0022-538X/06/$08.00⫹0 doi:10.1128/JVI.01421-05
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Optimal Long-Term Humoral Responses to Replication-Defective
Herpes Simplex Virus Require CD21/CD35 Complement
Receptor Expression on Stromal Cells
Mark A. Brockman,
1,3† Admar Verschoor,
2,4‡ Jia Zhu,
1§ Michael C. Carroll,
2,4and David M. Knipe
1,3*
Departments of Microbiology and Molecular Genetics1and Pediatrics and Pathology,2Program in Virology,3Harvard Medical School,
and CBR Institute for Biomedical Research,4200 Longwood Avenue, Boston, Massachusetts 02115
Received 8 July 2005/Accepted 27 April 2006
Replication-defective herpes simplex virus (HSV) strains elicit durable immune responses and protect against virulent HSV challenge in mice, despite being unable to establish latent infection in neuronal cells. Mechanisms for generating long-lived immunity in the absence of viral persistence remain uncertain. In animals immunized with replication-defective HSV, durable serum immunoglobulin G (IgG) responses were elicited. Surprisingly, Western blot analyses revealed that the specificities of antiviral IgG changed over time, and antibody reactivity to some viral proteins was detected only very late. Thus, some of the durable IgG activity appeared to be contributed by either new or significantly enhanced antibody responses at late times. Following immunization, radiation bone marrow-chimeric mice lacking complement receptors CD21 and CD35 on stromal cells elicited only short-lived serum IgG and failed to mount recall responses to subsequent HSV exposure. Our results suggest that complement-mediated retention of viral antigens by stromal cells, such as follicular dendritic cells, is critical for optimal maintenance of antibody responses and B-cell memory following vaccination with replication-defective HSV.
Replication-defective mutant strains of herpes simplex virus (HSV) induce immune responses that are as durable as those of replication-competent HSV strains (7, 30). This continues to be paradoxical because several studies have shown that repli-cation-defective HSV strains do not spread efficiently to sen-sory ganglia and establish latent infection to only a very low level (12, 16, 17, 21). In addition, replication-defective HSV mutants express many viral proteins (9, 10) but do not generate infectious progeny virus. Regardless, they are immunogenic in animal models (6, 9, 12, 29), including rhesus macaques (32), and elicit protection against virulent HSV challenge (5, 6, 10, 25, 26, 28, 29). A recent comparison of herpes vaccine candi-dates in mice indicated that replication-defective HSV strains were more effective than glycoprotein- or DNA-based modal-ities (16). A complete understanding of the mechanisms by which long-term immune responses are induced by these rep-lication-defective vectors, and viral vaccines in general, is nec-essary and remains an important area of investigation.
Antibody responses following many viral infections are very long lived, and several mechanisms for maintenance of serum immunoglobulin G (IgG) have been described. Lymphocytic choriomeningitis virus infection generates professional anti-body-producing plasma cells (PCs) that traffic to the bone
marrow and whose survival appears to be independent of an-tigen (23). Anan-tigen independence was also observed using an experimental Cre-loxsystem to alter the specificities of mem-ory B cells in vivo (24). Thus, B-cell memmem-ory has been consid-ered by some to be antigen independent. In vesicular stomatitis virus-infected mice, long-lived germinal centers and IgG re-sponses have been associated with antigen retention on follic-ular dendritic cells (FDC) (2, 3), likely in the form of immune complexes of antigen that is opsonized with antibody and com-plement. Therefore, retention of immune complexes on FDC can be mediated by complement receptor (Cr) CD21 or CD35 (1, 13, 36) or Fc gamma receptor II (33, 39) and presumably allows continued restimulation of memory B cells. Thus, some have argued that antigen is necessary for optimal IgG re-sponses.
In mice, complement receptors CD21 and CD35 are splice variants encoded by a single gene,Cr2, and transgenic mice disrupted for this locus (Cr2⫺/⫺) have been generated (1, 13,
27). The role of CD21/CD35 in antigen retention and antibody durability has been investigated previously through the use of radiation chimera approaches in which bone marrow cells were transplanted into irradiated wild-type (WT) or Cr2⫺/⫺animals
(13) or through adoptive transfer studies where antigen-primed cells were injected into naı¨ve WT or Cr2⫺/⫺mice (4). In these
systems, B cells expressed CD21/CD35, whereas stromal cells were deficient for these receptors, or vice versa. Using either methodology, signaling of the CD21/CD35 receptor on B cells was required to generate a short-term IgG response to purified protein antigen delivered systemically, but long-term mainte-nance of serum IgG required expression of Cr2 on stromal cells, such as FDC.
To investigate the mechanism by which immunization with
* Corresponding author. Mailing address: Department of Microbiol-ogy and Molecular Genetics, Harvard Medical School, 200 Longwood Avenue, Boston, MA 02115. Phone: (617) 432-1934. Fax: (617) 432-0223. E-mail: [email protected].
† Present address: Massachusetts General Hospital, Boston, MA 02129.
‡ Present address: Institute of Experimental Immunology, Zurich, Switzerland.
§ Present address: Fred Hutchinson Cancer Research Center, Seattle, WA 98109.
7111
on November 8, 2019 by guest
http://jvi.asm.org/
replication-defective HSV induces durable immunity, we eval-uated the antiviral IgG response in mice. We observed that the HSV protein specificities of the IgG response shifted over time, suggesting that the humoral response continued to evolve several months postinfection. To assess the role of antigen retention in this setting, we used a radiation-chimeric mouse model. In mice where CD21/CD35 was absent on stromal cells, antiviral serum IgG titers declined rapidly and recall responses to HSV were lost. We conclude that optimal maintenance of IgG responses following replication-defective HSV infection requires expression of CD21/CD35 on stromal cells.
MATERIALS AND METHODS
Cells and viruses.Vero cells (CCL-81; American Type Culture Collection)
containing the HSV-2UL5 and HSV-1UL29genes (V5-29 cells) have been
described previously (10). Cells were grown in Dulbecco’s modified Eagle’s
medium (DMEM; Cellgro) supplemented with 100 units/ml penicillin, 100g/ml
streptomycin, 2 mML-glutamine, 400g/ml G418, and 10% bovine calf serum
(vol/vol) (DMEM-10) at 37°C with 5% CO2. Following viral infection, cells were
maintained in DMEM containing penicillin, streptomycin,L-glutamine, and 1%
bovine calf serum (vol/vol) (DMEM-V) at 34°C until harvest.
Replication-defective HSV-2 strainsdl5,dl29 (9, 12), and 5BlacZ(10) have
been described previously. Thedl5 mutant was generated from the HSV-2 186
WT strain by deletion of theUL5gene. UL5 is an essential component of the viral
helicase-primase complex and is required for viral DNA synthesis. Thedl29
mutant was generated in the 186 strain by deletion of theUL29gene, which
encodes the viral single-stranded DNA binding protein ICP8, which is also
essential for viral DNA synthesis. The 5BlacZmutant was generated from the
186 strain by replacing theUL29gene (ICP8 protein) with a fusion protein that
contains the N-terminal 288 amino acids of ICP8 fused to theEscherichia coli
-galactosidase protein. All viruses were propagated on a complementing
Vero-derived cell line, V5-29, which expresses theUL5andUL29gene products. Stocks
of cell-free virus were prepared and their titers determined using V5-29 cells as described previously (19), and they were verified to be replication defective by titration on noncomplementing Vero cells. The attenuated,
replication-compe-tent HSV-2 186⌬Kpn strain contains a deletion in the viral thymidine kinase
(TK) gene (17) and was used in some experiments to compare the persistence levels of viral DNA following immunization. This virus was grown and its titer was determined using Vero cells.
Mice and bone marrow chimera construction.BALB/cJ mice were purchased from Jackson Laboratories (Bar Harbor, ME) and used for the experiments shown in Fig. 2 and 3. C57BL/6 mice (CD45.1 allotype) served as WT controls and bone marrow donors for the generation of radiation chimeras (see Fig. 4).
Mice deficient in complement receptors CD21 and CD35 (Cr2⫺/⫺), maintained
on a C57BL/6 background (CD45.2 allotype), were used as controls and bone
marrow recipients for radiation chimeras. C57BL/6 and Cr2⫺/⫺mice were bred
in-house at Harvard Medical School. Radiation chimeras were generated as
de-scribed previously (38). Briefly, 6-week-old mice were lethally irradiated (2⫻650
rads from a cesium-137 source), anesthetized by isoflurane inhalation (Abbott
Laboratories), infused with 1⫻107WT bone marrow cells, and rested for 6
weeks prior to the experiments. Bone marrow chimeras were evaluated by mea-suring the percentage of CD45.1-positive peripheral blood mononuclear cells using flow cytometry. Successful chimeras were defined as animals with at least 95% donor bone marrow-derived peripheral blood mononuclear cells. All mice were housed at Harvard Medical School in a specific-pathogen-free facility. Study procedures complied with NIH guidelines and institutional reviews by the CBR Institute and Harvard Medical School Animal Care and Use committees.
Virus inoculations.Mice were inoculated by either the subcutaneous (s.c.), the
intramuscular (i.m.), or the intraperitoneal (i.p.) route using a dose of 2⫻106
PFU of virus in a 20-l volume prepared by dilution in sterile, endotoxin-free
saline (Sigma). For initial experiments, groups of BALB/cJ mice were inoculated
s.c. at week 0 with eitherdl5 ordl29. At weeks 4 and 8, all mice were inoculated
s.c. with 5BlacZ. This immunization strategy reproducibly elicits protection
against challenge infection and induces robust and durable IgG responses (7, 29).
For radiation chimera experiments, C57BL/6 (WT), Cr2⫺/⫺mice reconstituted
with WT bone marrow (WTBM3Cr2⫺/⫺) or untreated Cr2⫺/⫺control mice
were infected by the i.m. route using a dose of 2⫻106
PFU of 5BlacZat weeks
0, 4, and 8. Recall responses in these mice were measured following i.p.
inocu-lation with 5BlacZat week 32 (6 months following the final immunization). The
i.p. route was chosen to elicit maximal stimulation of memory B-cell responses.
Serum collection and ELISA.Blood was collected from each mouse prior to inoculation (week 0) and throughout the course of each experiment by tail vein puncture or retro-orbital bleeding. Sera were prepared using Microtainer brand
serum separators (Becton Dickinson) and stored at⫺20°C prior to analysis.
Enzyme-linked immunosorbent assays (ELISA) to determine antigen-specific IgG titers were completed as described previously (11). Briefly, 96-well Maxisorp microtiter plates (Nunc) were coated with Triton X-100-treated, cell-free HSV-2
5BlacZlysate (250 ng total protein/well) in sodium bicarbonate buffer, pH 9.6
(Sigma). Plates were incubated with serial twofold dilutions of sera, and IgG was detected using rabbit anti-mouse IgG antibody conjugated to alkaline
phos-phatase (1:1,000 dilution; Sigma). Plates were developed withp-nitrophenyl
phosphate (Sigma) and read at 405 nm on a microplate reader (Molecular Devices). Antibody titer was calculated as the reciprocal of the final dilution resulting in an optical density at 405 nm 0.2 units above the background (naı¨ve
serum) level and is presented as the mean⫾standard error of the mean (SEM)
for each group.
Western immunoblots.Triton X-100-treated, cell-free 5BlacZvirions (10g
total protein) or Triton X-100-treated, 5BlacZ-infected V5-29 cells (50g total
protein) were fractionated using preparatory gels containing 9.25%N,N⬘
-dial-lyltartardiamide-cross-linked sodium dodecyl sulfate-polyacrylamide gel electro-phoresis gel and electroblotted to nitrocellulose (PROTRAN, Schleicher and Schuell). Membranes were washed in Tris-buffered saline (pH 7.4) containing 0.1% Tween 20 (TBST) and treated with blocking buffer (TBST containing 5% nonfat milk) for 2 h at 4°C. Serum samples from each group of mice were pooled in equal parts, diluted 1:200 in blocking buffer, and assayed using a Surf-Blot apparatus (Idea Scientific, MN) according to the manufacturer’s directions. After incubation at room temperature for 1 h, the membrane was washed in TBST, incubated with secondary rabbit anti-mouse IgG antibody coupled to horseradish peroxidase (1:10,000; Santa Cruz), washed, and detected using en-hanced chemiluminescence reagents (Amersham) and Kodak XAR-5 film.
Real-time PCR.HSV-2 DNA was detected in mouse tissues using a sensitive real-time PCR assay based on techniques described previously (17). Mice were
inoculated by the i.m. route with a dose of 2⫻106PFU of 5BlacZor 186⌬Kpn
or mock treated. At 5 months postinfection, the indicated tissues were collected
under sterile conditions and frozen at⫺80°C prior to processing as described
previously (17, 20). Total genomic DNA was extracted using a DNeasy tissue kit (QIAGEN). HSV DNA was detected using real-time PCR. In brief, TaqMan PCR amplification and detection were performed on an ABI 7700 sequence detector (Applied Biosystems) using the HSV-2 TK gene-specific primers F145 (CTG TTC TTT TAT TGC CGT CAT CG) and R263 (GTC CAT CGC CGA GTA CGC) and the fluorescence-labeled probe TK-pr (6-carboxyfluorescein-TTT GAA CTA AAC TCC CCC CAC CTC
GC-6-carboxytetramethylrhodam-ine). Reactions were performed in a 50-l volume containing 30 ng of DNA and
TaqMan universal PCR master mix (Applied Biosystems) with a final concen-tration of 250 nM of each primer and 200 nM of TaqMan probe and amplified for 40 cycles. For the viral DNA standard, purified HSV-2 186 strain DNA was
serially 10-fold diluted (from 106to 1 copy) in the presence of 30 ng genomic
DNA isolated from uninfected CV-1 cells. This standard curve indicated that the limit of detection for the assay was one copy.
RESULTS
Replication-defective HSV-2 recombinants elicit durable
humoral responses. We have shown previously that
replica-tion-defective HSV-1 mutant viruses elicit durable immunity in mouse models (7, 31). Because of the potential utility of mu-tant HSV-2 strains as genital herpes vaccines (9, 12), we wanted to verify these observations using HSV-2 mutant strains. We examined the serum IgG response following im-munization with replication-defective HSV-2 mutants derived from WT 186 strain virus that are deleted for either theUL5
helicase-primase component protein gene (dl5 strain) or the
UL29 single-stranded DNA binding protein gene (dl29 and
5BlacZstrains) (Fig. 1). Both of these genes are essential for productive HSV infection (10). Mice were inoculated at week 0 by the s.c. route with a dose of 2⫻106PFU ofdl5 ordl29.
At weeks 4 and 8, all mice were inoculated with a dose of 2⫻ 106 PFU of 5BlacZ. We collected serum from each animal
on November 8, 2019 by guest
http://jvi.asm.org/
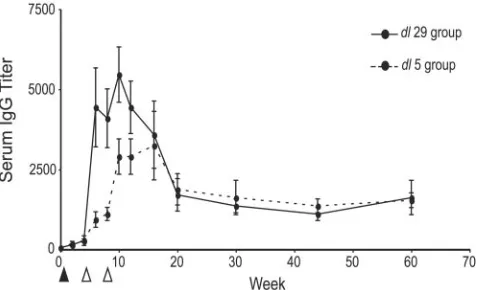
every 2 weeks up to week 12 and measured antiviral IgG by ELISA (Fig. 2). Primary inoculation with either dl5 or dl29 virus generated a low-titer, anti-HSV IgG response by week 4. Secondary infection with 5BlacZenhanced responses at weeks 6 and 8, and an additional boost with 5BlacZfurther increased antiviral IgG titers by weeks 10 and 12. Unexpectedly, serum titers in the dl5 group were consistently lower than in the
dl29-inoculated animals. This may indicate a difference in the ability of these viruses to produce viral antigen and induce primary IgG responses that were apparent only following a secondary exposure using 5BlacZ.
To assess the durability of these responses, we evaluated serum titers in these mice at weeks 16, 20, 30, 44, and 60. We observed that antiviral IgG titers peaked at relatively early times and then declined in all groups but stabilized by week 20 at approximately 30 to 50% of peak titers. Antibody titers were constant for at least 50 weeks postinfection, and these results paralleled our earlier findings of durable IgG antibody re-sponses following inoculation of mice using replication-defec-tive HSV-1 strains (7).
Some IgG responses are enhanced at late times
postinfec-tion.Productive HSV infection is characterized by the
expres-sion of at least 70 viral proteins. To assess the breadth of the IgG responses in these mice, we evaluated the HSV-2 antigen specificity using Western blot analysis. Pooled sera from the
dl29-infected group of mice were used to probe membranes prepared using cell-free HSV-2 virion protein lysate (Fig. 3A). Despite the fact that ELISA titers remained relatively stable at late times, we observed that the antiviral IgG specificities were not static over this period. Three patterns of antibody re-sponses, which we have labeled “continuous,” “declining,” and “late/enhanced,” were seen in these mice. Continuous IgG responses were usually generated immediately following infec-tion and remained detectable for the durainfec-tion of the experi-ment. This type of response could be seen for proteins with molecular masses of⬃200 kDa and ⬃35 kDa. Declining re-sponses were also induced early following infection but ap-peared to diminish over time (e.g., for proteins with molecular masses of⬃180 kDa,⬃120 kDa, ⬃110 kDa, and⬃50 kDa). Late/enhanced IgG responses appeared to be significantly im-proved at later times in the absence of new infection (e.g., for proteins with molecular masses of ⬃70 kDa, ⬃60 kDa, and
⬃40 kDa).
A significant component of the host immune response to
HSV is likely to be directed against nonstructural viral proteins that are highly expressed in infected cells and not incorporated efficiently into viral particles. Therefore, we repeated this Western blot analysis using membranes prepared with lysates from HSV-infected V5-29 cells (Fig. 3B). We observed similar patterns of responses regardless of the source of viral proteins. Continuous IgG responses were directed against proteins with molecular masses of⬃200 kDa,⬃110 kDa,⬃80 kDa,⬃65 kDa (Fig. 3B, broad band),⬃60 kDa, and⬃35 kDa. In addition, a late/enhanced IgG response against one protein with a molec-ular mass of ⬃55 kDa was observed. Next, we performed similar Western blot analyses using sera from thedl5 group of mice (Fig. 3C). In addition to several continuous responses in these animals (i.e., for proteins with molecular masses of⬃110 kDa and 65 kDa), we observed late/enhanced responses to proteins with molecular masses of ⬃60 kDa and ⬃55 kDa. Taken together, these results indicated that the IgG responses evolved over an extended time following immunization with replication-defective HSV mutants. This might be due to mat-uration of IgG avidity, ongoing isotype class-switch recombi-nation, or delayed expansion of low-magnitude cell popula-tions. Regardless of the mechanism, this observation indicates that sufficient viral antigen may persist following inoculation to drive IgG evolution. We observed similar late/enhanced re-sponses to a subset of viral proteins in both groups of mice, further implying that this result may be a general occurrence following infection with replication-defective HSV vectors.
Maintenance of serum IgG requires CD21/CD35 on stromal
cells.The apparent ongoing evolution of the humoral immune
response indicated that antigen persistence might play an im-portant role in the generation of durable immunity following infection by replication-defective HSV. To address this di-rectly, we used a radiation bone marrow chimera model in which Cr2⫹/⫹myeloid cells were transferred into Cr2⫺/⫺mice.
[image:3.585.303.543.70.215.2]We generated chimeric mice by reconstituting irradiated Cr2⫺/⫺mice with wild-type bone marrow (WTBM3Cr2⫺/⫺).
FIG. 1. Genomic maps of replication-defective HSV-2 strains used in this study. All viruses were derived from the wild-type HSV-2 186 strain. Thedl5 ordl29 replication-defective mutant virus contains a deletion of the entireUL5or UL29 gene, respectively. The 5BlacZ
mutant virus expresses a nonfunctional ICP8 protein (UL29 gene)
[image:3.585.55.272.71.140.2]fused in frame to theE. coli-galactosidase (gal) protein. Viruses were propagated on a complementing Vero-derived cell line, V5-29, which expresses theUL5andUL29products upon HSV infection.
FIG. 2. Durable IgG responses are elicited following infection with replication-defective HSV-2 strains. The induction and durability of HSV-2 specific IgG were measured using ELISA. Groups of mice were inoculated by the s.c. route at week 0 (black triangle) with 2⫻106PFU of either straindl29 (n⫽6) (solid line) or straindl5 (n⫽6) (dashed line). At weeks 4 and 8, all mice were boosted by inoculation with 2⫻ 106PFU of strain 5BlacZ(white triangles). Sera were collected at the indicated times, and IgG antibody titers were determined. Results are shown as the reciprocals of the mean endpoint dilutions⫾SEMs.
on November 8, 2019 by guest
http://jvi.asm.org/
The percentages of chimerism were examined by flow cytom-etry to distinguish donor cells (CD45.1 positive) from recipient cells (CD45.2 positive) and were greater than 98% in all mice (data not shown). Mice were inoculated at weeks 0, 4, and 8 with a dose of 2⫻106PFU of 5BlacZvirus by the i.m. route.
Sera were collected regularly, up to week 32, and antiviral IgG titers were determined using ELISA (Fig. 4A). Following in-fection, WT mice elicited IgG responses at weeks 3 through 10 that were similar in kinetics and magnitude to those in previous experiments (Fig. 2). Cr2⫺/⫺mice displayed impaired humoral
responses to HSV, as observed previously (11). This result was consistent with the loss of essential costimulatory signals pro-vided by CD21/CD35 on B cells (1, 13). WTBM3Cr2⫺/⫺
chi-meras induced robust IgG responses that were comparable to those in WT controls through at least 10 weeks. Between weeks 10 and 13, we observed a decline in serum IgG titers from peak values in all groups. The rates of IgG loss, however, were more rapid in the WTBM3Cr2⫺/⫺chimeric animals. By week 22,
only WT mice retained significant antiviral serum IgG (approx-imately 35% of peak titers). These results indicated that CD21/ CD35 on radiation-resistant cells, likely stromal cells, played an important role in the maintenance of IgG responses at later times following replication-defective HSV infection.
Stromal cell CD21/CD35 is critical for IgG recall responses.
[image:4.585.67.518.78.266.2]To assess memory B-cell activity at late times postinfection, we boosted the mice by immunization with 5BlacZvirus and
FIG. 3. Specificities of serum IgG responses change at late times following inoculation. Sera were collected as indicated in Fig. 2, and equivalent volumes from each mouse were pooled for use in Western blot assays to determine the breadth of IgG reactivities to HSV-2 proteins. Samples from thedl29 group (A and B) or thedl5 group (C) were used to probe nitrocellulose membranes prepared using cell-free HSV-2 virion protein lysate (A) or HSV-2 infected V5-29 cell lysate (B and C) as described in Materials and Methods. The times of serum collection (in weeks) are indicated at the top. Molecular masses (in kDa) of marker proteins for each blot are shown on the left. Continuous (C), declining (D), and late/enhanced (E) responses, as described in Results, are indicated to the right of each blot.
FIG. 4. Serum IgG responses to replication-defective HSV infection require stromal cell CD21/CD35. (A) WT mice (n⫽7) (black circles), WTBM3Cr2⫺/⫺chimeras (n⫽5) (white squares and dashed line), and Cr2⫺/⫺mice (n⫽5) (white triangles) were inoculated with 2⫻106PFU of 5BlacZby the i.m. route at weeks 0, 4, and 8 (black triangles). Sera were collected at the indicated times, and anti-HSV-2 IgG titers were determined by ELISA. Mean reciprocal IgG titers⫾SEMs are shown. (B) Recall responses in these mice were evaluated by i.p. inoculation with 2⫻106PFU of 5BlacZat week 32 (white triangle). Serum IgG titers were determined by ELISA, and results are shown as means⫾SEMs.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.102.489.517.676.2]measured their ability to elicit recall responses (Fig. 4B). To ensure a maximal recall response in these animals, we inoc-ulated mice by the i.p. route at week 32 and collected sera at weeks 33 and 34 for ELISA analysis. WT mice primed 6 months earlier by the i.m. route responded to the i.p. boost with robust antiviral IgG titers that were greater than pre-vious peak activity titers. Cr2⫺/⫺mice failed to elicit
signif-icant antiviral IgG following this additional exposure. No-tably, the WTBM3Cr2⫺/⫺chimeras also failed to mount a
significant recall response to this late HSV boost. These results suggested that the population of reactive memory B cells was significantly diminished in the absence of CD21/ CD35 expression on stromal cells.
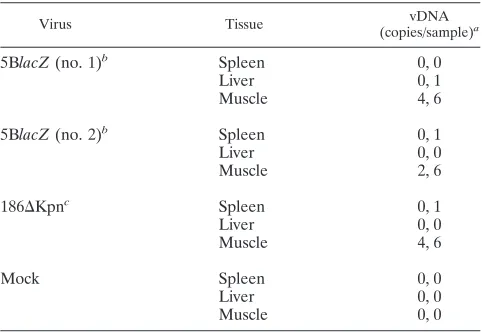
Evaluation of viral persistence in inoculated mice. The
previous results argued that retention of viral antigen on stromal cells, such as FDC, was necessary for long-term antibody responses. Another possible explanation for this observation was persistent viral infection in these animals. Therefore, we examined the levels of HSV DNA present in muscle tissue from two WT mice 5 months following infec-tion with 5BlacZ using a sensitive real-time PCR assay. In duplicate assays, only very low levels of HSV DNA (two to six molecules per sample) were observed in muscle at the site of inoculation with 5BlacZ(Table 1). HSV-2 DNA was not detected reproducibly in distal tissues from these mice (i.e., spleen and liver). Similar results were obtained follow-ing inoculation with the attenuated, replication-competent 186⌬Kpn strain, which contains a deletion of the viral thy-midine kinase gene. All tissues from a mock-treated animal were negative for HSV-2 DNA in these experiments. These data indicated that neither replicating nor nonreplicating strains of HSV-2 established ongoing infection efficiently. Instead, the limited amounts of viral DNA detected are more consistent with a model in which long-term antibody responses are maintained by mechanisms other than viral persistence, such as antigen retention on FDC.
DISCUSSION
We have shown here that immunization with replication-defective HSV-2 strains can elicit durable serum IgG responses and that antigen retention by CD21/CD35 appears to be very important for maintenance of long-term humoral immunity. We observed that the IgG response to replication-defective HSV-2 in WT mice was not static and that qualitative changes in antibody reactivity to viral proteins occurred at late times postinfection. In particular, many viral-antigen-specific IgG re-sponses were generated at early times and were maintained over the course of 1 year postinfection. Other responses were generated early but appeared to diminish over this time. Fi-nally, a subset of responses was enhanced at late times. This indicated that a portion of the durable antibody response to replication-defective HSV infection was due to antibody activ-ity that was either new or significantly enhanced late after infection. Inoculation of WTBM3Cr2⫺/⫺chimeric mice
indi-cated that complement receptors CD21 and/or CD35 ex-pressed on stromal cells play an important role in maintaining serum IgG titers and memory B-cell responses following im-munization. Together, these results indicate that antigen re-tention by CD21/CD35 is a necessary component of the dura-ble antibody response elicited by replication-defective HSV mutants.
Durable serum antibody responses to viral infection can be explained by antigen-independent mechanisms mediated by the generation of long-lived, antibody-producing PCs (34, 35) or by antigen-dependent mechanisms that require continued stimulation of memory B cells by immune complexes trapped on FDC (2, 3) that are present in the spleen or lymph node (14, 18, 22). The continuous responses that we observed might arise by either mechanism. Similarly, the declining responses might be merely a less stable activity resulting from either turnover of short-lived PCs (34) or a loss of memory B cells due to the decay of trapped antigen or diminished B-cell stimulation (8, 15). Therefore, the differences we detected between continu-ous and declining responses may indicate variation in B-cell or PC generation quantity or quality at early times following in-fection. The late/enhanced responses that we observed follow-ing replication-defective HSV-2 infection, however, allow us to speculate on the importance of antigen retention in this set-ting, because generation of new B-cell responses, expansion of existing low-magnitude populations, or ongoing IgG avidity maturation would not be expected to occur in the absence of antigen. Detection of these responses at late times provides compelling evidence that viral antigen remains accessible for an extended time following infection.
To test this hypothesis directly, we used a bone marrow chimera model. The fact that maintenance of serum IgG titers and B-cell recall activity required CD21/CD35 expression on stromal cells provides strong support for a model in which the durable immunity generated by replication-defective HSV in mice is largely antigen dependent. Association of viral proteins with FDC would allow naı¨ve B cells to encounter antigen and/or memory B cells generated in low abundance during initial infection to reengage antigen, thereby enhancing immu-nity without requiring ongoing infection. In the absence of efficient FDC-mediated antigen trapping in WTBM3Cr2⫺/⫺
[image:5.585.43.284.89.255.2]chimeric mice, early serum antibody titers were comparable to
TABLE 1. Real-time PCR quantification of HSV DNA at 5 months postinfection
Virus Tissue vDNA
(copies/sample)a
5BlacZ(no. 1)b Spleen 0, 0
Liver 0, 1
Muscle 4, 6
5BlacZ(no. 2)b Spleen 0, 1
Liver 0, 0
Muscle 2, 6
186⌬Kpnc Spleen 0, 1
Liver 0, 0
Muscle 4, 6
Mock Spleen 0, 0
Liver 0, 0
Muscle 0, 0
aReplicate samples from independent assays.
bReplication-defective strain.
cReplication-competent strain.
on November 8, 2019 by guest
http://jvi.asm.org/
those in WT mice, indicating that induction of humoral re-sponses was not altered significantly. Serum IgG titers, how-ever, declined rapidly, and recall responses were lost in these chimeric animals, indicating specific defects in antibody and memory B-cell maintenance.
Additional experiments will be required to identify the viral proteins recognized by the late/enhanced IgG responses. The predominance of these responses in purified virion lysates, however, indicates that they may be components of the virus particle. Potential candidates include the structural proteins VP19c (53 kDa) and VP22a (40 kDa) or the tegument proteins VP16 (55 kDa) and VP22 (38 kDa). It is unclear why an immune response to these proteins might differ from those of other viral antigens, though it may indicate that virus particles are preferentially captured and/or stabilized on stromal cells following acute infection. Ongoing viral infection might also provide a continued source of antigen over this extended pe-riod. We observed very low levels of viral DNA in muscle 5 months after inoculation with replication-defective HSV-2, but we did not detect viral RNA transcripts for either the ICP4 or the ICP27 immediate early proteins (data not shown) and therefore do not believe that persistent viral infection can explain the durable immunity seen in these studies. Regardless of the source of the viral proteins, our results demonstrate that complement-mediated retention of antigen by receptors CD21 and CD35 on stromal cells plays a significant role in IgG durability.
Although it could be argued that the amount of viral protein generated during a single-round infection by a replication-defective virus is significantly reduced and that humoral re-sponses in this context may be more dependent on comple-ment activity, we have observed a similar dependence on complement for induction of IgG responses following WT HSV infection (11, 37, 38). Those studies suggested that the amount of viral antigen generated during natural HSV infec-tion is limiting or inefficiently presented to B cells without the assistance of these innate factors. Our current results are con-sistent with a model in which opsonization by complement protein C3 and subsequent trapping of immune complexes by CD21/CD35 on FDC are a necessary step in the maintenance of IgG responses to replication-defective HSV, indicating that optimal durable immunity is dependent on in vivo antigen retention.
In conclusion, we have shown that the serum IgG antibody response following infection with replication-defective HSV-2 is durable but not static over time. Analyses of WTBM3Cr2⫺/⫺
chimeric mice indicated that CD21/CD35 expression on radi-ation-resistant stromal cells was necessary to maintain serum IgG titers. Furthermore, these chimeric mice failed to mount a recall response to subsequent HSV-2 infection, indicating a severe defect in B-cell memory. Our data support a model in which antigen retention on FDC is necessary to maintain op-timal serum IgG and memory B-cell responses following rep-lication-defective HSV infection. This may allow ongoing stim-ulation of naı¨ve and memory B cells and replenishment of a short-lived, antibody-producing plasma cell population. Addi-tional experiments will be required to distinguish these possi-bilities. We observed only low levels of HSV-specific antibod-ies in Cr2⫺/⫺mice, which were generated presumably in the
absence of significant antigen trapping by stromal cells and
receptor costimulation provided by Cr2 on B cells. It is appar-ent, however, that optimal long-term IgG responses in this experimental situation require Cr2 expression on stromal cells, which likely functions to allow viral antigen retention by FDC. A more detailed understanding of this CD21/CD35-mediated mechanism may assist in the design and development of vac-cines that induce durable and protective immune responses.
ACKNOWLEDGMENTS
We thank the members of the Knipe and Carroll laboratories for many helpful discussions, the Animal Care and Resources staff at Harvard Medical School for maintaining the animals used in this work, and Lisa Holik for assistance with manuscript preparation.
This work was supported by NIH grants AI042257 (M.C.C.), P01-AI046006 (D.M.K.), and AI57552 (D.M.K.).
REFERENCES
1.Ahearn, J. M., M. B. Fischer, D. Croix, S. Goerg, M. Ma, J. Xia, X. Zhou, R. G. Howard, T. L. Rothstein, and M. C. Carroll.1996. Disruption of the Cr2 locus results in a reduction in B-1a cells and in an impaired B cell
response to T-dependent antigen. Immunity4:251–262.
2.Bachmann, M. F., T. M. Kundig, H. Hengartner, and R. M. Zinkernagel.
1994. Regulation of IgG antibody titers by the amount persisting of
immune-complexed antigen. Eur. J. Immunol.24:2567–2570.
3.Bachmann, M. F., B. Odermatt, H. Hengartner, and R. M. Zinkernagel.
1996. Induction of long-lived germinal centers associated with persisting
antigen after viral infection. J. Exp. Med.183:2259–2269.
4.Barrington, R. A., O. Pozdnyakova, M. R. Zafari, C. D. Benjamin, and M. C. Carroll.2002. B lymphocyte memory: role of stromal cell complement and
FcgRIIB receptors. J. Exp. Med.196:1189–1199.
5.Boursnell, M. E., C. Entwisle, D. Blakeley, C. Roberts, I. A. Duncan, S. E. Chisholm, G. M. Martin, R. Jennings, D. Ni Challanain, I. Sobek, S. C. Inglis, and C. S. McLean.1997. A genetically inactivated herpes simplex virus type 2 (HSV-2) vaccine provides effective protection against primary
and recurrent disease. J. Infect. Dis.175:16–25.
6.Brehm, M. A., R. H. Bonneau, D. M. Knipe, and S. S. Tevethia.1997. Immunization with a replication-deficient mutant of herpes simplex virus
type 1 (HSV-1) induces a CD8⫹cytotoxic T-lymphocyte response and
con-fers a level of protection comparable to that of wild-type HSV-1. J. Virol.
71:3534–3544.
7.Brockman, M. A., and D. M. Knipe.2002. Herpes simplex virus vectors elicit durable immune responses in the presence of preexisting host immunity.
J. Virol.76:3678–3687.
8.Chen, Z., S. B. Koralov, M. Gendelman, M. C. Carroll, and G. Kelsoe.2000.
Humoral immune responses in Cr2⫺/⫺mice: enhanced affinity maturation
but impaired antibody persistence. J. Immunol.164:4522–4532.
9.DaCosta, X., M. F. Kramer, J. Zhu, M. A. Brockman, and D. M. Knipe.2000. Construction, phenotypic analysis, and immunogenicity of a UL5/UL29
dou-ble deletion mutant of herpes simplex virus 2. J. Virol.74:7963–7971.
10.DaCosta, X. J., N. Bourne, L. R. Stanberry, and D. M. Knipe.1997. Con-struction and characterization of a replication-defective herpes simplex virus 2 ICP8 mutant strain and its use in immunization studies in a guinea pig
model of genital disease. Virology232:1–12.
11.DaCosta, X. J., M. A. Brockman, E. Alicot, M. Ma, M. B. Fischer, X. Zhou, D. M. Knipe, and M. C. Carroll.1999. Humoral response to herpes simplex
virus is complement dependent. Proc. Natl. Acad. Sci. USA96:12708–12712.
12.DaCosta, X. J., C. A. Jones, and D. M. Knipe.1999. Immunization against genital herpes with a vaccine virus that has defects in productive and latent
infection. Proc. Natl. Acad. Sci. USA96:6994–6998.
13.Fang, Y., C. Xu, Y. X. Fu, V. M. Holers, and H. Molina.1998. Expression of complement receptors 1 and 2 on follicular dendritic cells is necessary for the
generation of a strong antigen-specific IgG response. J. Immunol.160:5273–
5279.
14.Fayette, J., I. Durand, J. M. Bridon, C. Arpin, B. Dubois, C. Caux, Y. J. Liu, J. Banchereau, and F. Briere.1998. Dendritic cells enhance the
differenti-ation of naive B cells into plasma cells in vitro. Scand. J. Immunol.48:563–
570.
15.Fischer, M. B., M. Ma, N. C. Hsu, and M. C. Carroll.1998. Local synthesis of C3 within the splenic lymphoid compartment can reconstitute the
im-paired immune response in C3-deficient mice. J. Immunol.160:2619–2625.
16.Hoshino, Y., S. K. Dalai, K. Wang, L. Pesniak, T. Y. Lau, D. M. Knipe, J. I. Cohen, and S. E. Straus.2005. Comparative efficacy and immunogenicity of replication-defective, recombinant glycoprotein, and DNA vaccines for
her-pes simplex virus 2 infections in mice and guinea pigs. J. Virol.79:410–418.
17.Jones, C. A., T. J. Taylor, and D. M. Knipe.2000. Biological properties of herpes simplex virus 2 replication-defective mutant strains in a murine nasal
infection model. Virology278:137–150.
on November 8, 2019 by guest
http://jvi.asm.org/
18.Klaus, G. G. B., J. H. Humphrey, A. Kunkl, and D. W. Dongworth.1980. The follicular dendritic cell: its role in antigen presentation in the generation of
immunological memory. Immunol. Rev.53:3–59.
19.Knipe, D. M., and A. E. Spang.1982. Definition of a series of stages in the
association of two herpesviral proteins with the cell nucleus. J. Virol.43:
314–324.
20.Kramer, M. F., and D. M. Coen.1995. Quantification of transcripts from the ICP4 and thymidine kinase genes in mouse ganglia latently infected with
herpes simplex virus. J. Virol.69:1389–1399.
21.Leib, D. A., D. M. Coen, C. L. Bogard, K. A. Hicks, D. R. Yager, D. M. Knipe, K. L. Tyler, and P. A. Schaffer.1989. Immediate-early regulatory gene mutants define different stages in the establishment and reactivation of
herpes simplex virus latency. J. Virol.63:759–768.
22.Ludewig, B., K. J. Maloy, C. Lopez-Macias, B. Odermatt, H. Hengartner, and R. M. Zinkernagel.2000. Induction of optimal anti-viral neutralizing B cell responses by dendritic cells requires transport and release of virus
particles in secondary lypmphoid organs. Eur. J. Immunol.30:185–196.
23.Manz, R. A., M. Lohning, G. Cassese, A. Theil, and A. Radbruch.1998. Survival of long-lived plasma cells is independent of antigen. Int. Immunol.
10:1703–1711.
24.Maruyama, M., K. P. Lam, and K. Rajewsky.2000. Memory B-cell
persis-tence is independent of persisting immunizing antigen. Nature407:636–642.
25.McLean, C. S., M. Erturk, R. Jennings, D. N. Challanain, A. C. Minson, M. E. Boursnell, and S. C. Inglis.1994. Protective vaccination against pri-mary and recurrent disease caused by herpes simplex virus (HSV) type 2
using a genetically disabled HSV-1. J. Infect. Dis.170:1100–1109.
26.McLean, C. S., D. Ni Challanain, I. A. Duncan, M. E. Boursnell, R. Jennings, and S. C. Inglis.1996. Induction of a protective immune response by
mu-cosal vaccination with a DISC HSV-1 vaccine. Vaccine14:987–992.
27.Molina, H., V. M. Holers, B. Li, Y. Fung, S. Mariathasan, J. Goellner, J. Strauss-Schoenberger, R. W. Karr, and D. D. Chaplin. 1996. Markedly impaired humoral immune response in mice deficient in complement
recep-tors 1 and 2. Proc. Natl. Acad. Sci. USA93:3357–3361.
28.Morrison, L. A., X. J. DaCosta, and D. M. Knipe.1998. Influence of mucosal and parenteral immunization with a replication-defective mutant of HSV-2
on immune responses and protection from genital challenge. Virology243:
178–187.
29.Morrison, L. A., and D. M. Knipe.1994. Immunization with replication-defective mutants of herpes simplex virus type 1: sites of immune
interven-tion in pathogenesis of challenge virus infecinterven-tion. J. Virol.68:689–696.
30.Morrison, L. A., and D. M. Knipe.1996. Mechanisms of immunization with
a replication-defective mutant of herpes simplex virus 1. Virology220:402–
413.
31.Morrison, L. A., and D. M. Knipe.1997. Contributions of antibody and T cell subsets to protection elicited by immunization with a replication-defective
mutant of herpes simplex virus type 1. Virology239:315–326.
32.Murphy, C. G., W. T. Lucas, R. E. Means, S. Czajak, C. L. Hale, J. D. Lifson, A. Kaur, R. P. Johnson, D. M. Knipe, and R. C. Desrosiers.2000. Vaccine protection against simian immunodeficiency virus by recombinant strains of
herpes simplex virus. J. Virol.74:7745–7754.
33.Qin, D., J. Wu, K. A. Vora, J. V. Ravetch, A. K. Szakal, T. Manser, and J. G.
Tew.2000. Fc gamma receptor IIB on follicular dendritic cells regulates the
B cell recall response. J. Immunol.164:6268–6275.
34.Slifka, M. K., and R. Ahmed.1998. Long-lived plasma cells: a mechanism for
maintaining persistent antibody production. Curr. Opin. Immunol.10:252–
258.
35.Slifka, M. K., R. Anitia, J. K. Whitmire, and R. Ahmed.1998. Humoral
immunity due to long-lived plasma cells. Immunity8:363–372.
36.van den Berg, T. K., E. A. Dopp, M. R. Daha, G. Kraal, and C. D. Dijkstra.
1992. Selective inhibition of immune complex trapping by follicular dendritic
cells with monoclonal antibodies against rat C3. Eur. J. Immunol.22:957–
962.
37.Verschoor, A., M. A. Brockman, M. Gadjeva, D. M. Knipe, and M. C. Carroll.2003. Myeloid C3 determines induction of humoral responses to
peripheral herpes simplex virus infection. J. Immunol.171:5363–5371.
38.Verschoor, A., M. A. Brockman, D. M. Knipe, and M. C. Carroll.2001. Cutting edge: myeloid complement C3 enhances the humoral response to
peripheral viral infection. J. Immunol.167:2446–2451.
39.Yoshida, K., T. K. van den Berg, and C. D. Dijkstra.1993. Two functionally different follicular dendritic cells in secondary lymphoid follicles of mouse spleen, as revealed by CR1/2 and FcR gamma II-mediated immune-complex
trapping. Immunology80:34–39.