Copyright 0 1975 American Society for Microbiology Printed inVol.16, No.U.S.A.
Polyoma Virus
Strain with Enhanced Synthesis
of
Capsid
Protein
THOMAS G. TACHOVSKY' ANDJ. D. HARE*
DepartmentofMicrobiology, University of Rochester SchoolofMedicineandDentistry, Rochester,
NewYork14642
Received forpublication 20February1975
Astudyof theimmunochemical characteristicsand thesynthesisofthe capsid proteinsoftwopolyomavirusstrains (3049and1pS)wascarriedout todetermine the mechanism responsible for the unique accumulation of those structural polypeptides in the cytoplasm ofcells infected with the 3049 strain. Antisera prepared against disaggregated virus peptides and whole virus were used to measurethequantityofvirus-specific antigensincellsinfected by thetwostrains by using an indirect radioimmunoassay technique. The 3049-infected mouse
embryocellswerefoundtocontainseveral-fold moreantibody-binding material
than those infected with thelpS strain. Furthermore,thecytoplasmicfraction of 3049-infected cells also containedmoreantibody-binding activity,supportingthe
hypothesisthat thephenotypeof the3049virus(cytoplasmiccapsidprotein)was areflection of the increasedsynthesisofthecapsid polypeptides.
Viruses are known to cause a number of changes in the metabolism of host cells. Cur-rently, investigations are concerned with the identification of virus-specific functions and the elucidation ofthe molecular basisofthe virus-cell interaction. Polyoma(Py) virus, becauseof its limited genetic material, is auseful tool for probing virus-cell relationships. Toward this end a series ofconditional-lethal mutants has beendescribed (4, 9).Inadditiontochemically induced mutant strains, several naturally oc-curring strains have been described (12, 15), withdifferences in plaque size (7, 8) orin the induction of virus-specifictransplant immunity
(14).
A new variation among Pyvirus strains has been studied in ourlaboratory (6, 15). Permis-sive mouse cells infected with 3049 strain dis-playboth nuclear andcytoplasmicfluorescence when stained forcapsid antigen bythe indirect immunofluorescence technique. All other Py strainsdisplayonly nuclear fluorescence (5, 17, 19, 23, 24). However, in the nonpermissive hamster embryo cell, the 3049 strain produces only nuclear fluorescence (15).This new variant is therefore unique among Py viruses in the accumulation ofdemonstable amounts of cap-sid antigeninthecytoplasmofpermissive cells. Studies were undertakento measure the quan-tityofvirus-specificproteinssynthesized bythe
' Presentaddress: Wistar Institute of Anatomy and Biol-ogy,Philadelphia,Pa. 19104.
newvarianttodetermine whetherthe cytoplas-mic fluorescence results from increased quan-tities of capsid polypeptide and possibly to elucidate the mechanism responsible for the expression of thisphenotype.
MATERIALS AND METHODS
Virus strains. The preparation andoriginofthe
3049(12)andlpSlarge plaque(8,20)strains andthe hemagglutination and plaque assays (11) were de-scribedpreviously. Thestock virus used wasproduced
in baby mouse kidney cultures (26). The rate of attachment and subsequent infection of mouse
em-bryo cellswas identical for the 3049 and lpSvirus strains.
Viruspurification.Virusused for antiserum
pro-ductionand asantigenforimmunologicalstudies was
purifiedaccordingtothemethodofRoblinetal. (21). This entailedlow-speed centrifugationofcrudevirus
suspension with subsequent release of cell-bound
virus by treatment of the cell debris with receptor destroying enzyme followed by fluorocarbon extrac-tion (Genetron 113-Allied Chemical, Morristown, N.J.).Supernatantvirusplusvirusfreed fromcellular
componentsweresubjectedtovelocity centrifugation
inKBrfollowed byisopycnic centrifugationinCsCl.
Purifiedvirus was concentrated eitherby pelletingat 100,000 x g for 2 horby lyophilization.
Virusdisaggregation.Purified viruswasdialyzed againstdistilledwaterandlyophilized.Theresulting
powder was dissolved inasolution of8Mureain1M Tris solution, pH 8.5, with 10-' M EDTA. The solution was degassed under vacuum and bubbled withnitrogen for30min.Two-mercaptoethanol was added to a final concentration of 0.75 M, and the 116
on November 10, 2019 by guest
http://jvi.asm.org/
POLYOMA
solution was allowed to stand for 2 to 4 h at room temperature. An aliquot of freshlyprepared solution of 1.2 M iodoacetamide in 1 M Tris, pH 8.5, was addedtoafinal concentration of 0.37 M iodoacetam-ide. The tube was covered with tin foil to prevent thelight-catalyzed formation ofI,andapH of 8.0was maintained by adding 2.2 M Tris base, pH 10.5. Between pH readings the solution was maintained under nitrogen. After 2 h the S-carboxymethylated peptidesweredialyzed against 0.15 M NaCl.
Cell culture techniques. Mouse embryo cell
cul-tures were prepared as described previously (13). Hamsterandratembryo cell cultureswereprepared from 12- and 15-daypregnantanimals, respectively, by using thesamemethod. The 3T3-Py6 cell linewas received from Thomas Benjamin (4); P388D cell line fromAllan Rabson;and Hy2a and Hy7a, mouse-ham-ster somatic cell hybrid lines, from C. Basilico (3). Cells were cultivated in modified Eagle minimal essential medium (AutoPow) with penicillin and streptomycin and supplemented with either 5% or 10% heat-inactivated bovine fetal serum. Cells for immunofluorescence were grown in chamber slides (Lab Tek Products). All other cell culture was in plastic petri dishes (Falcon) with or without glass
coverslips.
Production ofantisera. Antisera to whole virus wereprepared and absorbed asdescribed previously (12, 15). Antiseratodisaggregated virus polypeptides were prepared by homogenizing reduced, S-carbox-ymethylated virus peptides in incomplete Freund adjuvant and inoculating 0.1 ml, containing 12to 15 ;sgofprotein, intothefootpads of rabbits. An
addi-tional 0.4 mlof the same proteinsolution innormal saline was injected intraperitoneally. Each rabbit received a similar series of injections without adju-vant at 2-week intervals for 6 weeks. A second antiserum to disaggregated virus polypeptides (RIPCD) was a gift of P. S. Lombardi (P. S. Lom-bardi, Abstr. Annu. Meet. Am. Soc. Microbiol. 1973, V10,p.196).
Fluorescent antibody technique. Fluorescent antibody staining was carried out as previously de-scribed (6). Fluorescein-labeled immunoglobulin G (IgG) fraction ofgoatanti-rabbitgammaglobulinwas obtained commercially (Cappel Laboratories, Down-ington, Pa.).
Immunodiffusion. Double diffusion was carried outin 1%Nobleagarcontaining 0.5 Mglycine,0.15M NaCl, and 0.01% merthiolate. Whole virus was ap-plied to the plates 36 h before the application of capsid polypeptide preparations. The addition ofup to 3 M urea to the agar had no effect on the development of precipitin lines. The plates were allowed to develop for at least 48 h at 37C in a humidified chamber, washed thoroughly, dried and stained with0.1%Buffaloblackin 50%methanol and 10%acetic acidsolution;excessstainwasremovedin
2%acetic acid.
12II-labeling. The IgG fraction ofgoatanti-rabbit
gammaglobulinwaslabeled with Na121(Amersham/
Searle)by the iodine monochloride technique (2). Indirectradioimmunoassay.Cellsgrownonglass
cover slips were fixed and reacted with antiviral antibody as described for the immunofluorescence test. The cover slips were incubated for 30 min at room temperature with 125I-labeled goat anti-rabbit
gamma globulin, rinsed three times for 10 min in Tris-buffered saline and counted in a gamma spec-trometer.
Radioimmunoassay of cellular fractions. Mono-layers of mouseembryo cells were mock infected with
Tris-buffered salineorinfected with 3049or lpSat a multiplicity of 40PFU/cell. At24 hpostinfection the cellmonolayers were removed bytrypsinization,
cen-trifugedat 800x g for10min, andresuspendedat106
cells/ml in homogenization buffer-0.5 M Tris, pH 7.6, 0.15 M KCI, 3 x 10-3 M MgCl, 0.5% Nonidet, 10-3 Mdithiothreitol.Thecellsuspension was
homog-enized in atight-fitting Dounce homogenizer and the extent of cell disruption was followed by phase
microscopy. More than 90% of thecells weredisrupted
after 25strokes. The nuclei were removedby centrifu-gation at 1,500 x g for 20 min and thesupernatant
(cytoplasmic fraction) was saved. The pellet was rinsed twice with homogenizingbuffer less detergent. Both fractions were then frozen and thawed three times,sonicated for 15 s, andcentrifugedat1,500 x g for15 minbefore use. Supernatant fluids served as the source of viral or controlantigens.
Freshly prepared extracts proved to be better sources of antigen than materials which had been stored frozen. The loss ofantigenicallyactivematerial with time was alsoobserved in the immunofluores-cencetest.
Radioimmunoassaywasperformed asdescribedby Hayashietal. (16).Twenty-microliteraliquots of the samples were dried in wells of polyvinyl microtiter dishes(CookeEngineeringCo.,Alexandria, Va.) and fixed for 5 min with methanol. Nonimmune and antiviral sera (0.02 ml/well) were added and the plates were incubated for 1 h at 37 C. After rinsing with distilled water 10 times, 0.02 ml of1261-IgG frac-tionofgoatanti-rabbitgammaglobulin (about100,000
counts/min)inTrisbuffer with 1%BSAwasadded to
eachwell andincubationcontinued for an additional 1h at 37C. Theplateswerethen rinsed 10 times with distilled water, and the wells were separated and counted in a Packard gamma spectrometer.
RESULTS
Characteristics of antisera produced against disaggregated virus polypeptides. The detection of capsid antigen in the cyto-plasm of 3049-infected mouse embryo cells in the absence of detectable numbers of virus particles (6) suggested the accumulation of a precursor form of the antigen. To investigate this possibility, antisera were prepared against the reduced, S-carboxymethylated polypep-tides of both the 3049 and lpS virus strains. Care wastaken toreduce theamount of cross-reactivity of the antisera by initially injecting small amountsof antigen emulsified in
incom-VOL.16,1975 117
on November 10, 2019 by guest
http://jvi.asm.org/
pleteFreundadjuvant(25).
Table 1 lists the characteristics of the anti-sera produced in rabbits against intact 3049 virions (anti-3049 pool) as well as 3049 (anti-3049D) and lpS (anti-1pSD) polypeptides. Nei-ther of the antisera to disaggregated polypep-tides possessed neutralizing or hemagglutina-tion-inhibiting activity againstthe intact virus. Both sera, however, reacted with the viral antigensinthe infected cells detectedby immu-nofluorescence, as shown in Table 2. The distribution of fluorescence was found to be identical to thatproduced when the whole virus antiserumwasused.
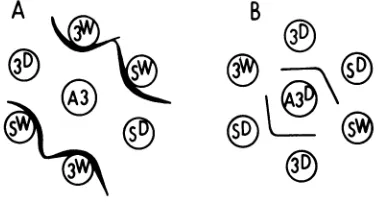
The immunological reactivities of these anti-sera were examined further by immunodiffu-sion. As shown diagrammatically in Fig. 1A, anti-3049 pool serum reacted with both 3049 andlpSvirusparticles toformaheavy precipi-tin linethat showed a reaction ofidentity. This whole virus antiserum, however,did not precip-itatewithdisaggregated virus proteins.Asmall spurfrequently formed inthe reaction between 3049 antiserumand the 3049antigen (Fig. 1A), demonstrating that therewas, in additiontothe major shared determinant, a second antigenic determinant present on the 3049 virion not representedonthelpSvirion.
By contrast, precipitin lines were formed in the reaction between antisera to disaggregated virus proteins and the virus polypeptide, but not with intact virions. These reactions are
shown inFig. 1B in which anti-3049Dserum is included; anti-lpSD serumshowed an identical
pattern.
Theprecipitin
lines between thedisag-TABLE 1. Somecharacteristics ofantisera against wholeanddisaggregated virus
Antibodytitera Test Anti-3049 Anti- | D
pool 3049D
Anti-1S
Hemagglutinationin- 1,260 40c 40c
hibitionb
Plaqueneutraliza- 400,000 loc loc
tiond
Immunofluorescencee 640 160' 80'
aReciprocal of the last dilution positive for each test.
"Dilution of serum sufficient to inhibit 8
hemag-glutinatingunitsofbothSE3049andlpSstrains. cPreimmunization serum displayed similar activ-ity.
dDilutionof serumnecessarytoproduce50% inhi-bitionofplaque formationby both virus strains.
tPreimmunization serum displayed no reaction withinfected cells.
TABLE 2. Distribution of fluorescence at 24 h
postinfectionin mouse embryo cells infected withSE 3049and IpSviruses
3049b lpSb
Antiseruma Nucleus Cyto- Nucleus
Cyto-plasm plasm
Anti-3049pool +c + + c
Anti-210 + + +
Rl-PCDd + + + _
Anti-3049De + + + _
Anti1pSDe + + + _
aAll sera used at 1:10 dilution in complete Tris buffer.
bMultiplicity of infection, 40 PFU/cell. c+, Fluorescence; -, nofluorescence.
dAntiserum prepared against disaggregated poly-omaand received from P. Lombardi.
eAntisera againstdisaggregated polyoma prepared forthisstudy.
B
e
FIG. 1. Double diffusion gel patterns. Symbols: A, anti-3049 pool antiserum; B, anti-3049 polypeptide antiserum. A3, anti-3049 pool; A3D, anti-3049
poly-peptide; 3W, 3049 virion; Sw, IpS virion; 3D, 3049
polypeptide; SD, IpS polypeptide.
gregated polypeptides of the two virus strains and their respective antisera showed reactions ofidentitywith noevidenceforthe formation of asimilarspur as seen inthe wholevirussystem. These reactions demonstrate that the intact virions of both virus strains carry one major antigenic determinant that is common in the two strainsbut which isdistinct fromthemajor determinanton thedissociated, S-carboxymeth-ylated polypeptides of the two strains. In spite of these differences, antisera to intact virus and to the polypeptides detect their re-spective antigenic determinants in the cyto-plasmand nucleus ofthe3049-infected cellsbut only inthe nucleus oflpS-infected cells(Table 2).
Expression ofthe 3049 phenotype in vari-ous host cells. Previous studies (6, 15) have shown that hamsterembryo cells infected with the 3049 strain of Py display only nuclear
J.VIROL.
A
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.498.275.467.97.251.2] [image:3.498.276.465.295.396.2] [image:3.498.71.265.453.591.2]fluorescence in the small proportion of cells in which lytic infection takes place. The failure of hamster embryo cells to accumulate capsid antigen in the cytoplasm suggested a unique virus-cell interaction in which the synthesis of capsid polypeptide was regulated by the host cell. Avariety of cells were theninvestigated for their effect on the expression of this unique phenotype and the results are summarized in Table 3.
Cells of mouse origin, including both primary cell cultures and established cell lines, all displayed cytoplasmic fluorescence when tested withanti-3049 pool serum for capsid antigens 24 h after infection. Of special interest was the expression of the 3049 phenotypic character in the 3T3-Py6 cells, a cell line establishedbythe transformation of 3T3 mouse cells by a host range mutant ofPy (4). The presumed presence ofanother Py genome had no effect on the 3049 phenotype.
Mouse-hamster somatic cell hybrid lines Hy7a and Hy2a (3) were also examined. Both virus strains replicated in each line (unpub-lished data); the samefluorescentpatternswere observed in these cell lines as in permissive mouse embryo cells. The presence of hamster chromosomes in these two cell lines did not suppress the cytoplasmic fluorescence pheno-type.
Twoother cell types were also tested.Primary babyhamsterkidneycellsdisplayed 0.1 to 0.2%
TABLE 3. Capsidpolypeptide distribution in various cell typesasdeterminedbyindirect
immunofluorescence
3049 lpS
Celltypea Nucleus Cyto- Nucleus
Cyto-plasm plasm
Mouse
MEF +b + +
-P 388D1 + + +
-BMK + + +
-3T3-Py6 + + +
-Mouse-hamster hybrid
Hy7a + + +
-Hy2a + + +
-Hamster
BHK + +
-Rat
REF + + +
aCellsweregrowninchamber slides and fixed and stained at 24 h postinfection, except baby hamster
kidneyand ratembryo fibroblasts, which werefixedat 48hpostinfection.
b+,Fluorescence; -, fluorescence negative.
positive cells in the immunofluorescence test. The distribution of fluorescence in these cells was identical to that observed in hamster em-bryo cells. On the other hand, ratembryo fibro-blastsnotonly displayedagreater percentageof cells (15 to20%) producing capsid polypeptide but the cells infected with 3049viruswere posi-tive in both nuicleus and cytoplasm.
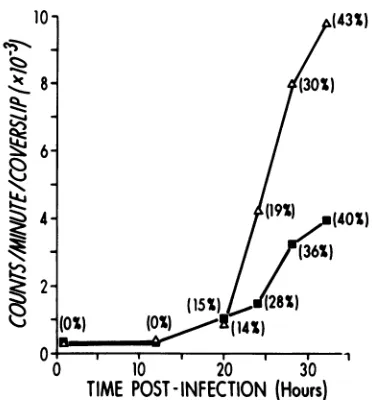
Measurement of capsidproteins by quanti-tative radioimmunoassay. To measure the relative quantity ofcapsid proteinsor polypep-tides produced in mouse cells infected with the 3049or lpS virus strains, the indirect radioim-munoassay technique described by Hayashi et al. (16)wasutilized.The binding of125I-labeled goatanti-rabbit gamma globulin to cells treated with rabbit anti-3049 pool serum at different times after infection is shown in Fig. 2. Capsid antigen was detected inequivalent amounts in cells infected with both viruses at 20 h. From that point on, however, 3049-infected cells bound increasingly greater quantities of the labeled antiserum than lpS-infected cells.This increased binding of 12I-antiglobulin was not the result of a greater proportion of cells becom-ing infected by the 3049 virus as indicated by the similar number of cells synthesizing capsid proteins in the two infections, as measured by immunofluorescence, but rather reflected a
1)c
10-
(43S)
8- 4{0%)
6-4- (19%)
(40)
-
)-(36X)
2-(154)
)28S
(1
0 ,. . . 1
0 10 20 30
TIME
POST-INFECTION
(Hours)
FIG. 2. Binding of 125I-antiglobulin to infected mouseenbryo cells treated with 3049 pool anti-serumin the indirectradioimmunoassay. Numbers in parenthesis indicate percentage of cells positive for
capsidantigen as determined by the indirect
immu-nofluorescencetest. Eachpoint represents the average
of five determinations. Symbols: (A) SE3049; (U)
ips.
16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.498.247.433.393.593.2] [image:4.498.37.232.424.627.2]greater amount of antigenic material in 3049-infected cells.
Radioimmunoassay of capsid proteins in subcellular fractions. The demonstration of more antibody-binding material in 3049-infected cells supported the hypothesis that more capsid polypeptide was synthesized. An attempt was made to demonstrate that more polypeptide was present in the cytoplasmic fraction of infected cells.
Table4showstheresultsobtained by indirect radioimmunoassay usingthe nuclear and cyto-plasmic fractionsofvirus-infected cells. In each case 3049-infected cell fractions, both nuclear and cytoplasmic, bound more '25I-antiglobulin than the lpSfractions. Significantly more anti-gen was detectable inthe cytoplasmic fraction of the 3049-infected cells by the anti-disaggre-gated virus antisera than by the anti-whole virus antiserum.
The nuclear fractions of both infected cell populations contained detectable viralantigen, thereactingspecies present in a form recogniza-bleby both the anti-whole virus antiserum and the anti-disaggregated virusantisera.
Inhibition of capsid protein synthesis by metabolic inhibitors. The effect of several metabolic inhibitorson thesynthesis ofcapsid proteins as measured by the indirect radioim-munoassay in 3049virus-infected cellswasthen studied. As shown inFig. 3,fluorodeoxyuridine, actinomycin D, a-amanitin and cycloheximide allstrongly inhibited synthesisofthe antibody-binding proteinwhen added 1 hafterinfection, followed by a progressive reduction in the de-greeofinhibition duringthe course of infection. Several interesting points should be noted. The synthesisofcapsid antigenwas asdependenton continuing DNA synthesis as it was on RNA synthesis.The effect ofca-amanitin, aninhibitor
TABLE 4. Subcellular distribution of capsid
polypeptideinIpS-and3049-infected mouse embryo cells asdetermined by indirectradioimmunoassay
Counts/minperwelIb Antigena Anti-3049 Anti-3049D Anti l SD
SE3049/cytoplasm 60±2 210 +9 210 +8
lpS/cytoplasm 40±2 0 0
SE3049/nucleus 550± 15 730± 16 400 + 6 lpS/nucleus 280± 11 320±9 1704 3
aCells were cultured in 100-mm petri dishes in-fected with virus at amultiplicity of 40PFU/cell,and harvested 24 hlater.
bMean±e 1standarddeviation for three determina-tions.
80- *FdU
0 ACYCLOHEXIMIDE
~60- 50-
40-O-'4,...
05 30- 20-
10-0 5 10 15 20 25 30
TIMEOFADDITIONOF INHIBITOR(Hrs post-infection)
FIG. 3. The effect of various metabolic inhibitors on the synthesis of capsid proteins in 3049-infected mouse embryo cells as measured by bindingof 125J_ antiglobulin to cells treated with 3049 pool anti-serum. The inhibitors wereadded at different times afterinfectionand all coverslipswereharvestedat30 hpostinfection.Results areexpressedas apercentage ofbinding tocontrol,untreated cellsat30handare charted atthe time when the inhibitorwasadded.
Ac-tinomycinD, 0.05,ug/ml; a-amanitin,1
Asg/ml;
fluoro-deoxyuridine (FdU), 15A.g/ml;
cycloheximide, 1 lAg/ml. Fiveduplicate cover slip samples were assayed for eachdetermination.
of nucleoplasmic RNA polymerase II (18), was lost morerapidlythanthatofactinomycin D,in spite of the fact that it was fully as active as actinomycin D when added early in infection. Finally, as would be expected, cycloheximide inhibited synthesis of the protein for a longer timethan anyofthe other compounds.
DISCUSSION
The 3049 strain of Py virus displays a uniquely different phenotype from other re-ported strains in that capsid polypeptide is detectable in the cytoplasm as well as the nucleus of the infected mouse cell. Several possible mechanisms for this phenotype were
considered, namely, altered diffusion ofcapsid protein from cytoplasm to nucleus, or enhanced synthesis of capsid protein (6). This study was carried out to measure the relative amountsof capsid polypeptide in the whole infected cell or
fractions thereof, following infection with the 3049 virus and a control, wild-type virus (1pS) using more quantitative, radioimmunoassay techniques.
Antisera to intact virions as well as the dissociated, S-carboxymethylated polypeptides derived frompurifiedvirionswerepreparedand characterized byseveraldifferent criteria.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.498.273.459.60.236.2] [image:5.498.69.263.528.614.2]serum to intact 3049 virions detected a major commonantigenicdeterminantonthe 3049 and lpS particle but did not react with the dis-sociatedpolypeptides. Antisera to the disaggre-gated, S-carboxymethylated polypeptides of either 3049 or lpS virions detected a major common antigenic determinant on the dis-sociated polypeptides of both virus strains. These seradidnot reactwith the intact virion of either strain as measured by neutralization of infectivity, inhibition of hemagglutination, or formation ofprecipitin bands. These data indi-cated that the capsid polypeptides ofthe two virus strains were immunochemically indistin-guishable and therefore the use of a radioim-munoassay to measurethe relative quantities of capsidproteins in the cells infected with thetwo virus strains was valid. Furthermore, these reagents could be utilized toidentify precursor polypeptides as well as the protein configura-tioncharacteristic of intact virions.
With these reagents it was then shown that the cytoplasm as well as the nucleus of the 3049-infected cellcontained virus-specific poly-peptides that reacted with antisera to both intact virionsandthedissociated polypeptides. The lpS-infected cell, however, showed
evi-dence
forthepresenceof polypeptides carrying the two antigenic determinants only in the nucleus. These findings support the concept that the capsid polypeptide is synthesized in thecytoplasmand israpidly translocatedtothe nucleus in thewild-type infection where parti-cle assembly takes place. The precursor mole-cule,however, neverattainsalevel sufficienttobe observed in thecytoplasmwith the indirect fluorescent antibody technique. In the case of the3049 strain, infection is associatedwith the production of the polypeptide resulting in a concentration in the cytoplasm sufficient to react positively with the fluorescent antibody technique. Furthermore, the demonstration that the antigenic determinants characteristic of both intact virions and precursor polypep-tides arefound inthecytoplasm aswellasthe nucleus suggests that some structural matura-tion mayalso takeplace inthe cytoplasm.
The next step was to measure the relative quantity ofvirus-specfic antigenic material in cells infected with 3049andlpSvirusesusingan indirect radioimmunoassay as described by Hayashi et al. (16). By using carefully con-trolled infectious doses of each virus strain to yield equal numbers of cells producing capsid protein detectedby immunofluorescence, it was possibletodemonstrate that 3049-infected cells bind several-fold morevirus-specific antiserum than lpS-infected cells. This was detected by
the specific bindingof 125I-labeled IgGfraction of goat anti-rabbit gamma globulin. Further-more, the cytoplasmic fraction of the 3049-infected cell was shown to contain antibody-binding antigen under conditions inwhich the cytoplasm of
lpS-infected
cells failed to bind antibody.These findings provide direct evidence that the synthesis ofcapsid protein is significantly higher in 3049- than in
lpS-infected
cells and that there is a significant accumulation of capsid polypeptide precursor molecules in the cytoplasm ofthe 3049-infected cell. Recently it has been demonstrated that there is also a twofold greaterquantity ofvirus-specific poly-adenylated RNAin both the nuclear RNA and polyribosomalfractions of3049-infectedcellsas comparedtothose infected with lpSvirus(22). Onthisbasis, then, itcan beproposedthat the phenotype of the 3049 virus, namely, the pres-ence of capsid polypeptide molecules in the cytoplasm, is the direct result of the increased numbers of mRNA templates specifying at least the major structural protein, VP2 (21), rather thansomevirus controlled enhancement of the translation rate fromthe mRNA. Direct -assess-mentof the rate of translation or thequantity of translatable mRNA has not been carried out, however.Twootherfindings bear on the question as to which level of genetic control is altered in the 3049 virus replication process. The first is the demonstration that the expression of the pheno-type is closely correlated with the presence of certain mouse chromosomes such that in the absence of mouse genetic information, i.e., in hamster cells, the phenotype issuppressed. On the otherhand, inmouse-hamster hybrids with sufficient mouse DNA represented, the virus replicates and the phenotype is expressed. The ratembryo cell is ofinterest in that a significant number of cells are permissive for the expres-sionof"late" functions and the 3049 phenotype is expressed. These findings suggest that some host cell function is intimately associated with the expression of thephenotype.
A second finding of interest is the prolonged sensitivity of the synthesis of capsid protein to inhibition of DNA synthesis by FdU, with a pattern quite similar to that produced by ac-tinomycinD(Fig. 3). This isreminiscent of the demonstration by Glover (10) that inhibition of Py DNA synthesis restricts the quantity and quality ofthe viral RNA transcribed "late" in infection to yield an RNA population that resembles "early"transcripts.
Taken together, these findings suggest that the rate of synthesis of this late virus gene
on November 10, 2019 by guest
http://jvi.asm.org/
122 TACHOVSKYANDHARE
product, the majorcapsidprotein, is regulated by thequantity ofmRNA available for transla-tion. Furthermore, the quantity of mRNA may in turn be controlled bysomehost cellfunction which determines the rate of transcription of the viral genome following the onset of viral DNA synthesis. This function could beeither a host DNA or RNA polymerase thefunction of which is modified by some factor associated with the synthesis of late viral functions. The reverse transcription co-factor postulated by Amati (1)would alsofitwiththispostulate. The elucidation of the mechanism of this unique phenotype should provideasignificant increase in the understanding of genetic regulation in eukaryotic host cell-virus systems.
LITERATURE CITED
1. Amati, P. 1974. A working model foroncogenic DNA virus replication. J. Theor. Biol. 46:221-227.
2. Bale, W. F., R. W.Helmkamp, T. P.Davis,M. J.Izzo, R. L. Goodland, M. A. Contreras, and I. L. Spar. 1966. High specific activity labelingofproteinwith 131I by
the iodinemonochloride method. Proc. Soc.Exp. Biol. Med. 122:407-414.
3. Basilico, C., Y. Matsuya, and H. Green. 1970. The interaction ofpolyoma virus with mouse-hamster
hy-brid cells. Virology 41:295-305.
4. Benjamin, T. L. 1970. Host rangemutantsofpolyoma
virus.Proc. Natl.Acad. Sci.U.S.A.67:390-399. 5. Bereczky, E., R.Hughes,J.M.Bowen,W.Munyon,and
L. Dmochowski. 1965. Study of DNA synthesis and antigen formation in polyoma virus-infected mouse embryo cells by autoradiographyand immunofluores-cence.Texas Rep.Biol. Med. 23:3-15.
6. Betts, R. F., T. G. Tachovsky, and J. D. Hare. 1972. Studiesonthe mechanism ofcytoplasmic antigen
ac-cumulation following infection with a new variant of polyoma virus.J. Gen.Virol. 16:29-38.
7. Crawford, L. V. 1962. The adsorptionofpolyomavirus. Virology 18:177-181.
8. Diamond, L.,and L. V.Crawford.1964.Some
characteris-tics oflarge-plaqueandsmall-plaquelines ofpolyoma
virus.Virology22:235-244.
9. Eckhart,W. 1969.Complementationand transformation
by temperature-sensitive mutants ofpolyoma virus. Virology38:120-125.
10. Glover,D. M. 1974.CouplingofpolyomaDNA andRNA synthesis. Biochem. Biophys. Res. Commun.
57:1137-1143.
11. Hare, J. D., P. Balduzzi, and H. R. Morgan. 1963.
Polyoma virusand L cell relationship. I. Some charac-teristics of a cell linepersistently infected with poly-oma virus. J. Natl.Cancer Inst. 30:45-56.
12. Hare, J. D., and J. Chan. 1966. Antigenic variation among polyoma viruses fromdifferent sources demon-stratedby plaque neutralization. Virology 30:62-73. 13. Hare, J. D. 1967.Localization and characterization of the
phenylalanine transport mechanism in normal and polyoma-transformed hamster cells. Cancer Res. 27:2357-2363.
14. Hare, J. D. 1967. Transplant immunity to polyoma-induced tumorcells. IV. A polyoma strain defective in transplant antigen induction. Virology 31:625-632. 15. Hare, J. D. 1970. Anew type of variation among polyoma
virus characterized by cytoplasmic accumulation of capsid antigen. Virology 40:978-988.
16. Hayashi, K., J. Rosenthal. and A. L. Notkins. 1972. Iodine-125 labeledantibodytoviral antigensbinding to the surface of virus-infectedcells. Science 176:516-518. 17. Henle, G., F. Deinhardt, and J. Rodrigues. 1959. The
developmentofpolyomavirus in mouseembryo cells as revealed by fluorescent antibody staining. Virology 8:388-391.
18. Lindell, T. J., F.Weinberg, P. W. Morris, R. G. Roeder, and W. J. Rutter. 1970.Specificinhibition of nuclear RNA polymerase II by a-amanitin. Science 170:447-449.
19. Malmgren, R. A., R. Rabatti, and A. S. Rabson. 1960. Intracellular localization of polyoma virus antigen demonstrated with fluorescein labelled antisera. J. Natl. Cancer Inst. 24:581-588.
20. McCulloch, E. A., A. F. Howatson, L.Siminovitch, A. A.
Axelrad, and A. W. Ham. 1959. Tumor induction in
Swissmicebyvirusobtainedoriginallyfrom a
mam-mary tumor inC3H/Jaxmouseand then cultivated in vitro. Proc. Am. Assoc.CancerRes. 3:41.
21. Roblin, R., E. Harle, and R. Dulbecco. 1971. Polyoma
virus proteins. I.Multiple virion components. Virology 45:555-566.
22. Rutherford, R. B., and J. D. Hare. 1974. Evidence fora
regulatory function related to the expression of the polyoma genome following the onset of virus DNA replication. Biochem. Biophys. Res. Commun 58:839-846.
23. Sachs,L., and M.Fogel. 1960. Polyoma virus synthesis in tumorcells as measured bythe fluorescentantibody
technique.Virology 11:722-736.
24. Williams, M. G., and R. Sheinin. 1961. Cytological
studies of mouseembryocells infected withpolyoma
virususing acridine orange and fluorescent antibody.
Virology 13:368-369.
25. Vestergard-Jorgensen, P. E. 1972. Freund's adjuvants: theirinfluence on thespecificityofviralantisera.Acta Pathol. Microbiol.Scand. Sect. B 80:931-933. 26. Winocour,E. 1963.Purification ofpolyomavirus.
Virol-ogy 19:158-168.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/