0022-538X/78/0026-0214$02.00/0
Copyright©1978 AmericanSocietyforMicrobiology PrintedinU.S.A.
Binding
of tRNA
to
Reverse
Transcriptase of RNA
Tumor Viruses
AMOS PANET* ANDHADASSAH BERLINER
DepartmentofVirology,The HebrewUniversity,Hadassah MedicalSchool,Jerusalem, Israel Received forpublication28November1977
The interactionof tRNA with the reversetranscriptase (RNA-dependentDNA
polymerase) of mammalian RNA viruses, such as Moloney murine leukemia virus andsimiansarcomavirus, has been studied. Whereas the purifiedreverse
tran-scriptase of mammalian virusessedimented in glycerol gradients as a globular protein with a molecular weight of 70,000, after interaction with tRNA the enzyme cosedimented with aprotein of 150,000 molecular weight. The twofold increase in molecularweight could be a result of either two reversetranscriptase molecules
complexed withatRNA or, alternatively, several tRNA molecules bound to a
single enzymepolypeptide.The enzymecomplexes were dissociated in part upon degradation of the tRNA moiety by pancreatic RNase A. The reverse transcrip-tasereleased from virions ofMoloney murine leukemia virus, simian sarcoma
virus, andavianmyeloblastosis virus, bynonionic detergent, migrated fasteron
glycerol gradients than purified enzyme preparation. This phenomenon was
probablydue tocomplex formation between part of the virionenzyme andthe
tRNA,whichisendogenousinvirions. Addition ofexogenoustRNA wasneeded,
however, to quantitatively complex all the virion reverse transcriptase ofMoloney
murineleukemia virus and simiansarcomaviruses. The reversetranscriptase of
Moloney murine leukemia virus did not show tRNA species specificity in the
binding reaction when glycerolgradients were used for assay. Thus, several tRNA
species ofEscherichia coli, yeast, chicken,andrat originwereable to complex
with theenzyme. The species specificityin the interaction between tRNA and
avianmyeloblastosisvirusreversetranscriptasewasalsoexamined. We
demon-strated thatunderourexperimental conditions,thisenzyme binds differenttRNA
species of E. coli and yeastaswellastRNA of chickenorigin.
Initiation of DNA synthesis in RNA tumor scriptase, shows both DNA polymerase and
viruses isprimed in vitro bya tRNA, which is RNaseHactivities(12, 22).
partof the70SRNAgenomecomplex (1). Two In this study we showthat the reverse
tran-groups ofvirus use different tRNA species of scriptase of two mammalian viruses, Moloney
celloriginasprimer. Thus, tryptophan tRNAis MuLVandsimiansarcoma (SiSV),forns
com-theprimer in avian myeloblastosis virus (AMV) plexes with tRNA. The binding of tRNA
and avian sarcomavirus (6), and proline tRNA changes the molecular weight of the mammalian
has recently been shown to be the primer in enzyme from 70,000 to 150,000, suggesting a Moloney murine leukemia virus (MuLV) (17). complexstructure oftwo enzyme subunits and Theprimer tRNATrP in AMV hastworecogni- atRNAmolecule.
tion sites: it is boundat a specific site to the
template35S RNAgenomesubunit(21), and it MATERIALS AND METHODS
complexeswithhighaffinitytothereversetran- Virusesandenzymes. Moloney MuLV was grown
scriptase (16). ThebindingoftRNAT"" topun- inSwiss mousecells(clone 1)and purified as described
fied AMVreversetranscriptaserequires the hol- before(3).AMVwas agiftfromJ.W.Beard ofLife
oenzyme,whichconsists oftwopolypeptides (a SciencesInc., St. Petersburg,Fla., and SiSVwas
ob-and
I?).
Theisolatedsmallerasubunit(molecu- tamed from J. Gruber of the National Institutes oflarweight, 65,000), which manifests both DNA Health, Bethesda, Md. Reverse transcriptasewas
pur-polymeraseand RNase H activities (4), cannot ifiedfrom these three viruses by sequential column
formastablecomplex with tRNATrP (5, 7). e- chromatography on DEAE-Sephadex followed by
forme
aian
transcriptamisolated from mammaphosphoceliulose
(23).All
buffersusedthroughouttheverse transcriptase isolated from mammalian purification procedure contained 20 mM potassium RNA tumor viruses is asinglepolypeptidewith phosphate, pH 7.5,ratherthan
Tris-hydrochloride,
pHa molecular weight of 70,000 (19). This poly- 7.9,since we found that phosphate bufferssignifcantly
peptide,like theasubunit of AMVreversetran- increased the stability of the reverse transcriptase
* ~~~~~~~~214
on November 10, 2019 by guest
http://jvi.asm.org/
duringpurification. The purified enzymewasdialyzed between enzyme andtRNA,we first sought
con-extensively against 50 mM Tris-hydrochloride (pH ditions to separate the complex from the
un-7.5), 0.1 M KCI, 0.1% (vol/vol) Triton X-100, 20% bound reverse
transcriptase.
Glycerolgradients (vol/vol)ethyleneglycol, 2%(vol/vol) glycerol,and 10 conting ethylene glycol were found to be mostmM mercaptoethanoL Pancreatic RNase A, electro- t bl
phoretically purified, was purchased fromWorthing- mixe with purified an after short
ton Biochemicals Co.,Freehold,N.J. mixed with
purified
enzyme, and after short Reagents.Chlcken liver tRNA and rat livertRNA incubation in thecold the mixture, togetherwith were isolated as described by Rogg etal. (18). Yeast twointemalprotein markers,waslayeredonto tRNAwas a product of Schwarz/Mann, Orangeburg, the gradient. After centrifugationfor 27h, theN.Y., and Escherichia coli WtRNA was a gift from positionsin the gradient of the reverse
transcrip-Y. Lapidot, The Hebrew University. These tRNA tase and the protein markers bovine serum
al-preparations were further purified onDEAE-celiulose
bunin
andhuman
IgG were determined.Figure columns (Whatman, DE-52). The tRNA was loaded 1shows
that tRNA affects the sedimentationontothecolumns,which werethen washed stepwise coefficientof thereverse
banscriptase.
Whereas
withpotassiumacetate buffer, pH 5.0, containing 0.1
and 0.3MNaCl. The tRNAwas theneluted with 1.0 free enzyme cosedimented with bovine serum M NaCland precipitated with 2 volumes of ethanol albumin (4.7S; molecular weight, 70,000), addi-Purified E. colitRNAVI and yeasttRNAph were tion of 30 pg oftRNA shifted the polymerase products of Boehringer Chemical Co., Mannheim, Ger- activity to the position of the IgG marker (7.2S; many. These purified tRNA preparations were found molecular weight, 156,000). A twofold increase to lack acceptor tRNA for proline, tryptophan, and in sedimentationrate of the enzyme cannotbe methionine.One absorbance unit of tRNA at 260nm accounted for by equimolarbinding of one en-wasassumedtobe 50
pg,
or 1.67 nmoL Poly(A)and zyme molecule (molecularweight,
70,000) with poly(dT)IOweregiftsfrom Y.Lapidot; [3H]dTTP, 48siyme
tNmolecule (molecular weight,7,0)wtCi/mmol, was obtained from the Radiochemical a single tRNA molecule (molecular weight,
Centre, Amersham, Engld; bovineserum albumin, 25,000). Such acomple
is
more likelytoconsistelectrophoretically purified,wasfromCalbiochem,L of twoenzyme polypeptides and one tRNA
mol-Angeles, Calif.; andpurifiedhumanimmunoglobulin eculeor, alternatively, several tRNA molecules G(IgG)was agift from G. Spira, The Hebrew Univer- bound to a single polypeptide. Addition ofan
sity. even larger excess of tRNA does not further
DNApolymerase assay. Reaction mixtures (100 increasethe sedimentation rate ofthe enzyme 1) contained50mMTris-hydrochloride (pH 8.3),10 (data notshown).However,smalleramounts of
mMdithiothreitol,1ugofpoly(A)-oligo(dT),( 10 of tRNA (3
ug)
cause themajority of reversetran-bovineserumalbumin6mM MgC12, tM
~[scriptase
moleculestomigrate
at an intermedi-(350cpm/pmol),andsamples(usually30Al)
from theglycerol gradient. Reaction mixtures were incubated ate position between the two internal protein
for 60min at37°C, andtrichloroacetic acid-insoluble
radioactivity wascollected on glass fiber filters (What- _
man). One unit of activity is the amount of reverse I 6
BSA
transcriptase needed tocatalyze theincorporation of 71pmol of[3H]dTMPpermin under the above condi- . 0 4
tions. ^ 8 -
~~~~~~~~~~~~~~~A
/f.atiNA.,
38jol 0.4 Etions. ' 8
tRNAbindingassay. Mixtures (200
pil)
contained £.4o 0.2 50mMTris-hydrochloride (pH7.5), 5 mMMgC2, 10mM dithiothreitol, and 0.2% (vol/vol) Triton X-100. a U
0.0
After 10min of incubation at 4°C, theinternalprotein 8 B3A,3, 0.4markers bovine serum albumin (0.6 mg) and human I z
IgG (0.6 mg) were added, and the mixtures were lay- X 4 I 0.2 ered onto5-ml glycerolgradients (5 to 25%, vol/vol) in o a 30 50 mMTris-hydrochloride (pH 7.5), 0.1 M KCI, 0.1% z o.oS
(vol/vol)TritonX-100, 10 mMmercaptoethanol, and 8C c a
20% (vol/vol) ethylene glycol. Gradientswerecentri- -0.4 z
fugedfor 27 hat49,000rpmat4°C inanSW 50.1rotor _ -0.2
i-ofaBeckman
ultracentrifuge
andfractionated from 1 2 30the bottom (usually 32 0.16-mi fractions were col- I -'0_
lected). Samples (30 1d) were taken for the DNA FRACTION NUMBER polymerase assay, and 30 p1 was taken for protein
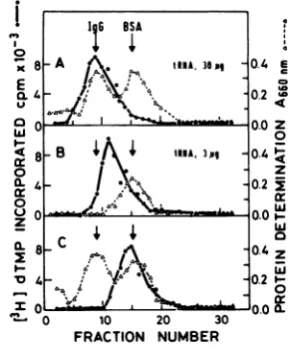
determination (11). Proteins were first precipitated FIG. 1. Binding oftRNA to Moloney MuLV re-with 10%(vol/vol)trichloroacetic acid to remove ma- versetranscriptase. Binding reactions and
centrifu-terials that interfere with theLowry assay(11). gation inaglycerol gradientwereperformedas
de-scribedin thetext. (A) Fortyunits ofreverse
tran-RESULTS scriptaseand 30 pg of rat liver tRNA;(B)40 units of
reversetranscriptase and3pg ofratlivertRNA; (C)
Binding of tRNA to Moloney MuLV re- 40 units of reversetranscriptase.A6w,Absorbance
versetranscriptase.Tostudythe interaction at 660 nm.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.509.285.431.423.598.2]216 BERLINER VIROL.
markers (Fig. 1B).The formation ofan appar- byRNaseA.The factthat RNaseAdissociates
entlysmaller enzyme
complex
may either be the preformedcomplexes (Fig.2B)
suggests that theresult ofpoor separation between the
complex
reverse transcriptase-bound tRNA is notpro-and free enzymeor
perhaps
limitedamountsof tected from RNaseAaction.tRNA induce formation ofa smaller
complex.
Specificity of the binding reaction. WeInclusion of only bovine serum albumin as a havestudied tRNAspeciesspecificityinbinding
marker in the gradient
(Fig. 1B)
demonstrates tothereversetranscriptase ofMoloney MuLVthat the purifiedreverse
transcriptase
doesnot and AMV by using unfractionated or purifiedcontribute enoughproteintointerfere with the tRNA species ofdifferent organisms. Figure 3
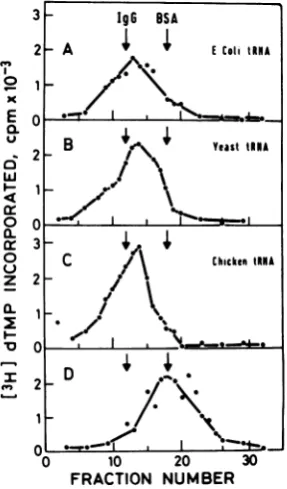
exactpositioningof the markers. shows that Moloney MuLV reverse transcrip-Toprovethat thereversetranscriptasecom- tase can interact with unfractionated chicken
plex indeed contains tRNA, we treated such livertRNA, yeast tRNA,or E. colitRNAwith
complexes with pancreatic RNase A, followed asubsequent increase in the sedimentationrate
bycentrifugation inglycerol gradients. RNaseA of theenzyme. Each of these tRNApreparations
wasmixedwith tRNA either before(Fig.
20)
or contains proline acceptorspecies, but thenu-after (Fig. 2B) binding to the enzyme, and in cleotidesequence of
tRNA'°
of mammalianor-bothexperimentsmostofthereversetranscrip- igin,whichservesas aprimer for DNAsynthesis
tase sedimented as free enzyme. Some of the in Moloney MuLV (17),differs from that of E.
complexes after RNase treatment sedimented coli
tRNAP'ro
(J. Dahlberg, personalcommuni-faster than the free reverse transcriptase (the cation). Purepreparations ofyeast
tRNAPhe
andposition ofbovine serum albumin marker) and E. coli
tRNAvaI,
whichcontain no proline andsimilartocomplexesmade withasmallamount tryptophanacceptor tRNA, also change the
sed-of tRNA (Fig. 1B). These enzyme molecules imentation coefficient ofthe reverse
transcrip-may still contain some intact, or partially de- tase (Fig.4).
graded, tRNAmolecules,since underour
exper-imental conditions 5 to 10% ofthe tRNA was
notconvertedintoacid-soluble
oligonucleotides
3-19G
BSA
2- A 4 ' E Coli flNA
196 BSA w 1
o E
4 *0 20 E0, *
A0E.
u 2 tDNAYeast
MoloneyMuLVreveseransripase40nit
glceo grdin cetiuainwr aredota * _
Cr4C
0I4
decieihet.()Ezyead5 zofra
lieRA. (B
Enyead50u
frtlvrtN RCINNwerinuae for
30mna
°,adte Nss
_0m
oa w c ChickentrNA_ C~~~~~~~~wL
0O 10 20 30 4 4
[image:3.509.293.437.336.581.2]FRACTION NUMBER 2
FIG. 2. Dissociation of reverse transcriptase- 2
tRNAcomplexesbyRNaseA.Bindingreactionswith 1Mo
MoloneyMuLVreverse transcriptase(40units) and
glycerolgradient
centrifutgation
werecarriedoutas C (Adescribed in thetext. (A) Enzymeand 50
pig
ofrat 0 10 20 30 livertRNA.(B) Enzymeand50pgofratlivertRNA FRACTIONNUMBER
wereincubatedfor10minat40C,andthenRNase A
(10pgJwasadded.After30minat40C,dithiothreitol FIG. 3. Reverse transcriptase complex
formnation
was addedto 10mM, andthe mixture was loaded with
unfr-actionated
tRNA of various organisms. ontoagradient.(C)A50-pugamountofratliver tRNA Conditions for binding reactions with Moloneywaspreincubated for15minat370CwithRNaseA MuLVreversetranscriptase (40 units) andglycerol (10
p.gJ
in the binding mixture, and then reverse gradientcentrifugationaredescribedinthetext.(A) transcriptasewasadded.Afteranadditional10min Enzymeand 29pg
ofE. colitRNA; (B)enzymeand at 4°C, the mixture was layered onto agradient. 34pgofyeasttRNA;(C)enzymeand33
pgofchickenA,m ,Absorbanceat660nm. livertRNA; (D) enzyme.
on November 10, 2019 by guest
http://jvi.asm.org/
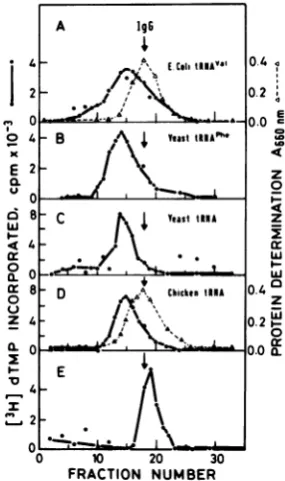
[image:3.509.92.231.363.528.2]lgG
BSAtRNATr
andtRNA4Mt
of origin.60 - Thus, total yeast tRNA, which contains
A I\ Yeast
tINAPhe
tRNATrp with differentprimary
structures(6,
8,
o0 _ /
\10)
orpure E. colitRNAVaJ
andyeasttRNAPhe,
020 I! \ can complex with AMV reverse transcriptase
and
change
itssedimentation
coefficient fromE
7.2S
(IgG
marker) to9S(Fig.
5).30 - \ Structureofreverse
transcriptase
indis-d20 B EColi
tUNA"'
rupted virions of Moloney MuLV, SiSV, andw ! \ AMV. Some 30% of the total RNA in virions of
< 10
Moloney
MuLV consists of small RNA (4-7S)0 150
containing
15species
of hostcell
tRNA (20)._ + Someof the small RNAmoleculesareboundin
o C s d11 the 70S RNA complex, and the rest arereleased
Z-10
_ d free in solution afterdisruption
ofvirions
witha. /.
\nonionic
detergent.
We haveinvestigated
X 5 - 1 \ whether the "free tRNA" is bound to the reverse
- o
.transcriptase,
l\_ | which isalso released from virionsI _
by
detergent
treatment. The results inFig.
630 - D illustrate that the enzyme in disruptedMoloney
20 - MuLV
migrates
inglycerol
gradients
faster thanbovineserumalbumin, whichmarks theposition
10_ / of thepurifiedreversetranscriptase.Addition of
exogenousratliver tRNAfurther increases the
C0 0 1010 ~2020 30 molecular
weight
of virionpolymerase,
with the30 FRACTION NUMBER
FIG. 4. Binding ofreverse transcriptase to purified
tRNAspecies. Conditionsfor bindingreactions with A 19g
MoloneyMuLVreversetranscriptase(60units) and
glycerol gradientcentrifugationaredescribed in the 4 E t tUNA"'Val 0.4 c
text. (A) Enzyme and 20 pgof yeast tRNAPhe; (B) 2r0
enzymeand25 pgofE. colitRNAVal; (C) enzyme and
10pg of oligo(dT);(D)enzyme.
0,
c-o
.0°64-B * YeasttUIA"o
From these results we conclude that under E 2
N
the
binding
conditionsused,
Moloney
MuLVQ.
Oreverse
transcriptase
doesnotselectively
binda, °unique tRNA species. In contrast, synthetic C Y.at toNA Z
oligo(dT),which isanefficientprimerfor DNA 4- /
synthesis with poly(A) as template, does not
o0
* . - waffect the sedimentationrateof theenzyme
(Fig.
8 D i+ tUNA 044). 0
Tofurther
investigate
thequestion
ofspecific-
Z 4- 0.2ity,wereexamined the tRNAbinding specificity a. - o
of AMV reverse transcriptase by the use of 2 .0
1-- E
glycerol gradients. In a previous study (16) it 4
-was shown that outof unfractionated chicken
cell
[32P]tRNA,
only two species, tRNATrP and 2tRNA4Met, were able to bind to AMV enzyme o
withhigh affinity.The
analysis
ofcomplex
for- 0 10 20 30 mation wascarriedoutbyfiltration of the mix- FRACTION NUMBERturesthrough SephadexG-100 columnsthatsep- FIG. 5. Interaction of various tRNA specieswith arated the enzyme-bound [32P]tRNA from un- AMV reversetranscriptase. Conditions for binding
bound[PtRNA
(molecular weight, 25,000). reaction and glycerol gradient centrifugation withbound
[lycerolgadin
etfgtorte AMVreversetranscriptase (50 units)aredescribedinWhen glycerol gradient
centrifugation,
rather thetext.(A)
Enzymeand20pg
ofE.colitRNAvalJ(B)than gel filtration, was used for analysis ofA.MV enzyme and 15pg ofyeast tRNAPhe; (C) enzymeand
reversetranscriptase-tRNA complex formation 52pgof totalyeasttRNA; (D) enzyme and47pgof
(Fig. 5), we found that the AMV polymerase totalchickenliver tRNA; (E) enzyme.A66,
Absorb-could also bind tRNA species other than ance at660 nm.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.509.82.224.63.335.2] [image:4.509.283.426.337.578.2]218 PANET AND BERLINER J. VIROL.
I 196 BSA the reverse transcriptase in virions of AMV.
30- A f Figure8A shows that all the reverse
transcrip-o 20 *; MvLV tasereleased from AMV after detergent
treat-x )'7\/ mentmigratesonglycerol gradients as a protein
E 10 i \_ $ with asedimentation coefficientof 9S,consistent
o
withacomplex
ofenzyme(7.2S)and tRNA(Fig.
i20
B ° 8C andD).
Addition ofexogenoustRNAtotheMuLV+
tIUA
virus
does notfurther increase thesedimenta-10
lo
if
\ z tionrate of thecomplex (Fig. 8B).Itisinterest-o '--.--.-. w ing to note that a virion of AMV contains
be-z 6- C a tween20and80molecules ofreverse
transcrip-4ulV
enzyme+
tUNA tase (14, 15) and some 35 to 70 molecules of 15MuLY 0
_4. tRNAspecies; ofthese, only 7 to 10 molecules
X2 / i 9Z \ 0.2 o are the primer tRNAT'P (20). The observation
I
0 I~~m S -3,00thatall
the released virionenzymeiscomplexed
0 10 20 30 to tRNA further strengthens the notion that
[image:5.509.87.246.49.239.2]FRACTION NUMBER many tRNA species, besides tRNATrP, can bind
FIG. 6.Binding of tRNA to reverse transcriptase to the reverse transcriptase when glycerol
gra-in virions. Conditionsforbinding reactionsare de- dients are beingused as an assay system.
scribed in the text. (A)MoloneyMuLV (250agJ was
treated with0.2%(vol/vol)TritonX-100 in a binding DISCUSSION mixture. After incubation for 10 min at 4°C, the
mixture wascentrifuged for 20 min at 6,000 x g, and In this study wehave shown that the reverse the clearsupernatant was layered onto a glycerol transcriptaseof mammalian RNA tumor viruses
gradient. (B) Virions(250gj were treatedwith Triton exists in two forms; the purified enzyme is a
X-1OO)asdescribed in(A)in thepresence of30()
lg
of single polypeptide with a molecular weight ofrat livertRNA. (C)Forty unitsofpurifiedMoloney 70,000,whichisconvertedinto a high-molecular-MuLV reverse transcriptase and 30 pg of rat liver weight complex after binding of tRNA. The tRNA. Protein determinations were not done with exactstructure of this complex is not yet
known,
gradients A and B, since virionproteins interfere butaccording to its molecular weight(150,000)
with the assayofthemarkers.A66o
,.
Absorbance at twoarrangementsarepossible:i)
anaggregate 660 nm.result that it sediments at the position of the A BSA
purified reverse transcriptase-tRNA complex 3 A
(Fig. 6B andC).Theseobservations suggest that X S
S'
some polymerasemolecules released fromviri- E 2 E
onsare
complexed
withtRNA and thusmigrate
1 - / > .faster in
glycerol gradients.
However, to quan-_ ctitatively complex
all of the virion enzyme, ad- i o-ditional exogenous tRNA is needed. It ispres- o + 4
entlyunknownwhether the enzyme is boundto a. 4 / z
tRNA in the virion or complexes are formed ° 2
subsequenttovirusdisruption. z p
w
To determine whethermammalian RNAtu- C a
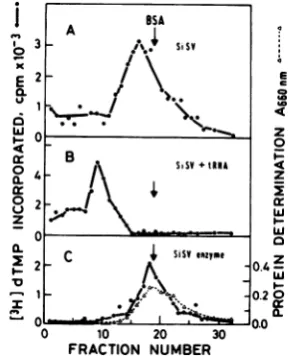
mor virus reverse
transcriptase
otherthan the 2 C 4sisY
@RFme
0.4z
Moloney MuLV enzyme can also bind tRNA, X
we chose a primate virus, SiSV. The results 10.20
presented
inFig.
7show that whereaspurified
,.o
0SiSVreversetranscriptasemigratesas aprotein 0 10 20 30
withamolecularweightof70,000 (usingbovine FRACTION NUMBER
serumalbuminas
marker),
theenzymereleased FIG. 7. Interaction between SiSV reversetran-fromSiSVvirions is somewhatheavier(Fig.7A). scritaseand tRNA. Conditionsforbindingreactions After addition ofexogenous tRNA, the DNA andglycerolgradientcentriAgationaredescribed in
polymerase of SiSV sediments as a proteinof the text. (A)SiSV(125 ag)wastreated withTriton
X-p1(Xecunr
150,00mlecuarwigh (Fi. 7B. Thstheexer-in
thedlegendlayeredtoontoFig.6;
aglycerolgradient(B)SiSV
(125
pg)aswasdescribedtreatediment demonstrates that thereversetranscrip- withTriton
X-.(X)
inthepresence of300jig
ofrat liver taseofaprimate
viruscanbindtRNAsimilarly
tRNA asdescribed in the legend to Fig.6;
(C)purifiedtotheenzymeofamurinevirus. SiSVreversetranscriptase(25units).
A6w,,
Absorb-We havefurtherinvestigatedthestructureof ance at 660nm.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.509.286.431.384.565.2]of two subunits with one or two tRNA molecules, to the enzyme. (ii) Thepattern of the
sediment-or(ii) several tRNA molecules bound to a single ing enzyme complex suggests a unique species
polypeptide. If the mammalian virus reverse with a molecularweight of 150,000; nonspecific
transcriptase-tRNA complex is indeed a dimer, aggregation is more likely to produce multiple
then it maybecomparedtotheholoenzyme of species. (iii) Proteins such as bovine serum
al-avian sarcoma leukosis viruses. The avian hol- bumin, IgG, oreven the isolated a subunit of
oenzyme is composedoftwosubunits, aand /1 AMVreversetranscriptase (5, 7)do notcomplex
(4), or
,B,,
in Rous sarcomavirus propagated in with tRNA. (iv) Reverse transcriptase of Rousduck celUs (9). In experiments in which tRNA sarcoma virus temperature-sensitive mutant
concentrations were low, the reversetranscrip- ts335is thermolabile for both DNA polymerase
tasecomplex sedimented at an intermediate po- activity (24) and the tRNAbinding function (A.
sition. Such sedimentation may reflect forma- Panet,G. Weil, and R.Friis, manuscript in
prep-tion ofa complexwith adifferent structure or aration). (v) Inhibitors of reverse transcriptase
dissociation of the dimer during centrifugation. activity such as monospecific antibody and
N-It may be argued that the association of re- ethylmaleimide also inhibit the tRNA binding
verse transcriptase and tRNA represents non- function (7).
specific aggregation of protein and nucleic acid; The fact that crudereverse transcriptase
re-there are,however,several lines of evidence that leased from virionssediments faster than
puri-indicatespecificity in this interaction. (i) Com- fiedenzymesuggests that it is bound to tRNA,
plexformation requires intact tRNA, whereas and, during enzyme purification, the tRNA is
fragments of tRNA(7), oligo(dT), or 16S rRNA removed and the two-subunitenzyme of
mam-of E. coli(unpublished observation) do not bind malian viruses dissociates. On the other hand,
thetwosubunits of theavian virus enzyme may
hold
togetherevenwithout tRNA. It is not clear60- 196 touswhy inaprevious studymouseliver tRNA
fi
A
AMVdid
not interact with MuLV reversetranscrip-T40
_A
A tase (7). The quality of the tRNA preparationo /, or the differentcomposition of the glycerol
gra-x 20 - dients used for theassay may have been
respon-Efi
O
sible for theseeming
lack of interaction.u There are several studies in the literature
d 40 B AMV +tllA
regarding
thestructureofthereversetranscrip-2 / tase in virions and
cells.
Mondal et al. (13) have<a
20- /\reported
the existence of two enzymes inlym-2
0 I .
./
phosarcoma cellsproductively
infectedwithgib-u c Al enzym the reversetranscriptase isolated from
virions,
z5 / *^ with a molecular weight of 70,000; and (ii) a
cL
5high-molecular-weight
enzyme(140,000)
that is x .__v._/able~ to copy the 70S RNA genome veryeffi-1 C
ciently.
Thislatter form could bedissociatedby
1 2 D r>t"1e
high-salt
and detergent treatment to yield thew
\70,000-molecular-weight
enzyme. Based on the1 _, results presentedhere, we suggest that the
high-I.
molecular-weight
reverseranscriptase
found in00 10 20 30
productively
infected
cells(13)
orinRoussar-FRACTION NUMBER coma virus-disrupted
virions
(2) is indeed an [image:6.509.83.224.307.544.2]enzymecomplexedtotRNA.The association of
FIG. 8. Binding of tRNA toreverse transcriptase tRNAwith the reverse transcriptase in
produc-in disrupted virions ofAMV. Conditionsfor tRNA tively infected cells could provide the
mecha-bindingare described in the text. IgGmarkerwas nism responsible for the incorporation ofcertain added to reactions (C) and(D) only, and glycerol
cell
tRNAspecies into thebudding
virus. Suchgradientswerecentrifuged for 20h (A)
MAMV(50)iw9)
amechanism,
however,
required some tRNAwas treated with Triton X-100 asdescribed in the * * ' * * r
legendtoFig. 6;(B) AMV
(5() g)
was treatedwithspecies
specificity lIn
thebidig reaction,isince
TritonX-1IK)in thepresence of125pgof chicken liver a specific subset of the celi tRNA's is incorpo-tRNA as described in thelegendtoFig. 6;(C) purified rated into virions (20).
AMVreverse transcriptase (50units) and 50pg of AMVreversetranscriptasewasshowntobind chicken livertRNA; (D) purified AMV reverse tran- only tRNATrp andtRNA4Met withhigh affinity
scriptase
(50units).
(16). In the present study we found that theon November 10, 2019 by guest
http://jvi.asm.org/
220
AMVenzymecanalso bindother tRNAspecies, 9.Hizi, A., J. P. Leis, and W. K.Joklik. 1977. RNA althoughpossiblywith lower
affinity.
Such bind- dependent DNA polymerase of ASV. Comparison ofing
was observed whenhigh
concentrations of the catalytic properties of the a, /2 and 8 enzymeingwasobservedwhen high ~~~~forms.J. Biol. Chem.252:2290-2295.
tRNAwereaddedtothepurifiedAMVenzyme 10.Keith, G.,A.Roy,J. P.Ebel,and G.Dirheimer. 1971.
andcomplexformationwasassayedusing glyc- The nucleotide sequence oftwo tryptophan tRNAs
erolgradient centrifugationrather thanSepha- from brewer yeast. FEBSLett.17:306-308.
dex gel filtration (16). This apparent lack of 11.Lowry,0.H.,N.J.Rosebrough,A. LFarr,and R. J.
dex.gelifitration(16).
This apparent lackof Randall. 1951. Proteinmeasurement with theFolinspecificity isreminiscent of the interaction be- phenol reagent.J.Biol. Chem.193:265-275.
tweenaminoacyl-tRNA synthetasesand tRNA. 12. Moellng,K. 1974. Characterization ofreverse transcrip-The latterenzymesbind theircognatetRNA's taseand RNase H from Friend-murine leukemiavirus.
tighlyut an lsoassociatewithother tRNA
1.Virology
62:465-59.tightly but can alsoassociate with other tRNA 13
Mondal, H.,
R. E.Gallagher,
andR.C.GaBo.
1975.specieswith loweraffinity (25).
Furthermore,
we RNAdirected DNApolymerasefromhumanleukemiccouldnotdemonstratetRNAspeciesspecificity bloodcells and fromprimate type-Cvirus-producing
inthecomplexfonnedwith MuLVreversetran- cells:high and low molecularweight forms withvariant
scriptae.Ifideedte affinty of te MuLV biochemicalandimmunological properties.Proc. Natl.
scriptase. If indeed the affinity of the MuLV Acad. Sci. U.S.A.
72:1194-1198.
enzymetoitsprimer tRNAP"oishigherthanto 14. Mosser,A.G.,R.C.Montelaro,andR. R.Rueckert.
other tRNA species, otherexperimental condi- 1977. Proteins of Rous-associated virus type61:
poly-tions willhave tobesought.Sephadex gelfiltra- peptide stoichiometry and evidence thatglycoprotein
tion, for example, failed to show any complex
gp35
is not a cleavage product ofgp85.
J. Virol.fonnationetween[3p]tRNA an the MuLV 23:10-19.
formation between [32P]tRNA and the MuLV 15. Panet, A.,D.Baltimore,and T.Hanafusa. 1975. Quan-reversetranscriptase. titation of avian RNA tumor virusreversetranscriptase
byradioimmunoassay.J.Virol.16:146-152.
16. Panet,A.,W.A.Haseltine,D.Baltimore,G.Peters, ACKNOWLEDGMENTS F.Harada,and J. E.Dahlberg.1975.Specific binding This workwassupportedbygrantsfromStiftungVolkswa- oftryptophan transfer RNAto avianmyeloblastosis genwerk and The IsraelCommissionforBasicResearchu virusRNA dependent DNA polymerase (reverse tran-Wethank N.Goldblum for hishelpf scriptase). Proc. Natl. Acad. Sci. U.S.A.72:2535-2539.
Wethak
NGdbumforhihep.17.
Peters,G.,F.Harada,J. E.Dahlberg,
A.Panet, W.A.Haseltine,and D.Baltimore.1977.
Low-molecu-lar-weightRNAs ofMoloney murineleukemiavirus:
LITERATURE CITED identification of the primer for RNA-directed DNA 1. Dahlberg,J. E., R. C. Sawyer,J. M.Taylor,A. J. synthesis. J. Virol.21:1031-1041.
Faras, W.E.Levinson,H. M.Goodman,and J. M. 18. Rogg,H.,W.Wehrli,and M.Staehelin.1969. Isolation Bishop.1974.TranscriptionofDNA from the 70SRNA ofmammalian transferRNA.Biochim.Biophys.Acta ofRoussarcomavirus.I.Identificationofaspecific4S 195:13-15.
RNA which serves asprimer. J. Virol. 13:1126-1133. 19. Ross, J., E. M. Scolick, G. J. Todaro,and S. A. 2. Duesberg, P.,K. V. D.Helm,and E.Canaani. 1971. Aaronson. 1971. Separationofmurine cellular and Properties ofasoluble DNApolymeraseisolated from murineleukemiavirus DNApolymerases.Nature (Lon-Rous sarcoma virus. Proc. Natl. Acad. Sci. U.S.A. don) New Biol. 231:163-167.
68:747-751. 20. Sawyer, R.C.,and J. E.Dahlberg.1973.Small RNAs 3. Fan, H.,and M. Paskind. 1974. Measurementofthe ofRoussarcomavirus:characterization by two-dimen-sequencecomplexityofclonedMoloneymurineleuke- sional polyacrylamide gel electrophoresisand finger-mia virus 60 to70sRNA: evidenceforahaploidge- printanalysis. J. Virol.12:1226-1237.
nome.J.Virol.14:421-429. 21.Taylor,J.M.,and R.Illmensee.1975.Site on the RNA
4. Grandgenett,D.P., G.F.Gerard,and M. Green. 1973. ofanaviansarcomavirusatwhich primer isbound. J. Asinglesubunit from avianmyeloblastosisvirus with Virol. 16:553-558.
bothRNA directed DNApolymerase andribonuclease 22. Verma, I. M. 1975.Studiesonreversetranscriptase of Hactivity. Proc. Natl. Acad. Sci. U.S.A.70:230-234. RNAtumor viruses.III.Properties of purified Moloney 5. Grandgenett,D.P.,A.Vora,andA. J.Faras.1976. murineleukemia virus DNApolymerase and associated
Different states of avian myeloblastosis virus DNA RNase H. J. Virol.15:843-854.
polymerase and their binding capacity to primer 23. Verma, I. M., and D.Baltimore. 1973. Purification of tRNA'P.Virology 75:26-32. theRNA-directedDNA polymerasefrom avian
mye-6. Harada, F., R. C. Sawyer, and J. E. Dahlberg. 1975. loblastosis virus and its assay withpolynucleotide
tem-Aprimer ribonucleic acid for initiation ofinvitro Rous plates. MethodsEnzymol.29:125-131.
sarcomavirusdeoxyribonucleicacidsynthesis.J.Biol. 24. Verma, I. M., W. S. Mason, S. D. Drost, and D. Chem.250:3487-3497. Baltimore. 1974. DNA polymerase activity from two
7. Haseltine,W.A.,A.Panet,D.Smoler,D.Baltimore, temperature sensitive mutants of Rous sarcoma virus is G.Peters,F.Harada,and J. E.Dahlberg.1977.The thermolabile. Nature (London) 251:27-31.
interaction oftryptophan tRNA and avian sarcoma 25. Yarus, M. 1972. Solvent and specificity. Binding and virus reversetranscriptase: furthercharacterizationof isoleucylationofphenylalaninetransfer ribonucleic acid thebinding reaction.Biochemistry16:3625-3632. (Escherichia coli) by isoleucyl transfer ribonucleic acid
8. Hirsh, D.1971.Tryptophantransfer RNA as theUGA synthetase from Escherichia coli. Biochemistry suppressor. J. Mol. Biol.58:439-458. 11:2352-2361.
on November 10, 2019 by guest
http://jvi.asm.org/