0022-538X/79/06-0668/06$02.00/0
Nucleotide
Sequence Analysis of Viable Deletion Mutants
Lacking
Segments
of the
Simian Virus
40
Genome
Coding
for
Small
t
Antigen
BAYAR THIMMAPPAYA AND THOMAS SHENK*
Department of Microbiology, University of Connecticut HealthCenter,Farmington,Connecticut06032
Receivedforpublication27September 1978
The deletions in nine viable simian virus 40 mutants have been mapped by direct DNA sequence analysis. The mutant DNAs lack small segments ofthe early region of the viral chromosome (between0.535 and 0.600 map unit). The deletionsarealllocated in the region which isremovedfrom thelargeTantigen transcript bysplicing. No onedeletion removes this entire region, but no partof thissegment isconservedin allof themutantsexcept for severalnucleotidesnear thesplice points ofthetranscript. Although the deletions do not alter theregion coding for the largeTpolypeptide,they dodeleteportionsof thesegmentcoding for the C-terminal half of the small tpolypeptide.
Twotranscripts have beenidentified insimian virus40 (SV40)-infectedcellswhich map in the early region of the viral genome (Fig. 1). Both transcripts extend (5' -+ 3') from near 0.66 to
0.16 onthe SV40map,andboth arespliced (1,
2). One transcript is believed to code for the SV40 small t antigen, a polypeptide of about 17,000 daltons. Thispolypeptidehasbeen iden-tified in bothlyticallyinfectedmonkeycellsand
transformed rodent cells (3, 10, 17, 18). The
second transcript codes for large T antigen, a
polypeptideof about 90,000 daltonswhichisalso
present in both lytically infected and
trans-formed cells (10, 13).
The T and t polypeptides are structurally
related. Most of the
[35S]methionine-labeled
trypticpolypeptides ofsmall tare alsopresent in large T antigen (10), and Paucha et al. (9) have shown the N-terminal amino acid se-quences of these polypeptides to be identical. Thisisconsistent with thestructureof the tran-scripts coding forthe Tandtpolypeptides.The
transcripts share the same 5'-nucleotide
se-quence, coding for the N-terminal portions of
thepolypeptides.But,asaresultofsplicing,the
mRNA coding for large T antigen lacks the sequence codingforboththeC-terminalportion ofsmall tantigen and a series oftranslational terminationcodons (0.534to0.600mapunit) (1, 12, 20).
Aseriesof deletion mutants has been isolated
withlesionslocatedbetween 0.535 and0.600 on
theSV40 map(16,18).Thisregionisnotpresent
in thelargeTtranscript,but iscontainedin the
smalltantigentranscript (Fig. 1).Aspredicted,
these deletions donotaffect the size oflargeT
antigen but do alter small t antigen. The
mu-tants areviable,growingat aslightly slowerrate
than wild-type virus (16, 18),butare markedly impaired in their ability to transform rodent
cells(2, 18). This suggests thatatleast portions
of the C-terminus ofsmalltantigenare dispen-sable for lytic growth of SV40, but an intact
small t antigen is required for efficient viral
transformation.
We havedetermined by directDNAsequence analysis the nucleotide sequence whichnine of these deletion mutants lack. All mutants lack segments of theregionbetween 0.535 and0.600 on theSV40 chromosome. Althoughno mutant lacks this entire region,there is noportionofthe sequence which is conserved in all ofthe
mu-tants, except small segments near the splice
points ofthelargeTantigentranscript.
MATERIALS AND METHODS
Cells and viruses. The origin andproceduresfor growth of the CV-1P line ofAfrican green monkey cells have beendescribedby Mertz and Berg (8). The wild-typeSV40(designatedwt830) is aplaque-purified derivative oftheSVS strain (19).Thedeletionmutants
d1883, d1884, d1890,andd1891werederived fromwt830
(16). Mutantd12006waskindlyprovided by William Topp (18). Mutants d11410 andd11440 werederived by expansion ofthedeletion ind1884, using the pro-cedure ofShenk(14).Mutantd11410containsasecond deletion at 0.70 to 0.71 map unit (15). Thedeletion in
d1891wasexpandedtoproduced11441andd11442.
DNA andenzymes. SV40DNAwasextracted(5) from CV-1P cells infected at a multiplicity of<0.05 PFU/cellwhen >90% of the cellsshowedcytopathic effect. Closed circularviral DNA[SV40(I)]was puri-fieddirectlyfrom thesupernatant byaddingCsCl to 668
on November 10, 2019 by guest
http://jvi.asm.org/
FIG. 1. DiagramoftheSV40chromosomerelating
themappositionof viable deletionmutants(0.60to
0.535 mapunit)tothelocationofthetwoknownearly
transcripts. Theportions ofthe transcriptsbelieved
tocodefor earlyproteinsareindicatedbythe blocked
segments (t=smalltantigen; T=largeTantigen).
Regions removed from transcripts by splicing are
represented by dottedlines. ORI = origin ofDNA replication.
1.56 g/cm3 andethidium bromide to 200,ug/ml and centrifuging toequilibrium.The band ofSV40(I) DNA
wascollected,and the ethidium bromide was removed
by passingthe DNAthrough AG50W-X8resin
(Bio-Rad Laboratories) (11). Restriction endonucleases
were either prepared in the laboratory or purchased from New England Biolabs and used according to
published protocols. Bacterial alkaline phosphatase
was purchasedfrom WorthingtonBiochemicals Corp., and T4 polynucleotide kinase was from Boehringer Mannheim Corp.
DNA sequence analysis. Restriction
endonucle-ase-generated fragmentswereendlabeled bythe
pro-cedure of Maniatiset al. (6). Sequence analysis of end-labeled fragments was perfonned as described by MaxamandGilbert(7), using 12%polyacrylamidegels (0.7 mmthick,40 cmlong).
RESULTS
Toformulateasimple strategy for the
nucleo-tide sequenceanalysisof alterations present in the deletion mutants, it was firstnecessary to determine whichrestriction endonuclease
cleav-agesiteswerepresent andwhich were missing
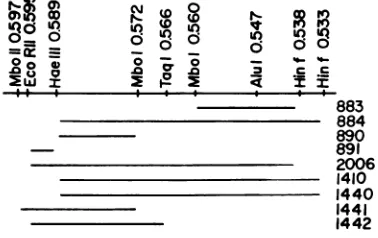
in each mutant DNA. Accordingly, each DNA was cleaved with several enzymes which cut SV40DNAbetween 0.53 and 0.60 map unit. The data from theseanalyses aresummarized in Fig. 2.Mutantd1884DNA, for example, retained the Hinfendonucleasecleavagesite at 0.533 and the
HaeIIIendonucleasecleavage site at 0.595 map
unit, but lost all of the cleavage sites assayed
between these points (Fig. 2). Mutant dl890
DNAretainedall of the cleavage sites assayed, but its HaeIII and MboI endonuclease A frag-ments were bothshortened, indicating that the deletion was contained between theHaeIII and MboI endonuclease cleavage sitesat0.589 and 0.572 mapunit, respectively (Fig. 2).
Utilizingtheinformation inFig. 2, appropriate restriction endonuclease-generated fragments from each mutant were prepared. The fragments
were end labeled (6) and finally cleaved with
anotherrestriction endonuclease togenerate a fragmentlabeledwith 32P at one 5' end. Mutant d1890DNA, forexample,was cleaved with the Hinf and MboIendonucleases, andthe 0.572 to 0.642 map unit fragment was end labeled and then cleaved with HaeIII endonuclease. This produced a fragment with a labeled 5' end at 0.572 map unit and anunlabeled 5' end at 0.589 mapunit for sequenceanalysis. The end-labeled
fragments prepared from the deletion mutant
DNAs are diagramedinFig. 3.This figure also indicates the sequence which was read (solid lines) on either side of the deletions (dashed
lines)aftersubjectingtheend-labeledfragments
tonucleotide sequenceanalysis by themethod
of Maxam andGilbert(7).
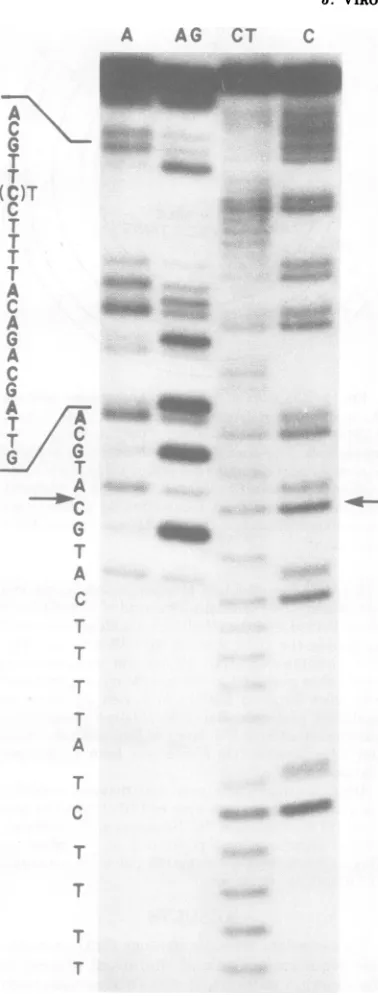
Anexampleof a sequencinggel (dl890DNA)
and thenucleotidesequencedetermined from it areshown inFig.4.The sequence starts from a
pointnearthelabeled, MboI
endonuclease-gen-erated 5' end(complementarytonucleotide 457/ 4704inFig. 5) and isreadfrom bottom to top. The locationofthe deletion isindicatedby an arrowwhere the sequence jumps ahead 27
nu-cleotides as compared with the wild-type
se-quence.
to 'Q o0 U) Uo<
0 I& .00 %t%..
0-0-c
=LLJ =
883 884 890 891 2006 1410 1440 1441 1442 FIG. 2. Location ofviable deletion mutations de-terminedbyrestrictionendonucleasecleavageof
mu-tantDNAs. The mappositionsatwhich the enzymes cleave weredetermined bylocatingthe known
rec-ognition sequencesoftheenzymes in theSV40
nu-cleotidesequence of Reddyetal.(12).Thesolid bars are bounded by restriction endonuclease cleavage
sitespresent inmutant DNAs and extend through
cleavage sites which themutantslack.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.505.263.452.461.580.2]p. o
o q .0 o
cli
I') 0 0 .0X.
"IC)I
co
c c
I I
A.
-_--. 883
--- -_ 884
- -_ 890
- -- -. 890
891
_--
-- - .* 2006- 1410
-- _ .1440
1441
-1442
FIG. 3. Strategyfor nucleotide sequence analysis
of mutantDNAs. The solidcircles represent 32P-la-beledends;the solid lines representregionsin which the sequence wasread;thedashedlines indicate the segmentsdeleted inmutantDNAs.
ThenucleotidesequenceofSV40intheregion
codingfor smalltantigenis
reproduced
inFig.5 (taken from the data of Fiers et al.
[4]
andReddyetal. [12]).The segments which various
deletionmutantslackareindicatedbybrackets.
The points at which Reddy et al.
(12)
havedetermined splicesto occur inthe
large
T and smallttranscriptsarealsoindicated. All of themutantslackportionsof thenucleotide segment
which are not present in the
spliced
large
T transcript.DISCUSSION
Itwas relatively easy to determine the
loca-tion ofthesedeletionsintheSV40genome since
the nucleotide sequence of the viral DNA is
known(4, 12). Ingeneral, thedeletionscouldbe
unambiguouslyfixed in thenucleotidesequence
byanalyzing only onestrand ofthe viral DNA
in the region ofinterest. Volckaert, Fiers, and Berg (personalcommunication) have also deter-mined the nucleotide sequence alterations in
d1883,d1884,d1890,andd1891.Theirresultsare
incomplete agreement with ours.
Thedeletionsrangein sizefrom25to253base
pairs.Theyareall
loeated
inthe region removed from thelarge Tantigentranscript bysplicing(Fig. 5). None removes this entire region (346
nucleotides), but nopart ofthis segment is re-tained in allofthe mutants except forseveral nucleotidesnearthe splicepoints.Although the deletions donotalterthe regioncodingforlarge Tantigen, theydodelete portionsofthe segment
codingfor thesmall tpolypeptide. Thisis
con-sistentwithreports that themutants synthesize normallarge Tbut alteredsmalltpolypeptides
(3,16, 18;G.Khoury,personalcommunication).
The alterations predicted by the nucleotide
sequence agreewellwith thesize of the mutant
i__-_t,.
.g
S-! \
s ZF'
tv .v.
x,
',,61 ..6..
r
:
l.;t.
..^R
.:s
.,
..7F
S,
R
U 1
... .4
.2--,. A
r
G
A
.:. .-I
mmrnm~
FIG. 4. Autoradiograph ofa sequencinggel
con-tainingdl89 DNA. The sequence, which is readfrom
bottomtotop,beginsattheresiduecomplementaryto
position 457/4704 (Fig. 5) and reads through the
deleted segment(indicated byarrows).The Cresidue,
which is six residuesfrom the top ofthe sequence listedbeside thegel,isartifactual. It shouldbea T. WefindanA residue when the opposite strand is
analyzedaspredictedbythedataofFiersetal. (4)
andReddyetal. (12).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.65.257.54.210.2] [image:3.505.264.453.64.562.2]-MOW ATGG T
9561@AT010
GAG
GAATCr
TTGCGC
GCITMG
MTGC TGG GMMT 80/5081 90/5071 100/5061 110/5051 120/5041 130/5031 140/5021 150/5011MET ASPLYS VAL LEUASN ARG GLU GLU SER LEU GLN LEUMETASP LEULEU GLY LEUGLUARG SERALA TRP GLY ASN
ATTCCT C AGKA A TAT
T1bA
AM TGC AA GAG m CATCU[fALM
GGA GCAGALCA
GM M4 AME
/5001 170/4991 180/4981 190/4971 200/4961 210/4951 220/4941 230/4931 240/4921 ILE PRO LEU MET ARG LYS ALA TYR LEU LYS LYS CYS LYS GLU PHE HIS PRO ASP LYS GLY GLY ASP GLU GLU LYS PET LYS LYSPiT
T SPLICE
AT
ATCTG TACMG A ATG G GATGGAG TAT GCT CTCMCC m MA a TTCTGG GATGI
GTAm250/4911 260/4901 270/4891 280/4881 290/4871 300/4861 310/4851 320/4841 330/ ASN THR LEUTYR LYS LYS PETGLU ASP GLYVALLYSTYR ALA HIS GLN PRO ASP PHE GLY GLYPHETRP ASP ALATRGLUVALPHE
r1441
F1442'2006
1441
r1440
r1410
88GCT TCT TCC TAMT CCT GT
GLAT
GCA ATGTAC TGCAMCMCGLG
TGT GCAAG
MA
ATGTCTGCIMC
TGC ATA TGC340/4821 350/4811 360/4801 370/4791 380/4781 390/4771 400/4761 410/4751 ALA SER SER LEU ASN PROGLYVAL ASP ALAMET TYRCYS LYSGLN TRPPROGLU CYS ALA LYS LYS PETSER ALA ASN CYS ILECYS
890i
14421TTGCTGTGC TTA
CI
AG
a&ACA-
AI-
AM TTA TAC MG AM GAT CCA MTGTG TGGGL
T aGTAC TGC TTC GAT /4741 430/4731 440/4721 450/4711 460/4701 470/4691 480/4681 490/4671 500/4661 LEU LEU CYS LEU LEU ARG MET LYS HIS GLU ASN ARG LYS LEU TYRARG LYSASP PRO LEU VAL TRP VAL ASP CYS T CYS PHE ASPr883
TGC
TUlMA
ATG TGG M WACTT M TGT%A GGA AC
TTAMT
CTGTGG
TL
AATT
MA CM AUACC
TACNAGAT
510/4651 520/4641 530/4631 540/4621 550/4611 560/4601 570/4591 580/4581 590/ CYS PHEARGMETTRP PHE GLY LEUASP LEU CYSGLUGLYTHlR LEU LEULEUTRP CYS ASP ILE ILEGLY GLN THRTM
YR ARG ASP2006it SPLICEF-i 883
183
88,44
W01
1T+t
SPLICE [image:4.505.63.458.121.382.2]-TILAPG
CTCJ,GG3T TAIA1TMTGTAL,TGTGTm
ACTACATr
CI&TTGM
GTATA1ATTMACC
TATGGCTG-600/4561 610/4551 620/4541 630/4531 640/4521 650/4511 660/4501 610/4491 680/4481 690/4471 LEULYS LEUFIG. 5. Nucleotidesequence of the portion of the SV40 genome coding for small t antigen. The region
extends fromabout 0.65 to 0.53 map unit. The sequence isfromFiers et al. (4) andReddy et al. (12), and the
nunberscorrespond to those assigned by Fiers etal./Reddyetal.Thesplicepointspresentin the large T and
small ttranscriptsarefrom Reddy et al. (12). The segments which variousdeletionmutants lack are indicated
by brackets.
t proteins which have been observed. Mutant
d1884lacks a DNA segment coding for 63 of the
174 amino acids which the wild-type sequence
predicts comprise the little tpolypeptide (Fig.
5). Three amino acids not found in the
wild-type
proteinshould be added to the C-terminal end
of the dl884polypeptide beforereaching a ter-mination codon (UAG) at position 669/4492
(Fig. 5). This predicts that the d1884 genome
should code for a 114-amino acid little t
poly-peptide.Sleigh et al. (18) and Khoury (personal
communication) haveobserved a 12,000-dalton
polypeptide in dl884-infected cells. Mutant
dl890,whose DNA lacks 27 base pairs, should
synthesize a 165-amino acid small t protein. Khoury (personalcommunication) has found a 16,000-dalton polypeptide in cells infected with
thismutant.Finally,therelativelysmall deletion
(25 base pairs) in d1891should alter the trans-lational reading frame (the deletion is not a
multiple of3 base pairs) toproduce a
dramati-cally shortened polypeptide. Synthesis of this
mutantpolypeptideshould
terminate
at aUGAcodon (Fig. 5, position 432/4729) after adding 19 amino acids in a new reading frame. Thus, the sequencepredictsa109-amino acidpolypeptide.
Khoury (personalcommunication) observes an
11,000-daltonpolypeptide.
Mutant d1884, among others, removes one
splicepointutilizedfor production of thesmall
t antigen transcript (12; Fig. 5, position 604/ 4557). It follows that this mutant cannot synthe-sizeasmall t antigen transcript which isspliced atthe normal position; yetd1884-infected cells
containa12,000-daltonsmalltpolypeptide (18;
Khoury, personalcommunication). Possibly, the
mutant polypeptide is produced utilizing an
unspliced transcript. Alternatively, a suitable
transcript may be produced which is spliced at
a newlocation.Analysis of the early viral
tran-scripts present in the cytoplasm of
d1884-in-fected cells should distinguish between these possibilities.
Several of the deletions described in this
on November 10, 2019 by guest
http://jvi.asm.org/
THIMMAPPAYA AND SHENK
portextend to positions very nearsplice points in both the large T andsmall t transcripts. Using such mutants, we can ask how manynucleotides, ifany, adjacent to a splice point must remain intact if a mature, spliced transcript is to be
generated.The d12006 deletion extends towithin
2 base pairs of thesplice point for the small t transcript at position 604/4557 (Fig. 5). If this mutant can produce a bona fide small t tran-script, then no more than two nucleotides on oneside of thesplicepoint are required by the splicing machinery, unless, of course, the nucleo-tides at the far end of the deletion which are fused to the region of interest can fortuitously
performtherequiredfunction. Ifd12006-infected
cells do not containa smallttranscript with a
normal splicepoint, itshouldbesafe toconclude
thatacritical segment has been deleted.
The d1884 deletion extends through position
659/4502 (Fig. 5) and isonly10basepairs from
the splice point (position 670/4991) utilized in
both the large T and small t transcripts. The mutant makes a normal large Tantigen (3, 16,
18) and,therefore, ispresumed tosynthesize a
properly spliced large T transcript. Thed1884 deletion was expanded by the procedure of
Shenk (14) to determine whether any of the
remaining 10 base pairs between the deletion
and the splice point could be removed while maintaining viability (and, thus, the ability to produce a functional large T transcript). Two expanded mutants were produced, d11410 and
d11440 (Fig. 5). Strikingly, neither deletion
ex-tended closer tothesplicepoint, but thed11410
and d11440 deletions extended 6 and 23 base
pairs in the opposite direction, respectively. Since the deletions did not move further toward
thesplicepoint, it is temptingtospeculate that
all or most of the 10 nucleotides between the ends of the deletions and the splice point are
required to produce a functional (i.e., spliced)
large T transcript. We are presently isolating additional expandeddeletion mutants, both vi-able and defective, to test this notion. Mutant
d11440hasacquiredtwoT/A base pairs at
po-sitions658-659/4502-4503 (Fig. 5) which are not presentinitsparent(d1884).Wecannot offer an explanation for this observation. Nucleotide se-quenceanalysisof additionalexpanded deletion mutants will tell whether this is a common event.
Mutant d1891 lacks a segment extending
through position 352/4809, 26 base pairs from thesecondlarge T transcript splice point (Fig. 5, position 325/4836). This deletion was ex-panded to generate d11441 and d11442. The
d11442deletion has notextendedfurther toward
the splice point, but the
d11441
deletion hasmoved anadditional 8 base pairs in this direction (Fig. 5). Thus, we set themaximum number of base pairsrequired on the deletable side of this
splice point at 18. Further experiments are in
progress to refine thisestimate.
ACKNOWLEDGMENTS
We acknowledge the competent technical assistance of BonnieMassey.
Thisworkwassupportedby Public Health Service research grantCA-19151 fromtheNationalCancer Institute and was
carriedoutduringthe tenureofanEstablished Investigator-ship of the American Heart Association to T.S.
We thank P. Berg, W.Fiers, G. Khoury, and G.Volckaert forcommunicatingtheirunpublisheddata to us.
LITERATURE CMD
1. Berk,A.J., andP. A.Sharp.1978.Spliced earlymRNAs of SV40. Proc. Natl. Acad. Sci.U.S.A.75:1274-1278.
2. Bouck, N., N. Beales, T. Shenk, P. Berg, and G.
diMayorca. 1978. New region of the SV40 genome required for efficient viraltransformation. Proc. Natl. Acad. Sci. U.S.A.75:2473-2477.
3. Crawford, L.V.,C. N.Cole,A. E.Smith,E.Pancha, P.Tegtmeyer,K.Rundell, andP. Berg. 1978.
Or-ganizationandexpressionofearlygenes ofSV40. Proc.
Natl.Acad. Sci. U.S.A.75:117-121.
4. Fiers,W.,R.Contreras, G. Haegeman,R.Rogiers,A.
VandeVoorde, H. VanHeuverswyn,J. Van
Her-reweghe, G. Volckaert, and M. Ysebaert. 1978.
Completenucleotide sequence of SV40 DNA. Nature
(London)273:113-120.
5. Hirt,B. 1967.SelectiveextractionofpolyomaDNA from infected mousecellcultures.J. Mol. Biol. 26:365-369. 6. Maniatis, T., A. Jeffrey,and D.Kleid.1975.Nucleotide sequenceof therightward operator ofphage A.Proc. Natl. Acad.Sci.U.S.A. 72:1184-1188.
7. Maxam,A., andW.Gilbert. 1977. Anewmethod for
sequencingDNA. Proc. Natl. Acad. Sci. U.S.A. 74:560-564.
8. Mertz,J.E., andP.Berg.1974.DefectiveSV40genomes: isolation andgrowthofindividualclones.Virology62: 112-124.
9. Paucha, E.,A.Mellor,R.Harvey,A. E.Smith,R. M.
Hewick, andM. D.Waterfield.1978.Largeandsmall
tumorantigensfromSV40 have identical aminotermini
mapping at 0.65 map units. Proc. Natl.Acad.Sci. U.S.A. 75:2165-2169.
10. Prives, C., E. Gilboa,M.Revel, andE. Winocour. 1977. Cell-free translation of SV40 early messenger RNAcoding for viralT-antigen.Proc.Natl. Acad. Sci.
U.S.A.74:457-461.
11. Radloff, R.,W.Bauer, andJ.Vinograd.1967.A dye-buoyantdensity methodfor the detection and isolation of closed circularduplexDNA: the closedcircularDNA in HeLacells. Proc. Natl. Acad. Sci. U.S.A. 57:1514-1521.
12.Reddy,V.B.,B.Thimmappaya,R.Dhar,K. N.
Sub-ramanian,B. S.Zain,J Pan,P. K.Ghosh,M.L.
Celma, andS. M.Weissman. 1978.Thegenome of SV40. Science 200:494-502.
13.Rundell, K.,J.K.Collins,P.Tegtmeyer,H.L.Ozer,
C.-J. Lai,and D. Nathans. 1977. Identification of simian virus40proteinA.J.Virol.21:636-646. 14. Shenk,T.1977. Abiochemical method forincreasingthe
sizeof deletion mutationsinSV40 DNA. J. Mol. Biol. 113:503-515.
15. Shenk,T. 1978. Construction ofa viableSV40 variant
containingtwofunctionaloriginsof DNAreplication. Cell13:791-798.
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 DELETION
16. Shenk, T.E., J. Carbon, andP.Berg.1975. Construc-tion andanalysis of viable deletionmutantsofsimian virus 40.J. Virol. 18:664-671.
17.Simmona, D. T., and M. A. Martin. 1978. Common
methionine-tryptic peptidesnearthe amino terminal
end ofprimate papovavirustumorantigens. Proc. Natl. Acad. Sci. U.S.A. 75:1131-1135.
18.Sleigh, M.J.,W. C.Topp,R.Hanich,and J. F. Sam-brook. 1978. Mutants of SV40withanalteredsmallt
proteinarereduced intheir abilityto transformcells.
Cell 14:79-88.
19. Takemoto, K. K., R.L Kirchstein, and K. Habel. 1966.Mutants of simian virus40differing in plaquesize, oncogenicity, and heatsensitivity. J. Bacteriol. 92:990-994.
20. Thimmappaya, B., and S. M. Weissman. 1977. The early region ofSV40 DNAmayhavemorethanone gene.Cell 11:837-843.
on November 10, 2019 by guest
http://jvi.asm.org/