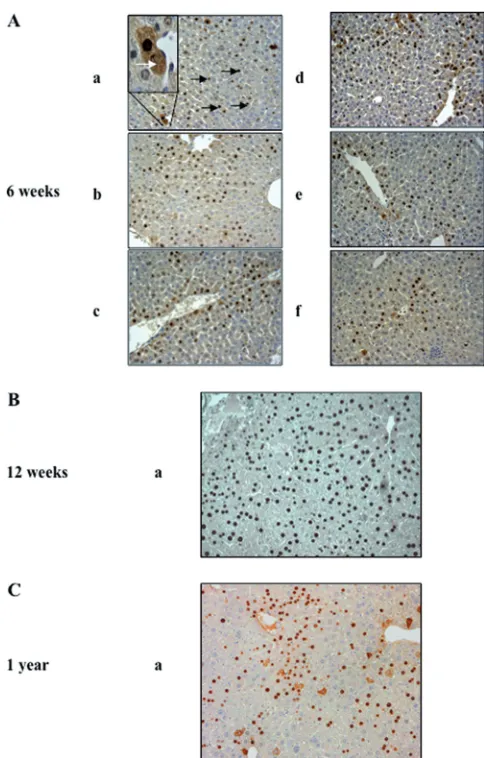
FIG 3Expression of HBcAg in the liver of AAV2/8-HBV-injected mice. Shown is immunohistochemical staining for HBcAg in liver sections from mice injected with 5⫻1010vg of AAV2/8-HBV at week 6 (A), week 12 (B), or
1 year (C) postinjection. Different parts of the liver were analyzed: left lateral lobe (a), left medial lobe (b), right medial lobe (c), right lateral lobe (d), caudate lobe (e), and papillary process (f). Nuclear and cytoplasmic localiza-tions of HBcAg are indicated by black and white arrows, respectively, on an enlarged magnification in panel Aa. Original magnification,⫻250. Results for one representative mouse are shown.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.298.540.63.442.2] [image:3.585.93.237.67.288.2]HLA-A2/DR1 female or male mice were given a single tail vein injection of 5⫻1010vg of the AAV2/8-HBV or the AAV2/8-empty (negative control) vector. Mice were regularly bled, and serum levels of HBsAg, HBeAg, and HBV DNA were determined (Fig. 1). All AAV2/8-empty-injected mice were negative for HBV antigens (data not shown), whereas all AAV2/8-HBV-injected mice were HBsAg and HBeAg positive on day 3 postinjection. In female mice, HBsAg (Fig. 1A) and HBeAg (Fig. 1B) levels increased to a mean concentration of 32.10⫾5.64g/ml and a mean S/CO ratio of 198⫾22.81, respectively, after 6 weeks. The HBsAg and HBeAg levels declined slowly thereafter, but all injected mice remained positive for 56 weeks. HBV antigen titers were higher in sera from male than in sera from female HLA-A2/DR1 mice (Fig. 1A, inset). To exclude detection of the AAV2/8-HBV inoculum, pre-S1-car-rying HBV particles were immunoprecipitated from serum sam-ples and assayed for the presence of HBV DNA by quantitative real-time PCR. HBV particles became detectable 2 weeks after AAV2/8-HBV injection. The virus titers then remained stable un-til at least 1 year postinjection (mean titers were 2.48⫻105and 1.50⫻ 106 copies/ml of HBV DNA in sera from females and males, respectively) (Fig. 1C). Thus, in this immunocompetent HLA-A2/DR1 mouse model, administration of AAV2/8-HBV re-sults in persistent high levels of HBsAg and HBeAg in the sera of all injected mice and is associated with viremia. This pattern is thus similar to what is observed during persistent HBV infection in humans (12).
HBV replication and gene expression in the liver of
AAV2/8-HBV-injected mice.To analyze the kinetics of HBV replication
and transcription, liver samples from AAV2/8-HBV-injected mice were collected at various time points. At 6 weeks, 12 weeks, and 1 year postinjection, HBV DNA replicative intermediates were de-tected by Southern blot analysis (Fig. 2A), and pregenomic
(3.5-kb) and subgenomic (2.4- and 2.1-(3.5-kb) HBV RNA transcripts were detected by Northern blot analysis (Fig. 2B) of liver samples. HBV DNA replicative intermediates and HBV RNA transcripts were still detectable at 1 year but at lower levels. As in HBV-transgenic mice, HBV covalently closed circular DNA (cccDNA) was not detected (data not shown).
Liver tissues were also tested by immunostaining for HBcAg expression (Fig. 3) at 6 weeks, 12 weeks, and 1 year postinjection. The liver of each mouse was divided into six parts to evaluate the HBcAg expression pattern (Fig. 3Aa to f). HBcAg was detected in both cytoplasm and nuclei, but mostly in nuclei, of hepatocytes (Fig. 3Aa, white and black arrows, respectively) in various parts of the liver. At each time point, the pattern of HBcAg expression was similar in the various lobes of the liver considered. HBcAg was expressed in about 60% of the hepatocytes at 6 weeks (Fig. 3A) and 12 weeks (Fig. 3B) post-AAV2/8-HBV injection. HBcAg expres-sion persisted for at least 1 year (Fig. 3C) but was detectable in only around 15% of the hepatocytes at this time.
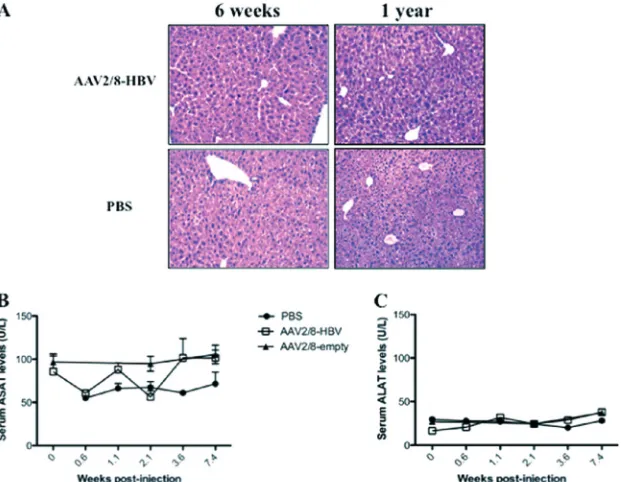
We further examined whether the HBV persistence in the AAV2/8-HBV-injected mice was associated with an inflamma-tory response. Livers from HBV carrier and control mice were stained with hematoxylin and eosin at 6 weeks and 1 year postinjection (Fig. 4A), and mononuclear cell infiltrates were compared. Livers from AAV2/8-HBV-injected mice presented normal architecture, with no obvious inflammatory infiltrates (Fig. 4A). Also, ASAT and ALAT levels were not higher in sera from AAV2/8-HBV-injected mice than in sera from PBS- or AAV2/8-empty-injected mice during the 1-year follow-up (Fig. 4BandCand data not shown). Thus, in our model, long-term expression of HBV antigens and HBV replication in liver following administration of AAV2/8-HBV did not cause appar-ent liver damage.
FIG 4Histological analysis of liver sections and transaminase activities in sera from AAV2/8-HBV-injected mice. (A) Hematoxylin and eosin staining of one representative lobe of liver sections from mice 6 weeks or 1 year after injection of 5⫻1010vg of AAV2/8-HBV or PBS. Original magnification,⫻250. Results are
for one representative mouse of five. (B and C) Mean ASAT (B) and ALAT (C) activities in sera from 10 AAV2/8-HBV-injected, 10 AAV2/8-empty-injected, and 4 PBS-injected mice, expressed as U/liter.
AAV-Mediated HBV Replication in HLA-Transgenic Mice
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.138.453.67.308.2]Defect in the HBcAg- and HBsAg-specific immune response
in HBV carrier mice. Both HBc- and HBs-specific T-cell
re-sponses may contribute to viral clearance (23). Therefore, we tested whether the HBV persistence observed after injection of AAV2/8-HBV into HLA-A2/DR1 mice was due to impaired HBc-or HBs-specific immunity. HLA-A2- HBc-or -DR1-restricted peptides derived from HBV core and envelope proteins were used to detect IFN-␥-producing T cells from spleen and liver from
AAV2/8-HBV-injected mice by an ELISPOT assay (Fig. 5A) and intracel-lular staining for cytokines (Fig. 5B). AAV2/8-empty-injected mice were used as negative controls. pCMV-S2.S and pCMV-HBc DNA immunization of HLA-A2/DR1 mice used as positive con-trols for IFN-␥-producing T cells gave rise to HBV-specific T cells. In contrast, neither HBc- nor HBs-specific IFN-␥-producing T cells were detected in spleen from HBV- or AAV2/8-empty-injected mice at early (6 weeks postinjection) (Fig. 5A) or
FIG 5HBV-specific T-cell responses of AAV2/8-HBV-injected mice. IFN-␥-secreting T cells detected by ELISPOT assay of splenocytes (A) and intracellular cytokine staining for IFN-␥of intrahepatic lymphocytes (B) were obtained from 5 to 10 HLA-A2/DR1 mice injected 6 weeks previously with AAV2/8-HBV or AAV2/8-empty or immunized 2 weeks previously with HBc- and HBs-expressing DNA vectors as positive controls. (A) HLA-A2- and HLA-DR1-restricted peptides derived from either HBcAg (18-27) or HBsAg (111-125, 179-194, 183-191, 200-214, and 348-357) were used to stimulate splenocytesex vivo. (B) Intrahepatic lymphocytes were analyzed after incubation with a pool of HLA-A2-restricted peptides (183-191, 204-212, and 348-357) or in medium alone (unstimulated). Representative fluorescence-activated cell sorter plots for IFN-␥-secreting T cells in the liver of DNA- or AAV2/8-HBV-injected mice are shown. Values in the upper right square of each panel represent the percentage of CD8⫹T cells that are also IFN-␥-positive cells. Results are representative of three independent experiments.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.137.448.68.535.2]late (12 weeks postinjection) (data not shown) time points. Intra-hepatic lymphocytes collected from AAV2/8-HBV-injected and DNA-immunized HLA-A2/DR1 mice were stained for intracellu-lar IFN-␥, TNF-␣, and IL-2 after stimulation or not with pools of HLA-A2/HLA-DR1-restricted HBV peptides (Fig. 5Band data not shown). Neither CD8⫹(Fig. 5B, right) nor CD4⫹(data not shown) HBV-specific T cells producing IFN-␥were detected in lymphocytes purified from liver of HLA-A2/DR1 mice following AAV2/8-HBV injection, in contrast to the findings for the DNA-immunized controls (Fig. 5B, left). Similar results were obtained for HBV-specific T cells producing IL-2 or TNF-␣ (data not shown). Also, anti-HBc and anti-HBs antibodies were not detected in sera of AAV2/8-HBV-injected mice (data not shown). This absence of HBc- or HBs-specific immune responses is in agreement with the persistence of HBV replica-tion in liver.
Numbers of hepatic Tregs and IL-10-producing cells are high
in AAV2/8-HBV-injected mice. To explore the possible role
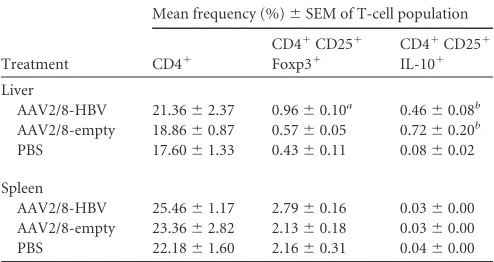
played by Tregs in the maintenance of HBV persistence, liver lym-phocytes and splenocytes were isolated from mice injected with AAV2/8-HBV, AAV2/8-empty, or PBS. The frequencies of CD4⫹ and CD4⫹CD25⫹Foxp3⫹T cells were evaluated by flow cytom-etry (Table 1). Two weeks after vector or PBS administration, the hepatic CD4⫹T-cell frequencies were similar for AAV2/8-HBV-, AAV2/8-empty-, or PBS-injected mice, whereas Tregs were signif-icantly more frequent in liver from AAV2/8-HBV-injected mice than in liver from PBS- or AAV2/8-empty-injected mice (Table 1). CD4⫹ T-cell and Treg frequencies in spleen did not differ between the three groups. To gain additional insights into the Treg response, we characterized their phenotypic changes following vector administration. Liver leukocytes and splenocytes collected from mice 2 weeks after administration of AAV2/8-HBV, AAV2/ 8-empty, or PBS were activatedex vivo. The AAV2/8-HBV and AAV2/8-empty samples contained a subset of CD4⫹CD25⫹T cells producing IL-10 that was absent from the liver of mice re-ceiving PBS (Table 1). However, in spleen samples, the percentage of CD4⫹CD25⫹T cells producing IL-10 did not differ between AAV2/8-HBV-, AAV2/8-empty-, and PBS-injected mice. This in-crease in Treg frequencies in liver was associated with the injection
of AAV expressing HBV antigens, whereas the presence of IL-10-secreting T cells was linked to the AAV vector.
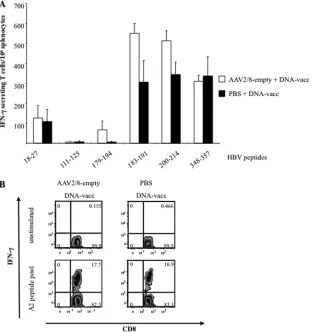
Immunization of AAV2/8-HBV-injected mice with HBc- and HBs-encoding DNAs breaks the observed T-cell tolerance to HBV.The liver microenvironment favors immune tolerance. To determine whether HBV persistence in AAV2/8-HBV-injected mice could be overcome by stimulating immune cells in the pe-riphery, we immunized HBV carrier mice by injection of plasmid DNAs encoding HBV antigens. HLA-A2/DR1 mice were first in-jected with AAV2/8-HBV or PBS, and 32 days later, they were immunized with pCMV-S2.S and pCMV-HBc DNAs via the i.m. route or not immunized. ELISPOT assays with individual pep-tides forex vivostimulation of spleen cells revealed that mice with persistent HBV infection developed a T-cell response to the HBc-derived 18- to 27-amino-acid peptide and HBs-HBc-derived 183-191, 200-214, and 348-357 peptides (Fig. 6A). However, the number of IFN-␥-secreting T cells was considerably lower than that for HLA-A2/DR1 mice receiving PBS instead of AAV2/8-HBV before DNA immunization.
Surprisingly, HBV-specific IFN-␥-producing CD8⫹T cells were detected in the liver of AAV2/8-HBV-injected mice (8.56%⫾4.20%) after DNA immunization, whereas these cells were not found in liver of mice receiving AAV2/8-HBV and PBS only. The mean frequency of CD8⫹ T cells in liver of control mice receiving DNA vaccines only was 4.67%⫾0.99%. However, the proportion of mice with HBV-specific CD8⫹T cells in liver was lower for HBV carrier mice (3/10 mice) than for control mice (7/7) (Fig. 6B).
To determine whether the decreased responses observed in the periphery and liver were due to the tolerogenic effect of the AAV2/8 vectorper se, mice were injected with either the AAV-empty vector or PBS, and 32 days later, they were immunized with pCMV-S2.S and pCMV-HBc DNAs. ELISPOT assays showed that mice receiving the AAV-empty vector before DNA immunization developed comparable IFN-␥-secreting-T-cell responses in the spleen (Fig. 7A). HBV-specific IFN-␥-producing CD8⫹T cells were detected at a similar level in the liver of AAV2/8-empty-injected (20.42%⫾3.12%) or PBS-injected (15.41%⫾6.12%) mice after DNA immunization (Fig. 7B). This suggests that the AAV2/8 vector had no tolerogenic effect on vaccine-induced im-mune responses and that the observed tolerance in AAV-HBV-transduced mice is due to transgene expression.
Although HBV-specific T-cell responses were detected in spleen and liver of HBV carrier mice after immunization, these responses had apparently no effect on HBeAg or HBsAg levels in sera (Fig. 6CandD), on HBcAg intrahepatic expression, or on HBV DNA replicative intermediates when tested at 2 weeks postimmunization (data not shown). Furthermore, this immune response was not associated with an increased level of transami-nase activity (data not shown).
DISCUSSION
[image:6.585.40.287.78.209.2]We report the development of a novel animal model for HBV persistence in an immunocompetent mouse strain expressing HLA-A2/DR1 molecules (13). This involved the use of the hepa-totropic hybrid serotype 2 and 8 AAV vector to deliver the entire HBV genome into mouse hepatocytes. AAV2/8 efficiently trans-duces hepatocytes (22). Mouse liver is permissive for HBV repli-cation (24), so the use of AAV2/8 vectors allows the entry step of HBV infection to be bypassed. We demonstrate that a single tail
TABLE 1Frequencies of hepatic and splenic T-cell populationsc
Treatment
Mean frequency (%)⫾SEM of T-cell population
CD4⫹
CD4⫹CD25⫹ Foxp3⫹
CD4⫹CD25⫹ IL-10⫹
Liver
AAV2/8-HBV 21.36⫾2.37 0.96⫾0.10a 0.46⫾0.08b
AAV2/8-empty 18.86⫾0.87 0.57⫾0.05 0.72⫾0.20b
PBS 17.60⫾1.33 0.43⫾0.11 0.08⫾0.02
Spleen
AAV2/8-HBV 25.46⫾1.17 2.79⫾0.16 0.03⫾0.00 AAV2/8-empty 23.36⫾2.82 2.13⫾0.18 0.03⫾0.00 PBS 22.18⫾1.60 2.16⫾0.31 0.04⫾0.00
a
The percentage of Tregs in AAV2/8-HBV-injected mice is significantly different from that in AAV2/8-empty- or PBS-injected mice (P⬍0.05).
b
The percentages of the CD4⫹CD25⫹IL-10⫹subset in AAV2/8-HBV- and in AAV2/ 8-empty-injected mice are significantly different from those in PBS-injected mice (P⬍ 0.05).
cValues represent the mean percentages⫾SEM of each subset of T cells in the living
lymphocyte population.
AAV-Mediated HBV Replication in HLA-Transgenic Mice
on November 7, 2019 by guest
http://jvi.asm.org/
vein injection of AAV2/8-HBV led to a persistent infection in all injected mice. This infection was characterized by the presence of HBsAg and HBeAg in serum for at least 1 year and the absence of seroconversion to antibodies to HBsAg or HBcAg. Viral particles containing HBV DNA were found in sera, and abundant HBV DNA replicative intermediates and HBV transcripts were found in liver both early (6 to 12 weeks) and late (1 year) after injection. The antigenemia rate and replication levels were higher in males than in females. Thus, our mouse model reproduced the epidemiolog-ical findings that serum HBsAg and HBV DNA levels are always higher in male than in female carriers (25,26). Our protocol re-sults in all injected mice becoming HBV carriers, for at least 1 year, after a single AAV2/8-HBV injection. In a mouse model of HBV persistence developed previously by Huang et al., only 40% of C57BL/6 mice showed the desired characteristics (8). Our model
therefore allows the use of a relatively small number of animals to establish HBV carrier status for future studies. Importantly, HBcAg expression was detected in 60% of hepatocytes in our model, contrasting with the 5 to 10% of hepatocytes expressing HBcAg observed previously after HDI with the HBV genome. In addition, AAV allows homogeneous transduction of the liver, whereas not all parts of the liver are reached after HDI (7). HDI clearly induces liver inflammation, possibly leading to the recruitment of immune cells to the liver and thereby to the clearance of HBV hepatocytes observed in most mouse strains, including ours (7,21). Our mouse model is more similar to the recently described model of HBV persistence based on a low-dose adenovirus vector carrying the HBV genome in immuno-competent C57BL/6 mice (10). However, the kinetics of early HBV antigen expression in sera and liver differ between the two
FIG 6HBV-specific T-cell responses of HBV carrier mice immunized with HBc- and HBs-expressing DNA vectors. (A and B) ELISPOT assays performed on splenocytes (A) and intracellular cytokine staining (B) of intrahepatic lymphocytes from 5 to 10 HLA-A2/DR1 mice immunized with HBc- and HBs-expressing DNA vectors and controls (PBS). Mice were previously injected either with AAV2/8-HBV or with PBS 32 days earlier. (A) Peptides derived from HBcAg (18-27) and HBsAg (111-125, 179-194, 183-191, 200-214, and 348-357) were used to stimulate splenocytesex vivo. (B) Intrahepatic lymphocytes were analyzed for CD8⫹ T cells producing intracellular IFN-␥after incubation with a pool of HLA-A2-restricted peptides (183-191, 204-212, and 348-357) or with medium alone (unstimulated). Representative fluorescence-activated cell sorter plots used to determine the percentages of IFN-␥-secreting T cells present in the liver of mice are shown. Values in the upper right squares are the percentages of CD8⫹T cells that are IFN-␥-positive cells. (C and D) HBeAg (C) and HBsAg (D) titers in sera of mice that were first injected with AAV2/8-HBV and then immunized 32 days later (arrow) with DNA or received PBS. The values reported are means⫾SEM, and results are representative of three independent experiments.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.115.474.65.445.2]models, and HBV antigens seem to persist longer at high levels in the AAV-HBV-injected mice.
Persistence of HBV in our mouse model was not deleterious for the liver and was linked to impaired HBV-specific immunity. In-deed, HBV antigenemia in AAV2/8-HBV-injected mice was high, mimicking the initial stage of natural HBV infection evolving to chronicity, in which significant levels of circulating HBV antigens but no virus-specific immunity is detected (27). The natural his-tory of chronic HBV infection in human has been divided into four different phases defined by virological, immunological, and clinical parameters (28). In our mice, the viral antigen levels in sera could reach approximately 2,500 IU/ml (32 g/ml) for HBsAg and 44 Paul-Ehrlich Institute units/ml (S/CO⫽200) for HBeAg in females and were even higher in males. This corre-sponds to values found in sera of patients in the “immune-toler-ant” phase of the disease (12). In these patients, normal or low levels of ALT and mild or no liver necroinflammation are ob-served, which is also a characteristic of our mouse model. Another feature of these patients is the apparent state of immune tolerance toward the virus, although this was revisited in a recent study (29). In our mice, neither HBc- nor HBs-specific T cells were detected
in the periphery or the liver at either early or late time points after AAV2/8-HBV injection. Given the crucial role of HBsAg- and HBcAg-specific immunity in clearance of HBV (2,23), this lack of antigen-specific immunity is in agreement with the chronic status and may result from the continuous exposure to high levels of HBV antigens in these mice or from priming of T cells in liver. Antigen presentation in liver can skew immune responses toward tolerance and limit the effector functions of T cells (30) or pro-mote T-cell death via the proapoptotic molecule Bim (31).
Liver-directed transgene delivery by AAV vectors in mouse models results in tolerance to a number of the proteins produced, both secreted (32,33) and cytosolic (34). In our model, we ob-served Treg populations larger than those in controls, and these cells may actively suppress the functions of HBV-specific T cells and maintain strong transgene expression. Several studies have addressed the role of Tregs in HBV infection and have shown a correlation of Treg frequencies in peripheral blood and liver with different disease stages (35, 36). Depletion in vitro of CD4⫹ CD25⫹Tregs from peripheral blood mononuclear cell samples taken from HBV-infected patients led to an increase of IFN-␥ production following HBV antigen stimulation (37). Moreover,
FIG 7HBV-specific T-cell responses of mice immunized with HBc- and HBs-expressing DNA vectors. (A) ELISPOT assays performed on splenocytes from 8 HLA-A2/DR1 mice immunized with HBc- and HBs-expressing DNA vectors. Mice were previously (⫺32 days) injected either with AAV2/8-empty or with PBS. The peptides derived from HBcAg (18-27) and HBsAg (111-125, 179-194, 183-191, 200-214, and 348-357) were then used to stimulate splenocytesex vivo. The values reported are means⫾SEM. (B) Intrahepatic lymphocytes were analyzed for CD8⫹T cells producing intracellular IFN-␥after incubation with a pool of HLA-A2-restricted peptides (183-191, 204-212, and 348-357) or with medium alone (unstimulated). Representative fluorescence-activated cell sorter plots used to determine the percentages of IFN-␥-secreting T cells present in the liver of mice are shown. Values in the upper right squares are the percentages of CD8⫹T cells that are IFN-␥-positive cells.
AAV-Mediated HBV Replication in HLA-Transgenic Mice
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.138.456.68.401.2]inhibition of viral replication reduces Treg counts and enhances the antiviral immune response in chronic hepatitis B patients (38). Tregs may also limit liver injury by controlling inflammation. We also found an increased number of T cells producing IL-10 in the liver following AAV transduction. IL-10 is a potent anti-inflam-matory cytokine that inhibits Th1-, Th17-, and Th2-mediated im-mune responses. Production of IL-10 is also associated with reg-ulatory responses such as Tr1 (39). Therefore, both increased numbers of Tregs and IL-10 production in liver may account for the absence of liver inflammation and the inhibition of HBV-specific immune responses in our mouse model. However, using eitherin vivoTreg depletion or a blockade of the IL-10 receptor, we did not find any improvement in HBV-specific immune re-sponses. This was consistent with previous studies that failed to rescue the AAV transgene-induced CD8 T-cell ability to prolifer-ate or to produce cytokines (40). In addition to the suppressive cytokine IL-10, a sustained high viral load (41) and the PD1– PDL-1 and Tim3– galnectin-9 inhibitory pathways are known to actively suppress T-cell responses (42). In our model, a complex interplay between the different pathways of T-cell inhibition may cooperate following AAV-mediated HBV gene expression in the liver, leading to the establishment of a chronic viral infection.
The AAV-HBV-induced persistence model that we have estab-lished is validated in mice transgenic for HLA molecules. Because of its similarities with persistent HBV infection in humans, these mice, in which mouse MHC genes have been replaced by human HLA genes, should be useful as a powerful preclinical model to evaluate antiviral and immunotherapeutic strategies. One key ob-jective of immune therapy of chronic HBV infection is to break the immune tolerance to HBV antigens and to induce functional HBV-specific T-cell responses that are able to clear the virus. In our model, we succeeded in breaking the observed tolerance by inducing HBV-specific T cells, which were found in the periphery and liver. We were able to prime HBV-specific T cells in the pe-riphery by i.m. injection of DNA vaccines encoding HBV antigens; this shows that the persistence of HBV antigens did not lead to the complete inhibition of HBV-specific T cells in our model. Clear-ance of HBV DNA in liver is mediated by at least two mechanisms involving noncytolytic inhibition of viral replication and cytolytic elimination of HBV-infected hepatocytes (43). However, despite the production of IFN-␥by DNA-primed T cells and their local-ization in the liver, the responses were not potent enough to de-crease the HBsAg and HBeAg levels in sera. Stronger vaccine pro-tocols may be needed to eliminate the virus. The increased number of regulatory T cells combined with an insufficient num-ber of HBV functional T cells or the absence of liver inflammation in our model might be responsible for HBV persistence.
We describe here a model of persistent HBV infection in im-munocompetent mice that reproduces many of the immunologi-cal features of patients with chronic HBV infection, notably defec-tive HBV-specific B-cell/T-cell responses and abnormally high numbers of Tregs and T cells producing IL-10. Despite the induc-tion of IFN-␥-secreting T cells following DNA-based vaccination, these mice failed to respond clinically to vaccination, as the vac-cine-activated T cells had no impact on HBV antigen levels, as was observed for patients enrolled in many clinical trials of therapeutic vaccination (44). Nevertheless, our mouse model of HBV persis-tence could be used to investigate the mechanisms underlying persistent infection following viral replication and antigen presen-tation in liver cells and to test potential therapeutic approaches to
counteract them. Finally, the viral vector AAV2/8 used to establish our HBV persistence mouse model could be easily manipulated to change the HBV genotype to allow analysis of, for example, the cross-reactivity of T cells induced by candidate therapeutic vac-cines.
ACKNOWLEDGMENTS
Histological sections and hematoxylin eosin staining were performed by Sophie Berissi (Histochemistry Facilities, INSERM, U845, Paris, France). Microscopy was done by the Microscopy Facilities of the Plateforme d’Imagerie Cellulaire (IFR94, Paris, France). We thank Christine Neu-veut, Sylvie Garcia, Genevieve Inchauspe, and Yu Wei for critical scientific discussions; Jean-François Meritet for advice on HBV DNA quantifica-tion; Yu Chun Lone for the HLA-A2/DR1 mice; and Katherine Kean for linguistic revision of the text.
This work and Sarah Dion’s fellowship were supported by grants from the Institut Carnot, Pasteur-Maladies Infectieuses. The Plateforme de Thérapie Génique of Nantes is supported by the AFM (France).
REFERENCES
1.Ganem D, Prince AM.2004. Hepatitis B virus infection—natural history and clinical consequences. N. Engl. J. Med.350:1118 –1129.
2.Rehermann B, Nascimbeni M.2005. Immunology of hepatitis B virus and hepatitis C virus infection. Nat. Rev. Immunol.5:215–229. 3.Maini MK, Schurich A.2010. The molecular basis of the failed immune
response in chronic HBV: therapeutic implications. J. Hepatol.52:616 – 619.
4.Guidotti LG, Matzke B, Schaller H, Chisari FV.1995. High-level hepa-titis B virus replication in transgenic mice. J. Virol.69:6158 – 6169. 5.Chisari FV.1996. Hepatitis B virus transgenic mice: models of viral
im-munobiology and pathogenesis. Curr. Top. Microbiol. Immunol.206: 149 –173.
6.Liu F, Song Y, Liu D.1999. Hydrodynamics-based transfection in ani-mals by systemic administration of plasmid DNA. Gene Ther.6:1258 – 1266.
7.Yang PL, Althage A, Chung J, Chisari FV.2002. Hydrodynamic injection of viral DNA: a mouse model of acute hepatitis B virus infection. Proc. Natl. Acad. Sci. U. S. A.99:13825–13830.
8.Huang LR, Wu HL, Chen PJ, Chen DS.2006. An immunocompetent mouse model for the tolerance of human chronic hepatitis B virus infec-tion. Proc. Natl. Acad. Sci. U. S. A.103:17862–17867.
9.von Freyend MJ, Untergasser A, Arzberger S, Oberwinkler H, Drebber U, Schirmacher P, Protzer U.2011. Sequential control of hepatitis B virus in a mouse model of acute, self-resolving hepatitis B. J. Viral Hepat.18: 216 –226.
10. Huang LR, Gabel YA, Graf S, Arzberger S, Kurts C, Heikenwalder M, Knolle PA, Protzer U.2012. Transfer of HBV genomes using low doses of adenovirus vectors leads to persistent infection in immune competent mice. Gastroenterology142:1447–1450 e1443.
11. Nguyen T, Thompson AJ, Bowden S, Croagh C, Bell S, Desmond PV, Levy M, Locarnini SA.2010. Hepatitis B surface antigen levels during the natural history of chronic hepatitis B: a perspective on Asia. J. Hepatol.
52:508 –513.
12. Jaroszewicz J, Calle Serrano B, Wursthorn K, Deterding K, Schlue J, Raupach R, Flisiak R, Bock CT, Manns MP, Wedemeyer H, Cornberg M.2010. Hepatitis B surface antigen (HBsAg) levels in the natural history of hepatitis B virus (HBV)-infection: a European perspective. J. Hepatol.
52:514 –522.
13. Pajot A, Michel ML, Fazilleau N, Pancre V, Auriault C, Ojcius DM, Lemonnier FA, Lone YC.2004. A mouse model of human adaptive immune functions: HLA-A2.1-/HLA-DR1-transgenic H-2 class I-/class II-knockout mice. Eur. J. Immunol.34:3060 –3069.
14. Pajot A, Michel ML, Mancini-Bourgine M, Ungeheuer MN, Ojcius DM, Deng Q, Lemonnier FA, Lone YC.2006. Identification of novel HLA-DR1-restricted epitopes from the hepatitis B virus envelope protein in mice expressing HLA-DR1 and vaccinated human subjects. Microbes In-fect.8:2783–2790.
15. Loirat D, Lemonnier FA, Michel ML.2000. Multiepitopic HLA-A*0201-restricted immune response against hepatitis B surface antigen after DNA-based immunization. J. Immunol.165:4748 – 4755.
on November 7, 2019 by guest
http://jvi.asm.org/
16. Ayuso E, Mingozzi F, Bosch F.2010. Production, purification and char-acterization of adeno-associated vectors. Curr. Gene Ther.10:423– 436. 17. Scaglioni PP, Melegari M, Wands JR.1997. Posttranscriptional
regula-tion of hepatitis B virus replicaregula-tion by the precore protein. J. Virol.71: 345–353.
18. Michel ML, Davis HL, Schleef M, Mancini M, Tiollais P, Whalen RG.
1995. DNA-mediated immunization to the hepatitis B surface antigen in mice: aspects of the humoral response mimic hepatitis B viral infection in humans. Proc. Natl. Acad. Sci. U. S. A.92:5307–5311.
19. Mancini M, Hadchouel M, Tiollais P, Pourcel C, Michel ML. 1993. Induction of anti-hepatitis B surface antigen (HBsAg) antibodies in HBsAg producing transgenic mice: a possible way of circumventing “non-response” to HBsAg. J. Med. Virol.39:67–74.
20. Cougot D, Allemand E, Riviere L, Benhenda S, Duroure K, Levillayer F, Muchardt C, Buendia MA, Neuveut C.2012. Inhibition of PP1 phos-phatase activity by HBx: a mechanism for the activation of hepatitis B virus transcription. Sci. Signal.5:ra1. doi:10.1126/scisignal.2001906. 21. Deng Q, Mancini-Bourgine M, Zhang X, Cumont MC, Zhu R, Lone YC,
Michel ML.2009. Hepatitis B virus as a gene delivery vector activating foreign antigenic T cell response that abrogates viral expression in mouse models. Hepatology50:1380 –1391.
22. Cooper M, Nayak S, Hoffman BE, Terhorst C, Cao O, Herzog RW.
2009. Improved induction of immune tolerance to factor IX by hepatic AAV-8 gene transfer. Hum. Gene Ther.20:767–776.
23. Bertoletti A, Ferrari C.2003. Kinetics of the immune response during HBV and HCV infection. Hepatology38:4 –13.
24. Chen SH, Hu CP, Chang CM.1992. Hepatitis B virus replication in well differentiated mouse hepatocyte cell lines immortalized by plasmid DNA. Cancer Res.52:1329 –1335.
25. Chen DS.1993. Natural history of chronic hepatitis B virus infection: new light on an old story. J. Gastroenterol. Hepatol.8:470 – 475.
26. Tian Y, Kuo CF, Chen WL, Ou JH.2012. Enhancement of hepatitis B virus replication by androgen and its receptor in mice. J. Virol.86:1904 – 1910.
27. Guidotti LG, Rochford R, Chung J, Shapiro M, Purcell R, Chisari FV.
1999. Viral clearance without destruction of infected cells during acute HBV infection. Science284:825– 829.
28. European Association for the Study of the Liver.2012. EASL clinical practice guidelines: management of chronic hepatitis B virus infection. J. Hepatol.57:167–185.
29. Kennedy PT, Sandalova E, Jo J, Gill U, Ushiro-Lumb I, Tan AT, Naik S, Foster GR, Bertoletti A.2012. Preserved T-cell function in children and young adults with immune-tolerant chronic hepatitis B. Gastroenter-ology143:637– 645.
30. Protzer U, Maini MK, Knolle PA. 2012. Living in the liver: hepatic infections. Nat. Rev. Immunol.12:201–213.
31. Knolle PA.2012. The liver’s imprint on CD8(⫹) T cell priming. J. Hepa-tol.57:718 –719.
32. Breous E, Somanathan S, Vandenberghe LH, Wilson JM.2009. Hepatic regulatory T cells and Kupffer cells are crucial mediators of systemic T cell tolerance to antigens targeting murine liver. Hepatology50:612– 621. 33. Cao O, Dobrzynski E, Wang L, Nayak S, Mingle B, Terhorst C, Herzog
RW.2007. Induction and role of regulatory CD4⫹CD25⫹T cells in tolerance to the transgene product following hepatic in vivo gene transfer. Blood110:1132–1140.
34. Martino AT, Nayak S, Hoffman BE, Cooper M, Liao G, Markusic DM, Byrne BJ, Terhorst C, Herzog RW.2009. Tolerance induction to cyto-plasmic beta-galactosidase by hepatic AAV gene transfer: implications for antigen presentation and immunotoxicity. PLoS One 4:e6376. doi:10 .1371/journal.pone.0006376.
35. Franzese O, Kennedy PT, Gehring AJ, Gotto J, Williams R, Maini MK, Bertoletti A.2005. Modulation of the CD8⫹-T-cell response by CD4⫹ CD25⫹regulatory T cells in patients with hepatitis B virus infection. J. Virol.79:3322–3328.
36. Stoop JN, van der Molen RG, Baan CC, van der Laan LJ, Kuipers EJ, Kusters JG, Janssen HL.2005. Regulatory T cells contribute to the im-paired immune response in patients with chronic hepatitis B virus infec-tion. Hepatology41:771–778.
37. Xu D, Fu J, Jin L, Zhang H, Zhou C, Zou Z, Zhao JM, Zhang B, Shi M, Ding X, Tang Z, Fu YX, Wang FS.2006. Circulating and liver resident CD4⫹CD25⫹regulatory T cells actively influence the antiviral immune response and disease progression in patients with hepatitis B. J. Immunol.
177:739 –747.
38. Stoop JN, van der Molen RG, Kuipers EJ, Kusters JG, Janssen HL.2007. Inhibition of viral replication reduces regulatory T cells and enhances the antiviral immune response in chronic hepatitis B. Virology361:141–148. 39. Saraiva M, O’Garra A.2010. The regulation of IL-10 production by
immune cells. Nat. Rev. Immunol.10:170 –181.
40. Lin SW, Hensley SE, Tatsis N, Lasaro MO, Ertl HC.2007. Recombinant adeno-associated virus vectors induce functionally impaired transgene product-specific CD8⫹T cells in mice. J. Clin. Invest.117:3958 –3970. 41. West EE, Youngblood B, Tan WG, Jin HT, Araki K, Alexe G, Konieczny
BT, Calpe S, Freeman GJ, Terhorst C, Haining WN, Ahmed R.2011. Tight regulation of memory CD8(⫹) T cells limits their effectiveness dur-ing sustained high viral load. Immunity35:285–298.
42. Jin HT, Anderson AC, Tan WG, West EE, Ha SJ, Araki K, Freeman GJ, Kuchroo VK, Ahmed R.2010. Cooperation of Tim-3 and PD-1 in CD8 T-cell exhaustion during chronic viral infection. Proc. Natl. Acad. Sci. U. S. A.107:14733–14738.
43. Guidotti LG, Ishikawa T, Hobbs MV, Matzke B, Schreiber R, Chisari FV.1996. Intracellular inactivation of the hepatitis B virus by cytotoxic T lymphocytes. Immunity4:25–36.
44. Michel ML, Deng Q, Mancini-Bourgine M.2011. Therapeutic vaccines and immune-based therapies for the treatment of chronic hepatitis B: perspectives and challenges. J. Hepatol.54:1286 –1296.
AAV-Mediated HBV Replication in HLA-Transgenic Mice