capped mRNA from their vertices. Analysis of the pseudoatomic model of the VP6 layer, obtained by placing the atomic structure of VP6 into electron microscopy reconstructions of the DLP, has identified the regions of the protein involved in interactions with the internal layer. To study the role of VP6 both in the assembly of DLP and in transcription, 13 site-specific substitution mutations of VP6, targeting the contacts between the two inner layers, were constructed and expressed in the baculovirus system. The effects of these mutations on VP6 expression, trimerization, and formation of macromolecular assemblies were investigated. Using either in vitro reconstituted DLP derived from purified viral cores and recombinant VP6 or in vivo self-assembled virus-like particles resulting from the coexpression of VP2 and VP6 in the baculovirus-Sf9 system (VLP2/6), we have identified the amino acids essential for recovery of transcription or assembly. All VP6 mutants formed stable trimers which, like wild-type VP6, assembled into tubular structures. The ability of VP6 to interact with VP2 was examined by several assays, including electron microscopy, coimmunoprecipitation, purification of VLP2/6, and monitoring of the transcriptase activity of reconstituted DLP. Of the 13 VP6 mutants examined, 3 were unable to assemble with VP2 and 3 others partially assembled. These mutants either did not rescue the transcriptase activity of core particles or did so only marginally. Four mutants as well as the wild-type VP6 assembled and transcribed very well. Three mutants assembled well on cores but, surprisingly, did not rescue the transcriptase activity of reconstituted DLP. Our results indicate that hydrophobic interactions between VP6 and VP2 residues are responsible for the stability of the DLP. They also show that subtle electrostatic interactions between VP6 and the underlying transcriptase machinery can be essential for mRNA synthesis.
Rotaviruses are members of the Reoviridae family of seg-mented, double-stranded RNA viruses. Viruses in this family are nonenveloped, and their complex capsids contain several concentric protein layers displaying icosahedral symmetry. Structural studies by X-ray crystallography have revealed the complex interactions between the innermost layers of some of
theReoviridae, like bluetongue virus (BTV) (9), as well as the
structure of the single-layered core particle of reoviruses (20). In the case of rotaviruses, the crystal structure of the double-layered particle (DLP) has not yet been reported but a pseudo-atomic model has been derived for the middle layer by use of an approach combining X-ray crystallography and electron mi-croscopy (18). As is the case for BTV, the two innermost layers of rotaviruses display different geometries; thus, there is a symmetry mismatch between them. The assembly of the DLP is thus a challenging subject to study in terms of both the protein-protein and protein-RNA interactions. The rotavirus capsid contains four major proteins (VP2, VP6, VP4, and VP7) (7). Coexpression of recombinant VP6 (middle layer) and VP2 (internal layer) in the baculovirus insect cell system results in the production of virus-like particles (VLP) that can be
puri-fied (13). The expression of VP2 alone results in the produc-tion of particles, called pseudo-cores, that have the same ge-ometry as viral cores (15). The two minor constituents of the viral capsid, proteins VP1 and VP3, which are present in low molar ratios within the virion, have RNA transcriptase and RNA-modifying properties (4, 17). In their mature form, the viral particles exhibit no transcriptase activity. Solubilization of the external layer, which occurs upon viral entry into the cy-toplasm of the target cell, activates the particle’s transcriptase activity. Metabolites can then enter the particle, and viral tran-scripts are synthesized and extruded (5, 14). Solubilization of VP6 from the DLP inactivates the endogenous transcriptase activity, which can be recovered by the addition of native or recombinant VP6 (2, 12). This feature allows testing of the effects of different VP6 mutants on the transcriptase activity of the DLP.
The outer layer of the DLP (i.e., the middle layer of the mature particle) is composed of 780 VP6 polypeptides, which are distributed as 260 trimers. Twenty VP6 trimers lie on icosahedral threefold axes; the remaining 240 trimers lie on local threefold axes of the T⫽13 surface lattice. The structure of the VP6 trimer consists of an elongated, tower-like molecule with its molecular threefold axis along its long dimension (19). The base of the tower contacts the inner layer, which is per-pendicular to the molecular threefold axis. The inner layer of the capsid is a thin shell made up of 120 polypeptides of VP2, * Corresponding author. Mailing address: UMR CNRS-INRA
Vi-rologie Mole´culaire et Structurale, 1 Ave. de la Terrasse Baˆt. 14C, 91198 Gif-sur-Yvette Cedex, France. Phone: 33(0)1 3465 2604. Fax: 33(0)1 3465 2621. E-mail: [email protected].
7822
on November 8, 2019 by guest
which form 60 asymmetric dimers which are, in turn, arranged with T⫽1 icosahedral symmetry (15). The VP6 and VP2 layers interact through predominantly hydrophobic surfaces, al-though the base of the VP6 layer displays an overall negative surface electrostatic potential. The mismatch in triangulation of the two layers, T⫽13 and T⫽1, leads to a variety of different environments for each of the 13 VP6 polypeptides at the con-tact region with VP2. Since VP6 forms trimers in solution, it is likely that trimers are the driving force for VP6 and VP2 interactions during core assembly (1, 8, 16, 18).
A 2.0-A˚ -resolution image of the crystal structure of VP6 revealed the intratrimer VP6 interactions, and a pseudoatomic model of the VP6 layer revealed the amino acids of VP6 that are involved in intertrimer contacts and also in the interactions with the other proteins that form the viral particle (18). Each VP6 monomer consists of two distinct domains along the long dimension of the tower-like molecule; the interactions between monomers involve both domains and are extensive. To better understand the interactions of VP6 during capsid assembly and its role in transcriptase activity, we focused our site-directed mutagenesis efforts on the base of the trimer, in the lower domain of VP6, i.e., the region directly in contact with the VP2 layer as identified in the pseudoatomic model (18). A series of substitutions of amino acids was designed based on this model. The effects of these mutations on the overall structure of VP6, on the VP2-VP6 interactions, and on the transcriptase activity of the DLP were examined by several assays, including coim-munoprecipitation, purification of in vivo self-assembled VLP resulting from coexpression of VP2 and VP6 in the baculovi-rus-Sf9 system (VLP2/6) in density gradient, analysis of the recovery of the transcriptase activity of the core, and electron microscopic observations of the integrity of the relevant mac-romolecular assemblies. Of the 13 VP6 mutants examined, 3 were unable to assemble with VP2 and 3 others partially as-sembled. These mutants were incapable of rescuing the tran-scriptase activity of core particles or rescued it only marginally. Four mutants assembled and transcribed very well, as did wild-type VP6. Three mutants assembled well on cores but,
surpris-ingly, did not rescue the transcriptase activity of the reconsti-tuted DLP. Our results indicate that hydrophobic interactions between VP6 and VP2 residues are responsible for the stability of the DLP. They also show that subtle interactions involving electric charges between VP6 and the transcriptase machinery are essential for transcript extrusion and for RNA synthesis.
MATERIALS AND METHODS
Viruses and cells.Spodoptera frugiperda(Sf9) cells were grown in suspension or as monolayers at 26°C in Hinks medium supplemented with 10% fetal calf serum. Derivatives ofAutographa californicanuclear polyhedrosis virus (AcNPV) containing the wild-type rotavirus VP6 gene (BacRF6), the VP2 gene (BacRF2), or the green fluorescent protein (GFP) fused to VP2 (GFPJA16) have been described previously (3, 13).
Isolation of recombinant baculoviruses expressing VP6 mutants and purifi-cation of VLP.VP6 was first subcloned in pFASTBAC, and a series of mutants of VP6 were obtained using the QuikChange kit from Stratagene. All the oligo-nucleotides used for mutagenesis are shown in Fig. 1 as the coding strands of rotavirus gene 6. Donor plasmids were transposed to obtain the corresponding bacmid in DH10BAC cells, and the recombinant bacmid DNA was used to lipofect Sf9 cells. The resulting recombinant baculovirus was propagated as described previously (4). Mutated nucleotides in recombinant baculoviruses were verified by sequencing the corresponding donor plasmid.
Baculovirus-expressed VLP were purified as described previously (13). In brief, Sf9 cells were coinfected with recombinant baculovirus BacRF2 expressing VP2 and with recombinant baculoviruses expressing various VP6 mutants or wild-type VP6 (multiplicity of infection, 5 to 10 PFU per cell). Alternatively, the baculovirus GFPJA16 expressing the GFP-VP2 fusion protein was used in place of BacRF2 for cesium chloride density gradient purification of assembled VLP2/6 (3). After incubation at 26°C for 5 to 7 days, cells were harvested and supernatants were clarified at 4,000⫻gand centrifuged for 60 min at 40,000 rpm in a Beckman 45 Ti rotor. Pellets resuspended in a buffer containing 20 mM PIPES [piperazine-N,N⬘-bis(2-ethanesulfonic acid)] and 10M Ca2⫹(pH 6.6)
were treated with Freon 113 and then adjusted to a density of 1.30 g/ml by the addition of CsCl. After centrifugation at 35,000 rpm in a Beckman SW55 rotor overnight, density gradients were illuminated with UV light to make bands of VLP visible.
The wild-type and mutant rotavirus VP6 proteins were purified as follows. Infected Sf9 cells were harvested 3 to 5 days postinfection, and clarified medium was centrifuged for 30 min at 35,000 rpm in a Beckman 45 Ti rotor. The pellet was resuspended in 50 mM MOPS (morpholinepropanesulfonic acid; pH 6) and then adjusted to 0.3 M CaCl2and centrifuged for 10 min at 13,000⫻g. The
supernatant containing purified VP6 was dialyzed against water at room tem-FIG. 1. Oligonucleotides used for mutagenesis. Designations for VP6 mutants with amino acid substitutions (indicated in bold type) at positions 32, 65, 70, and/or 71 are shown at left. Next to these are the corresponding sequences of gene 6 of bovine rotavirus strain RF (GenBank accession no. K02254). Mutated bases are indicated in lowercase letters. Silent mutations that created restriction sites (for screening) are underlined, and mutated codons are indicated by double underlining.
on November 8, 2019 by guest
[image:2.587.139.448.71.245.2]perature. Under these conditions, VP6 assembled in large polymers that were recovered by centrifugation and used for transcriptase reconstruction assay.
Immunoprecipitation and protein analysis.Sf9 cells infected with one or two baculoviruses at a multiplicity of infection higher than 5 PFU/cell were incubated in Hinks medium supplemented with 1% fetal calf serum and 120 Ci of [35S]Promix (Amersham) per ml. When an extensive cytopathic effect was
ob-served, cells were resuspended in radioimmunoprecipitation assay (RIPA) buffer (10 mM Tris-Cl, 1% Triton X-100, 0.5% sodium deoxycholate, 1 mM EDTA, 150 mM NaCl [pH 7.4]), incubated on ice for 15 min, and then centrifuged at 13,000
⫻gfor 5 min. An aliquot of supernatant was incubated with 1l of the ascitic fluid containing monoclonal antibody RV138 directed against VP6 (11). The complex was precipitated with protein A bound to Sepharose 4B, washed with RIPA buffer, and analyzed by sodium dodecyl sulfate-polyacrylamide gel elec-trophoresis (SDS-PAGE).
SDS-PAGE analysis was performed in the Laemmli gel system. Proteins were detected by Coomassie blue staining or immunodetected after blotting on poly-vinyl difluoride by transverse electrophoresis in 10 mM cyclohexylaminopropane-sulfonic acid (CAPS) buffer, pH 11. RV138 monoclonal antibodies were used for immunodetection of VP6.
Transcriptase assay and in vitro reconstitution of DLPs.The standard tran-scriptase assay contained 0.3g of DLPs, 2 mM concentrations of nucleoside triphosphates (except for UTP, which was used at a concentration of 0.4 mM), 6 mM Mg2⫹, 50 mM Tris [pH 7.7], 1 mM dithiothreitol, 16 U of RNasout (Gibco),
and 1.5Ci of [3H]UTP. The reaction mixture (50l) was incubated at 40°C for
40 min, and samples were taken in duplicate for measurement of the synthesis of macromolecules precipitated by 5% trichloroacetic acid. Typically, the incorpo-ration of [3H]UTP was 10,000 cpm for a 10-l sample. For reconstitution of
DLPs, cores were prepared as previously described (2). The concentrations of cores and VP6 added to core preparations were controlled by SDS-PAGE by use of the NuPage system (Invitrogen) in MOPS buffer according to the instructions of the manufacturer. The amount of DLPs used for transcriptase activity was adjusted in order to keep the amount of VP2 constant in all assays. Purified cores (corresponding to 1g of DLPs) were incubated for 10 min at room temperature with various concentrations of purified VP6 in 6 mM Mg2⫹. Then one-third of
the reconstituted particles was assayed for transcriptase activity as described above. For each mutant, a standard with DLPs and a standard with cores alone were included in the assay. One-third of the reassembled particles was directly analyzed by electrophoresis in 0.6% agarose gel in Tris-MOPS buffer as previ-ously described (21). The last one-third was analyzed by SDS-PAGE to deter-mine the stoichiometry of VP6.
Electron microscopy.Specimens for electron microscopy were prepared with purified VP6, appropriate CsCl density gradient fractions containing VLP, or in vitro reconstituted DLPs. Fractions of density gradient were desalted through a
Sephadex G-25 spin column equilibrated in 20 mM Tris (pH 7.4)–20 mM NaCl–1 mM CaCl2. DLPs were reconstructed as described above in 6 mM Mg2⫹and
incubated at 40°C for at least 10 min. Samples were applied to air-glow-dis-charged carbon-coated grids, blotted immediately with filter paper for 1 to 2 s, and negatively stained with 1% uranyl acetate solution. The grids were examined in a Philips CM12 electron microscope operated at 80 kV.
RESULTS
Rationale for the selection of VP6 residues for site-specific mutagenesis.The amino acid sequence of protein VP6 is well conserved, with about 85% identity, among group A rotavi-ruses, but the conservation is less when comparison is made to VP6 proteins of the other rotavirus serogroups that have been sequenced so far. The amino acid sequence of VP6 of the bovine rotavirus strain RF shows only 40% identity with that of porcine group C rotaviruses and only 16% identity with that of group B rotaviruses (10) (Fig. 2). The lower domain of VP6 consists of a bundle of eight alpha helices (helices␣a through
␣h) derived from two segments of the polypeptide chain. The first segment, containing helices␣a through␣e, is formed by the 150 N-terminal amino acids. The second segment, residues 335 to 397 at the C-terminal end, contains the remaining three helices.
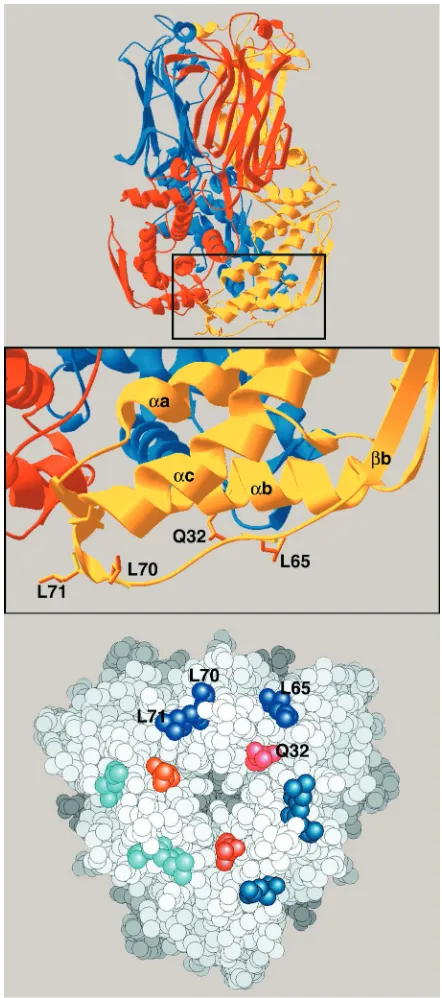
[image:3.587.91.497.73.250.2]The model of the intermediate layer shows that VP6 resi-dues participating in the interactions between VP6 and VP2 come essentially from the extendedb-␣c loop and also from the bottom side of helix␣b, which runs horizontally at the base of the trimer (Fig. 3). Theb-␣c loop (residues 60 to 74) is actually the most conserved region of the polypeptide chain, particularly the segment containing residues 65 to 71 (Fig. 2). The side chains of Leu 65, 70, and 71 point outward and are exposed to the solvent in the isolated trimer. This loop is likely to adapt its fold to the precise type of contact made with VP2, given that the symmetry mismatch between the corresponding layers in the viral particle requires several different types of FIG. 2. Important features of the rotavirus VP6 sequence. The alignment of amino acid sequences of protein VP6 from rotavirus groups A and C is shown. Within group A, the most distant viruses have a VP6 amino acid sequence identity of 65%; the identity of sequences between proteins from groups A and C is 40%. The domains implicated in VP6-VP2 interaction are shown in boxes, and the positions of the site-directed mutations (positions 32, 65, 70, and 71) are indicated by downward-pointing arrowheads. The uppercase black letters denote strictly conserved residues in group A rotavirus VP6 (from analysis of 39 available sequences). For group C rotavirus VP6, uppercase black letters indicate residues having strict identity with those of group A rotaviruses. Uppercase gray letters indicate positions displaying only conservative mutations. Secondary-structural elements are labeled above the sequences: rectangles indicate␣-helices and arrows indicatestrands. The alignment was obtained initially with the PILEUP program and was then manually adjusted based on the crystal structure.
on November 8, 2019 by guest
interactions with VP2. The pseudoatomic model shows that the side chains of the above-mentioned leucine residues point to-ward VP2, as does the side chain of residue Q32 in helix␣b. Consequently, we mutated the four amino acids at positions
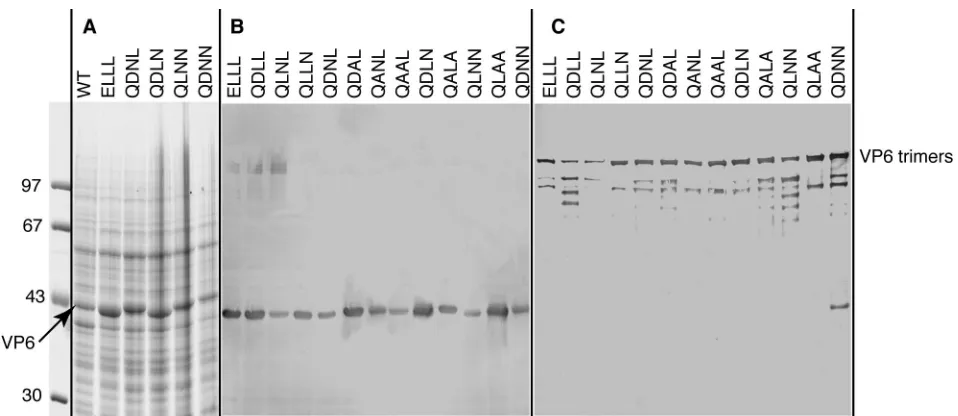
Ala (D29A in the conventional notation) was constructed as a control, since the side chain of this residue does not point to VP2. DNA from each VP6 derivative was then cloned into the baculovirus, and the VP6 mutants were expressed in insect cells. SDS-PAGE analysis of infected cells showed that all VP6 proteins, both mutant and wild type, were expressed to similar levels (Fig. 4A and B). Each VP6 mutant was assayed for trimerization by SDS-PAGE analysis of the unboiled sample as previously described (8, 22). Under these conditions, a major band at an apparent molecular mass of 150 kDa and corre-sponding to the trimer was present for all mutants (Fig. 4C). Minor bands of lower apparent molecular weight, correspond-ing probably to degradation products, were also visible. The QDNNtriple mutant also showed, in the absence of boiling, a band migrating like the monomer. This suggests that perhaps in this case the trimer is not as stable as the wild type. Electron microscopy examination (data not shown) of cell lysates at a pH of 6.6 revealed typical 75-nm-diameter VP6 tubes for all the mutants (16).
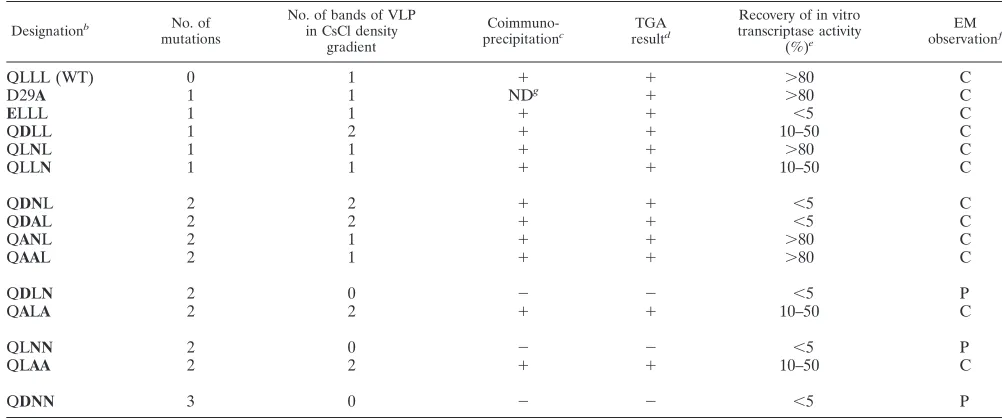
VP6 substitutions at certain specific contact points prevent the assembly of stable capsids in insect cells.To investigate whether VP6 mutants could assemble into the rotavirus capsid, we first used an assembly assay based on the coexpression of VP2 and VP6 in insect cells. As previously described, when these two proteins are expressed it is easy to purify assembled VLP from insect cell lysates by use of CsCl density gradients (13). Each recombinant VP6 was coexpressed with GFP-VP2 protein (3) in insect cells as described in Materials and Meth-ods. At 5 days postinfection, cells were treated with fluorocar-bon and the assembled VLP were purified through a cesium chloride density gradient. The use of a VP2 protein fused to GFP, which has been shown previously to efficiently assemble into VLP (3), simplified the detection of the band of VLP in CsCl density gradient. When expressed alone, VP2 or GFP-VP2 formed hydrophobic aggregates that in the absence of detergent remained associated with cell membrane debris and did not give rise to a band in density gradients (13).
[image:4.587.52.273.72.571.2]Three VP6 single mutants (ELLL, QLNL, and QLLN), two VP6 double mutants (QANL and QAAL), and the D29A con-trol mutant were able to assemble into particulate structures that could easily be visualized as a single band in CsCl density gradients at the same density as that of native VLP. One VP6 single mutant (QDLL) and four VP6 double mutants (QDNL, QDAL, QALA, and QLAA) gave rise to two bands in CsCl FIG. 3. VP6 trimer, with a close-up of the domain that interacts
with VP2. In the top panel, the threefold symmetry axis is vertical and each subunit is displayed in a different color. The side chains that point to VP2 are shown in orange. In the middle panel (a close-up of the boxed portion of the top panel), the numbers of the residues are indicated and secondary structural elements are labeled by lowercase letters preceded by␣or, which denote␣-helices orstrands (see the legend to Fig. 2). The bottom panel shows the VP6 trimer from the interior of the capsid. Residues of a single subunit are labeled, with corresponding residues of the other subunits displayed in similar hues.
on November 8, 2019 by guest
density gradients. The upper band looked like the band ob-served with wild-type VLP. The lower band was sharper and contained flocculate material. The relative intensity of the two bands was not the same for the five VP6 mutants and, in addition, varied from experiment to experiment. Finally, two double mutants (QDLN and QLNN) and one triple mutant (QDNN) gave rise to no visible band after density gradient centrifugation. Representative density gradients for each of the three cases described are shown in Fig. 5. The morphology of VLP purified in CsCl density gradient was characterized by electron microscopy. Wild-type VP6 and all of the VP6 mu-tants that gave rise to a single band in CsCl density gradient appeared similar in the micrographs, with a regular arrange-ment of VP6 trimers on the outer layer of these particles. Those mutants whose VLP gave rise to two bands in CsCl density gradient looked more heterogeneous and were aggre-gated. The upper band contained partially assembled VLP2/6, while the particles contained in the lower band were even more partially decorated by VP6 and often resembled VP2 pseudo-cores (the representative QDNL mutant is shown in Fig. 6). This is probably due to the loss of VP6 trimers from these particles during purification and/or electron microscopy prep-aration. SDS-PAGE analysis confirmed that the stoichiometry of VP6 and GFP-VP2 was altered in the lower band (Fig. 6C). The efficiency of VP6 mutants in assembling with VP2 was further assessed by coimmunoprecipitation. Sf9 cells were in-fected simultaneously by the baculovirus allowing the expres-sion of VP2 and by the baculovirus allowing the expresexpres-sion of various VP6 mutants. The medium was then supplemented with [35S]methionine. Cell lysates prepared in RIPA buffer containing detergent were incubated with an anti-VP6 mono-clonal antibody, and the immunoprecipitates were analyzed by SDS-PAGE (Fig. 7). Under these conditions, two double mu-tants (QDLNand QLNN) and a triple mutant (QDNN) did not
show bands of VP2 coimmunoprecipitating with VP6, which was as expected based on the CsCl centrifugation experiments.
[image:5.587.53.532.69.277.2]Assembly of VP6 with the core is necessary but not sufficient for transcriptase activity. The experiments described above suggested important differences in the affinities of the VP6 mutants for VP2. To better evaluate the affinity of mutated VP6 for VP2, we designed an in vitro assay where interactions between both proteins were evaluated for variable amounts of VP6. A suspension of purified cores was mixed with increasing concentrations of purified VP6 and then assayed for transcrip-tase activity as described above. Simultaneously, an aliquot of the mixture of cores and VP6 was analyzed by electrophoresis FIG. 4. Analysis of the expression of recombinant VP6 mutants and their ability to form trimers. Sf9 cells were infected with recombinant baculovirus, and the infected-cell lysates were analyzed by SDS–10% PAGE and stained with Coomassie brilliant blue (A) or immunostained with an anti-VP6 monoclonal antibody (B and C). The positions of the VP6 monomers (A and B) and trimer (unboiled sample) (C) are indicated. Above the lanes, the site-directed mutations are indicated by the names of the residues (single-letter code) at positions 32, 65, 70, and 71. Molecular weight markers are indicated at the left.
FIG. 5. VLP2/6 formation by rotavirus VP2 and mutated VP6 pro-teins. Sf9 cells were either infected with a recombinant baculovirus allowing the expression of a chimeric protein (enhanced GFP fused to a VP2 protein with the first 92 amino acids deleted) (C) or coinfected with this recombinant baculovirus and a baculovirus expressing the wild-type VP6 (WT) or recombinant baculoviruses expressing various VP6 mutants (QDLL, QLNN, QDNL). The VLP2/6 particles were purified on CsCl density gradients as described in the text.
on November 8, 2019 by guest
[image:5.587.311.530.520.653.2]in 0.6% agarose gel in order to evaluate the percentage of cores that was converted into DLPs. The total amount of VP6 put into the reaction mixture was also estimated by use of SDS-PAGE and Coomassie blue staining. This assay was per-formed for all mutants, and Fig. 8 illustrates the results ob-tained for mutants (QLNL, QLLN, QDNN, andELLL) repre-sentative of the four classes observed. Mutants in the first class, QLNL, QANL, QAAL, and the D29A control mutant, allowed a complete recovery of transcriptase activity when added in at least stoichiometric amounts. The second class includes QDLL, QLLN, QALA, and QLAA. For these mutants, the recovery of transcriptase activity was partial and never higher than 50%, even when VP6 was added in large excess amounts. Agarose gel electrophoresis analysis of these samples showed that they contained a significant amount of cores, suggesting that there was equilibrium among cores, VP6, and DLPs. Three VP6 mutants (QDLN, QLNN, and QDNN) that consti-tuted the third class did not assemble with cores and did not allow the recovery of transcriptase activity. Finally, the fourth class of mutants (ELLL, QDAL, and QDNL) assembled well with cores, as estimated by agarose gel electrophoresis, but showed no transcriptase activity.
All the reaction mixtures were examined by electron micros-copy after negative staining. It appeared that at the highest concentration of VP6 it was always possible to identify some particles resembling DLPs, but in various proportions. With the QDLN, QLNN, and QDNNmutants, the number of DLPs was extremely low and they were aggregated and in many cases partially decorated. With all of the other mutants, we observed high numbers of apparently well-structured DLPs. In particu-lar, micrographs of DLPs from wild-type VP6 and from the
ELLL mutant were indistinguishable (Fig. 9), yet this mutant was transcription incompetent.
DISCUSSION
The crystal structure of rotavirus VP6 revealed a molecule organized in two domains: the top domain consists of a 
sandwich structure that has been observed in many other viral
capsid proteins, and the bottom domain has a characteristic fold made essentially of␣-helices. The pseudoatomic model of the VP6 layer of the viral particle suggested that VP6 residues participating in the interactions with VP2 come essentially from the b-␣c loop and from the bottom side of helix ␣b, which runs horizontally at the base of the trimer (18). The DLP model suggests that the VP2-VP6 assembly relies mainly on hydrophobic interactions. Four amino acids were thus selected on the basis of their locations in the three-dimensional struc-ture, their conservation among group A and group C rotavi-ruses (since it is known that VP6 from group C rotavirotavi-ruses can assemble on top of group A rotavirus cores [22]), and the orientations of their side chains (pointing towards VP2). Each of the four residues (Q32, L65, L70, and L71) was replaced by a hydrophilic (L➟N), charged (Q➟E or L➟D), or small apolar (L➟A) residue. Because these residues are located away from
[image:6.587.122.467.79.234.2]the monomer and the trimer interfaces, we predicted, and and the QDNL mutant of VP6. The VLP were purified by CsCl density gradients, and both the upper (A) and lower (B) bands were analyzed by negative staining. The protein content of each band was analyzed by PAGE (C). Shown are the results for the upper (lane 1) and lower (lane 2) bands; the results for wild-type VLP2/6 are also shown for comparison (lane 3).
FIG. 7. Coimmunoprecipitation assay of VP2 and VP6. Sf9 cells were infected simultaneously with a baculovirus allowing the expres-sion of VP2 and a plasmid directing the expresexpres-sion of wild-type VP6 (WT) or of a site-directed VP6 mutant. The mutations are indicated at the top by the residues (single-letter code) present at positions 32, 65, 70, and 71. At 72 h posttransfection, cell lysates were immunoprecipi-tated by use of anti-VP6 monoclonal antibodies.
on November 8, 2019 by guest
[image:6.587.302.539.519.660.2]verified in this study, that they did not affect VP6 trimerization or the ability of VP6 to assemble in tubes.
The autoassemblies of VP2 into pseudocores and of VP2 and VP6 into VLP by baculovirus-mediated coexpression of the respective genes in insect cells (13) provide a useful model for studying rotavirus protein-protein interactions occurring during virus morphogenesis. VP2 pseudocores are highly sta-ble when assemsta-bled in the absence of VP6 molecules but do not give rise to a band after CsCl density gradient centrifuga-tion (13). The coexpression of VP2 and wild-type VP6 leads to the formation of VLP which do give rise to a band in CsCl density gradient centrifugation. Most of the single (ELLL, QLNL, and QLLN) and two of the double (QAAL and QANL)
mutants that have slightly more hydrophilic residues than does the wild type at positions L65, L70, and/or L71 assembled well with VP2. VLP2/6 obtained with these mutants were stable in CsCl density gradient. Most of the double mutants (QDNL, QDAL, QALA, and QLAA) interacted with VP2, as evidenced by coimmunoprecipitation. However, the interaction appeared to be weak and was affected during the purification process either during the extraction of Freon or by the high concen-tration of salt in CsCl density gradient. In contrast, other mu-tants of VP6 (QLNN, QDLN, and QDNN) were not coimmu-noprecipitated with VP2 when coexpressed in insect cells and did not give rise to any band after CsCl density gradient cen-trifugation. Analysis of the various mutants (Table 1) sug-FIG. 8. In vitro assembly of cores with mutated VP6 and recovery of transcriptase activity. An equivalent amount of DLPs or cores was mixed with either water or variable amounts of purified VP6 mutants, respectively, in 6 mM Mg2⫹. The mixture was incubated for 10 min at room
temperature, and then one-third of the mixture was mixed with glycerol and loaded on a 0.6% agarose gel (results shown in the upper set of panels), one-third was assayed for transcriptase activity (results shown in the graph), and one-third was analyzed by PAGE (results shown in the lower set of panels). The agarose gel was stained with a Silver Stain II kit (Bio-Rad), and the polyacrylamide gel was stained with Coomassie blue.
on November 8, 2019 by guest
gested that an alanine residue at position 65, 70, or 71 has a minimal effect on VP2-VP6 interactions. Polar or charged res-idue(s) at these three positions severely perturb VP2-VP6 as-sembly, demonstrating that hydrophobic interactions are es-sential for the assembly of VP6 through several residues. An oversimplified summary of our data could indeed be stated as follows: VP6 single mutants assemble with VP2 but double mutants do not. However, comparison of the QLAA,QALA, and QAAL mutants suggests that the last of these has the highest affinity for VP2, given that it was the only one of the
three to give rise to a single band in CsCl density gradients (Table 1). This double mutant behaved effectively like the QLNL single mutant.
It has been shown that VP6 of group C rotaviruses can assemble to VP2 of group A rotaviruses when coexpressed in insect cells, leading to the formation of chimeric, empty, dou-ble-layered VLP (22). However, group C rotavirus VP6 does not restore the transcriptase activity of group A rotavirus cores (12). The possibility of core reconstitution allowed the identi-fication of the residues essential to the transcriptase activity of FIG. 9. Electron micrographs of negatively stained in vitro reconstructed DLPs. Purified DLPs (center panel) were freed of VP6 by treatment with 1.5 M CaCl2(2), resulting in viral cores (upper left panel). The cores were reassembled with wild-type VP6 and the ELLL and QDNN VP6 mutants (upper right, lower right, and lower left panels, respectively). Note that DLPs that reassembled with wild-type VP6 and the ELLL VP6 mutant are morphologically indistinguishable from each other, although they present completely different transcriptase activities.
on November 8, 2019 by guest
DLPs. The QDLL mutant, which contains a single change in comparison with the VP6 protein from group C rotaviruses (and which would be referred to as QQLL in the notation used here), assembles and complements the core for transcription poorly, suggesting that position 65 could be critical. As sum-marized in Table 1, it is clear that the assembly of VP6 with the core is an absolute requirement for restoring transcriptase activity and that mutants that did not assemble in vitro (QDNN, QLNN, and QDLN) did not induce any activity. How-ever, the complete assembly of three VP6 mutants (ELLL, QDAL, and QDNL) with the core, which was confirmed by electron microscopy, migration in agarose gel, and sedimenta-tion in CsCl density gradient, is not sufficient to restore tran-scriptase activity. These results illustrate the complex role of VP6 in the transcriptase activity. Several hypotheses can be considered. First, VP6 could directly or indirectly, via VP2, interact with VP1. It has been shown that VP6 trimer can bind to VP1 polymerase in a far-Western assay (23). The pseudo-atomic model of the rotavirus DLP, however, shows that VP6 is not in direct contact with the polymerase. Second, it can be noted that mutants that assembled but did not induce tran-scription have an additional negative charge at the base of the trimer. The nascent mRNA transcripts generated within DLPs by endogenous transcriptase complexes have been shown to be translocated through channels at the icosahedral fivefold axes of the capsid (14). One possibility is that the extra charge at the interface between VP2 and VP6 could block the channels to the nascent mRNA. An alternative and more likely interpre-tation is that transcript extrusion requires some conforma-tional change of VP2, which otherwise closes the channel at the fivefold axes. This conformational change of VP2 is likely to be
done in a concerted manner with VP6. Addition of an extra negative charge at the VP6-VP2 contact region could therefore inhibit the necessary conformational change of VP2. A recent study of the viral transcription complex of bluetongue virus (6) clearly shows that VP7 binds nucleoside triphosphates provid-ing a substrate sink. This bindprovid-ing is associated with slight con-formational changes. VP7 also has a phosphate-binding site at the bottom part of the trimer close to the inner shell. It is possible that rotavirus VP6 has a similar role and that its conformational changes are altered when interactions with VP2 are not subtly adjusted. Further work is clearly needed to elucidate the exact function of VP6 in the large and complex transcriptional unit constituted by the DLP.
In conclusion, our study has clearly identified a series of key residues that are responsible for VP6-VP2 interactions during DLP assembly. Moreover, we have shown that subtle changes in the lower domain of VP6 are sufficient to block the tran-scription without affecting the interaction with VP2. In partic-ular, our results reveal that a single point mutation of VP6, Q32E, has no effect on particle formation but leads to a tran-scription-incompetent DLP. Finally, our results have supple-mented what is known about the atomic structure of VP6 and have generated biochemical and biological information reveal-ing the importance of hydrophobic interactions in capsid as-sembly.
ACKNOWLEDGMENTS
We thank Cynthia Jaeger and Inge Erk for their outstanding tech-nical help. We thank Carlos Arias for the D29A mutant.
This work was supported in part by a fifth PCRD grant from UE (QLRT 1999-00634) and by “Programme de Recherche Fondamentale
QALA 2 2 ⫹ ⫹ 10–50 C
QLNN 2 0 ⫺ ⫺ ⬍5 P
QLAA 2 2 ⫹ ⫹ 10–50 C
QDNN 3 0 ⫺ ⫺ ⬍5 P
aAll VP6 proteins analyzed showed trimerization and tubule assembly.
bExcept for the D29Amutant, the designations indicate the residues at positions 32, 65, 70, and 71, with substitutions shown in bold type. WT, wild type. cPresence (⫹) or absence (⫺) of coimmunoprecipitation with VP2 in Sf9 cells.
dTGA, Tris-glycine agarose gel electrophoresis. Symbols indicate presence (⫹) or absence (⫺) of a band migrating as DLPs.
eIndicated recovery of in vitro transcriptase activity as measured at the higher concentration of VP6; 100% transcriptase activity corresponds to DLPs in an amount
equal to the amount of cores present in the assay.
fVP6 was mixed with purified cores and observed by electron microscopy (EM) after negative staining. C, completely decorated and nonaggregated cores were
observed; P, partially decorated and aggregated cores were observed. See the lower right and lower left panels, respectively, of Fig. 9.
gND, not done.
on November 8, 2019 by guest
[image:9.587.40.541.84.294.2]7.Estes, M. K., and J. Cohen.1989. Rotavirus gene structure and function. Microbiol. Rev.53:410–449.
8.Gorziglia, M., C. Larrea, F. Liprandi, and J. Esparza.1985. Biochemical evidence for the oligomeric (possible trimeric) structure of the major inner capsid polypeptide (45K) of rotaviruses. J. Gen. Virol.66:1889–1900. 9.Grimes, J. M., J. N. Burroughs, P. Gouet, J. M. Diprose, R. Malby, S.
Zientara, P. P. Mertens, and D. I. Stuart.1998. The atomic structure of the bluetongue virus core. Nature395:470–478.
10.Jiang, B., H. Tsunemitsu, J. R. Gentsch, R. I. Glass, K. Y. Green, Y. Qian, and L. J. Saif.1992. Nucleotide sequence of gene 5 encoding the inner capsid protein (VP6) of bovine group C rotavirus: comparison with corresponding genes of group C, A, and B rotaviruses. Virology190:542–547.
11.Kohli, E., L. Maurice, J. F. Vautherot, C. Bourgeois, J. B. Bour, J. Cohen, and P. Pothier.1992. Localization of group-specific epitopes on the major capsid protein of group A rotavirus. J. Gen. Virol.73:907–914.
19.Prasad, B. V., G. J. Wang, J. P. Clerx, and W. Chiu.1988. Three-dimensional structure of rotavirus. J. Mol. Biol.199:269–275.
20.Reinisch, K. M., M. L. Nibert, and S. C. Harrison.2000. Structure of the reovirus core at 3.6 A˚ resolution. Nature404:960–967.
21.Ruiz, M. C., A. Charpilienne, F. Liprandi, R. Gajardo, F. Michelangeli, and J. Cohen.1996. The concentration of Ca2⫹that solubilizes outer capsid
proteins from rotavirus particles is dependent on the strain. J. Virol.70:
4877–4883.
22.Tosser, G., M. Labbe, M. Bremont, and J. Cohen.1992. Expression of the major capsid protein VP6 of group C rotavirus and synthesis of chimeric single-shelled particles by using recombinant baculoviruses. J. Virol. 66:
5825–5831.
23.Zeng, C. Q. Y., M. K. Estes, A. Charpilienne, and J. Cohen.1998. The N terminus of rotavirus VP2 is necessary for encapsidation of VP1 and VP3. J. Virol.72:201–208.