0022-538X/79/12-0796/07$02.00/0
Polypeptides
of
Hepatitis
B Virus Surface
Antigen Produced
by
a
Hepatoma Cell Line
PATRICIA L.MARION,' FELIX H.SALAZAR,' JENNIFER J.ALEXANDER,2 ANDWILLIAM S.
ROBINSON'*
Departmentof Medicine,Stanford University, Stanford, California94305,1and National Instituteof
Virology, Johannesburg, SouthAfrica2
Received for publication25April1979
ThePLC/PRF/5cell linederived fromahumanhepatomaproduces hepatitis
Bsurface antigen (HBsAg) in 22-nm particles of the same buoyant density as
thosefound in the serum of infectedpatients.TheHBsAg particlesfrom this cell
linewere labeled with[3S]methionine andpurified,and the polypeptideswere
compared by sodium dodecyl sulfate-polyacrylamide gel electrophoresis with those of serum-derivedparticles. The two major polypeptides of serum-derived
HBsAg particles (p20and p23)werefound in the same relative amountsin the
particles from the cell line. The three smallest of the five minor components
observed inHBsAg particlesfrom serum were present inparticlesfromthe cell
line. These polypeptides (p31, p36, and p43), as well as p20 and p23, were
precipitated with anti-HBs-containing serum. The two largest polypeptides of serum particles (p49 and p66) were not detected in particles from these cells. When the PLC/PRF/5 HBsAg particles were radiolabeled with tritiated sugars, p23,andnotp20,wasfoundtocontainradioactivity, indicating that thepattem
ofpolypeptide glycosylation is similartothat ofserumHBsAg. None of the other possible gene products of hepatitis B virus was detected in the
PLC/PRF/5-derivedHBsAg particles,inthecells,orinthe cellsupernatants.
HepatitisBvirus(HBV)infection inhumans
isaccompanied bythe appearance of viral anti-gens in liverand blood. Several viral forms in
the blood contain hepatitis B surface antigen (HBsAg).Oneforn,the Daneparticle,contains
an intemal antigen designated the hepatitis B coreantigen (HBcAg), DNA, andDNA
polym-eraseactivity and is consideredtobe the
com-plete virion (18).Athird antigen, hepatitisB e
antigen (HBeAg), canbe detected in the blood ofsomeinfectedpatients(13). Recent evidence indicates thatHBeAg is foundin acryptic form in the interior of the Dane particle (22). The
mostabundantHBsAgformsare22-nm
spheri-calparticlesandfilamentous formsconsisting of
protein, carbohydrate, andlipid, and these are considered to be defective or incomplete viral forms withoutHBcAgorDNA.Several
polypep-tides isolated from these particles have been
shown tocontain both group- and type-specific
HBsAgdeterminants (4,5, 19). Ininfectedliver,
HBsAgisdetected incellcytoplasmand atcell surfaces, and HBcAg is detected in nuclei by immunofluorescence (1, 6).
Numerous studies have demonstrated an as-sociation betweenpersistentHBV infection and
primaryliver cancer in some parts of theworld
(2, 8, 21).Itis not known whether the
relation-shipisacausalone.In1976,atissue culturecell
line designated PLC/PRF/5 was isolated from
a primary liver carcinoma of an African man
withpersistentHBVinfection. The cell linewas
subsequentlyshowntoproduce smallamounts
of HBsAg, apparently in the form of 22-nm
particles (12). The antigenic subtype has been
reportedtobe ad (12). The other antigens
asso-ciated withHBV, HBcAg and HBeAg,were not
detected in the cells. These findings indicate that thecells containatleastsomeHBVgenes.
Since cellsin culture havenotbeen
success-fully infected with HBV,PLC/PRF/5cells offer the firstopportunity tostudyHBsAg formation in tissueculture.
HerewedescribeexperimentsinwhichPLC/ PRF/5 cells were incubated with radiolabeled methionine orsugars, the HBsAg particles were
purifiedfromculturemedium, andthe
radioac-tive polypeptideswereanalyzedby sodium
do-decylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) andcompared withHBsAg parti-clespurifiedfrompatientserum. Thecells and culture medium werealsoassayed for Dane
par-ticles, HBcAg,and HBeAg.
MATERIALS AND METHODS Ceilculture. ThePLC/PRF/5cellswere grown in Dulbecco-modifiedEagle medium supplementedwith 10% fetalbovine serum.Thecellshavebeen passaged 796
on November 10, 2019 by guest
http://jvi.asm.org/
HBV POLYPEPTIDES PRODUCED BY HEPATOMA CELLS 797 weeklyand are nowatpassage 44 in our laboratory.
Production of HBsAg has remained at a constant level. Thecells are infected with mycoplasma.
Labelingof cells. At2 to 3days after reaching confluency in 10-cm culture dishes, PLC/PRF/5cells
were incubated with 50
lACi
of L-[3S]methionine (Amersham Corp., 800 to900 Ci/mmol) in 10 ml of methionine-free medium supplemented with 1% me-thionine-containing medium and 10% fetal bovine se-rum.After3days, the medium was replaced with fresh medium containing L-[3S]methionine and incubated foranother2days. Thepooledradioactivecell super-natants werecentrifuged at 12,000xgand4°C for 30 min to removecellular debris.Similar cultureswereincubated withL-[6-3H]fucose (AmershamCorp., 17Ci/mmol) and
D-[6-3H]glucosa-mine(AmershamCorp.,20Ci/mmol) (20uCieach)in 10 ml ofDulbecco-modified Eagle medium for the sametimesdescribed for[3S]methionine.
DetectionofHBsAg. Solid-phase radioimmunoas-say(AusriaII,AbbottLaboratories)wasusedtodetect HBsAg. Samples from CsCl gradient fractions were diluted in TNE buffer(0.01MTris-hydrochloride,pH 7.5; 0.15 MNaCl;0.001MEDTA)atleasteightfold to avoid the effects of concentrated CsCl onthe assay.
Purification of HBsAg. Clarified cell superna-tants from PLC/PRF/5 cells were centrifuged at 25,000 rpm and4°Cfor24h inaSpinco SW27 rotor to pellet the HBsAgparticles. All but0.5 ml of the supernatantswereremovedbysuction; the remaining 0.5mlcontained concentrated fetal calfserumproteins andlooselypelleted HBsAg particles.Thepellets were suspended in the remaining supematant and were made up to 7 ml with TNE buffer. Solid CsCl was addedtogiveadensity of1.2g/ml, and the material wascentrifuged at 45,000 rpm and 4°C for 68 h in a
Spinco50Tirotor.Fractionswerecollectedfrom the bottomof the tube andassayedforradioactivity and HBsAg. Peak fractions ofHBsAgwerepooled,diluted to 7 ml with CsCl at 1.2 g/ml in TNE buffer, and
centrifugedasecondtime. TheHBsAg peakfractions were again pooled and dialyzed at 4°C overnight
against TNE buffer to remove CsCl. The material
(approximately2ml)wasthenlayeredontop ofa 33-mlpreformed 5 to20%sucrosedensity gradient con-tainingTNE buffer and1mgof bovineserumalbumin per mlandcentrifugedat25,000rpm and4°Cfor5h inaSpinco SW27rotor. HBsAgpeak fractionswere
dialyzedovernightagainstTNEbuffertoremovethe sucrose.TheHBsAgwasthen banded twicemorein CsCldensity gradientsasdescribed above. The frac-tions containing purified HBsAgparticles from the finalgradientwerethenpooledandkeptat4°Cuntil used.
HBsAgfromserum waspurifiedin thesame man-ner,but aLowryproteinassay(10)wasusedtodetect total protein in gradient fractions, and HBsAg was
detectedbycomplementfixation.
Antisera. Aguinea pig anti-HBs/adw-containing
serumwas agiftfromJohnGerinof the OakRidge
NationalLaboratory. Theserumwasabsorbed with rabbit liver powder and has a complement fixation titerof1:32.
The immunoglobulin G fraction of serum from HBsAg carrierswithhightitersofanti-HBcwas pre-pared by ammonium sulfate precipitation and
em-ployed in asolid-phase radioimmunoassayforHBcAg (16).
Sera from HBsAg carriers with high titers of anti-HBe were used to detect anti-HBeAg by double immuno-diffusion (detection to a serum dilution of 1:128) and by anenzyme-linked immunosorbent assay (14) used in thislaboratory (detection to adilutionof1:64,000). PAGE. Purified HBsAg particles in CsCl were di-luted 1.5-fold with TNE buffer, and 20
jig
each of bovineserumalbumin and cytochrome c were added. Atotal of5to 10volumes of ethyl alcohol (EtOH) was added, and the precipitated protein was pelleted bycentrifugation for 10min at 10,000 x g. The pellets
weresuspended insample buffer (0.07 M Tris-hydro-chloride, pH6.8; 11% glycerol; 0.0015% bromophenol blue; 3%SDS; 10% ,B-mercaptoethanol) and heated at 100°C for 3 min.
Thepolypeptides were separated by electrophoresis invertical slabgelsasdescribed by Laemmli (9). Gels of13or 8 to20%polyacrylamide were runusuallyat 19mA/13.5 cmfor 5 h. Gelswerefixed in 20% meth-anol-7% acetic acid, stained in 0.2% Coomassie bril-liant blue in 50% methanol-7% acetic acid, and de-stained in the fixative. To enhance detection of radio-activity, we performedfluorography as described by Bonner andLaskey(3)onallgels before drying with a slab gel dryerfrom Hoefer Scientific Instruments. The driedgels were exposed toXR-5X-rayfilm (East-man Kodak Co.) at -70°C and developed. Protein standards of known molecular weightswere fB-galac-tosidase, bovineserumalbumin, ovalbumin, aldolase,
chymotrypsinogenA, and cytochrome c.
Immunoprecpitation.Antigenswere
immunopre-cipitatedwithstaphylococcal protein A prepared by the method of Kessler (7). One volume of HBsAg
particleswas diluted with either 3 volumes of TNE bufferplus0.1%,8-mercaptoethanoland1mgof bovine serumalbumin per ml(TNEMEBSA)or2volumes of 1XRIPA buffer(0.01 M Tris, pH 7.2; 0.15 M NaCl; 1%
deoxycholate; 1% Triton X-100; 0.1% SDS) plus 1 volume of2x RIPA buffer. Serum containing
anti-HBs(251d/ml)wasadded,and the mixturewas
incu-bated for1hat37°C.Asuspensionofprotein A (10%,
vol/vol)wasthen added(50
p1/5
p1
ofantiserum), and the mixturewasincubated for30minat room temper-ature.Aftercentrifugation (10,000rpm inaBrinkman 3200 centrifuge for 10 min), the pellet was washed oncewith0.5ml ofTNEMEBSAor lx RIPAbuffer,repelleted,andresuspendedin50pl ofsample buffer
containing6%SDS.Sampleswereheatedat100°C for 3 to 5min, theproteinAwasremovedby centrifuga-tion, andsampleswereanalyzed bySDS-PAGE.
RESULTS
Purification of HBsAg particles from PLC/PRF/5 cells. As culture fluids of PLC/
PRF/5 cells contained greater amounts of
HBsAg than did disrupted cells, these fluids
wereusedasthe sourceofHBsAgparticles for
purification.After removalof cellular debris
by
low-speed centrifugation, the HBsAg particles were pelletedin theultracentrifuge. The
parti-cles were then purified as described above by
fourequilibriumcentrifugationsinCsCl andone
VOL. 32,1979
on November 10, 2019 by guest
http://jvi.asm.org/
798 MARION ET AL.
rate sedimentation in a sucrose gradient. The
finalCsCl gradientservedmainlyto concentrate
the particles without the losses incurred when other concentratingmethods, such aspelleting orultrafiltration,wereused.
The firstand lastCsCl centrifugationsteps of
[35S]methionine-labeled HBsAg particle purifi-cation are shown in Fig. 1. Figure 1A shows
trichloroacetic acid-precipitable 35S andHBsAg
measured by solid-phase radioimmunoassay in the fractions of the firstgradient. The HBsAg particles accounted forsucha smallfraction of theacid-precipitable[35S]methioninethatthese
particles couldnotbedistinguishedas aseparate 35S component in this early purification step.
35S-labeled HBsAg particlesbecame detectable
onlyatthe third or,rarely, secondpurification
step. In the final gradient (Fig. 1B), the major 35Speakandthat ofHBsAgcoincided.Thepeak
fractions ofthe final gradientwere pooled and
stored at 4°C. Several purification steps for HBsAg were needed because of thelargeamount
ofnon-HBsAg 35S-labeled material released by these cells. 35S-labeled HBsAg represented about1% of thetotaltrichloroacetic acid-precip-itableradioactivityinthe clarified cell
superna-tants. SDS-PAGE analysis of the non-HBsAg 35S-labeled material revealed a continuous
dis-tribution ofpolypeptideswithmolecularweights
of 5,000(5K) to100K,and nopredominant com-ponents were visible.
HBsAg particles from serum were found at
the same buoyant density (1.23 g/ml) in CsCl
densitygradients.
Comparison of the polypeptides of [36S]-methionine-labeled HBsAg with HBsAg from human serum. The polypeptides of pu-rified [35S]methionine-labeled HBsAg particles
(subtype ad) fromPLC/PRF/5cellsand of
pu-rifiedHBsAg(subtype adw) from patient serum that was HBeAgpositivewereanalyzed by
elec-trophoresisin an 8 to20%SDS-polyacrylamide gradient gelasdescribed above.
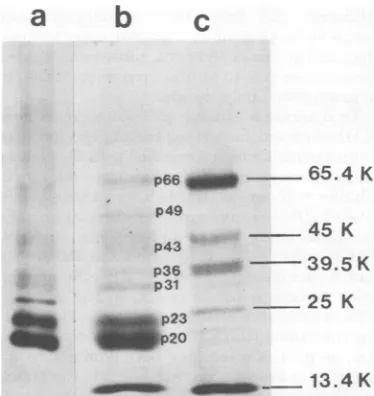
The polypeptides of the serumHBsAg prep-aration, shownwithcytochrome c in Fig. 2, track b, appear to be similar to previous findings of
others (18).Twopolypeptidescorresponding to
molecular weights of 20 and 23K (p20 and p23)
constitute the majority ofthe Coomassie
bril-liantblue-staining material. p20was presentin
greater amounts thanp23. Several minor
Coo-massiebrilliant blue-stainingcomponents in
po-sitionscorrespondingtomolecularweights of 31, 36, 43,49,and66K(p31, p36, p43, p49, and p66) were also regularly observed. p66 comigrates with human serum albumin. An eighth
poly-peptide migrating at 97K was frequently
ob-served, althoughnotin thegelshown.
5 l0 15 20
m z U,
3
0 0
u 0
o
0
E
CL)
C)
5 10 15 20
FRACTION NUMBER
FIG. 1. Isopycnic centrifugation of 35S-labeled HBsAg particles in CsCl gradients. (A) The first equilibrium CsCldensitygradient centrifugation for 35S-labeled HBsAg purificationwascarriedoutasdescribedin the text,andfractionswerecollectedfromthe bottomof the tube. The trichloroaceticacid-precipitable
radioac-tivitywasdeterminedon1,il of each fraction, and theamountofHBsAg activitywasmeasuredineveryother
fraction by radioimmunoassay after 100-fold dilution of2.5gil.(B) The final equilibrium CsCl density gradient
centrifugationstep in 5S-labeled HBsAgparticle purificationwascarriedoutasdescribed in thetext, and fraction collection,radioactivity, andHBsAgassayswerecarriedoutasdescribed in (A), using 5 ,ul for the 35S
determinationand 5,uldiluted50-fold fortheantigenassay.
0
o
0x
E
C')
C/)
U-ur
i m z *I
--3
0 0
u
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.70.457.376.575.2]HBV POLYPEPTIDES PRODUCED BY HEPATOMA CELLS 799
In Fig.2, tracka, thepositions of
[3S]methi-onine-labeledpolypeptidesfrom the PLC/PRF/
5 HBsAg preparation electrophoresed in the samegelcanbe observedafterautoradiography
tobesimilartothepositions ofthe polypeptides
of serum-derived HBsAg, with certain
excep-tions.p20 andp23are presentin thesame
gen-eral proportions.p31,p36, and p43 are also pres-ent, although the high background makes it
difficult todistinguish them. The two prepara-tionsdifferedbythepresence of a 27K (p27)
3S-polypeptide less heavily labeled than p20 and p23, butmoreinevidence than the otherminor componentsandnotdetected by Coomassie
bril-liant bluestaining of the serum HBsAg
polypep-tides. p27 was present in the same relative amountsintwootherpreparations of 3S-labeled HBsAgparticles.
CarbohydrateinHBsAgfromPLC/PRF/
5cells. One of themajor polypeptides of puri-fiedserumHBsAg (our p23) has been observed by others (18)tostain withperiodic acid-Schiff,
c
p66 _I 65.4 K
p49
p43
u.,.A.i ...~~~~~1.
p31
.*..w
_p~~~23
_p20
45 K 0
I
0
--- -39.5K Ci
0
_~25 K x
E
-t)
[image:4.504.53.240.303.501.2]_,1 .413.4K
FIG. 2. Comparison ofthepolypeptides of HBsAg particles from PLC/PRF/5 cells and from human
serum.HBsAg particles fromserumand
[uS]methi-onine-labeledparticles fromPLC/PRF/5 cellswerepurifiedandprecipitatedwith EtOHasdescribed in the text, and thepolypeptides ofeachwereseparated on an 8 to 20% SDS-polyacrylamide gradient gel.
Electrophoresis was carried out for 16 h at 40 V. (Track a) Polypeptides of 3S-labeledHBsAg parti-cles(8,000 cpm) after fluorography for3weeks; (track b) polypeptides ofserum HBsAg particles stained with Coomassiebrilliantblue; (track c)marker
pro-teinsrunin thesamegelandstained with Coomassie brilliant blue were bovine serum albumin (65.4K), ovalbumin(45K),aldolase(39.5K), chymotrypsinogen A(25K),andcytochromec(13.4K).
suggesting thatit was glycosylated. Minor com-ponents migrating at 27, 32 or 35, and 53.3K
have also appeared to contain carbohydrate in
previous studies (18).
Todetermine whether any of the polypeptides in the HBsAg preparation from PLC/PRF/5
cellswere glycosylated, we incubated the cells with both
L-[3H]fucose
andD-[3H]glucosamine
andpurified the HBsAgparticles in the culture
supernatants as described above. The 3H and
HBsAgprofilesinthe finalCsCl gradient
coin-cidedasshown inFig.3.The3H-labeledHBsAg
particleswere EtOH precipitated andanalyzed
by SDS-PAGE alongside EtOH-precipitated
[35S]methionine-labeled HBsAg.Amajor
prob-lem with this analysiswastheverylowlevel of
radioactivity incorporatedintoHBsAgparticles
in these experiments. Although 2.5 mCi of
[3S]methionine
incubated with 108 cellsas de-scribed yielded a purified particle preparation containingapproximately150,000 cpm,thesame amountof tritium in thesugarsusedatthesamespecific activity yielded no -more than 37,000
cpminpurified particlesinanyexperimentand
required fluorography for detection by
autora-diography. Thepattem of3H-labeled
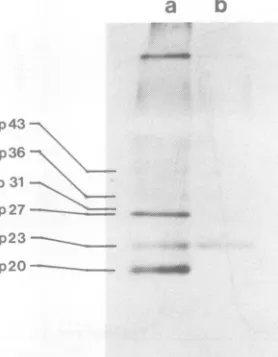
polypep-tidescanbeseenin track b ofFig.4and
com-pared with the 3S-labeled polypeptides shown intracka.Amajoramountof3Happearsinp23,
i1.4
1.3
a
1.2 zn
z
en
1.1
0
3 I
o
5 10 15 20 25
FRACTION NUMBER
FIG. 3. Final isopycnic banding ofHBsAg parti-cles labeledwithL-[6-3H]fucose andD
-[6-3H]gluco-samine. Theparticles were radiolabeled and
par-tially purified as described in the text. Fractions collectedfromthefinalCsCl density gradient were
analyzed for radioactivityandHBsAgasdescribed in thelegendtoFig.IB,exceptthat the3Hcontentof 10,ul ofeachfractionwasmeasured.
a
b
VOL. 32,1979
A
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.253.448.367.579.2]800 MARION ET AL.
andnoneis detectedinp20.Althoughnotvisible
inFig.4,radioactivityis alsopresent inp3l and
p36,indicating that thepattemofglycosylation
inthePLC/PRF/5 HBsAgpolypeptidesis
sim-ilar tothat ofHBsAg from patientserum.
Immunospecificity of labeled
polypep-tides ofPLC/PRF/5 HBsAg. To determine which35S-labeledpolypeptides haveHBsAg
de-terminants, wedissociated 35S-labeled
polypep-tides ofpurifiedparticleswith RIPA bufferand
immunoprecipitatedthem withaguinea pig
se-rum containing anti-HBs/adw and protein A.
Forty to fifty percent of the radioactivity in various particle preparations was precipitated
and recovered bythis method, comparedwith
50 to70%recoverybyEtOHprecipitation.The
SDS-polyacrylamidegelpattern ofpolypeptides
precipitated by HBsAg-specific antiserum is
shown inFig. 5, trackb,andcomparedwith the
same amountofEtOH-precipitatedHBsAgfrom
thesamepreparationin tracka.Thehigh
back-ground along the track of EtOH-precipitated
polypeptides is not present with the
immuno-precipitated
material, and only p20, p23, p31,p36, and p43aredetected in similarproportions
tothepolypeptidesofserumHBsAg.Thethird
major polypeptide found in EtOH-precipitated
HBsAg,p27, was notdetected, and thereare no
[image:5.504.289.434.62.206.2]visible bands corresponding to the serum-de-rived HBsAg components at 49, 66, and 97K.
FIG. 4. Patternofglycosylationofthepolypeptides
of HBsAgparticles produced byPLC/PRF/5cells. HBsAgparticles labeled withL-[3H]fucose andD
-[3Hlglucosamine (track b) and [35S]methionine-la-beledparticles(track a)werepurified,EtOH precip-itated, andanalyzed bySDS-PAGE(13% gel,19mA for5h) asdescribed in thetext. Bothsamples con-tained12,000 cpm, and thefluorogramswereexposed for3weeks.
a
ww
..:.
[image:5.504.92.231.387.566.2].-b
..d:05::.
:
p43--p36' p31
p27
-p23 p20
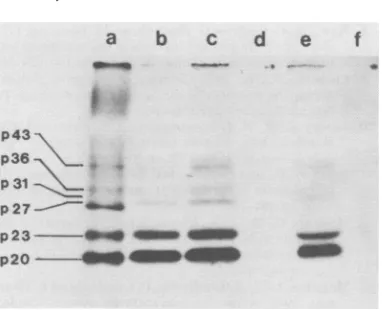
FIG. 5. Immunoprecipitation of35S-labeled HBs-Agpolypeptides with guinea pig anti-HBs/adw. Pu-rified HBsAg particles labeled with [3S]methionine weredivided intotwoparts. Thefirstpart(track a) wasconcentrated by precipitation withEtOH, and thesecond(trackb)wasprecipitated in RIPAbuffer with the anti-HBs andproteinAasdescribedin the text.The polypeptideswereanalyzed bySDS-PAGE asdescribed in thelegendtoFig.4.
Although p23 from the immunoprecipitate
seems to bepresent in agreater concentration
thanp20 in thisexperiment, subsequentstudies have shownp20 tobe thepredominant immu-noprecipitatedpolypeptide.
To determine whetherp27 was a component
ofHBsAgparticleslackingHBsAgspecificityor acontaminantwhichcopurified with the HBsAg particles,wecarriedoutimmunoprecipitationin abuffer withoutdetergents. Comparisonof35S-. labeled HBsAg afterimmunoprecipitationin the
non-dissociating buffer and analysis on SDS-PAGE withEtOH-precipitated and
RIPA-pro-tein A-immunoprecipitated
[35S]methionine-la-beledantigen is shown in Fig.6. The polypep-tides immunoprecipitated
under.non-dissociat-ingconditions (trackb) appear tobe thesame
(i.e.,nop27)aswhenthey were
immunoprecip-itated afterdissociation with RIPA buffer(track
c). Therefore,p27isprobablynot a component
ofHBsAg particles, but a contaminant of the
purifiedHBsAgpreparations.
Todeterminewhether p27 or anyof the other
polypeptidescontainedHBcAgorHBeAg
spec-ificities,weincubatedequal amounts of purified
t[S]methionine-labeled
HBsAg particles withpurified human immunoglobulin G containing
anti-HBc and with human serum containing
anti-HBe. The latterserum alsocontained
anti-HBs. After precipitation with protein A, the
polypeptides were compared by SDS-PAGE
with polypeptides from equal amounts of the sameparticle preparation precipitated,
respec-tively,withEtOH, anti-HBs(Fig. 6, tracks a, b, andc), and normal human serum (track f). It
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HBV POLYPEPTIDES PRODUCED BY HEPATOMA CELLS 801
P43
p36
P31
p27
- p23-p20 -4
FIG. 6. Im Agwithanti humanserun
ticles were d
wereprecipit
(track b) ant non-dissocial proteinA in
anti-HBc an
anti-HBe pit buffer, and (ti A in RIPA I thepellet wa
inthetextan
canbeseen
precipitated human seru
tides precipi by theserun
e). Thus, th HBcAg-reac HBsAg par
ments.
Wedidn(
ucts of HB cells. NoHI
noassay ofs
fold by ultr by freeze-tl cellsperml
materials A
bent assay.
measurable fluids.
We havei
particlespri
derived fron
the polypep
from human labeledpoly the minorp
imrlmunopre
distinguisha
a b c d e f from
polypeptides
inserum-derivedparticles
de-tected by Coomassie
brilliant
blue staining after SDS-PAGE. However, thehigher-molecular-weight minor polypeptides of serum HBsAg,
whichmigrate at49, 66,and 97K in oursystem, were not detectedinthePLC/PRF/5-produced HBsAg. One of the HBsAg polypeptides, p66, has been found
by
others to comigrate with humanalbumin, a protein foundinpreparations ofHBsAg
particles
purified
fromserum(20). p66__m
has been the componentvarying
the most inwe __ Z_ amount in different preparations (20). Its
ab-senceinthe hepatoma HBsAg raises the possi-bility that p66 has
actually
represented only human albumin in HBsAg preparations frommunoprecipitation of
35S-labeled
HBs-
serum in previous studies and that the HBsAg-HBs, anti-HBc, anti-HBe, and normal reactivity of p66 found in some experiments was
n.Purifiedand'S-labeled
HBsAg
par- only due to a trailing of lower-molecular-weightiivided into six equal fractions which antigen along the gel track. p27, a component
ated with the following: (track a)EtOH, fnd
in
ged
trag p27,aa
from
ti-HBs/adw serum and protein A in a found in
purified
HBsAgpreparations
from
ting buffer, (track c)anti-HBs/adw
with PLC/PRF/5 cellsandnotinHBsAg from serum, a dissociating buffer(RIPA),
(track d)appeared
tobe acontaminant,
since it failedtod protein A in RIPA buffer, (track e) precipitate when intact HBsAg particles were
us anti-HBs with protein A in RIPA immunoprecipitated with anti-HBs. Thus, it did rack f) normal human serum andprotein not react with anti-HBs and was not a
compo-5uffer. Each was then centrifuged, and nent of HBsAg particles.
sanalyzedbySDS-PAGEasdescribed The presenceof five separate
"S-labeled
pol-z4in thelegendtoFig.4. ypeptides in animmunoprecipitate
of purifiedthat nodiscerniblepolypeptide was particles, using anti-HBs-containing serumand Iwith anti-HBc (track d) or normal a
dissociating
buffer,
indicates thatmultiple
iin (track f), and only the polypep-
HBsAg-reactive
polypeptides
are found initated by anti-HBs were precipitated HBsAgparticlesfromhepatomacells, aswellas nwith anti-HBe and anti-HBs (track in serum
particles.
Thismultiplicity
isprobably
Lere was no evidence for HBeAg- or notdueto
degradation
by
serumproteases(11),
ctive polypeptides in the purified unless such proteases are also present in these ticle preparations in these experi-
PLC/PRF/5 cells.
Peterson et al. (15) have proposed that the ot find any other possible gene prod- two main
polypeptides
in the serumantigen
V associated with the PLC/PRF/5 particles
(our
p20
andp23)
differonly by
the BcAg was detectable by radioimmu-carbohydrate
moiety
found in p23. Thispost-supernatant fluids concentrated150- translational modification ofa
polypeptide
with acentrifugation or of cells disrupted HBsAgreactivity
canaccount for the presence iawing at a concentration of 2 x107 of at least these two components of different 1.No HBeAg was found in the same apparent molecular weight and similarantige-vith an enzyme-linked immunosor- nicity.Consistent with this conceptare our find-No DNA polymerase activity was ings that the hepatoma cells incorporate
3H-in the concentrated
supernatant
labeled sugars into p23 and notp20
and that both are precipitated by anti-HBs. As therela-DISCUSSION tive
proportions
ofp20
andp23appeartobethesame in HBsAg particles from both PLC/PRF/ radiolabeled and purified the HBsAg 5 cells and serum, these two components in this oducedby the PLC/PRF/5 cell line ratio may be a necessary feature of the particle n a human hepatoma and compared structure. How the remaining threeminor
com-tides with those found in particles ponents
(p31,
p36, and p43) may be related to iserum. The major[3S]methionine-
p20,
to p23, and to each other remains to be!peptides
(p20and p23) and threeofdetermined.
Since others have found similar an-olypeptides(p31,p36, andp43)
were tigenic specificity in several polypeptides iso-cipitated with anti-HBs and were in- lated from HBsAg particles (4, 19), different Lble in position and relative amount polypeptides must share some amino acidse-VOL. 32,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.61.252.58.213.2]802 MARION
quences,andone ormoreof thelarger
polypep-tides maybe uncleaved precursors of the smaller ones.
Although a large proportion of HBV (Dane
particle) DNA base sequences appears to be present in PLC/PRF/5 cells (unpublished
ob-servations),only HBsAg,andnototherprobable
HBV gene products, e.g., HBcAg, HBeAg, or
Daneparticles, has been found in the cells.
ACKNOWLEDGMENTS
We thank Aleem Siddiqui for helpful discussions and George Scullard and Janet Kahle for assistance with the
antigen assays.
Thiswork wassupported by Public Health Service grant AI 13526 fromtheNational InstituteofAllergy and Infectious Diseases. P.L.M.istherecipient of postdoctoral fellowship5
F2CA06083fromthe NationalInstitutesofHealth. LITERATURE CITED
1. Barker, L. F., F. V.Chisari,P. P. McGrath,D.W. Dalgard, R. L. Kirschstein, J. D.Almeida,T. S. Edgington, D. G. Sharp,and M. R. Peterson. 1973.
Transmission of type B viralhepatitistochimpanzees. J. Infect. Dis.127:648-662.
2. Blumberg, B.S.,B.Larouze,W.T.London,B. Wer-ner,J. E.Hesser, I. Millman, G. Saimot, and M. Payet. 1975. The relationship ofinfection with the hepatitisBagent to primaryhepatic carcinoma. Am. J. Pathol.81:669-682.
3.Bonner, W.M.,andR.A.Laskey.1974.Afilmdetection method for tritium-labeledproteins andnucleicacids in polyacrylamidegels. Eur. J. Biochem.46:83-88. 4. Dreesman,G. R., R.Chairez, M. Suarez, F. B.
Hollin-ger,R. J.Courtney, and J. L. Melnick. 1975. Pro-duction ofantibody to individual polypeptidesderived
frompurified hepatitis B surface antigen. J. Virol. 16:
508-515.
5. Gold, J. W. N., J. W.Shih, R. H. Purcell, and J. L. Gerin.1976.Characterization ofantibodiestothe struc-turalpolypeptides of HBsAg: evidence for subtype-spe-cific determinants. J. Immunol.117:1404-1406. 6. Gudat, F., L.Bianchi,W.Sonnabend, G. Thiel, W.
Aenishaenslin, and G. A. Stalder. 1975.Pattern of
coreand surfaceexpressioninliver tissuereflects state
ofspecificimmuneresponse inhepatitis B. Lab. Invest. 32:1-9.
7. Kessler, S. W.1975.Rapid isolation of antigensfromcells with a Staphylococcal protein A-antibody adsorbent: parametersof the interaction ofantibody-antigen com-plexes with proteinA. J.Immunol.115:1617-1624.
8. Kew, M., R. Geddes, G. Macnab, andI. Bersohn. 1974. HepatitisBantigen and cirrhosis in Bantu patients with primary liver cancer. Cancer(Philadelphia) 34:539-541.
9. Laemmli, U. K. 1970. Cleavage of structural proteins duringtheassembly of the head of bacteriophage T4. Nature(London) 227:680-685.
10.Lowry,0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall. 1951. Protein measurement withthe Folin
phenol reagent. J. Biol. Chem. 193:265-275.
11. MacKay, P.,and C. J. Burrell. 1976. Examination of the polypeptides of hepatitis B surface antigen. J. Gen. Virol.33:181-191.
12. Macnab, G. M., J. J.Alexander, G. Lecatsas, E. M. Bey,and J. M. Urbanowicz.1976.Hepatitis B surface antigenproduced by a human hepatoma cell line. Br. J. Cancer 34:509-515.
13. Magnius, L. O., A. Lindholm, P. Lundin, and S. Iwar-son. 1975. A new antigen-antibody system. Clinical significanceinlong-termcarriers of hepatitis B surface antigen. J.Am.Med. Assoc. 231:356-359.
14. Mathiesen, L. R., S. M. Feinstone, D. C. Wong, P. Skinhoej, and R. H. Purcell. 1978. Enzyme-linked immunosorbent assay for detection of hepatitis A anti-gen instool andantibody to hepatitis A antigen in sera: comparison with solid-phase radioimmunoassay,
im-muneelectronmicroscopy, and immune adherence he-magglutination assay. J. Clin. Microbiol. 7:184-193. 15. Peterson,D.L., I.M.Roberts, and G. N. Vyas. 1977.
Partial amino acidsequence of two major component polypeptides ofhepatitisBsurfaceantigen. Proc. Natl. Acad.Sci. U.S.A.74:1530-1534.
16. Purcell, R. H., J. L. Gerin, and J. B. Almeida. 1973/
74.Radioimmunoassay forthedetection of the core of
the Daneparticleandantibodyto it.Intervirology 2: 231-243.
17. Rao, K. R., andG. N. Vyas.1973.HepatitisBantigen activity in protein subunits produced by sonication. Nature(London)NewBiol.241:240-241.
18. Robinson, W.S. 1977. The genome of hepatitis virus.
Annu.Rev.Microbiol.31:357-377.
19. Shih, J. W., and J. L. Gerin. 1975. Immunochemistry of hepatitisBsurfaceantigen (HBsAg): preparation and characterization of antibodies totheconstituent poly-peptides. J. Immunol.115:634-639.
20. Shih, J. W., and J. L.Gerin.1977.Proteins ofhepatitis
Bsurfaceantigen:aminoacidcompositions of the major polypeptides. J. Virol.21:1219-1222.
21. Szmuness, W. 1978.Hepatocellularcarcinoma and the hepatitis B virus: evidence for a causal association. Prog. Med.Virol.24:40-69.
22. Takahashi, K., Y. Akahane,T.Gotanda, T. Mishiro, M.Imai, Y.Miyakawa, andM.Mayumi. 1979. Dem-onstration ofhepatitisBantigenin the coreof Dane particles. J. Immunol.122:275-279.
on November 10, 2019 by guest
http://jvi.asm.org/