JOURNALOF VIROLOGY, Mar. 1975,p.645-653 Copyright 01975 AmericanSocietyforMicrobiology
Vol.15,No. 3 Printed in U.SA.
Formation of
Nucleoprotein Complexes
Between
Polyoma
Empty Capsids
and
DNA
H. VASKEN APOSHIAN,* RONALD E. THAYER, ANDPRADMAN K. QASBA'
Departmentof Cell Biology andPharmacology, University of Maryland School of Medicine, Baltimore, Maryland21201
Received forpublication18October1974
Purified polyoma empty capsids and polyoma type I DNA interact in a
cell-freesystem toformnucleoprotein complexes. Complexes that consist ofone, two, three, and four empty capsids per DNA molecule have been detected.
Polyoma
virions or capsomers do not react with added DNA to form such complexes.Productive infection
of mousecells
by
poly-oma virusyields mainly
complete virionsand
emptycapsids
(3, 26)having
densitiesin
CsCl
ofabout
1.32and
1.29g/cm3, respectively
(26).The production
ofother
polyoma-related
parti-cles has
been summarized elsewhere (28).
Thepurified
emptycapsids
arebelieved
tobe devoid
ofnucleic acid (2,
3,17). Whether
polyoma
emptycapsids
are precursorsof
complete
vi-rions,
by-products
of
virus
synthesis,
orthe
result
ofthe
disruption
of mature virionshas
neverbeen clarified.
In a
number
ofphage
systems,there
is increasingevidence
forthe interactions
ofviral
DNA and
emptycapsids. For
example, the
experimental results of Pruss
etal.
(21), using
the
phage P2-P4
system, arecompatible
with
DNA
packaging via the filling
ofpreformed
emptycapsids.
Hohn and Hohn
(11) have
shown that
empty,headlike
particles
of
phage
X areactive
inpackaging DNA
invitro. In the
caseof
phage T4,
atleast
someDNA
entersthe
phage head
afterthe
head has
been
constructed
(13).
Finally, experiments
by
Serwer (25)
indi-cate the formation in vivoand
in vitro ofcomplexes between phage T7 capsids and T7
DNA.
The evidence of this
paperindicates that
inacell-free
systempolyoma
emptycapsids and
polyoma
typeI DNA interact
to formcom-plexes.
Such
complexes
are notformed
between
polyoma virions and added polyoma DNA.
MATERIALS AND METHODS
Media. EC medium (18) was used with primary
mouse embryo cells. Dulbecco's modifiedEagle me-diumcontaining 10%calfserumwasused with baby
mousekidney cells.
' Present address: National CancerInstitute, National In-stitutes ofHealth, Bethesda,Md.20014.
Virions and empty capsids used for binding studies. Polyoma virions and empty capsids used in the bindingexperiments wereproduced byinfecting primary mouse embryo cells, at a multiplicity of infection ofabout 1 PFU/cell, with thislaboratory's
stock of the smallplaquevariantof the Toronto strain
of polyoma virus. The method of Crawford (1) was
used for virus production and harvesting.
Toobtain radioactive virions and emptycapsids,10
,gCi
ofL-[fH
]tryptophan
(3Ci/mmol)wereadded perml of medium at 18 h postinfection. The medium, after infection, contained 1% dialyzed calf serum. Virions and empty capsidslabeled with
[3H jarginine
wereproduced by adding to the cells, after the virus adsorption period, medium containing citrulline and
1%dialyzed calf serum butlackingarginine. L- ['H Jar-ginine (20.6Ci/mmol)wasadded 20 h after infection. The viruswasharvested 6 days postinfection.
The crudeextracts werepurified essentiallybythe method of Winocour(26). The peak of virions and the peak of empty capsids obtained after equilibrium centrifugation in CsCl were each subjected to two
additional CsCl equilibrium centrifugations. The finalpeak of virionsoremptycapsidswasseparately dialyzed, sedimented through the 10to40%standard
sucrose gradient, and dialyzed against 0.01 M Tris (pH7.5)-0.05 M NaCl. Virionsoremptycapsidswere
located in all of the abovegradients byradioactivity and/or hemagglutination assays. The nonradioactive polyoma empty capsids had 44 hemagglutination
units/gg
ofprotein.Preparation of radioactive polyoma DNA. Ra-dioactive DNA was obtained by adding [4C
Ithymi-dine (56mCi/mmol) or [3H]thymidine (20Ci/mmol)
to babymousekidney cells 18 h after infection with
the large plaque strain of polyoma virus. Polyoma DNAwasextracted from the cells48h afterinfection, using the method ofHirt(9). The DNAwaspurified with phenol, precipitated with alcohol, and passed through twocyclesofcentrifugation in cesium chlo-ride-ethidium bromidegradients(23). The peak
con-taining polyoma typeI DNAwaspooled. The ethid-ium bromide was removed by four extractions with isopropanol and dialysis against0.1xstandard
saline-645
on November 10, 2019 by guest
http://jvi.asm.org/
APOSHIAN,THAYER, ANDQASBA
citrate. The DNA solution was dialyzed finally against 0.02 M Tris (pH 7.5)-0.02 M NaCl and had a 280:260absorbance ratio of 0.538.
Standard assay method for measuring complex formation. The assay iscarried out in a glass tube, which has been treated with Siliclad (Clay Adams Co.) 1 h before use. The standard reaction mixture (0.20 ml) contains 1.5 x 10-2 M Trisbuffer (pH 7.5), 10-2MMgCl2,0.10 mg ofcrystalline bovine albumin (Pentex), 0.61
lAg
ofpolyoma type I[3H
]DNA (3.48 x 10' counts/min perjg),
and designated amounts of purified polyoma empty capsid protein. (Pentex crys-talline bovine albumin was usedbecause it is free of endonuclease activity. Two different lots of Armour crystallizedbovine albumin were found to contain an endonuclease activity that converts polyoma DNA from 53S to 14 to 16S material.) After incubation at 37Cfor60min, the reaction mixture is chilledto 4C for 10 minand transferred to the top of the standard sucrosegradient at4C in aSpincoSW41 polyallomer tube. Thestandard
sucrosegradient consists of10ml ofaneutral 10 to 40% sucrosegradientcontaining0.01 MTris (pH 7.5)-0.15 M NaCland 0.5 mg of crystal-line bovine albumin (Pentex) per ml. The sucrose gradient is preformedover0.30ml ofaCsClcushion having a density of 1.77 g/ml. Sedimentation is carried out in theSpincoSW41rotor at 4C for2hat25,000 rpm. After centrifugation, the bottom ofthe tube is punctured, and 20drops/fractionarecollected directly into plastic scintillation vials. Scintillation fluid containing Triton X-100 is added, and the radioactivity in each fraction is determined with a PackardTri-Carbscintillation counter.
Gradients. In addition to the standard sucrose
gradient, described above, the following gradients were used. (i) CsCl isopycnic gradients (26) were performed at 32,000 rpm for 22 h at 25C with a Spinco SW50.1 rotor. (ii) Alkaline 5 to 20% sucrose
gradients were prepared by mixing in a Buchler gradient-forming apparatus 2.3 ml of 5% sucrose containing 0.1 NNaOH-0.5 MNaCl-0.025M EDTA and 2.3 ml of 20% sucrose containing 0.4 N NaOH-0.5 M NaCl-0.025 M EDTA. Centrifugation was for 115 min at49,000 rpm, 4C, in the Spinco SW50.1 rotor. (iii) All gradients were fractionatedby puncturing the bottomof thetubes andcollectingdrops. The radioac-tivity in each fractionwasdeterminedwithaPackard Tri-Carb scintillation counter by using a toluene-based scintillation fluid which contained 0.55% 2,5-diphenyloxazole, 0.01% 1,4-bis[2-(4-methyl-5-phen-yloxazolyl)
]-benzene,
10% water, and 33% Triton X-100(Rohm and Haas).Other methods. Hemagglutination assays were
performed using guinea pig erythrocytes (1). Protein
wasdetermined by the method of Lowry et al. (12). Concentrationsofpolyoma DNA were determined by ultraviolet absorption, using the extinction coefficient E,,%=200. Electrophoresis was doneby the method of
Maizel (14), using 0.1% sodium dodecyl sulfate-7.5% polyacrylamide gels, 7.5 cm in length. Virions or empty capsids were incubated in a previously de-scribed,sodiumdodecyl sulfate-containing disruption bufferfor 2hat 55Cbefore electrophoresis (14).
Other materials. The following materials were obtained from the listed sources: [sH]thymidine (20 Ci/mmol), New England Nuclear; [14C ]thymidine (116mCi/mmol),Mallinckrodt,or
[14C
Jthymidine (56 mCi/mmol), New England Nuclear; Pronase (nu-cleasefree), Calbiochem; pancreatic DNase and pan-creatic RNase, Worthington Biochemicals; Pentex bovine albumin, crystallized, Miles Laboratories; L-[8HJtryptophan (3 Ci/mmol), Schwarz-Mann; L-[3H]arginine (20.6 Ci/mmol) and L-[14C ]arginine (0.313Ci/mmol), New England Nuclear.RESULTS
Criteria of purification. The two principal criteria usedto indicate thedegree of purity of the emptycapsids were gel electrophoresis and sedimentation throughsucrose. Gel electropho-resis of purified preparations of empty capsids (Fig. 1A) indicated the presence of polyoma polypeptides P1, P2, P3, and P4 and the ab-senceof detectable amounts ofP5, P6, and P7. Thenumbering system of Roblinetal. (24) has been used to designate the polypeptides. Poly-peptides P5, P6, and P7 are "internal proteins,"
which
containarginine but lack tryptophan
(4, 24). They are believed to represent host cellhistones,
probably
bound topolyoma DNA,
in the virions (4, 24). P5, P6, and P7 representabout
15 to 20% of theradioactivity
found in arginine-labeled virions (Fig. 1C; ref. 24). Fif-teen percent of the 12,000counts/min
recovered in the analysis of the[I-H]arginine-empty
cap-sids
ofFig. 1A would amount to1,800
counts/
min. If
significant
amounts ofP5, P6,
and P7 werepresent in the [3H]arginine-empty
capsids,
they
would beexpected
infractions 40to 46 ofFig.
1A.However,
fractions 40 to 46 do not represent adiscretepeak
andcollectively
repre-sentonly 60counts/min
or 0.5% of the recovered countsper minute.The [3H]tryptophan-labeled
emptycapsidsalsodonotappeartobe
contami-nated with
significant
amounts of [3H ]nonviral protein (Fig. 1B).Sedimentation of
purified
radioactiveempty capsid preparations through the standard su-crose gradient indicates that virtually all theradioactivity
is inthemajorpeak (Fig. 2).
Whencompared
to polyoma virions (240S), added as asedimentation
marker,
the emptycapsids
sediment at 189S. Otherinvestigators
havereported
values of 140 and134S
forpolyoma
emptycapsids
(2,17).
Either ouremptycapsids
areaggregated
orthey
containsubstantial
im-purities bound
tothecapsids.
The electrophore-sis results(Fig.
1)
indicate that the latterpossibility
isunlikely. On the
otherhand,
the140S value reported
foremptycapsids
by
others646 J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
POLYOMA EMPTYCAPSID-DNACOMPLEXES
30
o.1
_
I0
647
C. P2
P4
La t5
Pi P3
0 10 20 30 40 50 0 10 20 30 40 50 0 10 20 30 40 50 FRACTION FRACTION FRACTION
FIG. 1. Sodium dodecylsulfate-polyacrylamide gel electrophoresis of purified polyomaemptycapsids and virions. (A) [3H]arginine-empty capsids; (B) [3H]tryptophan-empty capsids; (C) ["4C]arginine virions.
3
2
10
E
UL
1
10 20 30 40
Fraction
FIG. 2. Sedimentationofpurified [3H]arginine
pol-yomaemptycapsids in the standardsucrosegradient.
When[14C polyomavirionsareaddedas a
sedimenta-tion marker (240S), the peak of
[14C]radioactivity
is atfraction 23.may be due to the presence of DNA bound to
theempty capsids.
Increase in the sedimentation ofpolyoma type I DNA after incubation with polyoma empty capsids. The incubation of polyoma emptycapsids with radioactive polyomatype I
DNAcauses amarkedchangeinthe
sedimenta-tion behavior of the DNA (Fig. 3). When the DNA is incubated in the absence of theempty capsids, all of the radioactivity remains at the
top of the sucrose gradient (Fig. 3A). The
incubation of the DNA with nonradioactive polyoma empty capsids results in the
appear-ance ofanumber ofnew peaks of radioactivity
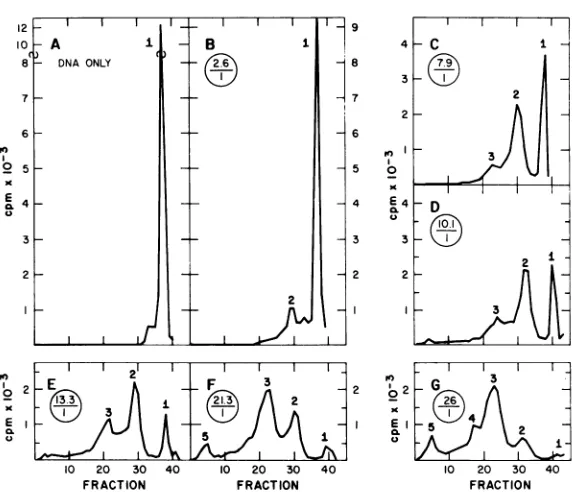
(Fig. 3B-G). At a low ratio of protein toDNA (Fig. 3B), a single new peak appears. As the
protein-to-DNA ratio increases to about 13:1, the radioactivity of peak 1 decreases, that of peak 2 increases, andathirdnewpeakappears. Eventually, as peak 3 increases, peak 2
de-creasesandtwonewpeaks,4 and5, appear(Fig. 3F andG). Further characterizations ofsomeof thesepeaks will be described later in thispaper. When radioactive empty capsidsare incubated
inthe standard reaction mixture without DNA,
aprofile identicalto that of Fig. 2 is seen.
The addition of either 7 or lOS polyoma
capsomers, intheplace ofemptycapsids,tothe standard reaction mixtureresulted in all of the radioactive DNA remaining at the top of the standard sucrose gradient. The profiles of the
gradients were identical to that of Fig. 3A. Furthermore, theaddition ofcapsomersto
reac-tion mixtures containing empty capsids and DNAdidnotinhibittheformation of theempty
capsid-DNA complexes. The 7 and lOS
capso-mers wereprepared and purified by the method
ofFriedmannand David (6),exceptthat capso-mers in thepresentexperimentswereprepared
frompurifiedemptycapsids instead of polyoma
virions.
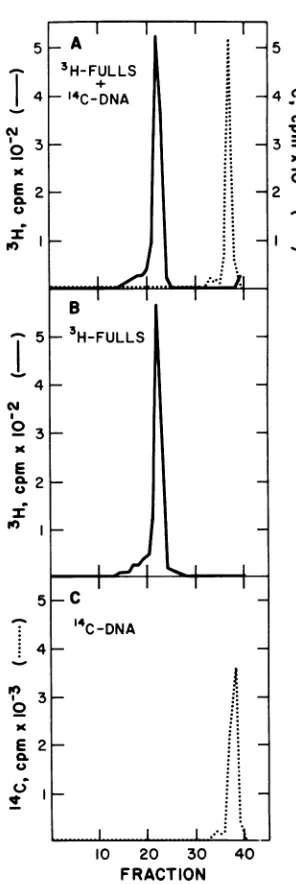
Failure ofpolyoma virions to change the sedimentation behavior ofnon-encapsidated DNA. Figure 4A presents thesucrose gradient
profile obtained after incubation of [3lH]trypto-phan polyoma virions (fulls) with polyomatype
VOL. 15,1975
30 25 20
'.
10
05
A.
P2
Pi
p3 P4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.105.403.66.227.2] [image:3.504.51.244.261.522.2]APOSHIAN, THAYER, ANDQASBA
9
4
8 3
7
2 6
IX
5 O
x
4 E4
3 3
2 2
"I
2 2
E
cx
J.VIROL.
10 20 30 40 10 20 30 40 10 20 30 40
FRACTION FRACTION FRACTION
FIG. 3. Change in the sedimentation ofpolyoma type I [8HIDNA after incubation with polyoma empty capsids and the influence of the ratio ofproteintoDNA concentrationson the reaction. Thestandardassay
method withtypeI[9H1DNAwasused. In the reaction ofA,emptycapsidswereomitted. The reactions of B, C,
D, E, F, and G had1.6, 4.8, 6.5, 8.1, 13, and16,ug,respectively. Theencircled numbersaretheprotein-DNA
ratiosineach reaction mixture. Recoveries of radioactivity from thesegradients and others in thispaper were greaterthan96%,unlessotherwise stated.Direction of sedimentationisfrom righttoleft in this and all figures.
I ["4C
]DNA.
The single peak of [3H ]radioactiv-ity that is found doesnotcontainany[14C
]DNA radioactivity. Unlike polyoma empty capsids, full capsids do not change the sedimentation behavior of added DNA.The addition of [3H ]tryptophan virions to a
reaction mixture containing nonradioactive empty capsids and type I [4C ]DNA did not change the multipeaked sedimentation profile that is observed when empty capsids and
["4C
]DNAare incubatedtogether. Suchmixingexperiments indicate that the failure of virions to react with addedpolyomaDNA is notdueto the presence ofsomeinhibitory factor(s) in the
virion preparation.
Characterization oftheDNA-empty capsid complex. The approximate Svalues ofpeaks2 and 3 (Fig. 3) formed in the reaction between polyoma type I [3H]DNA andunlabeled empty
capsids were determined by the method of
Martin and Ames (15). As a sedimentation
marker,
[14C
polyoma virus (240S) was added to the reaction mixture immediately prior tosedimentation in thestandardsucrosegradient.
The approximate S value of peak 2was 144S.
Peak 3 cosedimented exactly with the 240S marker.
When the 144S complex formed between [3H ]arginine-empty capsids and
[4C
]type I DNA is isolated from the initial sucrosegradi-ent and either diluted in or dialyzed against
0.01 M Tris (pH 7.5)-0.15 M NaCl, resedimen-tation in the standard sucrose gradient results
intherecoveryof all the 3H and 14C
radioactiv-ity in asingle peak sedimenting at 144S.
Simi-larly, dilution or dialysis of the 240S complex
results in asingle peak sedimenting at
240S.
The 144Sproduct of the reaction contains the polyoma polypeptides found in polyomaempty capsids, indicating that the reaction involves capsid and notnoncapsid protein (Fig. 5). The 14C peaks near the origin represent the["4C ]DNA thatwas in thecomplex.
Theprotein and DNAcontentsofsomeof the
complexes formed were calculated using the
specific activity of the ["4C ]DNA and 3Hempty capsids (Table 1). The peak fraction of the144S complex is foundtocontain the leastamountof protein relative to DNA, whereas the 370S complex contains four times asmuch. Table 2
summarizes experiments using three different preparations of 3H emptycapsidstoobtain the percent DNA and protein-DNA ratio of the 144S complex.
648
12
10
8
7
6
t5
3
2
to
E
- A i _ B I
- DNAONLY
2
II
3 Ii
I
-1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.119.405.69.315.2]POLYOMAEMPTY CAPSID-DNA COMPLEXES
Examination
by electron microscopy of the
reactionmixture, after
incubation but without
sedimentation
through
sucrose,shows the
pres-ence ofemptycapsid-DNA
complexes consist-ing of one, two, three, and four empty capsids permolecule
ofDNA. When the 144, 240, and320S
complexes are isolated from the sucrose gradient, electron microscopy showed there were one, two, and three empty capsids,respec-tively,
perDNA molecule.
Complexes
contain-ing one, two, or three empty capsids per DNA molecule are seen in Fig. 6. Nocomplexes
are seen if the DNA or the empty capsids areomitted
fromthe reaction.The DNA in the
144and
240S complexes
is not protected fromhydrolysis by
pancreatic DNase (Fig. 7). After complexesbetween
emptycapsids and DNA
areformed,
incubation
ofthem with nuclease-free Pronase destroys them
(Fig.
8).They
arealso dissociated
by
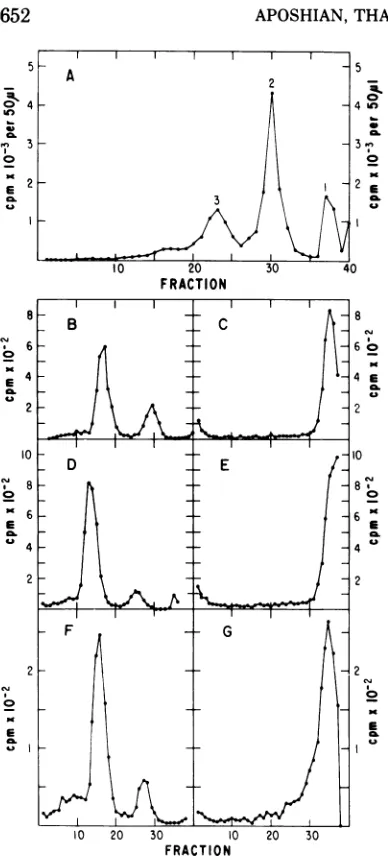
1M NaCl.Determination of the ratio of polyoma type
I
DNA to types II plus III DNA after
incuba-tion with
polyoma empty
capsids. After
incu-bation in the standard
assayreaction
mixture,
followed
by sedimentation through alkaline
su-crose,89% of
type I DNAand
11% of typesII
plus
III werefound
tobe
present(Fig.
9).Since
the
same percentages wereobserved
forthe
DNA
prior toincubation (Fig. 9B),
itis
con-cluded
that whenthese
purified
emptycapsids
areincubated with
type IDNA
anendonu-cleolytic
cleavage
of type IDNA does
not occur.Miscellaneous
agents and their effects. The
presence of 0.50 MNaCl
during the reactioncompletely
preventsthe reaction
between
type IDNA and
emptycapsids. No
inhibition
isdetected
inthe
presence of up to 0.25M
NaCl.
The
appearance ofpeaks
2, 3, 4,and
5ofFig.
3 is preventedby
either pretreating the type I[3H
]DNA with pancreatic DNase
orpretreating
the
purified
emptycapsids with nuclease-free
Pronase. Purified
emptycapsids treated with
RNase and
DNaseand then
repurified
still
reactwith
polyoma
DNA.Mercaptoethanol
(0.5
x 10-3M),
EDTA
(0.04
M),
orpreincubation
of type I[3H]DNA with
pancreatic RNase did
notaffect the
appearance ofpeaks
2,3,
4, or 5of
Fig.
3.When
["C
]dTMP
wasused
inthe standard
reaction mixture in place of radioactive poly-oma type I DNA,all
ofthe radioactivity
re-mained at the very top of the sucrosegradient
and there was nodetectable
binding
of the[14C
]dTMP
tothepurified
emptycapsids.
DISCUSSION
These
experiments
demonstrate that poly-omaDNA andpolyoma
emptycapsids
interact5
14
N
6
3x
I'
14
02
3
£'
I0
toI
5
3 10
2
II A 3H-FULLS
-14C-DA-DNA
B 3H-FULLS
i
C-'4C-DNA
i
- 1 .10 20 30
FRACTION
40
FIG. 4. Incubation of polyoma type I DNA with polyoma virions does not change its sedimentation behavior. The standard assay method was used, except emptycapsidswereomitted. Inthe reaction of
A, [3H]tryptophanvirions("fulls"),8.1ug,andtypeI
["CIDNA (1.18 x 10' counts/minperug), 0.62 Mg,
wereused; B, ["4C]DNAwasomitted;C, virionswere omitted. Thespecific activity ofthevirions was1.63
x 102counts/minperAgof proteinand 74
hemaggluti-nationunits/agof protein.
to form complexes. At a low ratio of empty
capsidproteintoDNA,a1:1complexis formed.
As the ratio increases, two capsids per DNA molecule and three capsids perDNA molecule
arefound.There isaspecificitytothereaction;
VOL. 15, 1975
649
5
0
3 3
x
5
2 i,,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.272.420.65.507.2]APOSHIAN, THAYER, AND QASBA
E
0.
u
0 20 30 40 50
FRACTION
FiG. 5. Polyacrylamide gel electrophoresis of the 144S complex formed between 'Hemptycapsids andtypeI ["4CJDNA. (Left panel) Standard assay method except that the reaction mixture contained 7.2 Mig of
['HJarginine-empty capsids (2.25 x 10' counts/minper Mg) and 0.74 Mg of type I ["4CJDNA (1.04 x 10'
counts/minper Mg). The 144S peakwaspooled, ["4C]arginine polyoma virionswereaddedasamarker, and the
proteinswereprecipitated with10%otrichloroacetic acid. The precipitatewaswashedwithacetoneandanalyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. (Right panel) As above,exceptreaction mixture contained 10 Mg of [3H]tryptophan-empty capsids (3.14 x 10' counts/minper MAg) and 0.74 Ag oftype I
[image:6.504.121.407.60.258.2][14C DNA.
TABLE 1. Protein and DNA analysis of complexes formedbetween empty capsidsand type I DNAa
ompex
D~PrtenA
EstimatedComplmx'
DNAc
DNA amt ofcapsiDs
(%oftotal) raid poen perDNA
molecule
144S 10.7 8.3 (1) 1
240S 6.3 14.9 1.8 2
320S 3.7 25.9 3.1 3
370S 3.0 32.8 4.0 4
aThestandard assay methodwasused, except that
each collected fraction of the sucrose gradient
con-tained10drops.The reaction contained
['H
Jarginine-emptycapsids,7.2ug(1.85 x 10'counts/min
perjg
of protein), and type I [4C]DNA,
0.74 ,ug (1.01 x 104counts/minperjAg).Recoveries of radioactivitieswere
90% for "ICand 88% for IH.The values in thetables
are for the peak fraction of each of the complexes collected from the sucrosegradient.
bS values were determined insucrose.
c(Micrograms ofDNA/microgramsofDNA +
mi-crograms ofprotein) x 100.
dMicrogramsofprotein/microgramsofDNA.
that is, the added
DNAcombines
with emptycapsids but
not withcomplete
virusparticles.
We do
notwish
toimply
aspecificity
for the type ofDNA, since
polyoma
types I or IIplus
IIIDNA,
aswell
as mouseDNA,
bind the emptyTABLE 2. Relative content ofproteinand DNAin the DNA-empty capsid144Scomplexa
Empty capsids 1 DNA Protein-DNA Expt labeled with (%)' ratioc
1 [3H]tryptophan 9.4 9.6
2 [SH ]tryptophan 10.6 9.4
3 [SH
]arginine
10.2 8.7aThe standard assay method was used. In each
experiment, 0.74 ug of polyoma type I
["4C
]DNAwasused. The amounts of empty
'H
capsids used in experiments 1, 2, and 3 were 6.2, 10.0, and 7.2Ag,
respectively.
b,C Calculatedasshown inlegendtoTable1.These values are basedon all thefractions obtained in the 144S peak of the gradients.
capsids (Aposhian
and Thayer,unpublished
data). However,
itshould
be remembered that
pseudovirions,
which contain mouse DNA frag-ments,also
areproduced
during productiveinfection
by polyoma
virus (16, 22, 27).The
results of theseexperiments
suggest thatthe
polyoma
emptycapsids
contain somesite(s)which has
an affinity for DNA. These sites do not appear to exist on the outer surface ofcomplete
virionsand, therefore,
probably
may notbe
onthe
outer surface ofthe emptycapsids.Rather,
thesite(s)
maybe
attached to orpart of650
J.VIROL.P2'
0
9'+
84
7'1
6'45
,I-4,
21
I P3 P4P5-7
&O'
10
9
8 7
cl-6
6
5E
0.
4..Cs, 3 2 0
FRACTION
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.504.63.255.353.467.2] [image:6.504.265.458.354.437.2]POLYOMA EMPTY CAPSID-DNA COMPLEXES
the inner surface of the
emptycapsids
or,alternatively,
at apossible point
of
incomplete-ness
of
someof
the
emptycapsids.
However,
we do not know of anypublished
evidence in
support ofsuch
apostulated
incompleteness.
The
possibility
that such
sites are present onthe
outer surface ofthe
complete virions,
but
areoccupied
already
by
somenucleic
acid,
appears
unlikely.
The
virionshave
been
par-tially purified
by
atleast three
CsCl
equilib-rium
centrifugations. The
DNA-empty
capsid
complex is dissociated
insuch concentrations of
.CsCl
(Aposhian,
unpublished data).
It
seemsreasonable
to assumethat
anycomplex
formed
between
unencapsidated
nucleic acid and the
possible sites, if
they did exist
onthe
outersurface
ofvirions, also would be dissociated and
separated
in
CsCl.
Also,
empty
capsids
purified
from
cells
that
werelabeled before
or afterinfection with radioactive
thymidine
werefound
tobe free
ofradioactivity.
Although
wehave
been
able
todetect,
by
the
presentmethods,
only
polypeptides P1,
P2,
P3,
and P4
inthe
purified
preparations of empty
capsids
and
inthe
DNA-empty
capsid
complex,
we areaware,of course,
that
as yet no onehas
been able
topurify
polyoma
virions
absolutely
freeof
tracesof
contaminating proteins.
Ho et
al.
(10)
have
reported
that
polyoma
empty
capsids
do
notappear
to reactwith
mouse[3H
]DNA,
asjudged
by
the failure
oftheDNA
toreduce the
hemagglutination
activity
of
the
emptycapsids and
by the failure
to precipi-tateradioactivity after
mixing [3H
]DNA,
empty
capsids, and
polyoma
virusantiserum. Hoetal. (10) used very small ratios ofprotein and DNA
(0.02:1 orless).
Complex
formationwould
notbe expected
to occur atsuch
small ratios(Fig.
3).Friedmann
(5)
has
disrupted polyoma
virions tocapsomers and DNA
by
using carbonate
buffer
(pH 10.6)and dithiothreitol.
Attempts
(5) atreassembly of the disrupted virus
by
overnight
dialysis of the preparations resulted
inthe formation
of a100S DNA-protein
com-plex consisting
mostly of linear
aggregates ofcapsomers,
indicating
toFriedmann
that
poly-omacapsomers
bind polyoma DNA. Our
experi-mentsdemonstrate that
purified,
naturally
oc-curring
emptypolyoma capsids also bind
poly-oma
DNA.
We do
notknow
at presentwhether the
formation
ofthese
emptycapsid-DNA
com-plexes will
be useful
as amodel
system forthe
study
ofthe
assembly
ofpolyoma virions.
How-ever,experiments
are in progress todetermine
whether the 55S polyoma nucleoprotein
com-plex, isolated
by
Green
etal.
(8) and
Goldstein
etal.
(7), also
binds
polyoma empty
capsids.
There
isevidence supporting
aprecursor role for emptycapsids
inthe
maturation of another papovavirus,
simian virus40,
as hasbeen
2
;
,'
> 4, ' 4J4 4 r ..
X,>t
¢,X@A:U^i¢8~4
Wat7S~~~~~~4
.EL ,4.C, 'I
;,*
; tb+X
.. lb.
;=,;,,; t.v.*,-.x !
-F | t > ^ * ^ ' Ja | ' ; \tC^i;>-<-~~~4'
, -,x -ieP '9>|
a' 5;r * s
444.~~~~~~r
`5jf 'tS e t' '|aX,'
'it~~~~~~~~~~~~~i
4.
FIG. 6. Complexesof polyomaemptycapsidsandpolyomaDNAformedin the standard reactionmixtureas seen byelectronmicroscopy.Reaction mixture contained type IDNA,0.80Ag, and12.0lAg ofemptycapsids. Protein monolayer technique was used.Magnification, 72,000.
VOL.15, 1975
651
on November 10, 2019 by guest
http://jvi.asm.org/
APOSHIAN, THAYER, AND QASBA
In
0.
E
0
E
cq
6
Ei
E
0
k-)
4In
ion
6
06
FRACTION
FIG. 7. Sensitivity ofthe DNA in theDNA-empty capsid complex topancreatic DNase. (A) The
stan-dard assay method containing 11.2 ug of polyoma
empty capsid protein and 0.68 Mlgof polyoma typeI [8HIDNA wasused. Thefractions from thestandard
sucrose gradient were collected inplastic minivials
and 50gliters ofeach fractionwasplaced on
What-man GF/A glass filterpaper. Thefilters were dried
andplacedinplastic countingvials. Toluene scintilla-tion fluidwas added and thesamples werecounted.
(B and C) A 50-Mliter amount of fraction 37ofthe abovegradientwasplaced in each oftwo test tubes.
To one ofthe test tubes (C) were addedMgCI2, to giveafinal concentration of5x 10-3M, and pancre-atic DNase (1
gg/ml).
MgCI2 and DNase were notadded to the other tube (B). The tubes were
incu-batedfor15minat37C andthenchilledat4 C. To
each tubewereadded 0.22 ml of2.7 x 10-2MNaK
10 20 30 4
E5 4
3
2
0
10 20 30 40
FRACTION
FIG. 8. Digestionof the capsid-DNA complexes by Pronase. The standard assay method was used, except thatserum albuminwasomitted from the incubation mixture and the gradient. The reaction mixture contained 11.2 gg of polyoma empty capsid protein and0.68ugof polyoma type I[3HJDNA.After 60 min at 37C, the reaction mixture was divided into ali-quotsof 0.10ml. To onealiquot nuclease-free Pronase (100ug/ml)was added. Each aliquot was incubated at
37Cforanadditional30minand thenplacedonthe standard sucrose gradient. Symbols: With Pronase, (0); without Pronase, (a).
pointed
outby Ozer and Tegtmeyer
(19,20).
The
observation that polyoma
emptycapsids
formcomplexes
withpolyoma
DNAraises
anumber
ofother
interesting
questions. Do the emptycapsids
protectthe DNA
inthe
complex
fromthe action
ofbacterial
restrictionen-zymes?
Arethe
emptycapsids attached
tospecific
segments ofpolyoma DNA? These
questions
arebeing investigated.
ACKNOWLEDGMENTS
Wearegratefuland indebted toThomas J. Kelly, Jr., for the electron microscopy. We thank Maybritt Doelp for excellent technical assistance.
This workwassupportedbyPublic Health Service grants GM-19209 from the National Institute of General Medical Sciences and CA-10497 from the NationalCancerInstitute, andbyagrantfromthe JohnA.HartfordFoundation, Inc.
EDTA and 0.03 ml of 1 NNaOH. The contents of
each tube were quantitatively transferred to the top
ofanalkaline 5 to 20%sucrosegradientand centri-fuged. Fractions (7 drops) werecollected directly in
plasticscintillation vials and counted in scintillation
fluid containing Triton 100-X. (DandE) SameasB
and C, except that 50,Lliters of fraction 30, instead
offraction 37,wasused.(FandG)SameasB andC except that 50 ,uliters of fraction 23, insteadof
frac-tion37,wasused.
FRACTION
652 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.64.258.52.483.2] [image:8.504.269.459.60.237.2]POLYOMA EMPTYCAPSID-DNA COMPLEXES
20 30 40 10
FRACTION
FIG. 9. Determination of theratio of polyomatype I DNA to typesII plus III DNA after incubation with polyoma emptycapsids. The standardassaymethod
was used. In each experiment, 0.61 ug of polyoma [3H]DNA typeI(3.43 x 104counts/minpergg)and 6.5 ugof polyoma empty capsid protein were used.
Afterincubation for1hat37C, the reaction mixture
was chilled to 0C and 0.02 ml of0.2MEDTA and 0.025 mlof1N NaOHwereadded withgentle mixing.
Then 0.20 ml of the mixture was placed on a
pre-formed alkaline 5 to 20%sucrosegradient and
cen-trifuged. Each fraction contained 7drops. (A) after incubation; (B) before incubation.
LITERATURE CITED
1. Crawford, L. V. 1962. The adsorption of polyoma virus. Virology18:177-181.
2. Crawford, L. V., and E. M. Crawford. 1963. A
compara-tivestudy of polyoma and papillomaviruses.Virology
21:258-263.
3. Crawford, L. V., E. M. Crawford, and D. H. Watson.
1962. Thephysical characteristics of polyoma virus.I. Two typesofparticle. Virology 18:170-176.
4. Frearson, P.M., andL. V.Crawford. 1972.Polyomavirus
basicproteins. J. Gen. Virol. 14:141-155.
5.Friedmann, T. 1971. In vitro reassembly of shell-like particles from disrupted polyoma virus. Proc. Nat.
Acad.Sci. U.S.A. 68:2574-2578.
6.Friedmann, T., and D. David. 1972.Structural roles of
polyomavirusproteins.J.Virol. 10:776-782. 7.Goldstein, D. A., M. R. Hall, and W. Meinke. 1973.
Propertiesofnucleoprotein complexes containing repli-cating polyomaDNA. J.Virol.12:887-900.
8. Green, M. H., H. I. Miller, and S. Hendler. 1971. Isolation of a polyoma-nucleoprotein complex from
infected mouse-cell cultures. Proc. Nat. Acad. Sci.
U.S.A. 68:1032-1036.
9. Hirt,B.1967. Selective extraction ofpolyomaDNA from
infectedmousecell cultures.J. Mol.Biol. 26:365-369.
10. Ho, J. K., G. Y. Chan, and J. C. Chan. 1970.Lackof
interaction between polyoma virus capsid andmouse
cell DNA in vitro. Life Sciences 9:1255-1260.
11. Hohn, B., and T. Hohn. 1974. Activity ofempty,headlike particles for packaging of DNA of bacteriophagesAin
vitro.Proc.Nat. Acad. Sci. U.S.A. 71:2372-2376.
12. Lowry, 0. H., N. J.Rosebrough, A. L. Farr, and R. J.
Randall. 1951. Protein measurement with the Folin phenolreagent.J. Biol. Chem. 193:265-275.
13. Luftig, R. B., W. B. Wood, and R. Okinaka. 1971. Bacteriophage T4 head morphogenesis. On thenature
ofgene49-defective heads and their roleas
intermedi-ates.J.Mol. Biol. 57:555-573.
14. Maizel, J. V., Jr. 1971.Polyacrylamide gel electrophoresis ofviralproteins, p. 179-246.In K. Maramoroschand
H. Koprowski (ed.), Methods in virology, vol. V.
AcademicPress Inc., New York.
15. Martin, R. G., and B. N. Ames. 1961. A method for determining the sedimentation behavior ofenzymes:
application to protein mixtures. J. Biol. Chem. 236:1372-1379.
16. Michel, M. R., B. Hirt, and R. Weil. 1967. Mouse cellular DNA enclosed in polyoma viral capsids (pseudovir-ions). Proc. Nat. Acad. Sci. U.S.A. 58:1381-1388.
17. Murakami, W. T., R. Fine, M. R. Harrington, and Z. Ben Sassan.1968.Properties and amino acid compositionof
polyomaviruspurified by zonal ultracentrifugation. J. Mol. Biol. 36:153-166.
18. Osterman, J. V., A. Waddell,and H. V.Aposhian. 1970. DNA andgenetherapy: uncoating of polyoma pseudo-virus in mouseembryo cells. Proc. Nat. Acad. Sci.
U.S.A.67:37-40.
19. Ozer, H. L.1972.Synthesis and assembly ofsimian virus 40. I.Differentialsynthesis of intact virions andempty
shells. J. Virol. 9:41-51.
20. Ozer, H. L., and P. Tegtmeyer. 1972. Synthesis and assembly of simian virus 40. II. Synthesis of the major capsid protein and its incorporation into viral particles. J. Virol. 9:52-60.
21. Pruss, G., R. N. Goldstein, and R. Calendar. 1974. In vitropackaging of satellite phage P4 DNA. Proc. Nat. Acad. Sci.U.S.A. 71:2367-2371.
22. Qasba, P. K., and H. V.Aposhian. 1971. DNA andgene
therapy: transfer ofmouseDNAtohuman andmouse
embryonic cells by polyoma pseudovirion. Proc. Nat. Acad. Sci.U.S.A.68:2345-2349.
23. Radloff, R., W. Bauer, and J. Vinograd. 1967. A dye-buoyant-density method for the detection and isolation ofclosedcircular duplex DNA: the closed circular DNA in HeLa cells. Proc. Nat. Acad. Sci. U.S.A. 57:1514-1521.
24. Roblin, R., E. Harle, and R. Dulbecco. 1971.Polyoma virus proteins. I. Multiple virionscomponents.Virology 45:555-566.
25. Serwer, P. 1974.Complexes between bacteriophage T7 capsids and T7 DNA. Virology 59:89-107.
26. Winocour, E. 1963.Purification ofpolyoma virus.
Virol-ogy19:158-168.
27. Winocour, E. 1968. Further studiesontheincorporation
ofcell DNA into polyoma-related particles. Virology
34:571-582.
28. Yelton, D. B., and H. V. Aposhian. 1972. Polyoma pseudovirions.I.Sequenceofeventsinprimarymouse
embryo cells leading to pseudovirus production. J.
Virol. 10:340-346.
VOL.15, 1975
![FIG. 2.yomaattionWhen fraction Sedimentation ofpurified [3H]arginine pol- empty capsids in the standard sucrose gradient](https://thumb-us.123doks.com/thumbv2/123dok_us/1575706.110160/3.504.105.403.66.227/yomaattionwhen-fraction-sedimentation-ofpurified-arginine-capsids-standard-gradient.webp)

![FIG. 9.polyomaAfterformedI0.0256.5incubation;waswastrifuged.Then[3H]DNA DNA Determination of the ratio ofpolyoma type to types II plus III DNA after incubation with empty capsids](https://thumb-us.123doks.com/thumbv2/123dok_us/1575706.110160/9.504.53.247.60.235/polyomaafterformedi-incubation-waswastrifuged-determination-ratio-ofpolyoma-incubation-capsids.webp)
