Copyright ©1976 American Society for Microbiology Printedin U.S.A.
Localization and Functional Role of the Pseudomonas
Bacteriophage 2 Depolymerase
FRANCISCO J. CASTILLO AND PASQUALE F. BARTELL*
Department of Microbiology, College of Medicine and Dentistry ofNew Jersey, New Jersey Medical School, Newark, New Jersey 07103
Received for publication 24 November 1975
The adsorption apparatus of phage 2 consists of a symmetrical base plate of
snowflake appearance, composed of six droplike spikes 7.0 to 7.5 nm in length
with a maximum diameter of 4.5 to 5.0 nm. The spikes are attached by their
narrow ends to a central ring 7.0 to 7.5 nm in diameter. Phage 2 depolymerase, a phage 2-induced hydrolytic enzyme, was found to be a structural protein of
phage 2 or in close association with the base plate.
Pdp,,
a phage 2 mutant,possesses apolypeptide that is antigenically similar to the depolymerase, but
devoid ofhydrolytic activity. This polypeptide was found to be located in the
region of the base plate of
pdp,.
Treatment of intact cells of strain BI withpurified phage 2 depolymerase inhibited the adsorption of phage 2. When phage
receptor-containing fractions of slime glycolipoprotein and lipopolysaccharide
were hydrolyzed by the depolymerase, amino sugars were released, and the phage-inactivating activities of these fractions were lost. The depolymerase was
also observed to induce the lysis ofstrain BI cells in hypotonic medium. The phage2depolymeraseappears toplay a role in adsorption and releaseof phage.
Bacteriophage2specificallyinfects and
repli-catesonPseudomonas aeruginosa BI.
Adsorp-tion to viable cells ofstrain BI has been
ob-served to occur on the entire cell surface in
typical tail-first arrangement (9). Phage 2 is
believed to directthesynthesis ofa
depolymer-ase (5) thatspecifically hydrolyzes the
glycoli-poprotein, previously referred to as SPB,
con-tained in the extracellular slime ofstrain BI,
reducing theviscosityand releasingaminoand neutralsugars (8).Experimental evidence
sup-ports the viewthat thedepolymerase occurs in
twodifferentstates: (i)free, and responsible for the spreading halo that surrounds phage 2 plaquesonlawns ofstrainBI, and(ii)boundto thephage2particle, asindicatedby the results
ofsedimentation in cesiumchloridegradients.
In these studies, despite exhaustive
purifica-tion of the phage particles, the depolymerase activity was found to band with phage at a
density of 1.49 to 1.51 g/ml (6). Although the
depolymerase isknown to becloselyassociated
with the structured phage particle, its exact
location has not been determined.
The possible functional role of the phage 2 depolymerase in the initial attachment of the
phagetohost cell issupportedbyexperimental
data (27). Analysis of several strains of P.
aeruginosa indicated that only those strains
containing substrate for thespecific
depolymer-asecarriedby phagewere capableofadsorbing
phage. In these studies, two phage-associated depolymerases, differing in their specificities,
were tested against various strains and mu-tants.
In the present study, a more precise
localiza-tionof thephage2depolymerase isoffered, and additional information has been assembled to
provide a keener insight into thepossible
func-tional role of this enzyme in the life cycle of phage2.
MATERIALS AND METHODS
Microorganisms. Pseudomonas phage 2, its mu-tant, pdp,, and theirhost, P. aeruginosa BI, have been previously described (5, 8). The propagation andpurification of phagewereinaccord with
proce-dures detailedinapreviouspaper(13).
Extraction and purification of slime
glycolipo-protein (GLP) and lipopolysaccharide (LPS). De-tailedprocedures concerning the preparation of GLP and LPS for use in these studies have been
pub-lished previously (7-9).
Purification ofphage 2 depolymerase. Phage 2
depolymerase wasobtained and purified according tothe method already described (4), withsome
mod-ification.After precipitation anddialysis,the
depo-lymerasewastwicesubjectedtochromatographyon
Sephadex G-200 andonce inBio-GelA-5m, eluting with 5 mM sodium phosphate buffer, pH 7.4.
Fi-nally, theenzyme wasfiltered seventimes on the Amicon ultrafiltration system through a SM-300
membrane (exclusion limit, 300,000 molecular
weight)to removeresidualphage,andconcentrated
701
on November 10, 2019 by guest
http://jvi.asm.org/
through a PM-10 membrane in the same system. Homogeneity of the purified enzyme preparation
wastestedbyimmunodiffusion techniques.Asingle
band was detected when purified or crude lysate preparations were reacted with rabbit antiserum
prepared against purified enzyme, and also when the purified enzyme was reacted with antiserum
preparedagainstwholephage particles.These crite-riawereusedtoindicate thatpurifiedenzyme prep-arations were devoid of detectable contaminating
proteinantigens.
Preparation of antisera. Each antigen
prepara-tionwas emulsifiedinan equal volumeofFreund
incomplete adjuvant. White rabbits, 4 to 5 kg in
bodyweight,weregivenweekly injectionswith4ml ofantigen emulsion subcutaneouslyintheneck
re-gionbehind theears, for 2to4consecutive weeks. One weekafter thefinal injection, theanimalswere test-bled fromthe marginal veinsof the ears, and finallybledbycardiacpuncture.The phage antigen consisted ofpurified phageat2 x 10"° PFU/mlin 5 mMTrisbuffer, pH7.4. Serum neutralizationrate constants(K values)werecalculatedasdescribedby
Adams (1). Thepurifiedenzyme solutions usedfor
antigenicstimulation contained 85,ugofproteinper
mlin 5mMsodium phosphate buffer, pH7.4.
Immunodiffusion. Immunodiffusion was per-formed on glass slides and petri dishes overlayed with1%Nobleagarinbarbitalbuffer,pH8.6(14).
Determinationofenzymeactivity.Hydrolytic ac-tivityofthe phage depolymerase involvedreaction mixturescontaining0.25 mlof slime (2.44mg/ml)or
0.25 mlofLPS (2.18 mg/ml) in 0.2 M sodium phos-phate buffer(pH 7.4)and0.25ml ofenzymesolution (0.4 mg of protein/ml) in the same buffer. After
incubation for different time intervals, the tubes were immersed in ice water to stop the reaction. Release of amino sugars was determined by the
method of Belcheretal. (10).
Phage adsorption. Log-phasecultures were
sedi-mented at6,000 x g for 10 min and suspended in
freshTrypticasesoybroth (TSB)ataconcentration of107cells/ml. Thecellularsuspensionsweremixed
withanequalvolumeofphagecontaining 105PFU/
mlat37C. Aliquotswere removedatvarious time
intervals, diluted1:10incoldTSB,andimmediately centrifugedat6,000 xgfor 10 mintosediment cells and adsorbed phage particles. The supernatant fluidsweredilutedandplatedonwild-typestrain BI to determine the number of unadsorbed or free
phageparticles.Anadsorptionmediumcomposed of
5mMTris buffer(pH 7.4),supplemented with0.1M sodium chloride and 0.01 Mmagnesiumsulfate,was
alsoused inplaceof TSB (27).
Phage inactivation. Phage 2 suspensions (105
PFU/ml) were mixed with the substances to be tested and incubatedat37C; after the required time
intervals, the mixtures werediluted in cold buffer
andtitratedtodeterminethenumber ofremaining infectivephageparticles.
Effectof depolymerase onphage 2 adsorptionto
wholeBIcells.Strain BI in the log phase of growth
was centrifuged at6,000 xgfor 10minat 4Cand
resuspended inTSBto aconcentration of 108cells/
ml. The reaction mixtures contained 1 ml of cells
and0.15 mlof enzymesolution (70tkgofprotein/ml) in 0.2Msodiumphosphatebuffer (pH 7.4). Controls were eitherdevoid of cellsorenzyme. The enzyme wasadded20 minbefore phagein onecase,andat
the sametime as phage inthe other. After phage addition(105PFU/ml), themixtures wereincubated 10 min at 37 C, and residual phage was titered as
described above.
Effect ofdepolymerase on phage 2 inactivation byGLP and LPS. GLP and LPS solutions (200 ,gI ml)in5mMTrisbuffer (pH 7.4),supplemented with
0.1 Msodium chloride and0.01 Mmagnesium sul-fate,werereacted for60minwithequal volumes of enzyme (70 ,ug ofprotein/ml) in the same buffer. Controls contained buffer instead of enzyme
solu-tion.Phage2(105PFU)wasaddedtoeachtubeand, after incubation for 15min,sampleswereremoved, andphagetiters weredetermined.
Effect ofdepolymerase onintactbacterial cells. Four-hour log-phase cultures ofP. aeruginosa BI
were centrifuged at 3,000 x g for 10 min, washed
twice in 5 mM sodium phosphate buffer (pH 7.4), andresuspendedtohalf the original volumeinthe
samebuffer. For studies oflysis, 1-ml portions of the suspension ofwashed cellswere dilutedin 6 ml of sodium phosphate buffer and exposed to the lytic agents singly and in various combinations at the following concentrations:phage2depolymerase,300
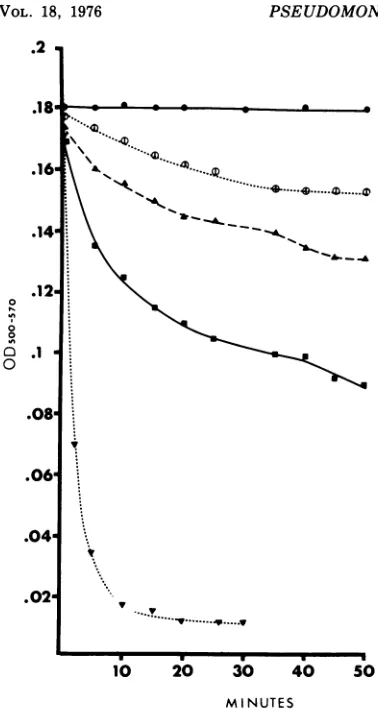
jtgof protein per ml; egg white lysozyme, 350,ug/ml; EDTA, 320 /Lg/ml. Controls contained 1 ml of cell suspension in 6 ml ofphosphate buffer. Lysis at roomtemperaturewasfollowed by measurement of optical density at 500 to 570 nmwith a Klett-Sum-mersoncolorimeter.
Electron microscopy. Drops ofspecimen
suspen-sions wereplacedongrids with carbon-coated Form-varfilm for1 min, and excess material was blotted off. The samples were examined, after negative stainingwith 2%sodiumphosphotungstate (pH 7.6),
inanHitachiHU-12electronmicroscope at 75 kV.
RESULTS
Structure of phage2tailtip. To gain
infor-mation onthe structures) involvedin the
ad-sorption process, astudy of the morphology of the phage particle, especially the tail tip, was undertaken.
The gross morphological characteristics of
phage 2 have been described (9). This phage belongstoBradley'sgroup B (12),andpossesses
ahead withhexagonal outlineandalong,
non-contractile tail. A close examination of the tail
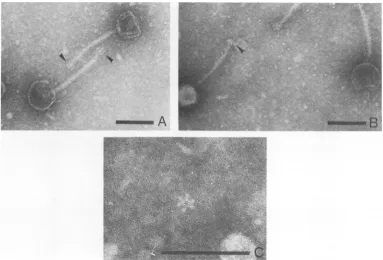
structure revealed finger-like projections or spikesatthe distalend,asseeninFig. 1A,that
measured between 13 and 16nm in length and
5.5 to 8 nm in diameter, or shorter spikes as
seen inFig. 1B. Phage 2base plates were
ob-served to have a snowflake appearance with
radial symmetry, consisting of sixdrop-shaped
spikes7.0 to 7.5 nminlengthwithamaximum
diameter of4.5 to 5.0nm,withthe narrow ends
oriented toward the center and attached to a
centralring7.0to7.5 nm indiameter (Fig. 1C).
on November 10, 2019 by guest
http://jvi.asm.org/
PSEUDOMONAS BACTERIOPHAGE 2 DEPOLYMERASE '703 Localization ofthe phage-associated
depo-lymerase. Evidencehas beenpresented
indicat-ing a close structural association of the
depo-lymerase with the phage 2particle(6), and the
likelihood that the depolymerase functions in the initial attachment ofphage (27). It
there-fore becameofgreatimportancetodeterminea
more precise location of the enzyme on the
phage particle.
If the phage-associated depolymerase is
lo-cated on a region of the virus involved in the
adsorptive process, theninteraction of
anti-de-polymerase serum with the phage particles
should render them incapable of initiating
in-fection, and therefore inactive. Furthermore,
underappropriateconditions,theinteractionof
phage 2 with anti-depolymerase serum might be easily visualized under the electron
micro-scope, thus providing additional information concerning the exact location of the enzyme.
Table 1 shows the results of the first
experi-ment. GLP and LPS, known to contain phage
receptors, were included in the experimentas
positive controls since they readily inactivate phage2.When phage 2 wasmixedwithpurified
GLP orLPS fromstrain BI, 85 and 97%
inacti-vation occurred, respectively, after 10 min at
37 C. When the viruswasreactedwith
antise-rum
prepared
againstpurified
phage
particles,
100%inactivation tookplace. Likewise,
antise-rum prepared against purified
depolymerase
brought
aboutcomplete
inactivation of thephage, indicating that depolymerase is indeed
associated with the virus
particle,
and essentialto its
infectivity.
When
phage
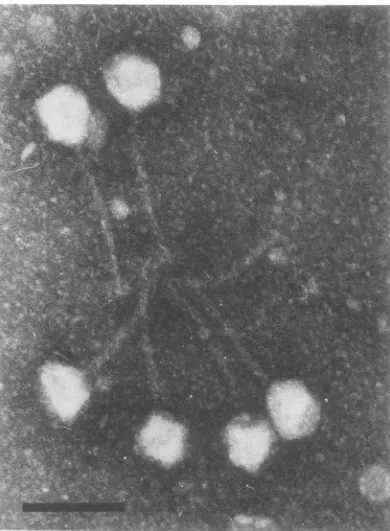
2 wasinteracted with theanti-depolymerase serum, electron
micrographs
showed thatthe virusparticleswerebound
to-gether in a rosette arrangement, with the
phage
tailsorientedtowardthecenter(Fig.2).
In contrast, when phage 2 was reacted with
antiserum prepared against purified phage 2
particles, agglutination
ofthe virus occurred ina completely disorganized pattern. These
re-sultsindicatethat the polysaccharide
depolym-eraseis located atthetipofthe tail ofphage
2,
close to, orformingpartofthe baseplate
struc-ture.
Pdp,,
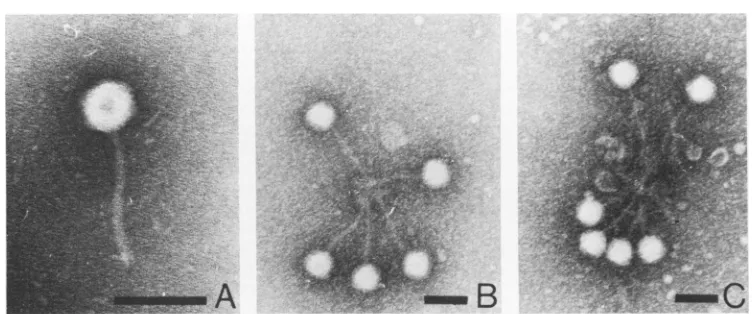
aphage2depolymerase mutant. Pdp1isahalo-lessmutantofphage2(Fig. 3A)that is
abletoinfect andreplicateonP.aeruginosa
BI;
however, depolymerase activity has not been
observedtobeassociated with this mutant (5). Thegrowth curveof
pdp,
wasfound to besimi-FIG. 1. Pseudomonasphage2.In(A.) thearrowspointtofinger-like projectionsorspikesatthetailtip,and
in(B) thearrowindicates the shorter spikes. Thephage2 baseplateisshownincenterof (C).Notethesix
drop-shaped spikesoriented towardthecenter.Barrepresents 100nm.
VOL. 18,
.11 -,.
V ...
low
IS.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.59.442.358.618.2]TABLE 1. Inactivation ofphage 2 by anti-phage 2 andanti-depolymerase sera
Phage in- Phage re- % Inac-Medium put(PFU/ covered
tiva-ml) (PFU/ml) tion
Buffer 1.3 x 104 1.3 x 104 0
TSB 1.3 x 104 1.3 x 104 0
GLPI 1.3 x 104 1.9x 103 85
LPSr 1.3 x 104 3.8 x 102 97
Anti-phage 2se- 1.3 x 104 0 100
rum
Anti-depolym- 1.3 x 104 0 100
erase serum
aTris, 5 mM, pH 7.4, supplemented with 0.1 M
sodium chloride and 0.01 M magnesium sulfate. bPurified from P. aeruginosa BI (100
jig/ml).
e Purified from P. aeruginosa BI (100 ,ug/ml).lar to thatof phage 2, having a latent period of
55 min, a rise period of 20 to 25 min, and an
average burst size of 10. The absence of halo
around the plaquesof this phage might be
in-terpretedintwoways: (i) absenceof
depolymer-ase or(ii)thestructural presence of
depolymer-asethat lacks hydrolyticactivity. Experiments
were performed toexamine these possibilities,
andthe results arepresented inTable2.
Anti-serum prepared against purified phage 2 was
found tocompletelyinactivate
pdp,,
indicatingastrongserologicalrelatedness between phage
2and its mutant. Antiserum prepared against
thepurifieddepolymerasesimilarly inactivated
the mutant. This was taken as an indication
that a cross-reacting polypeptide(s),
antigeni-cally similarto the depolymerase, was present
in
pdp,
particles. Further examination underthe electron microscope revealed that
pdp,
in-teractedwith the anti-depolymerase, producing
a tail-centered rosette arrangement (Fig. 3 B,
C) similartothat observed when thewild-type phage 2 was reacted with anti-depolymerase
serum (Fig. 2). These results suggest that the
mutant,
pdpl,
possesses tail regionpolypep-tide(s) thatisantigenically relatedtothe
wild-type phage 2 depolymerase polypeptide(s).
Al-though enzymatically inactive, this
polypep-tide(s) appears to be in the same structural
location as thedepolymerase of phage 2.
It is of interest to note that GLP and LPS,
although able to readily inactivate phage 2,
showed noability toinactivate
pdp,.
Effectof purified phage 2 depolymerase on
cellularphagereceptors. Since the possibility
thatphage 2depolymerase may play a role in
theadsorptive process has been considered (27),
theeffect of enzyme treatment on the ability of
whole cells of strain BI to adsorb phage 2 was
examined. It was observed that whereas 69%
adsorption of phage 2to BIcells occurred after
10min ofincubation, only51% adsorption took
place when enzyme wasadded simultaneously
with the phage. Furthermore, when the cells
weretreated with theenzymefor 20min before
phageaddition,noadsorption of phage occurred
(Table 3).
GLP and LPS from strain BI have been
shown to contain receptors forphage2(9), and
it wasof interest to study the effect of the phage
2 depolymerase on GLP and LPS. It Was
ob-served thatpurified depolymerase had the
abil-ity to hydrolyze not only slime, as previously
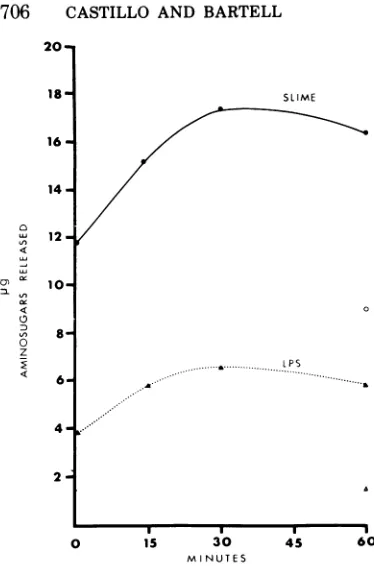
reported (8), but also LPS extracted from strain BI, inducing the release of amino sugars (Fig. 4). The effect of enzyme treatment on the
abil-ityof GLP and LPS to inactivate phage 2 was
then determined. As shown in Table 4, when
GLP and LPS were treated with the enzyme
before addition of phage, they completely lost phage 2 inactivating activity. These
experi-ments suggestthat phage 2 receptors are either
modified and/or destroyed by the action of the
enzyme.
Depolymerase treatment of strain BI cells, suspended in a hypotonic medium, resulted in a
FIG. 2. Bacteriophage 2 after interaction with
anti-depolymeraseserum.Note the rosette formed by
thephage with their tails oriented toward the center. Bar represents100 nm.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.266.461.346.612.2]PSEUDOMONAS BACTERIOPHAGE 2 DEPOLYMERASE
A
FIG. 3. Phage 2 halo-less mutant pdp (A) after interaction with anti-depolymerase serum(B, C). Note rosette arrangement similar to that observed withwild-type phage 2. Bar represents 100nm.
TABLE 2. Inactivationofphagemutantpdplby
anti-phage2and anti-depolymerase sera
Phage input Phagere- %
Inac-Medium (PFU/ml) covered tiva-(PFU/ml) (PFU/ml) tion
Buffer 1.95 x 105 1.95 x 105 0
GLP` 1.95 x 105 2 x 105 0
LPSr 1.95 x 105 2.08 x 105 0
Anti-phage2 1.95 x 105 0 100
serum
Anti-depo- 1.95 x 105 0 100
lymerase
serum
aTris, 5 mM, pH 7.4, supplemented with 0.1 M
sodium chlorideand 0.01 Mmagnesiumsulfate.
bPurified fromP. aeruginosaBI (100
/xg/ml).
c Purified from P. aeruginosa BI (100
,.g/ml).
TABLE 3. Effect of depolymeraseonadsorptionof
phage2toviable cellsofP.aeruginosaBI
Phage Phagere- % Ad-Medium added covered
(PFU/ml) (PFU/ml) sorption
TSB +buffer" 2.56x 105 2.56 x 105 0
TSB + depolymerase 2.56x 105 2.60x 105 0 Cells" +buffer 2.56x 105 0.80x 105 69
Cells + depolymerase 2.56 x 105 1.25x 105 51
(at zerotime)
Cells +depolymerase 2.56 x 105 2.56x 105 0
(at-20min)
aTris, 5 mM, pH 7.4,supplemented with0.1 Msodium
chloride and0.01Mmagnesiumsulfate.
bP.aeruginosa BI(10"cells/ml).
reduction ofturbidity, indicatingcellularlysis.
Similarlytic activitywasobservedinthe
pres-enceofEDTA, lysozyme,orboth(Fig. 5).
Thus,
the phage 2 receptors appear to be intimately
related to essential structural components of
the cell wall, and thehydrolytic activityof the
phage 2depolymerase is capable of disrupting
thisintegrityof the cell envelope.
DISCUSSION
Phage 2 has been previously described as
possessing a knoblike structure at the distal
end of itstail, and observationshavealso been
made of tail spikes in the region ofinteraction
of the tail tips with slime GLP from strain BI
(9). Inthe present work, it was possible not only
tocorroborate the presence of these spikes, but
also to provide information on the structural
detail of the phage 2 base plate.
It is of interest to note that the phage spikes
were observed in forms that suggest the
possi-bility of two states: extended and retracted.
The physiological implication of this
dimor-phism is not understood. However, it may
explain the earlierreportsofaknoblike
struc-ture at the tail tip ofphage 2. On the other
hand, the differences observed might also be
accounted for by the fact that the specimens
were viewed in different planes, resulting in
different two-dimensional projections of the
same type of structure. Isolated base plates
were observed as a symmetrical snowflake
structure. Although in the present study only one structural stateofthe isolated base plates
was observed, the possibility of alternative
states is not disregarded. Electron microscope
examinations of othersystems, such as T-even
coliphages, have suggested that basal plates
mayexistintwo states (2, 17, 20, 25, 26, 28). In
the intact T-evenphage particles with extended
tail sheath, the base plates occur as fine hexag-onalstructureswitha maximumdiameter of31
to 40 nm; with contraction of the sheath, the
appearance of the basal plates changes from
hexagons to six-pointed stars with a maximal
diameter of50 to 60 nm.
705
VOL. 18,
ZSC;_.
;M.l
B
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.64.441.63.220.2] [image:5.503.54.246.278.388.2]706 CASTILLO AND BARTELL
20-J
LU Q
0 z
Is.
16'
14-12.
10'
4-SLIME
lPS
0 i5 30 45
M NUTES
FIG. 4. Release of aminosugarsfrom I
nosaBIslimeGLP (0.61 mg) (@) and LPS(
(A) duringtreatmentwithphage2depolym
mg).ControlsofslimeGLP(0)aridLPS (L
additionofdepolymerase arealsoshown.
The fact that only the tail tips of
particles interacted with antiserum I
against purified depolymerase, with
comitantloss ofphage infectivity,sugg
thisenzyme islocatedinclose associat
or is one of the structural component
base plate. This observation indicat
again, thepossible importance of theer
the infectious process. Localization o
associated enzymes at the tail tip
phages is welldocumentedinthe litera
22, 23, 29-31). It thus appears very
that the phage 2-depolymerase exist,
states: free, and boundtothephagepa
one of the structural proteins of thebk
(6, 8). The relatedness of the free an
depolymerasewasdetermined by their
cal cross-reactivity, since antiserum I
against the purified free enzyme reach
thephage-associatedenzyme.
Ifthe depolymerase activity plays
the attachment ofthe wild-typephag
cells, thenpdp,,amutantdevoid of de
aseactivity, shouldbe incapable ofac
orshowalowerrateof adsorption, as
by longer latent and rise periods than
phage2. However,thesamevalues,55minfor
thelatentperiods and20 to 25min for therise
periods,wereobservedwithbothphages.These
resultssuggestthatactiveenzymeisnot
essen-tialintheadsorptionprocess.Nevertheless,the
presence of structural enzyme in an inactive
form might still be involved inthe adsorptive
process. This hypothesiswassupported by the
factthatthe mutantphage,
pdp,,
althoughde-void of depolymerase activity, reacted with
anti-depolymerase serumtoproducea
tail-cen-tered rosette arrangement similar to that
ob-served with the wild-type phage 2. These
re-0 sultsclearly indicatedthepresenceofwild-type
depolymerase cross-reacting polypeptides in
the tail tip of
pdp,,
which, although devoid ofenzymatic activity, might retain the abilityto
recognize complementary receptors and allow
phageto attachtothecell surface.
In this connection, it should be mentioned
that coliphage T4D possesses an enzyme,
di-hydrofolate reductase, as oneof the structural
components of the base plate, and that this
enzyme has been shown to play a role in the
, adsorption process. Two mutants ofT4D,
nei-60 ther of which induces the production of active
dihydrofolate reductase, are fully infective.
P. aerugi- Thesemutants induce the productionofa pro-(0.55 mg) tein that resembles theenzymeinthat it binds
Erase(0.1 tothesubstrate, but doesnotreducefree
dihy-\)without drofolate. Kozloffetal. indicated that the
pro-tein may serve a structural rather than an
enzymatic roleinthephage tail plate (24). This
phage 2 hypothesis has also been proposed by Dawes
prepared
and Goldberg(18),whostated that thedihydro-the con- folatereductase plays noenzymatic roleinthe
,ests
that virion,but isstructurallyimportantduring theionwith earlystagesof phage adsorption. A similar
ex-ts of the planation may apply to the role of phage 2
;es, once depolymerase in the adsorption process,
enzymein TABLE 4. Effect of depolymeraseonphage2
f phage- inactivation bypurified GLP andLPS from P.
of other aeruginosaBI
Lture(11,
certain
s in two
articleas
aseplate
id bound
serologi-prepared
ted with
a role in
e 2to BI
polymer-lsorption reflected
those of
Phage input Phage re- % Inacti-Medium (PFU/ml) covered vation
Medium
~~~(PFU/ml)
Buffer
6.75 x 105 6.75 x 105 0Buffer + depo- 6.75 x 105 6.80 x 105 0
lymerase
GLP'+buffer 6.75 x 105 1.24 x 105 82
GLP + depo- 6.75 x 105 6.8 x 105 0
lymerase
LPSC
+buffer 6.57 x 105 1.75 x 103 99.8LPS+depolym- 6.75 x 105 6.9 x 105 0 erase
aTris, 5 mM, pH 7.4, supplemented with 0.1 M
sodiumchloride and0.01Mmagnesium sulfate.
bPurified fromP.aeruginosa BI.
ePurified fromP. aeruginosa
BI.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.60.247.52.335.2]PSEUDOMONAS BACTERIOPHAGE 2 DEPOLYMERASE .2
01
0
08
0
.0
.06
.04
.02
.04..~~~~...
10 20 30 40 50
MINUTES
FIG. 5. Changes in optical density ofP.
aerugi-nosa BI suspensions during treatment with 5 mM sodiumphosphate buffer, pH7.4(0),depolymerase
(300 pg/ml) (0D), lysozyme (350
Mg/ml)
(A),EDTA (320 pg/mi) (U),andEDTA (320 pg/ml)pluslyso-zyme (350 pg/ml) (V).
thereby clarifying the fact that pdp, is fully
infective. Alternatively, the phage 2
depolym-erase may possess a bifunctional activity,
in-volving theabilitytohydrolyze the slime GLP
and LPS and to recognize and attach to the
phage receptors presentinslime GLP and LPS.
Itisof interest to notethat whereaswild-type
phage 2 is irreversibly inactivated by slime
GLP and LPS, the mutant phage
pdp,
is notinactivated bythese cellular components. It is
possiblethat the mutantdepolymerasecarried
by
pdp,
has more stringent requirements forrecognition of receptors inpurified slime GLP
andLPS, orthatpurification of GLP and LPS
induces modifications in thetertiary structure
of these substances which limit or modify the
availability of the receptors. Another
possibil-ity is that secondary receptors may be
neces-saryfor the inactivation of
pdp,,
and these areeithermissing or hidden inthe purified slime
and LPS. Alternatively, the free and bound
enzymes, although serologically similar, may
have a differentmechanism of interaction with
the phage receptors.
Purified phage 2 depolymerase was found to hydrolyze not only slime GLP from strain BI, as previously reported (8), but also LPS from the
same strain. This hydrolytic activity resulted
in the release of amino sugars and destroyed
the phage-inactivating activity of GLP and LPS. Furthermore, cells of strain BI treated
with the enzyme lost the ability to adsorb
phage 2. These results indicate that the
sub-stratefor the enzyme is present in LPS as well
as in GLP. This is of great significance since
these two cell fractions possess the functional
receptors for phage 2. The enzyme is able to
destroy the receptor activityof GLP and LPS,
notonly in vitro, but also in situ.Thus, the free
depolymerase produced in excess during the
process ofphage replication may protectnewly
formedprogenyfrominactivation by GLP and LPS thatis present onthe surfaceof the cell.
Gram-negative bacteria containseveral
sur-face layers of differentcomposition and
physi-cal nature (15). Each of these layerspresentsa
mechanical barrierthat might impedethefree
release of mature phages from infected cells.
Phage release maybeaccomplishedby the
pro-duction of specific enzymes that disrupt the
structuralintegrity of these layers. The pepti-doglycan layer is usually considered to be re-sponsible for maintaining the
rigidity
ofthe cellin the majority of gram-negative bacteria.
However, in P. aeruginosa the situationseems
somewhat different.
Although
some lesionswere observed aftertreatmentofcells with
ly-sozyme, the cellwallstructureremained intact
(19). This indicates that peptidoglycan is not
solely responsible for the
rigidity
of the cell.Furthermore, if the outer membrane is
dis-rupted bytreatmentwith Trisbuffer, EDTA,or
acombination of
both,
cell lysis ensues (3, 16,19, 21, 33). As reported in the present
work,
treatment ofBIcells with
phage
2depolymer-ase in hypotonic media induced cell
lysis.
Un-der natural conditions, depolymerase activity
may facilitate the release of
phage
particles
by
eliminating a portion of the physical barriers
represented bythe structural layers ofthe cell
envelope. Inaddition, progeny
phage
2maybeprotectedfromthe receptors presentinLPSand
GLP which are
hydrolyzed
orpossibly
blockedby thedepolymerase.
Thus,
thephage
2depo-lymerase appearsto play arole intwo
impor-tantstepsof thephagelife
cycle:
adsorption
andrelease.
707
VOL. 18,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.52.241.49.409.2]ACKNOWLEDGMENTS
This studywassupported by Public Health Servicegrant
AI-08504from the National Institute of Allergy and Infec-tiousDiseases.F. J.Castillowassupported byapredoctoral
scholarship from the Instituto Venezolano de
Investiga-cionesCientificas.
LITERATURE CITED
1. Adams, M. H. 1959.Bacteriophages. Interscience
Pub-lishers, Inc., NewYork.
2. Anderson,T. F.,and R.Stephens.1964.Decomposition ofT6 bacteriophage inalkaline solutions. Virology
23:113-117.
3. Asbell,M. A.,andR.G. Eagon.1966.Role of multiva-lent cationsin the organization, structure, and
as-sembly of the cell wall ofPseudomonas aeruginosa.J.
Bacteriol.92:380-387.
4. Bartell, P. F., G. K. Lam, andT. E.Orr.1968. Purifica-tionand properties ofpolysaccharide depolymerase
associated withphage-infectedPseudomonas
aerugi-nosa.J.Biol. Chem. 243:2077-2080.
5. Bartell, P. F., and T. E. Orr. 1969. Origin of
polysac-charide depolymerase associatedwithbacteriophage infection.J.Virol.3:290-296.
6. Bartell, P. F., and T. E. Orr. 1969. Distinct slime polysaccharide depolymerase of bacteriophage-in-fectedPseudomonasaeruginosa:evidence of close
as-sociationwiththestructured bacteriophage particle. J.Virol. 4:580-584.
7. Bartell,P.F.,T.E.Orr, and B.Chudio.1970.
Purifica-tionand chemicalcomposition of the protectiveslime
antigen ofPseudomonas aeruginosa. Infect. Immun.
2:543-548.
8. Bartell, P. F., T. E. Orr, and G. K. H. Lam. 1966.
Polysaccharide depolymerase associated with bacte-riophage infection. J. Bacteriol.92:56-62.
9. Bartell,P.F.,T. E.Orr, J.F.Reese,andT.Imaeda.
1971. Interaction ofpseudomonas bacteriophage 2
with the slimepolysaccharide and lipopolysaccharide
ofPseudomonasaeruginosastrain BI. J.Virol. 8:311-317.
10. Belcher, R.,A.J. Nutten, and C. M.Sambrook.1954.
The determination of glucosamine. Analyst 79:201-208.
11. Bessler, W., E. Freund-Molbert, H. Knufermann, C. Rudolph, H. Thurow, and S. Stirm.1973. A bacterio-phage-induced depolymeraseactiveonKlebsiellaKll
capsularpolysaccharide. Virology56:134-151. 12. Bradley, D.1967.Ultrastructureof bacteriophages and
bacteriocins. Bacteriol. Rev. 31:230-314.
13. Castillo, F. J.,and P. F. Bartell. 1974. Studiesonthe
bacteriophage 2 receptors ofPseudomonas
aerugi-nosa.J.Virol. 14:904-909.
14. Chase, M. W.1968. Buffers,p.365-408.In Methodsin
immunology and immunochemistry, vol. II. Aca-demicPressInc., New York.
15. Costerton,J. W., J. M. Ingram, and K. J. Cheng. 1974.
Structureand functionof the cell envelope of
gram-negativebacteria.Bacteriol. Rev. 38:87-110. 16. Cox,S.T., and R. G. Eagon. 1968. Actions of
ethylene-diaminetatra-acetic acid, tris
(hydroxymenthyl)-ami-nomethane,and lysozymeoncell walls of Pseudomo-nasaeruginosa. Can. J. Microbiol.14:913-922.
17. Cummings,D.J.,V. A.Chapman, S.S.DeLong,A. R.
Kusy,and K. R. Stone. 1970. Characterization of T-evenbacteriophage substructures II. Tailplates. J.
Virol.6:545-555.
18. Dawes, J., and E. B.Goldberg. 1973. Functions of base-platecomponentsinbacteriophageT4infection.
Vi-rology55:380-390.
19. Eagon, R. G., and K. J. Carson. 1965.Lysis of cell walls and intact cells ofPseudomonasaeruginosaby ethyl-enediamine tetraacetic acid andby lysozyme. Can.J.
Microbiol. 11:193-201.
20. Fernandez-Moran, H. 1962. New approaches in the
study of biological ultrastructurebyhigh-resolution electron microscopy, p. 411-427. In R. J. C. Harris (ed.),Symposia of theInternational Society for Cell Biology, vol. 1.Theinterpretationofultrastructure.
Academic Press Inc., New York.
21. Gray, G. W., and S. G. Wilkinson.1965. The effect of ethylenediaminetetraacetic acidonthe cell walls of some gram-negative bacteria. J. Gen. Microbiol. 39:385-399.
22. Iwashita, S., and S. Kanegasaki. 1973.Smoothspecific phage adsorption. Endorhammosidase activity of tail parts ofphage P22. Biochem. Biophys. Res. Com-mun. 55:403-409.
23. Kanegasaki, S., and A. Wright. 1973. Studies of the mechanismof phage adsorption: interaction between phageE'5and itscellular receptor.Virology 52:160-173.
24. Kozloff, L.M., C. Verses,M.Lute,and L. K.Crosby. 1970. Bacteriophage tail components. II. Dihydrofol-atereductaseinT4Dbacteriophage.J. Virol.
5:740-753.
25. Poglazov, B.F. 1973.Morphogenesis of T-even bacteri-ophages.Monographsindevelopmental biology,vol.
7.S. Karger, Basel.
26. Poglazov, B.G.,L.P.Rodikova,andR. A.Sultanova. 1972.Isolation and characterization ofbacteriophage T4 baseplates.J.Virol.10:810-815.
27. Reese, J. F., G. Dimitracopoulos, and P. F. Bartell.
1974. Factorsinfluencing the adsorption of bacterio-phage2tocells of Pseudomonas aeruginosa. J. Virol. 13:22-27.
28. Simon, L. V., andT. F.Anderson.1967. The infection of Escherichiacoli by T2 and T4 bacteriophagesas seen in the electron microscope. II. Structure and functionof thebaseplate. Virology 32:298-305. 29. Stirm, S., W. Bessler, F. Fehmel,E.Freund-Molbert,
and H. Thurow.1971.Isolationofspike-formed parti-cles frombacteriophage lysates. Virology45:303-308.
30. Stirm, S., W. Bessler,F.Fehmel, andE.
Freund-Mol-bert.1971.Bacteriophageparticles with endo-glycosi-dase activity.J.Virol.8:343-346.
31. Stirm, S.,W.Bessler, F. Fehmel, E.Freund-Molbert, and H. Thurow. 1974. Uber Eine Bakeriophagen-Induzierte Colansaure-Depolymerase. Zentralbl. Bakteriol. Parasitenkd. Infektionskr. Hyg. I. Abt. Orig.A.226:26-35.
32. Takeda, K.,and H. Uetake. 1973. In vitro interaction between phage and receptor lipopolysaccharide: a novel glycosidase associated withsalmonellaphage E15.Virology 52:148-159.
33. Voss,J. G.1964.Lysozyme lysisofgram-negative
bac-teriawithoutproductionofspheroplasts.J. Gen.
Mi-crobiol. 35:313-317.