JOURNAL OFVIROLOGY, Mar.1978, p. 811-823 0022-538X/78/0025-0811$02.00/0
Copyright X 1978 AmericanSocietyforMicrobiology
Vol.25; No. 3
PrintedinU.S.A.
Groups of Adenovirus Type 2 mRNA's Derived from
a
Large
Primary
Transcript: Probable Nuclear
Origin
and
Possible
Common
3' Ends
J. R.NEVINSANDJ.E. DARNELL*
TheRockefeUerUniversity,NewYork, New York 10021
Received forpublication 27 September 1977
Late in adenovirus type 2infection, a number of mRNA's apparently arise by
processing
alarge
nucleartranscript
thatrepresents theright-hand
85% of thegenome
(smzed
in Evans et al., Cell 12:733-739, 1977). Hybridization of labeled late mRNA to a series of DNA restriction fragments representing the right-hand 70% of the genome demonstrates atleast 12 discrete mRNA's that appeartofall into five groups, eachpossibly
containing acommon3'terminus. The processing necessaryto generate these mRNA's apparentlyoccurs in thenucleus.
All the mRNA's appear to contain a sequence of -100nucleotides
complementary
to afragment
with coordinates 25.5-27.9. Thisfragment
containsone of the
regions
foundby Berget
et al.(Proc.
Natl. Acad. Sci. U.S.A. 74:3171-3175, 1977), ChowetaL(Cell
12:1-8, 1977), andKlessig (Cell
12:9-22,1977)
tobe"spliced"
ontothe 5' end of late adenovirustype 2 mRNA's.Becausethe sequencestobe
spliced
existonly
onceperlarge transcript,
any of themRNA-specific regions might only
bepreserved
fromasmall fraction of thetranscripts.Measurement of the
tansport efficiency
ofregions
ofthelarge
nucleartranscript,
in
fact,
shows thatonly
15 to25%ofanyparticular region
istransported
tothecytoplasm.
The overall conclusion of theseexperiments
is that thelarge late
nuclear transcript
canbeprocessed
in the nucleustoyield
anyoneof many (-12)mRNA's;
theunused
portions
of theprimary
tanscript
then accumulate in thenucleus
or aredestroyed.
Information about the
synthesis
of virus-spe-cific mRNA inadenovirus-infected cells haspro-gressed
veryrapidly
sincerestrictionendonucle-ases became availabletodivide the virus DNA
intoa
reproducible,
orderedsetoffragments
(29,31). The
approximate
sitesonthegenome thatcorrespond
tospecific early
and late mRNA's(32, 34), the determination of which strand of the DNA is
transcribed,
and the direction oftranscription
ofearly
andlate mRNA'swerealldetermined
by hybridization
of unlabeled RNAto
fragmented
labeled DNA(for
areview,
seereference
9).
Morerecently
(6, 26), adetailed
mapof
mRNA
sites inspecific regions
of ade-novirus type 2(Ad2)
DNA was obtainedby
observing
"Rloop"
formation-Rloops
(40) areRNA-DNA
hybrids
in which onestrand of the DNA duplex is displaced by the mRNAmole-cule-ini
specific
regionsofthegenome.In
paralel
with themapping ofmRNAsiteson the Ad2 genome, biochemical studies of mRNA formation revealed thatAd2mRNA,in common with
celular
rmRNA,
requires
a num-ber ofnuclearpost-transcriptional
modifications of the mRNA. These include the addition ofpolyadenylic
acid[poly(A)]
to the 3' terminus(32) and
specific methylations
(28, 37);internal
adenylate
residues aremethylated
togivem6A
approximately
onceevery200A's, in addition tothe formation ofa5'-terminal methylated struc-ture,
termed
a"cap"
(35). Theprimary
RNA transcripts that undergo this nuclear processing are notsimply
the samelength
as the mRNA itself. Forexample,
late in infection a single largetransciptional
unit from -16 to 100 on therightward-reading
strand iscopied
into apri-mary
trnscript
-28,000 nucleotides long (theconvention
[9]
forthe description
of the Ad2genome divides into 100 units
the
double-stranded
DNA,
each -350nucleotide
pairs or0.35 kilobase
[kb] long.
Therightward
tran-scribedrstrand has0asitsleft end and100at itsright,
with thereverse true forthe leftward strand 1. The coordinates ofparticular regions of the genome for either restrictionfagments
or mRNA coordinatesisgiven by two numbers, the percentages from lefttoright;
forexample,
the 36.7-40.5fragment
orthe 50-62 mRNA) (2, 8, 14, 43). Over 95% of the synthesis of RNA fromthe
regions
ofthe genomeencoding
the major811
on November 10, 2019 by guest
http://jvi.asm.org/
812
late proteins (8, 9, 21), themajorcore, penton, hexon, "1OOK," and fiber polypeptides,occurs as part of thislong RNA molecule. Moreover, la-beling of mRNA that originates in this region is equally sensitive to UV irradiation as the for-mation of the large nuclear transcript itself, thereby identifying the large transcript as the obligatoryprecursor to the mRNA lying within its boundaries (13, 14). Thus, the processing procedureappears toincludesite-specific cleav-age of alarge transcript as well as additionof capandpoly(A) tothe mRNA.
In studying the origin of the 5' ends of late mRNA'sinthe 50-100region, Berget etal. (4), Chowet al. (5), Gelinas and Roberts (12), and Klessig (19) have discovered a mostsurprising fact. The 5'-terminal 150-200 nucleotides ofa number of the late Ad2 mRNA's do notcome fromregions of the Ad2 genome contiguousto the remainder of the mRNA but from three distant sites. Becausethe
large
nuclear Ad2 pri-marytranscript appearsto be the mRNA pre-cursor for mRNA molecules from thisregion,
they conclude that 50 to 100 nucleotides from each of three
sites,
16.7,19.7,
and26.0,
arecleaved out
separately
andligated
onto the 5' ends of eachmRNA molecule derivedfromthe large transcript. This astounding development haspromptedus tomap asaccuratelyaspossible the number and relative amounts of various mRNAmoleculesderiving
fromthelarge
tran-script andtodetermine whether the formation ofall of the relevant mRNA's appears tobe anuclear event. To
satisfy
these goals, mRNA mapping by hybridization of labeled RNA is necessarybecause ofthedifficulty
ofobtaining
quantitative
andkinetic dataon mRNAdistri-bution
by using
electronmicroscopic
techniques.
Our conclusions are that at least 12 different mRNA'scanbe derivedfromnuclear
processing
of the16to100
transcript.
TheseRNAsfall into groups where two to four mRNA's may share the same 3' terminus. The mappositions
as-signed by
hybridizing
labeled molecules agreewell with the electron
microscopic
mapping (5,6, 26). A measurement of the conservation of
various parts of the
large
nuclear transcript is described that suggests that eachlong
RNA transcriptpossibly gives
risetoonly
onemRNA,
presumably
because each transcript containsonlyone setof 5'
spliced
sequencesnecessarytoformthemRNA.
MATERLALS AND METHODS
Celland virusgrowthaswellasthepurificationof
adenovirushave beendescribedpreviously (42).
Labeling
and extraction of RNA. HeLa cellswereinfected at multiplicities of2,000 particles per
cell. At theappropriate time afterinfection,the
in-fectedcelLswere concentratedto 10 cells perml in
fresh medium, and [3H]uridine was added (10 to 20
itCi/ml,
20mCi/pmol).
Afterlabeling, thecellswerechilled and collected by centrifugation. Cytoplasmic
andnuclearextracts werepreparedasdescribed
pre-viously.The extraction of RNAaswellasthe
prepa-ration ofpoly(A)-containingRNAby polyuridylic acid
Sepharose chromatography has been described (27).
Single-stranded RNAwasprecipitated from2MLiCl
overnightat4°C.
Electrophoresis of RNA.3H-labeled RNA
sam-ples (including 32P -28S and -18S rRNA markers) were
dissolved in ETS (0.01 M Tris-0.01 M EDTA-0.2%
sodium dodecyl sulfate [pH 7.4]) and made 50%
(vol/vol) formamide. After heatingto65°C for2mi,
bromophenolblue andglycerolwereadded, and the
samplewasappliedto a 2.7%acrylamide-0.14%
bis-acrylamide gel (0.6 by 10 cm) prepared in Loening
buffer(23).Electrophoresiswasat6mA/gelfor3 to
4 h. Gels were sliced into 2-mm fractions, and the
RNAwaseluted in 0.5 ml of TESS buffer [0.01 M
N-tris(hydroxymethyl)methyl-2-aminomethane-sulfonic
acid(pH7.4)-0.3 M NaCl-0.01 M EDTA-0.2% sodium
dodecyl sulfate] at 650C overnight. RNA was
frag-mented with alkaliasdescribed(16).
Electrophoresis in 98%formamide-containing gels
wasperformedasdescribedby Duesberg and Vogt (7).
Hybridizations. Hybridizations of RNA to
filter-bound Ad2 DNA fragmentswere carried out as
de-scribedpreviously (2).The RNAfrom each 107 cells
wasexposedtoDNAfragments equivalentto 1,ugof
total Ad2 DNA. Excess of DNAwasprovenby
rehy-bridizingsupernatantRNAtofreshfiltersrevealingat least 80% of the hybridizable RNA to be removed in the initial exposure.
RESULTS
Experimental design. LateinAd2 infection, the majority ofAd2-specific cytoplasmic RNA ispoly(A) terminated [poly(A)+;22,33]. More-over,almost all
(>90%)
of thelabeledpoly(A)+
RNAthat reaches thecytoplasmis Ad2specific (22, 32). Thus, by collecting the labeled poly(A)+ RNA lateininfection, the total virus-specific mRNAcanbeexamined without further purification. Because the firststepinthe design of thepresent work was to separateAd2-specific molecules on the basis ofsize, the electropho-reticmigrationofthetotal poly(A)+ RNA late in infectionwascompared under two conditions: (i) electrophoresis of denatured RNA in 98% formamide and (ii) denaturation followed by electrophoresis through gels not containing formamide. With both procedures (Fig. 1), dis-cretesize classesoflabeledpoly(A)+RNA mi-grating between 18S and 28S rRNA were ob-served aspreviously reported for Ad2-infected cells (22, 30, 39).The approximate distribution of labeled RNA compared with 28S and 18S rRNA markers wassimilar, but the resolution wasconsiderablybetter in thegelswithout the formamide. This technique was therefore adoptedforthepresent work.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
PROCESSING OF Ad2 mRNA's 813
The experiments to be described were
de-signedtomapthe variousmRNA'sindifferent
regions of the genome by first separating the mRNA according to size, followed by hybridi-zation of the labeled RNA toasufficientarray
of Ad2 DNA restrictionfragments (in DNA
ex-cess)sothatmostmRNAwouldhybridizeacross
a boundary between two fragments (Table 1). The percentage of labeled RNA ofaspecific size that hybridized to each DNAfragment would then allowassignment of theposition of the 5' and 3' termini of the mRNA exclusive of
post-transcriptionally addedsequences.Insomecases anmRNAspecies mightspan anentire fragment
andhybridize tobothright-hand and left-hand neighboring fragments. Such asituationwould
offer a measure ofthe length of mRNA
inde-pendent of its electrophoretic migration because the size of thefragmenttotallyspanned by the mRNA would be known from the DNA restric-tion fragment maps. The accuracy ofthe map
assignmentsmade with thesetechniques would
dependonthe errorinassaying the RNA:DNA
hybridization,the sizedetermination ofRNAby gel analysis, and theaccuracyoftherestriction fragment map ofthe Ad2 genome. An overall
accuracyof atleast1 to2%onthephysicalmap
would beexpected for the relative positioning of the variousmRNA's, especially when reviewed in light of the positioning of termini reported
A
10
5-10 -B
a-0 10 20 30
SLICE
FIG. 1. Electropherograms of
13HJuridine-labeled
late Ad2 RNA(0) together with 32P-rRNA (0) in98% formamide-containing polyacrylamide gel (A) and acrylamide gel preparedinaqueousbuffer (B). Elec-trophoresiswasfrom lefttoright.Aportion (20td)
of each elutedgel slicewascounted in triton scintilla-tionfluid.TABLE 1. DNA restriction fragmentsofAd2usedin
mapping mRNA locations
Map cooi
zyme(s)
used Name offrag-natee" ment
25.5-27.9 Hpa I Hpa F
29.1-36.7 Sma I/BamI SmaB/BamD
36.740.5 Sma I Sma I
40.5-52.6 Sma I Sma D
40.5-42.7 SmaI/Kpn SmaD/Kpn H
42.7-47.4 Kpn Kpn I
47.4-52.6 SmaI/Kpn SmaD/KpnD
52.6-56.9 Sma I Sma H
56.9-58.5 Sma I/EcoRI SmaA/EcoA
58.5-70.7 EcoRI Eco B
70.7-75.9 EcoRI EcoK,
75.943.4 EcoRI Eco D
83.4-89.7 EcoRI Eco E
89.7-100 EcoRI Eco C
aMap coordinates of Hpa I, Bam
I,
andEcoRI
cleavage sitesaretakenfrom datacollectedby Marc
Zabeauand distributed at the 1976 Cold Spring
-Har-bor Workshop on DNA tumor viruses. The Sma I
cleavage sitesaretaken from Carel Mulder (personal
communication). TheKpncleavage sites within the
Sma Dfragment were determined from the size of the
fragmentsproduced aswellastheir sensitivity to Sal
Idigestion.
from microscopic techniques (5, 6). The tech-nique ofhybridizing labeled mRNA
molec,ules,
ofcourse, also allows a means ofstudying the metabolismofspecificmRNA's anda quantita-tionof therelativeamountsofeachmRNA that reach the
cytoplasm.
Aspointed
outbelow,
nu-cleotide analysis of RNA and DNA is finally necessary tolocateexactly
theregion
of DNA correspondingto aparticular RNA.General results.The datafromseveralsuch RNA:DNAhybridizationsofelectrophoitically separatedRNA from cells labeled from15 to 18
h after infection areshown in Fig. 2, 3, and4.
The general results to note iniexam iing the data of
Fig.
2 and 3 are asfollows.(i)
mRNA hybridizations were carriedout to manydiffer-ent DNA
fragments
from 36.7 to89.7, and thesize distribution of RNA that
hybridized
toeach DNA fragment was distinctive; manydifferent specific mRNA'sfromapproximately lOSto28S (about -0.8 to 5 kb) wereclearly
present.(ii)
The reproducibility of the electrophoretic pat-ternisquite good; for example,compareFig.2A and
3A,
which represent two differentRNA-preparations
hybridized
to twodifferentprepa-rations of the DNA fragment 36.7-40.5. (iii) Some DNA
fragments
hybridize
two or three mRNA's, each of which is greater than the length ofthefragment. (Note
that above the mRNA was broken with alkali to chain sizes -0.2 to 0.3kb and thatall the DNAfragments
exceeded0.7kb andwerepresent in excess;
thus,
any
single
DNAmolecule wouldactually
hybrid-VOL. 25,1978
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.244.437.66.237.2] [image:3.501.75.184.376.589.2]814 NEVINS AND DARNELL
X 30* a.
u
SLICE
FIG. 2. Electropherograms oflate
rH]uridine-la-beled Ad2 RNAhybridizedto(A) Sma I(36.7-40.5)
filtersandto (B) Sma D(40.5-52.6) filters.
Electro-phoresiswasfrom lefttoright, with thearrows
indi-catingthepositions of 28Sand 18SHeLa rRNA's.
ize lessthan its fulllength of RNA, anecessity
in filterhybridization experiments if the DNA is
not to be released from the filter [15].) For
example, Fig. 3B shows that fragment 42.7-47.4, which is1.64 kblong,hybridized three mRNA's, 27S, 23S, and 19S RNA, all larger than 18S
rRNA, which is 2.1 kb long. Clearly, several different discretemRNA's share atleastapart
of their sequences. This proved to be true in virtuallyeveryregion of the large transcript that gives risetomRNAmolecules.(iv) Eachoneof
theeight fragmentstotheleftof83.4hybridized large RNA migratingtogether with 28S rRNA,
which is5kblong. Because the DNA fragments used in thepresenthybridization workrepresent
about 18 to 20 kb intotallength,atleastthree
tofour suchlarge mRNA's mustexist. (v) The DNA fragment that hybridized the largest
amount of labeled -27Sto 28S mRNAwas in
the 52.6-56.9region where the hexon mRNA is knowntomap(9, 21). This finding is consistent with the earlierobservation that hexonprotein is themostprominently synthesized protein late in infection(45). (vi) Althoughtotalcytoplasmic poly(A)+RNAwasused for theanalyses of Fig.
2and3,poly(A)+ RNA from polysomes yielded
patterns after hybridization identical to those
shown inFig.3(datanotshown).
Sortingoutthe27S to28S mRNA's. The
large 278 to 28S mRNA's that hybridized to
virtually every DNA fragment (Fig. 2 and 3)
werepositionedasfollows.
(i)The 38-50 mRNA. A27SmRNA
hybrid-ized both to the 36.7-40.5 region and to the 40.5-52.6region.It didnot, however, hybridize
tothe29.1-36.7region (datanotshown), placing
its 5' end inthe 36.7-40.5 region. About three
10
20
C 474 52.G
20
D 52 6 56,9
80
40-2 SL
(.) E 707 75.9
15
-10
5-F 175.9 834
10
L-
~
5-G 83.4 897
40
--
20-0 10 20 30 40 SLICE
FIG. 3. Electropherogramsoflate
"Hiuridine-la-beled Ad2 RNAhybridizedto(A)Sma I(36.7-40.5),
(B) KpnI (42.7-47.4), (C)SmaD/Kpn D
(47.4-52.6),
(D) Sma H(52.6-56.9),(E)Eco F(70.7-75.9), (F)Eco
D(75.9-83.4),and(G)Eco E(83.4-89.7)DNAfilters. Electrophoresiswasfrom lefttoright.
times as much of this labeled 27S mRNA
hy-bridized
to the 42.7-47.4region
as to the36.7-40.5 region. Because 42.7-47.4 is 1.64 kb long and is
entirely
traversed by the largemRNA,
then -0.5to0.6kb of thelong
mRNAon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.103.205.47.218.2] [image:4.501.309.408.57.546.2]PROCESSING OF Ad2 mRNA's 815
10
0v 2 a.
8
6
4
2
a. u
0 10 20 30 40 50
SLICE
FIG. 4. Electropherogram of late
3HJuridine-la-beledAd2RNAhybridizedtoSma H(52.6-56.9) (0)
andtoEco E(83.4-89.7) (-)DNAfilters.
Electropho-resiswasfrom lefttoright;arrowsindicatepositions
ofHeLa28Sand 18SrRNA's.Inthisexperiment the
gelwasfractionated into1-mmslices rather than
2-mmslicestoafford higher resolution.
should lie to the left of 40.5 in the 36.7-40.5 region(Table 2). The 5' end of this RNA
(exclu-siveof the spliced sequences) is thus placedat
about38 to 39. If this mRNA is 4.5 to 5.0kblong and contains -0.3 kb of 5' and 3' sequences added duringprocessing, then it would occupy
about4.2 to4.7kb,or12 to 13%ofthegenome. Its encoded 3' end would then lie around 50-52. (ii) The 50-62 mRNA and the 68-80 mRNA. Thenextpossible 4.5to5.0 kbRNAto
the right of -50-52, without extensive overlap with the previous large mRNA, would begin around50and extend toaround62.
Hybridiza-tion to DNA fragments in this region, in fact,
identified such anmRNA. The 52.6-56.9
frag-mentfrom the hexon coding region (9, 21)
hy-bridized averylarge amount ofmRNA in the 27S to 28S size range as previously noted. In
additionto theveryprominent27Speakinthe hybridization patterntothe 52.6-56.9region, a
shoulder of 288 was noted. Further analysis under conditions of higher resolution demon-strated aseparation of two RNAsnominallya
27S anda28S species, withapredominance of label in the 27S species (Fig. 4). These promi-nently labeled mRNA's extend through 56.9-58.5 (a short fragment for which
hybridi-zation data is not shown inFig. 3,but isgivenin
Table2).Inaddition, the neighboringrightward fragment 58.5-70.7 also hybridized a large
amount ofRNA in the 27S to 28S size range.
Thus,itappearsthatthesetwomRNA'sbegin
around 50-52, span the 52.6-58.5 region, and
terminate in the left portion of the 58.5-70.7
fragment. The proportion of the radioactive
RNA hybridized to 58.5-70.7 that was to the 50-62 mRNA's land therefore the number of basescovered in the58.6-70.7region) couldnot
a
llcJ
IW
.
.4
.
m3
E,
IC..
.2
L..
.0
EN4
-6.
a 43
2
0. 10
.0
C-. t-.
t--
C--coi
LOes
C--:
r-to
0._
4)
4,
z
E-CO _
oCD
cO CwD
Q0
8
O es0~ oe
oLO Co
-4
qeq -4eq -4 q -4 -4 v
X
--04
A
.s
eq C 7 eN
C-.
eq
4,4
4-4
0
o
04
b.
XC.)
.
-* cx
Bz
VOL. 25, 1978
28S 18S- 4
4
4
.1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.269.453.52.654.2] [image:5.501.73.218.56.196.2]be simply calculated (see Table 2), however, because thenextfragmenttotheright, 70.7-75.9, alsohybridizedanmRNA ofapproximately27S. However, the 70.7-75.9 fragment is only 2.1 kb longitself,soalong27S mRNA mustprojecton
eitherorbothsides. The 75.9-83.4fragment did
indeedhybridizea27SRNA butalesseramount
ofradioactivitythan the 70.7-75.9fragment (Ta-ble 2). Fromthe proportion of -27S RNA hy-bridized to 70.7-75.9 and 75.9-83.4, it could be concluded thatatleast 2 kb of thelong mRNA spanningthe 70.7-75.9regionmustlie beforethe
70.5-75.9region,thatis,in the58.5-70.7region. Therefore, two different long mRNA's derive
sequences from either endofthe 58.5-70.7 re-gion.
Insummary, wecanidentify four large
-4.5-to 5-kb mRNA's, one from 38-50, two ofvery similar size in the 50-62 region, and one from
68-80.
Common 3' termini: the 36.7-62 region. As pointedoutabove,severalregionsofthe Ad2
genomeappeartoencodemultiple mRNA'sthat
sharesomeof thesamesequences.Suchagroup of mRNA's might be arranged in any of the
followingways: (i)code for thesame or apartof
the same polypeptide and share either5' or3'
termini; (ii) share a terminus, but the longer mRNAmightbe translatedonlyover aportion
of its totallengthtoproduceadifferentpeptide from the shortermRNA(reminiscentofSindbis
virus [36]);or (iii) existas "overlapping genes"
that might be translated in different-reading framesaswith 4X174(3)and simian virus40(S. Weissman, personal communication). An effort
wasmade, therefore,to calculateterminal
posi-tions ofsuchoverlappingmRNA's fromthe size
estimate affordedby electrophoretic migration
plus a determination of the fraction of such
mRNA's that hybridized to contiguous DNA
fragments.
The data suggestanumber ofregionswithin
which from two to four mRNA's seem to
ter-minate. In one of these regions, -50 on the genome,ourconclusionsaregreatlystrengthened
by the data of Ziff and Fraser (46) that the
oligonucleotidesof three different-sizedmRNA's
areindistinguishable. Because we reached the
sameconclusiononthe basisof theproportion
of hybridization of various sized mRNA's to
DNAfragmentsbetween 40 and52,wefeelmore
confidentthatwecan place3' endsreasonably accurately.Inaddition,ourdataagreewiththe
assignment of 3' ends by electron microscopic data (5, 6, 26). Nevertheless, inthe absence of
fingerprintdataonvarious-sized mRNA'sfrom
each putative 3'-coterminal site, we cannot
firmlyconclude 3'identity.
We will first consider the mRNA's
comple-J. VIROL.
mentary to fragments between positions 36.7 and 52.6.Thereare three species ofRNA, 27S, 25S, and 20S, complementary to 36.7-40.5, a fragment that is itselfonly 1.33 kblong.Thus, each of these three mRNA'sshould also hybrid-ize to DNA on one or the other side of the 36.7-40.5 region. The examination of labeled RNAcomplementary totheneighboring right-handfragment,40.5-52.6, showed thatthree dis-tinctive RNA peaks were also observed, 27S, 23S, and 19S, only the longestofwhichcoincided with the mRNA's complementaryto36.7-40.5. Because all the mRNA's in the entire region (16-100) aretranscribedtotheright(5'--3'),the twosmaller mRNAhybridizedto the36.7-40.5, therefore, mustderive sequences to the left of 36.7-40.4, i.e., from the 29.1-36.7 region, and bothmusthave their3' ends within36.7-40.5. A separatepreparation of RNAwasprepared and hybridized to the 29.1-36.7 fragment. No 28S RNAhybridized, butboth a25S and 20S RNA were detected. Thus, these two mRNA's, the 25S and 20S, have sequences in the 29.1-36.7 region and end inthe 36.7-40.5,but we cannot determine precisely their3' termini. Thelarge 27S mRNAbegins within 36.7-40.5and contin-uesinto40.5-52.6.
Todetermine moreaccurately the map posi-tions of the three RNAs in the40.5-52.6 region, further subdivision of thisDNAfragmentwith another restrictionenzyme(Kpn)wasrequired. DNA fragments with coordinates 40.5-42.7, 42.7-47.4, and47.4-52.6 wereobtained,and the proportion of radioactivity hybridizedfrom each of the 27S, 23S, and 19S mRNA's to each of thesefragmentswasdetermined(Fig.3B and C, Table 2). The 23S and 19S mRNA's failed to hybridize to the 40.5-42.7 fragment (Table 2), but both mRNA's as well as the 27S RNA hybridizedto42.7-47.4and47A-52.6 fragments (Fig. 3B and C). However, no hybridization of 23Sand 19S mRNA'soccurred to the 52.6-56.9 fragment, demonstrating that the encoded 3' ends of these molecules must lie within the 47.7-52.6region.If a27S RNAdoesbegin in the 36.7-40.5 asconcluded above, italso could not extendbeyond approximately 50-52on the ge-nome. Thus, it appears likely that three mRNA's,27S, 23S,and19S,endnearthe 50-52 region.
Ziff and Fraser (46) have identified three mRNA's of approximately the same lengths (27S, 23S, and 19S) asdescribed here whose 3' terminal 100 nucleotides, i.e., the nucleotides next topoly(A), hybridizetothe40.5-52.6 DNA fragment. Asmentioned previously,
all
three of these 3'endshave the sameoligonucleotide com-position; moreover, thesedistinctive oligonucle-otidesarealsopresent inEscherichia coliRNAon November 10, 2019 by guest
http://jvi.asm.org/
VOL. 25, 1978
polymerasetranscripts of the47.4to52.6region. Thus,we canconclude that thesethree mRNA's (27S, 23S, and
19S)
all terminate very closeto thesamesite.Common
terminii:
the 50-62, 68-80, and 83-92 groups. Finally, it appears that three other groups of mRNA that, within a group, maysharecommon3'-terminal regions exist in the 50-62, 67-83, and 83-92 regions of the ge-nome. As discussed above, two large RNAs, a 28S and 27S species, can be resolved by high-resolutiongel analysis in the50-62region (Fig. 4). Asdiscussedlater, if thesetwomRNA's have nonoverlapping 5'regions thatmight be neces-sary for each to be translated into different proteins, theymayhavecommon3'termini.Next, both the 70.7-75.9 and 75.9-83.4 frag-ments(Fig. 3E, F, and G)hybridizeda27S, 22S, 19S mRNAgroup. A small -1.5kb mRNAmay
also
hybridize
to the 75.943.4 fragment. Byestimating the proportions of these various mRNA's
hybridizing
toeach ofthesetwofrag-ments
coupled
with the gel estimates of the mRNAsizes, itseemslikely
that all four of these mRNA's have 3'-terminal sequences inthe re-gion of79-80.
All wouldlikely terminate before the beginningof yetanothermRNAaround83 that continues across into theneighboring
83.4-89.7region.
The80-90regionhas beenshownbyavariety
of
experiments
to encode thefiberprotein (21,25).Aprominent 24SpeakofRNA
lying
mainly
in the 83.4-89.7 region was resolved into two mRNA's by
high-resolution
analysis (Fig. 4). These mRNA's extend into the 89.7-100 frag-ment, adistanceequal
toabout one-fifth of that whichliesinthe83.4-89.7region
(Table
2). The larger RNA of the two maybegin
in the 75.9-83.4 fragment because a small amount of labeledRNAof thecorrectlengthhybridized
toPROCESSING OF Ad2 mRNA's 817
both thisfragment and the 83.4-89.7 fragment (Fig.3F andG). Possibly,it isthesmallerRNA that encodes the fiberproteinbecause theregion of from -80 to 85 is nonessential for virusgrowth (9), whereas thefiberprotein is most likely an essentialgeneproduct.
Summary of mRNA detected. The tech-niquesusedfor thesehybridization studies dis-tinguishaverylargenumber of mRNA's, prob-ably asmanyas 12,that aredistinct in sizeand in map position. The earlier data on protein synthesis programmed by Ad2 mRNAhas also identifiedatleast ninespecificproteins encoded in thisarea (9, 20, 21). In additionthe in vitro protein synthesis studies identified the size of various mRNA's that gave maal program-ming for particular proteins. Thepresent work indicates many overlapping mRNA's, and it seems
possible
thatthe
5'mostsection of each Ad2 mRNA isublizedintranslationas, in fact, is the case for togaviruses (18, 36) and RNA tumorvirus(17, 38) mRNA's. These RNAs con-tain two or morepotential entrysites for ribo-somes,butonly the 5' site is active.By making theassumptionof 5'regionsbeing the translated portions ofoverlapping mRNA's and using all the availabledata, themapassignments shown inFig.5 weremade.Table3givesasummary of thecalculationsemployinggel-estimated lengths andproportions of RNAhybridizing tospecific regions that have been used to determine themap
positions
presented
inFig.
5. We wish toagain emphasize that only in theregion of50is thereoligonucleotide evidence ofexact 3'-coter-minal
mRNA's,
butthe dataonthisregion
ob-tainedby hybridization predicted
the coterminal natureof thesemRNA's inthat region.Our bestestimate
wouldbe,
therefore,
that theother
groupsofmRNA'smayalsoterminateat
com-mon3'
poly(A)
addition sites.0
I
P[
p II pvInitiation site
0 10 20 30 40 50 60 70 80 90 100
F Hpo I
c---I D H Sm
(Ec
Kpn H I D (Sma D)
Eco RI B F D E C
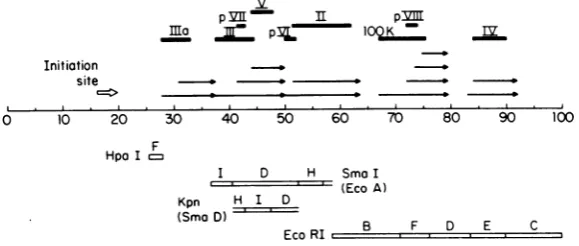
FIG. 5. Schematic representation of late adenovirus RNAs generated in region of 27-92fromthe adeno
genome.Positions of RNAs werederived fromdatapresentedin Fig. 2 through 4 andTable3. Solid bars
above RNAs indicatepossiblemaplocation of late proteins, based on approximate genome locations and
sizes of mRNA's encoding respective proteins (20). Initiation site at 16 for the major latetranscriptionunit is
fromdataofEvans etal.(8).
0
--w
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.107.397.484.607.2]TABLE 3. Sizes andcoordinates of late adenovirus RNAs
Approximate Estimated Coded size Genome Coordinates
Fragment Svaluea length(kb)b
(kb)'
(%)d3SmaI 25 3.8 3.5 10.0 28.5e 38.5f
20 2.7 2.4 7.0 31.5e 38.5f
SmaD 27 4.7 4.4 12.5 38.5(±0.5)' 51.0e
23 3.3 3.0 8.5 42.5e 51.0(±0.9)'
19 2.4 2.1 6.0 45.0(±0.4)eg 51.0(±0.9)'
SmaH 28 5.1 4.8 13.5 49.5e 63.0(±1.7)'
27 4.6 4.4 12.0 51.0e 63.0(±1.7)'
EcoF 27 4.7 4.4 12.5 67.Oe 79.5'
22 3.0 2.7 7.5 72.0 79.5
19 2.4 2.1 6.0 73.5 79.5
EcoD 13 1.5 1.2 3.5 76-79.5 80-83.5"
Eco E 24 3.4 3.1 8.5 83.0e 91.5
(±0.2)'
23 3.1 2.8 8.0 83.5e 91.5(±0.2)'
aNominalS value determined from gel analysis by comparison to 28S and
18S
to bethe same S value relativetorRNA insucrosegradientsforhexon mRNA(SmaHspecific)and fiber mRNA (Eco E specific).
b
Estimated
lengths given by Loening (23), assuming 28S and 18S to be 5.1 and 2.1 kb, respectively (44).cSize from footnote b minus150nucleotides for the poly(A) segment and 150 nucleotides for the 5' noncoded
leader sequence.
dValue in footnote c dividedby 35,500 and recorded to the nearest 0.5%.
eLength of RNA fromgel analysisand one terminus calculated as in footnote e allowed calculation of second
terminus.
fAssumption has been made that where an RNA terminates and a new RNA begins, no gap or overlap exists
between thetwo.
' Determined on basis ofoverlap of RNA in two adjacent fragments (see Table 2).
hBecause thisparticularRNA liesentirelywithin the Eco D (75.9-83.4) fragment and no overlaps were
obtained,theexactcoordinatesareunknown.
Presence of RNA sequences from
25.5-27.9 invarious mRNA's. Because each
mRNA deriving mainly from sequences tothe right of30 is thought to contain three ligated oligonucleotide regions (4, 5, 12, 19), labeled
RNA was hybridized to one small DNA
frag-ment that contains one of these regions. The
25.5-27.9DNAfragment,itselfonlyabout0.8 kb
long, hybridized labeled RNA from every size
class between 2 and 5 kb, with broad peaks corresponding to the major size classes of
mRNA (Fig. 6). The total amount of labeled
mRNAhybridized tothe 25.5-27.9 DNA
com-paredtothe total of all otherfragments to the
right of30wassmall(-2 to4%) anddistinctly higherin the 2- to 3-kbrangethan in the5-kb range.Thus,the amountshybridizedto25.5-27.9
areinagreementwith thepossibilityof approx-imnately 100 nucleotides from 25.5-27.9existing ineverymRNA(Table 4).
Inaddition asmall (8S) speciesofRNAwas
found hybridizing to the 25.5-27.9 fragment (slices33 to35 in bothtotalmiRNAandRNA
hybridizingto25.5-27.9) thatwasnotfound to
hybridize to any fragment in the region from
29-92. The possibility that a specific mRNA
existsthat is encodedbytheDNA in thisregion ofthegenomeis under furtherinvestigation.
Cellular location of processing event.
TABLE4. Hpa F(25.5-27.9)hybridizationa
25.5-27.9 30 to100 % Nu-RNAspe- hybridi- hybridi- 2%55n ticdeos cies zation zation 27.9
25.5-(cpm) (cpm) 27.9
27Sto28S 860 37,650 2.3 104
22Sto24S 330 11,200 2.9 84
300 10,900 2.8
19S to 20S 250 6,800 3.7- 81
8S 120
'Onepreparationofpoly(A)+RNAwasseparated
bygelelectrophoresisandwasusedtoobtain data in
Fig.3and6.Thecountsperminutehybridizedtothe
HpaF DNA
fragment
(25.5-27.9)arecomparedin thistabletothe total hybridizedtoall other DNA frag-mentsbetween30and 100. Slice10represents27Sto
28SRNA,15and16represent22Sto 24SRNA,and
19 represents 19S to 20S RNA. The sizes of the
mRNA's[poly(A) and 5'leader]weretaken fromgel
migrationtobe4,500,2,900, and 2,200.
Aloni et al. (1) have described results late in simianvirus 40 infectionthatsupport a possible cytoplasmic processing of a 19S into a 16S mRNA. Because so many of the mRNA's de-scribed in this reportalso overlapin sequence, asdo the simian virus 40mRNA's, the possibil-ity of cytoplasmic processing might also be raised for Ad2 mRNAformation.Infectedcells
818
NEVINS AND DARNELL J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.271.460.375.473.2]PROCESSING OF Ad2 mRNA's
werelabeled for 45min (in contrast to3or4h
used for all previous experiments); one-half of
the culture was harvested, and the remainder wastreated ("chased") withactinomycin Dfor an additional 3 h to suspend further mRNA labeling. The mRNAwasanalyzedasbeforeby
hybridizationtothe40.5-52.6 andthe 70.7-75.9 regions from which several apparently
3'-coter-minal mRNA'sarederived (Fig. 7). All species of mRNAwere present after the 45-min label
time in approximately the same ratios as
ob-served for the 3-h labeltime (see Fig. 3 and 4), and nosignificant changewas observed in the
absence of further RNAsynthesis for 3 h. Fur-thermore, the polysomal RNA fromcelLslabeled
continuously from 12 to 28 h of infection
con-tainedvirtuallyidenticalproportionsofthe
var-ious mRNA'sineachregion (datanotshown). Next, celLs werelabeled for 12 min, and nu-clear,poly(A)+ RNAwasisolatedandsubjected
togelelectrophoresisandhybridizationtoDNA
in the 40.5-52.6region (Fig. 8). All the species previously detected in the cytoplasm, the 27S,
238,
and 19SmRNA's,werealsopresentinthenucleus. Itappears,therefore, thatthe process-ing of RNA to yield mRNA from the Ad2-in-fectedcells occursinthe nucleus.
Efficiency of mRNA formation from pri-marytranscripts.One oftheprimereasonsfor
obtaining detailed information onthe map po-sition ofspecific Ad2 mRNA's wasto allow a
A 28S 18S
10
10 -B
0
5
0 10 20 30 40
[image:9.501.273.415.168.351.2]SLICE
FIG. 6. Electropherograms oftotal unhybridized late 3HJuridine-labeled Ad2 RNA (A) and RNA
hybridizedtoHpaF(25.7-27.9)DNAfilters(B).
Elec-trophoresiswasfrom lefttoright. Aportion(10 1d)of
eachelutedgel slicewascountedtoobtainthe profile in(A).
study of the efficiency of utilization as mRNA
of differentsegments ofthelarge late primary
nuclearRNA transcript. Suppose thata
partic-ular mRNAportion ofaprimarynuclear
tran-scriptwasall convertedtocytoplasmic mnRNA within 15to20min and this particular cytoplas-mic mRNAwasrelativelystable. The increase
in that fraction oflabeled nuclear RNA
corre-spondingtothesequencepresentin the
partic-0
0.
4
2
6
4
2
10 L
[image:9.501.84.203.388.590.2]SLICE
FIG. 7. Electropherograms of late
pH]uridine-la-beledAd2 RNAhybridizedtoSma D(40.5-52.6) DNA
filters (AandB)andtoEco F(70.7-75.9)DNAfilters
(CandD). (A)and(C)representRNAlabeledduring
a45min pulse,and(B)and(D)representRNAfrom
ceUslabeledfor45minandexposedtoactinomycin D(5pg/ml)foranadditional 3 h.
6
N
I0
0.
4
2
28S 18S
Is
i
I20
I 10 20
0
SLICE
FIG. 8. Electropherogram oflate
f3HJuridine-la-beledpoly(A)-containingnuclearAd2RNA hybrid-ized to Sma D (40.5-52.6) DNA filters. RNA was
labeledfor12 min at15 hpostinfectionandprepared
asdescribedinthetext.Electrophoresiswasfrom left
toright.Arrowsindicatepositions ofHeLa 28Sand
18S rRNA's. The nuclei in this preparation were
detergentwashedtoremoveallcytoplasm.
A 28S 18S C 28S 18S
4 4 4 4
I
-t
B 4 0 4
0 10. 20 0 10 20
819 Voi. 25,1978
3
2
5
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.501.299.394.424.573.2]ular mRNA would be followed in 15 to20min byaparallel increase inthis particular labeled
mRNA. A timecourseofincorporation of
[3H]-uridine into nuclearandcytoplasmic RNAwas first determined for RNA complementary to
Ad2 DNA fragments 52.6-56.9, 70.5-75.9, and 83.4-89.7; all of these DNA regions are either
entirely ormostly represented in mRNA. The curveof labeled nuclear RNA for eachfragment rose rapidly, as expected, reaching a plateau
after about 40 to 60 min (Fig. 9). The labeled mRNA begantoappearapproximately linearly after about 20 min. Therate oflabeling of
nu-clear andcytoplasmic RNA specific for the three regions wasestimated from the maximalslope
of increase and is shown in Table 5. From the maximum rates of labeling of the same
se-quencesinthe nucleus andthenthecytoplasm, the utilization of thenuclearRNA foranyofthe threeregions wascalculated; about 20% of the
52.6-56.9 region and about 15% of each of the othertworegions wereconserved. Because the
nucleiwerenotstripped of cytoplasm by
deter-gent treatmentinthisexperiment, atleast 15%
of the cytoplasmic RNA might remain in the nuclear fraction (41). Therefore, the poly(A)+ nuclear RNA of thecorrectsizeformRNAwas
measured by hybridization to 52.6-56.9 and 83.4-89.7. In each case, an amount oflabeled
RNA equalto about 20to30% ofthatalready detected in the cytoplasm was found. If this
poly(A)+ RNA is added to that in the free cytoplasm,the maximum numbers forthe
trans-portefficiency wouldbebetween 20 and 26% for
these three sections of the Ad2 genome. An
eJ
20-15-8
0 '0x
10.
5
0 40 800 40 80 0 40 80
MINUTES
FIG. 9. Kineticsofsynthesis ofadenovirusnuclear
RNA and the appearance oflabeled RNA in the
cytoplasmspecific forDNAfragments52.6-56.9(A), 70.7-75.9(B), and 83.4-89.7(C). HeLa ceUs infected for 15h with adenovirus were concentrated to 10i ceUsperml,and
fIHluridine
wasadded to100#0Ci
ml.Samples of2x107cellswereremoved at indicated
intervals, and thenuclear (0) andcytoplasmic (0)
RNAswereprepared.Each RNAsamplewas
precip-itatedwithLiCItoremoveviral DNA.Hybridizations
wereperformed with filters containingDNA
frag-ments equivalent to anamountpresentin 5 pgof adenoDNA.
TABLE5. Efficiency oftransportoflateAd2 RNAs
DNA fagment %Transport"
36.7-40.5 5
40.5-42.7 4
42.7-47.5 19
52.6-56.9 17
70.7-75.9 13
83.4-89.7 13
aDetermined from rate of
labeling
of cytoplasmicRNA between 45 and 90 min (counts per minute
increase per45min) dividedbytherateoflabelingof
nuclearRNA between 5and 20 min (counts perminute
increaseper15min).Theincreaseinradioactivitywas
approximatelylinear duringthetimechosen for the
estimation(see Fig. 9).
analysis was also made of regions of theDNA genomewheremultiple mRNA's arise, namely, 40.5-52.6. In such regions where the primary transcriptappears tobe used for more than one type ofmRNA, the transport efficiency should vary for RNA complementary to contiguous DNAfragments, being higherwhere sequences wereshared byallmRNA'sandlower for frag-mentsthat providesequences exclusively for one ofthegroup ofmRNA's.The efficiency of trans-port of nuclear RNA from the 40.542.7
frag-ment that
hybridizes
the 27S but notthe 23Sand 19S mRNA's(seeFig. 3)wascompared with the efficiency oftransport ofthe nuclearRNA from the 42.7-47.4 region that is completely includedin two andpartially includedin athird mRNA.
Only
4% of the 40.5-42.7 region was transported compared with19% ofthe42.7-47.4 region. Another region, 36.7-40.5, which pro-vides sequencesfor several mRNA's, including along
27S mRNA with several shorter 3'-co-terminal mRNA's, also showed a transport ofonly
5%.These dataare mostcompatible with the in-terpretation that each
primary
transcript gives rise to a limited number ofmRNA's, possibly one; thus, any one ofthepotential mRNA re-gions existswithonlya5 to30%success rate.DISCUSSION
Themajorfacts
established
bythis work are shown inFig.5-alarge number,perhaps 12 or more, mRNA species of distinct size (electro-phoreticmigration)
and genomepositioncanbe distinguishedfrom 29.1through100.McGrogan and Raskas (24) also have recently demon-stratedmultiple speciesof RNA inthisregion. These mRNA'saresynthesized(labeled) latein infection, and all appear to derive from one primary nuclear RNA transcript (2, 8, 13, 14, 43).All themRNA'scontaina3'poly(A),
several have beendefinitely demonstratedtocontain aJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.501.269.459.76.150.2] [image:10.501.65.256.438.533.2]PROCESSING OF Ad2 mRNA's 821
5'cap(12), and from theaveragecontentofcaps
in the total late Ad2 mRNA's,most molecules
mustcontainacap(37). In each specificcase so
farexamined, the nuclear processing also
appar-ently involves breakage and reunionat16.7,19.7
and 26 in theprimary transcript toproducean
mRNA(4, 5, 12, 19). Additional supporting evi-dence is offered on this point by the datum reported here than DNA from 25-27 hybridizes
anamountoflabeled RNAequivalent to -100
nucleotide portions ofevery size class of Ad2
mRNA's.
If this picture of multiple recombinational possibilities for the late 16-100 primary
tran-script is correct, manyintriguing questionsare
posed.
(i) Do all of theseeventsoccurin the nucleus?
At leastapreliminaryanswertothisquestion is afforded in Fig. 8, where it is shown that all ofa
group ofoverlapping mRNA's observable after long labels in thecytoplasmare labeled within
12min in the nucleus. Moreover, the proportions of these mRNA's in the cytoplasm appears to
remain fixed in the absence of further RNA synthesis; the processing events, at least for
someof the Ad2 mRNA's, would appeartobe nuclear.
(ii) Canaprimarytranscript give risetoonly
onemRNA?Iftheputative ligation sitesat16.9, 19.7, and 26.0 arenot repetitive at those sites (and thiscanonlybetoldbysequenceanalysis), then one transcriptcould donate the 5' 150 to
200 nucleotides to only one mRNA. Such a
mechanism wouldallowanyspecific mRNA
re-gion toexitto the cytoplasm withafrequency
sustantially less thanone. Anattempt to
mea-suretransportefficiencyofseveralregionsof the primary transcript known to encode major mRNA speciesshowsapproximately 15to20% conservationfor three of the fourand 20 to 30%
conservationforone (thehexonregion).These
resultsareinaccord,butclearlydo notestablish
the proposal that one transcript can be
con-verted intoonlyonemRNA.
(iii) Whatlocatestheprimary cleavage sites?
The moststriking generalfact aboutthe
distri-butionofcleavagesites(or mRNA termini) con-cerns the 3' termini. There may be only five
different 3' regions that exist for 12 different mRNA's-one group of mRNA's (ending at
'50) share theexact same3'regions (46), and
theremaybe fourothersuchcommon 3' sites.
However, each of the 12 mRNA's may have differentsequencesin its5'portions, excluding, ofcourse,theterminal -200nucleotidesthatare
presumablycommontoallmRNA's.
Themeaning ofthisarrangementof presumed coding regions may possibly parallel the mode
of polypeptide synthesis in certain togaviruses
(18,36),the retroviruses (17, 38),andthe papo-vaviruses
(S.
Weissman,
personalcommunica-tion). For
example,
withSindbis
virus (35) and Rous sarcoma virus(38),
mRNA's ofvarious
lengths exist that are translated into a single polypeptide. Thus, we might expect the 28S mRNA from 50-62 to encode the
small
pVI
protein from its5'region andthe 27SmRNA,a slightly shorter molecule, to lack the pVI se-quence and encode the hexon protein. In this case nooverlappingcodingsequenceswould oc-cur.Lewisetal. (21) suggested suchapossibility
several
years agobut didnotattempt toeffectacomplete
separation
ofthetwomRNA'stoprove thepoint.Intheregion that encodes thepentonprotein, 40.5-52.6,theoverlapping mRNA'sandthe pro-teins encoded by eachpresent greater
difficul-ties. The penton protein is -70,000 daltons, whichrequiresatleast2,300nucleotidesorabout 7.0% of the genome.The overlapping mRNA's in this region appear not to leave that much space free at the 5' end of any of the three mRNA's in the region. Thus, these mRNA's might be readin adifferent frameasis thecase for certainregionsin
4X174
(3) andsimianvirus40
(Weissman,
personalcommunication).
Alter-natively,thesemRNA'smight encode polypep-tides thatwereidenticalattheirCOOH-terminal segments but
possessed
differentNH2
termini. (iv) Whatarethe relativeproportions
ofthe various mRNA's and how are these decisions mediated?The one clear fact is that themRNA's from different
regions
neitherexitnorexist
atsteady
state in
equal
concentration in thecytoplasm;
e.g., there ismuch more labeled hexon mRNA than any other mRNA.
However,
Flint and Sharphave shownmRNA from70-76tobe themost
populous
atsteady
state(10).
Does thismeanthat the
cleavage
schedule favors certain regions over others? Notnecessarily.
If therewerefive
equally probable
initialcleavages,
sayto createfivedistinct 3'
termini,
followedby
asubdivision within these groups based on the number of mRNA's within each
section,
thennoprimary
cleavage
mechanism would berequired
todivide the
primary
transcript
unequally.
Such amodel ofprocessing
accordswith,
butby
nomeans is
proved
by,
the present data. In fourdifferentregions,RNAappearstobe conserved with about 20%efficiency.Thegreater
accumu-lation of hexonmightthen be
explained
bythe existence ofonly
twomRNA's fromthisregion. Clearly this discussion of thesecomplicated
eventsshould
only
betaken asan introductionto the
problem.
Forexample,
no mention hasyet been made of
cytoplasmic stability
of differ-entmRNA's,possiblyamostimportant
pointinVoi. 25,1978
on November 10, 2019 by guest
http://jvi.asm.org/
822 NEVINS AND DARNELL
controlling relative concentrations of the differ-ent mRNA's. The point worth making is that eventhougharemarkably complex setof
post-transcriptional
events is involved in producingthewelter of Ad2 mRNA's from the large tran-script, there is as yet no need to hypothesize that these are coordinated into a regulatory function. Suchsteps maybenecessaryand de-liver different amounts of primary transcript into differentmRNA's, but thismay occurwith afixed
probability
andnotbearegulated
setof eventscapable
ofresponding
to change. Only moreexperimentscansettle thisissue.ACKNOWLEDGMENTS
WethankMarianneSalditt-Georgieff and Patricia White forexcellent technical assistance and NigelFraser for the generousgiftofKpnDNA restriction fragments.
This work wassupportedby Public HealthServicegrant CA 16006-04from theNationalCancer InstituteandAmerican CancerSocietygrant NP213G. J.R.N. isaFellow of the Jane Coffin Childs MemorialFund for MedicalResearch.
LITERATURE CITED
1.Aloni, Y., M.Shani, and Y. Reuveni. 1975. RNAs of simian virus40inproductivelyinfectedmonkeycells: kinetics offormationanddecay in enucleate cells. Proc. Natl. Acad. Sci. U.S.A. 72:2587-2591.
2. Bachenheimer, S., and J. E.Darnell.1975.Adenovirus 2mRNA istranscribed aspart of ahigh molecular weight precursor RNA. Proc. Natl. Acad. Sci.U.S.A. 72:4445-4449.
3.Barrell, B. G.,G. M.Air,and C. A. Hutchison m. 1976. Overlapping genes in bacteriophageOX174. Na-ture(London)264:34-41.
4. Berget,S.M., C.Moore,and P. A.Sharp.1977.Spliced segments at the 5' terminus of adenovirus 2late mRNA. Proc.Natl. Acad.Sci. U.S.A. 74:3171-3175.
5. Chow,LT.,R. E.Gelinas,T. R.Broker,and R. J. Roberts.1977.Anamazingsequence arrangementat the 5' ends of adenovirus2messenger RNA. Cell12:1-8. 6. Chow, LT.,J. M.Roberts, J. B.Lewis,and T. R. Broker.1977. Amap ofcytoplasmicRNAtranscripts fromlytic adenovirus type 2, determined by electron microscopy of RNA:DNAhybrids.Cell11:819-836. 7. Duesberg,P.H.,and P. K.Vogt.1973.Gel
electropho-resis of avianleukosis andsarcomaviral RNA in form-amide:comparisonwith other viral andcellularRNA species.J. Virol. 12:594-599.
8. Evans, R.,N. W.Fraser,E.Ziff,J.Weber,M.Wilson, andJ. E.Darnell.1977.The initiation sites for RNA transcriptioninAd2 DNA.Cell12:733-739.
9. Flint,J. 1977.Thetopographyandtranscriptionof the adenovirusgenome.Cell10:153-166.
10.Flint, S.J.,andP.A.Sharp.1976.Adenovirus transcrip-tion. V.Quantitationof viral RNAsequencesin AD-2 infected and transormed cells. J. Mol. Biol. 106:749-771.
11.Flint,S.J., Y.Werwerka-Lutz,A.S.Levine, J.
Sam-brook,and P. A.Sharp.1975.Adenovirus
transcrip-tion.II.RNAsequencescomplementarytosimianvirus 40and adenovirus2DNA inAd2+NDr-and Ad2+ND3-infectedcells.J.Virol.16:662-673.
12.Gelinas, R.E., and R. J. Roberts.1977.Oneprominent 5'-undecanucleotide in adenovirus 2 late messenger RNAs.Cell11:533-544.
13.Goldberg, S.,J.NeAins,and J. E.Darnell. 1978. Evi-dence from UVtrnscriptionmappingthat late
adeno-virus type 2 mRNA is derived from alarge precursor molecule.J. Virol.25:806-810.
14.Goldberg,S., J. Weber, and J. E. Darnell. 1977. The definition of alarge viral transcription unit late in Ad2 infection of HeLacells:mapping by effect of ultraviolet irradiation. Cell 10:617-621.
15.Haas, M., M. Vogt, and R. Duibecco. 1972. Loss of simian virus 40 DNA-RNAhybrids fromnitrocellulose membranes; implications for the study ofvirus-host DNA interactions. Proc. Natl. Acad. Sci. U.S.A. 69:2160-2164.
16.Jelinek,W.,G.Molloy, R. Fernandez-Munoz, M.
Sal-ditt, and J. E. Darnell. 1974. Secondary structure in hnRNA:involvement of regions from repeated DNA sites. J. Mol. Biol.82:361-370.
17.Joho, R.H., M. A.Billetes, and C.Weisaman.1975. Mapping of biological function on RNA of avian tumor vinres:location of regions required fortransformation anddetermination of host range. Proc. Natl. Acad. Sci. U.S.A. 72:47724776.
18.Kennedy, S. I. T. 1976. Sequencerelationships between thegenome and theintracellularRNAspecies of stan-dard anddefective-interfering Semliki forestvirus.J. Mol.Biol. 108:491-512.
19. Klessig,D. F. 1977. Twoadenovirus messenger RNAs have a common 5'terminal leader sequence which is encodedatleasttenkilobasesupstream from the main codingregions for these messengers.Cell12:9-22. 20. Lewis, J. B., C. W. Anderson, J. F. Atkins, and R. F.
Gesteland.1974.Theorigin and destiny of adenovirus proteins. Cold Spring Harbor Symp. Quant. Biol. 39:581-590.
21. Lewis,J. R., J. F. Atkins, C. W. Anderson, P. R.
Baum,andR.F. Gesteland. 1975. Mapping of late adenovirusgenesby cell-free translation of RNA se-lected by hybridization to specific DNA fragments. Proc.Natl.Acad. Sci. U.S.A.72:1344-1348.
22. Lindberg, U., T. Persson, and L Philipson. 1972.
Isolation andcharacterizationofadenovirus messenger ribonucleic acid in productive infection. J. Virol. 10:909-919.
23. Loening,U. E.1967.The fractionationofhighmolecular weightribonucleicacidbypolyacrylamide gel
electro-phoresis.Biochem.J. 102:251-257.
24. McGrogan,M.,and IL J.Raskas.1977.Species identi-ficationand genomemappingofcytoplasmicadenovirus type 2RNAs synthesized late ininfection. J. Virol. 23:240-249.
25.Mautner,V.,J.Williams,J.Sambrook,P.A.Sharp, andT.Grodzicher. 1975.The location of the genes codingfor hexon and fiberproteinsinadenovirus DNA. Cell 5:93-99.
26. Meyer,J., P. D.Neuwald, S.-P. Lai,J. V.Maizel,and ELWestphal. 1977.Electronmicroscopyof late ade-novirus type2mRNAhybridizedtodouble-stranded viral DNA. J.Virol.21:1010-1018.
27. Molloy,G.R.,W.Jelinek,M.Salditt,and J. E.
Dar-nell. 1974. Arrangement ofspecific oligonucleotides within poly(A) terminated hnRNA molecules. Cell 1:43-53.
28.Moss, B., andF.Koczot.1976.Sequenceofmethylated nucleotides atthe 5'-terminus of adenovirus-specific RNA.J.Virol. 17:385-392.
29. Mulder,C.,J.R.Arand,W.Keller,U.Petterseon, R.J.Roberts,and P. A.Sharp.1974.Cleavagemaps of DNA from adenovirus types2and5byrestriction endonucleasesEcoRI andHpa1.ColdSpringHarbor Symp.Quant. Biol.39:397400.
30. Parsons,J.T.,J.Gardner,andM.Green.1971.
Bio-chemical studies on adenovirus multiplication. XIX. Resolutionoflate viral RNAspeciesinthe nucleus and
cytoplasm.Proc.Natl.Acad.Sci.U.S.A.68:557-560.
31. Pettersson,U.,C.Mulder,H.Delius,andP. A.Sharp.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL 25,1978
1973.Cleavage of AD-2 DNA into sixuniquefragments byendonuclease R.R1. Proc. Natl. Acad. Sci. U.S.A. 70:200-204.
32. Philipson, L, U. Petteron, U. Lindberg, C. Tib-betta, B. Venstrom, and T. Persson. 1974. RNA synthesis andprocesinginadenovirus infected cells. ColdSpringHarborSymp. Quant. Biol.39:447-456. 33.Philipson,L,R.Wall,G.Glickman,and J. E.Darnell.
1971.Addition ofpolyadenylatesequences to virus-spe-cific RNA during adenovirus infection. Proc. Natl. Acad.Sci.U.S.A. 68:2806-2809.
34.Sharp,P.A.,P. H.Gadlimore, and S. J. Flint. 1974. Mapping of adenovirus 2 RNA sequences inlytically infected cells and transformed cell lines. ColdSpring HarborSymp. Quant.BioL39:457-474.
35. Shatkin,A.J. 1976.Cappingofeukaryotic mRNAaCell 9:645-664.
36. Simmo D.T.,and J.H. Strauss.1974.Translation ofSindbis virus 26S RNA and 49S RNA inlysatesof rabbitreticulocytes.J.Mol. Biol. 86:397-409. 37. Sommers,S., MLSalditt-Georgieff,S.Bachenheimer,
J.E.Darnell,Y.Furuichi, AL Morgan,and A. J. Shatkin.1976.Themethylationofadenoviuns-specific nuclear and cytoplasmic RNA. Nuc. Acids Res. 3:749-766.
38. Stacey,D.W.,V. G.Affrey,andH. Hanasa 1977. Microinjection analysisofenvelopeglycoprotein mes-senger activitiesof avian leukosis viral RNAs Proc.
PROCESSING OF Ad2 mRNA's
823
Natl. Acad. Sci. U.S.A.74:1614-1618.
39. Tal, J., E. A. Craig, S. Zimmer, and H. J.Raskas. 1974.Localization ofadenovirus2 mRNAs tosegments of the viralgenome defined by endonuclease EcoR.R. Proc.Natl. Acad. Sci. U.S.A.71:4067-4061.
40.Thomas, AL, R. Iv White, and R. W. Davis. 1976. Hybridization of RNA to double-strandedDNA: for-mation of R-loops. Proc. Natl. Acad. Sci U.S.A. 73:2294-2298.
41. Vaughan,ML H.,J.R. Warner, and J. E. Darnell. 1967. Ribosomalprecursor particles in the HeLacell nucleus. J. Mol. Biol.25:235-251.
42.Wall, R., L Philipson, and J. E.Darnell. 1972. Proc-essing ofadenovirus-specific nuc!ear RNA during virus replication. Virology 50:27-34.
43.Weber, J., W.Jelinek,and J. E. Darnell. 1977. The definition of alargeviraltranscriptionunit late in AD-2infection of HeLacells: mapping of nascent RNA molecules labeled inisolatednuclei. Cell 10:612-617. 44.Wellauer, P. K., and I. B. Dawid. 1973. Secondary structuremaps ofRNA:processingofHeLaribosomal RNA. Proc. Natl. Acad. Sci.U.S.A. 70:2827-2831. 45.White,D.D.,MLD.Scharff,and J. V.MaizeL1969.
The polypeptides ofadenovirus. HI. Synthesis in in-fectedcells.Virology38:3956406.
46. Zi E.,and N.Fraser. 1978. Adenovirus type 2 late mRNA's:structuralevidence for3Tcoterminal species. J.Virol.25:897-906.

![FIG. 2.phoresisfilterscatingbeled Electropherograms of late rH]uridine-la- Ad2 RNA hybridized to (A) Sma I (36.7-40.5) and to (B) Sma D (40.5-52.6) filters](https://thumb-us.123doks.com/thumbv2/123dok_us/1532289.105831/4.501.309.408.57.546/fig-phoresisfilterscatingbeled-electropherograms-late-uridine-rna-hybridized-filters.webp)


![FIG. 7.filtersDceUsabeled(C 45 (5 and Electropherograms of late pH]uridine-la-Ad2RNA hybridized to Sma D (40.5-52.6) DNA (A and B) and to Eco F (70.7-75.9) DNA filters D)](https://thumb-us.123doks.com/thumbv2/123dok_us/1532289.105831/9.501.299.394.424.573/filtersdceusabeled-electropherograms-uridine-hybridized-dna-eco-dna-filters.webp)
