characterized genetically in previous reports are important for optimal packaging efficiency. A fragment covering a minimum of four potential stem-loop structures is required for the shortest packaging element compatible with gene transfer. Our results reveal the extent to which each of the segments of the packaging sequence contribute to packaging efficiency.
The assembly of infectious retroviral particles requires the selective encapsidation of two unspliced viral genomic RNA molecules (11, 35). This process, termed RNA packaging, de-pends upon specific sequences in the unspliced viral RNA and
in the Gag polyprotein (2, 6, 19, 32, 36). Thecis-acting signals
involved in retroviral RNA packaging include the essential
encapsidation sequences (⌿) located in the genomic RNA
between the first splice donor site and thegaginitiation codon
(35). There are additional regions of the genome that facilitate
encapsidation in the presence of ⌿, and these are located
proximal to the major subgenomic splice donor and at the 5⬘
end of thegagcoding sequence (7, 25, 26).
Packaging of retroviral RNA involves dimerization via RNA-RNA contacts as well as association with the Gag polyprotein. A palindromic element, called the kissing loop,
and stretches of guanines located within the 5⬘ untranslated
leader sequence of the RNA genome have both been sug-gested to be key elements for RNA dimerization (3, 10, 12, 13, 15, 18, 23, 27, 31, 34, 38–40). Dimerization might be linked to efficient packaging, although some retroviral packaging can occur in the absence of significant dimerization, as mutants that package monomeric genomic RNA have been observed (9, 10, 12, 17, 22, 34, 38).
The Moloney murine leukemia virus (MoMLV) packaging signal has provided an excellent system to study the determi-nants of retroviral packaging (reference 35 and references therein). In vivo and in vitro studies showed that a critical fragment of the MoMLV packaging signal lies within 520
nu-cleotides located at the 5⬘end of the MoMLV genome (5, 13,
34). Moloney murine sarcoma virus (MoMSV) is a replication defective virus that was derived from MoMLV and in which
most of the pol and envopen reading frames have been
re-placed by that of the c-mosgene. In MoMSV the encapsidation
signal has been localized to a 333-nucleotide sequence located
downstream of the 5⬘ splice donor site, corresponding to a
portion of the MoMLV ⌿ sequence. This region has been
previously shown to produce virus titers comparable to those
of wild-type MoMLV when inserted into a gene transfer vector
(1). The ⌿ sequences of MoMLV and MoMSV are almost
identical with the exception of a 50-nucleotide highly divergent
fragment located within the ⌿ sequence of MoMLV. In
MoMSV, this 50-nucleotide fragment is replaced by a 32-nu-cleotide sequence. This MoMSV fragment is virtually identical to virus-like 30S (VL30) sequences, and this hybrid MoMLV-VL30 sequence is likely to have resulted from the recombina-tion of MoMLV with the endogenous virus-like retrotranspo-son (21). VL30 can copackage with the retroviral genome, and therefore, recombination between these elements can occur (20, 45).
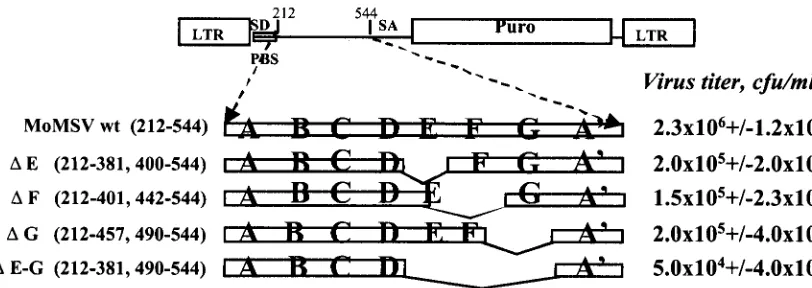
Deletion analysis of sequences included in predicted stem-loops E, F, and G. The ⌿ sequence of MoMLV folds into several stem-loop structures (28, 29, 42). When a subset of these stem-loops, named A, B, C, and D, were individually deleted, the virus titer and level of RNA packaging were re-duced 5- to 10-fold but not abolished, indicating that none of these elements is necessary for RNA packaging (29). Previous genetic studies have not focused on the contribution to RNA
packaging of sequences located 3⬘of the D stem-loop (bp 374
to 544), which were proposed to fold into additional stem-loop structures (42). Similar stem-loop structures are predicted for the corresponding fragment of the MoMSV sequence (bp 827 to 934 of the MoMSV clone 124 sequence, GenBank accession no. J02263), and we named those stem-loops E, F, and G.
To investigate the role of these sequences in RNA packag-ing, we carried out deletion mutagenesis of the proposed stem loops E, F, and G and tested these mutant viruses in a gene
transfer assay (Fig. 1). A linker containing two differentSfiI
sites replaced the MoMLV packaging sequence in plasmid pMFGpuro (37). A MoMSV packaging vector carrying the
wild-type ⌿ sequence was generated by amplifying the
MoMSV⌿fragment from plasmid pLNL6 (nucleotides 799 to
1132, BalI-PstI fragment of pLNL6, accession no. M63653,
corresponding to nucleotides 657 to 989 of the MoMSV clone 124 sequence, accession no. J02263) and cloning it into the modified pMFGpuro. To generate the deletion mutants,
por-tions of the deleted⌿sequences were amplified by using the
PCR-based splice-overlap extension method as described pre-viously (32) and cloning them into the modified pMFGpuro. The gene transfer assay was carried out by infecting CHO cells
* Corresponding author. Mailing address: Children’s Hospital, End-ers 609, 300 Longwood Ave., Boston, MA 02115. Phone: (617) 355-8426. Fax: (617) 566-4711. E-mail: [email protected].
4643
on November 8, 2019 by guest
according to previously published procedures (30) and by eval-uating the number of puromycin-resistant colonies 9 days after infection. Infectious titers of wild-type and mutant viruses were normalized according to equal amounts of reverse transcrip-tase (RT) activity in the supernatants used in infectivity assays. To evaluate the contribution to packaging of the entire se-quence predicted to fold into stem-loops E, F, and G, we generated a mutant in which the DNA fragment that includes
these stem-loops was deleted (bp 382 to 489, mutant⌬E-G)
(Fig. 1). This deletion resulted in a virus titer that was approx-imately 50-fold lower than that of the wild type, indicating that this sequence is required for high efficiency of viral packaging, although it is not necessary for virus viability.
Mutant viruses carrying a deletion of the sequences included in each of the potential stem-loops showed a reduction in virus
titer of 11- to 14-fold (mutants ⌬E, ⌬F, and ⌬G) (Fig. 1),
implying that each of these sequences contributes significantly to high-efficiency packaging. This level of reduction is similar to the reduction observed by Mougel et al. when stem-loop A, B, C, or D was deleted individually (29), indicating that the sequences included in stem-loops E, F, and G contribute to a similar degree to the packaging process and are critical to optimal RNA packaging and virus titer.
It was previously reported that when the 333-bp MoMSV fragment that we have investigated was compared to its ho-mologous fragment from MoMLV in a gene transfer assay, the titer from the MoMSV virus was 10-fold higher (1). The dif-ference was attributed to a highly divergent fragment that in MoMSV is part of the sequence that we assigned to stem-loop F and that shows high homology to sequences of VL30 (1). However, we were not able to detect significant titer differ-ences between the two viruses carrying the MoMSV and MoMLV packaging sequences when these sequences were in-serted in our gene transfer vector and transfected individually (data not shown). These data suggest that the sequences in-cluded in stem-loop F, proposed for the divergent fragment of MoMLV, are functionally equivalent to the sequences present in MoMSV (8, 14). It is possible that the different context of
these sequences might have played a role in the different out-come of the experiment reported by Adam and Miller (1) or, alternatively, that the differences in packaging efficiency of the two sequences can only be revealed in competition experi-ments (20, 41).
Identification of the minimal ⌿ domain by gene transfer analysis.Previous studies were not able to define conclusively the MoMLV or MoMSV elements sufficient for minimal pack-aging function. Stem-loops B, C, and D are virtually identical in MoMLV and MoMSV. Insertion of the 205-nucleotide frag-ment covering stem-loops B, C, and D into a 712-nucleotide fragment derived from the human immunodeficiency virus type
1 (HIV-1) leader sequence permits packaging of this hybrid⌿
sequence into MoMLV particles at 75% of the level allowed by the wild-type MoMLV packaging sequence (28). The extent of packaging function provided by this fragment in the absence of HIV-1 leader sequences was not established. In this study, Mougel and Barklis reached the conclusion that stem-loops B, C, and D are crucial for encapsidation and comprise the pack-aging core element for MoMLV. It is nevertheless unclear whether these structures can support RNA packaging and virus replication in the absence of heterologous packaging elements that may have been provided by HIV or whether additional MoMLV elements might be required.
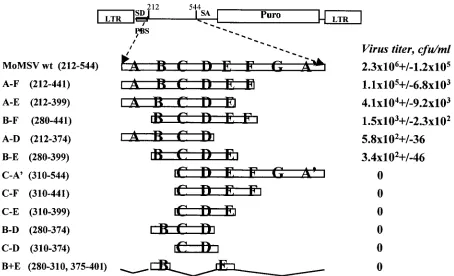
To identify the shortest MoMSV ⌿ fragment compatible
with RNA packaging and virus viability in a gene transfer assay, we generated a series of deletion mutants which
in-cluded only certain parts of the MoMSV⌿region and
evalu-ated their infectious titers in CHO cells (Fig. 2). Stem-loops C
and D were previously described as critical elements of the⌿
sequence of MoMLV (16, 28, 29). The contribution of these sequences to packaging was evaluated in mutants C-D, B-D,
C-E, C-F, and C-A⬘(Fig. 2). None of these viruses was
infec-tious. Construct C-A⬘ contains 70% of the packaging signal,
[image:2.587.87.493.75.219.2]and the mutant virus derived from it is unable to support virus replication. Thus, additional sequences upstream of stem-loop C are required to yield infectious virus and the sequences included in stem-loops C and D are not sufficient to provide
FIG. 1. Contribution of sequences at the 3⬘end of the⌿element of MoMSV to retrovirus-mediated gene transfer. The positions of the primer binding site (PBS) and splice donor (SD) and splice acceptor (SA) sites of the gene transfer vector are indicated. The deletion mutant constructs are named according to the stem-loops deleted in the constructs; numbers in parentheses indicate the nucleotides of the MoMSV sequence that are not affected by the deletion. The numbers following the mutant names indicate which nucleotides of the MoMSV fragment were amplified by PCR and inserted in the plasmid pMFGpuro⌬SfiI. The segment indicated with numbers 212 to 544 corresponds to nucleotides 657 to 989 of the MoMSV clone 124 sequence with accession no. J02263 (44). This numbering begins with the nucleotide at the viral RNA start site (43). LTR, long terminal repeat; Puro, puromycin.
on November 8, 2019 by guest
http://jvi.asm.org/
positive gene transfer. When two overlapping fragments that include sequences assigned to stem-loops A, B, C, and D (bp 212 to 374) or of stem-loops B, C, D, and E (bp 280 to 399) were investigated (mutant A-D or B-E) (Fig. 2), each
corre-sponding virus yielded a titer of approximately 102 CFU/ml.
One virus, carrying the fragment that covers stem-loops B, C, and D (bp 280 to 374), was noninfectious. Since the addition of either stem-loop B to construct C-E or stem-loop E to con-struct B-D appears in these experiments to be crucial to yield infectious virus, we generated a construct that contained only
sequences of stem-loops B and E (mutant B⫹E). The virus
from this construct was noninfectious. This suggests that
se-quences located at the 5⬘ and 3⬘ ends of C-D can provide
packaging function when they are linked to the C-D sequence and therefore, like C and D, are necessary but not sufficient.
The addition of the sequences located 5⬘of stem-loop B or in
stem-loop E to the sequence that covers stem-loops B, C, and D leads to virus titers that are very similar, suggesting that the contribution of these sequences to function is equivalent. When additional sequences are added to this core sequence, the titers of the corresponding viruses increase, indicating that each of the additional stem-loops can contribute to packaging function (mutants B-F, A-E, A-F). Only one portion of the sequence previously assigned to stem A is present in the pack-aging sequences that extend to nucleotide 212 in mutants A-D, A-E, and A-F. Therefore, it is more likely that one or both of
the two stem-loops located 3⬘of A and 5⬘of the B stem-loop,
suggested by Tounekti et al. (42), are important for the phe-notype of these mutants.
Our data indicate that multiple structural elements are re-quired in vivo for packaging function. Although sequences in stem-loop B can be deleted from the optimal sequence without losing virus viability (29), they are required when a shorter
sequence is evaluated. We found that ⌿ fragments that
in-cluded sequences assigned to stem-loops C and D (or B, C, and D) did not permit successful gene transfer when included in a viral genome. The smallest sequence compatible with virus viability includes sequences that have the potential to fold into at least four stem-loop structures.
Encapsidation of mutant viral RNAs. As the efficiency of gene transfer depends upon the virus infecting the target cell and integrating its genome into the cellular genome, the effi-ciency of gene transfer correlates well with the effieffi-ciency of viral RNA packaging. Because gene transfer also depends on successful reverse transcription and integration, viruses whose mutations affect these processes might have an infectious titer lower than the actual amount of packaged RNA.
To investigate whether our mutations affected steps other than RNA packaging, we determined the levels of viral genomic RNA present in particles by RT-PCR (32, 33). The results suggest that the MoMSV deletion mutations that we analyzed modulated viral infectivity through a reduction in the amount of encapsidated viral RNA, as we found a good, al-though imperfect, correlation between the amount of
incorpo-FIG. 2. Identification of the minimal⌿element of MoMSV that is compatible with retrovirus-mediated gene transfer. The deletion mutant constructs are named according to the stem-loops preserved in the constructs; numbers in parentheses indicate the nucleotides of the MoMSV sequence that are not affected by the deletion. The structure of the mutant⌿sequence is illustrated in the schematic representation of the mutants, and the corresponding infectious titer is reported. PBS, primer binding site; SD, splice donor site; SA, splice acceptor site; LTR, long terminal
repeat; Puro, puromycin.
on November 8, 2019 by guest
[image:3.587.66.520.73.349.2]rated RNA and the viral titer of the mutants (Fig. 1, 2, and 3B). It is unlikely that other steps of the virus life cycle, such as reverse transcription, were significantly altered in these mu-tated viruses.
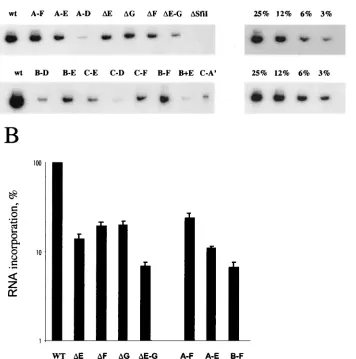
When stem-loops E, F, and G were deleted individually, the amount of incorporated viral genomic RNA was approximately
20% of that of the wild-type virus (mutants⌬E,⌬F,⌬G) (Fig.
3). When these three stem-loops were simultaneously deleted
in mutant⌬E-G, the genomic RNA incorporation was reduced
to 7% of that of the wild-type virus (Fig. 3). These data support the conclusion that the lower virus titer observed for these mutants is due to decreased genomic RNA incorporation and indicate that these sequences contribute significantly to opti-mal RNA incorporation.
We did not detect measurable amounts of RNA in any
mutants that were noninfectious (C-A⬘, B-D, C-E, C-F, and
C-D) or in the constructs that yielded the lowest viral titer
(B-E and A-D; 102 CFU/ml) when the RT-PCR assay was
carried out under our standard conditions (data not shown). This result indicated that these mutants incorporate less than 6% of viral RNA. When the total number of PCR cycles was increased, we could detect levels of RNA incorporation around 3% (which is our lowest standard) for mutants B-E and A-D
and below 3% for mutants C-A⬘, B-D, C-E, C-F, and C-D (Fig.
3A). The RNA incorporation data for mutants B-D and C-D further confirm the findings of the infectivity assay and indicate
that the⌿sequences included in these mutants do not support
incorporation of significant levels of RNA.
[image:4.587.120.476.103.462.2]RNA incorporation for MoMLV mutant viruses carrying B-D and C-D motifs was reported to be 74 and 48%, respec-tively, by Mougel and Barklis (28). In our experiments, RNA packaging for both B-D and C-D was less than 3% of the RNA incorporation observed for the wild-type mutants. The discrep-ancies between the data of Mougel and Barklis and those
FIG. 3. Relative RNA incorporation into particles by MoMSV⌿deletion mutants determined by reverse transcriptase PCR. (A) Analysis of RNA samples from pelleted virions. The results of one representative experiment are shown. (B) Genomic RNA incorporation into the wild-type and mutated virions. The histogram shows the average values and the standard errors of the means of RNA incorporation measured in three independent experiments. Mutants with RNA incorporation values that were around or below 3% (which is our lowest standard) are not reported in this histogram, as a precise calculation of the levels of RNA incorporation and the standard error was not possible. wt, wild type.
on November 8, 2019 by guest
http://jvi.asm.org/
aging signal, the HIV-1 sequences fold into elements that, although not sufficient for RNA packaging when tested alone, can provide packaging function together with the B, C, and D sequences. In our case, there were no sequences in addition to B, C, and D to support packaging function, and efficient RNA packaging and gene transfer could not be achieved with these sequences alone.
To exclude the possibility that the reduced RNA incorpora-tion by packaging mutants results from decreased intracellular transcription of the viral RNA, reverse transcriptase PCR was carried out on cellular RNA with primers specific for the MoMSV genomic RNA. We found that all of the mutants produced similar amounts of intracellular viral RNA and that, therefore, the reduced rate of packaging of the mutants results from reduced packaging efficiency (data not shown).
Phylogenetic analysis, computer modeling, and genetic anal-ysis were also used in the investigation of the minimal elements involved in the packaging of avian leukosis-sarcoma virus (4). In that study, the term minimal was used to indicate the small-est fragment that has a level of RNA incorporation similar to the wild type and not, as we use it here, to indicate a fragment compatible with the minimal level of function in a gene trans-fer assay. In avian sarcoma virus, an element composed of three stem-loops could provide a level of RNA incorporation only threefold lower than the wild-type sequence. In our case, a larger RNA fragment that spans at least seven stem-loops appears to be necessary for optimal RNA incorporation. The deletion of each of the seven stem-loops has a significant im-pact on the same parameters, reducing RNA incorporation fivefold or more (reference 29 and this report). Therefore, 82 nucleotides are sufficient for avian sarcoma virus wild-type RNA incorporation levels while a much larger fragment seems necessary in MoMSV for the same level of RNA incorporation efficiency. It is unclear why elements that are so different in their structural features carry out the same function in these different viruses. It is possible that differences in the confor-mation of the proteins they interact with might play a role in determination of the critical RNA structure.
In summary, the results obtained from the deletion analysis
extend our understanding of the MoMSV ⌿ sequence and
indicate that this sequence is composed of multiple elements, all of which are necessary for optimal packaging efficiency. This
is also true for the elements contained in the 3⬘portion of the
⌿sequence that had not been analyzed previously. A fragment
that is likely to fold into at least four stem-loop structures provides the minimal sequence that is compatible with RNA packaging and gene transfer.
minimal avian leukosis-sarcoma virus packaging signal. J. Virol.74:456–464. 5.Bender, M. A., T. D. Palmer, R. E. Gelinas, and A. D. Miller.1987. Evidence that the packaging signal of Moloney murine leukemia virus extends into the gagregion. J. Virol.61:1639–1646.
6.Clever, J., C. Sassetti, and T. G. Parslow.1995. RNA secondary structure and binding sites forgaggene products in the 5⬘packaging signal of human immunodeficiency virus type 1. J. Virol.69:2101–2109.
7.Clever, J. L., D. A. Eckstein, and T. G. Parslow.1999. Genetic dissociation of the encapsidation and reverse transcription functions in the 5⬘R region of human immunodeficiency virus type 1. J. Virol.73:101–109.
8.Clever, J. L., R. A. Taplitz, M. A. Lochrie, B. Polisky, and T. G. Parslow.
2000. A heterologous, high-affinity RNA ligand for human immunodefi-ciency virus Gag protein has RNA packaging activity. J. Virol.74:541–546. 9.Clever, J. L., M. L. Wong, and T. G. Parslow. 1996. Requirements for kissing-loop-mediated dimerization of human immunodeficiency virus RNA. J. Virol.70:5902–5908.
10.Darlix, J. L., C. Gabus, M. T. Nugeyre, F. Clavel, and F. Barre-Sinoussi.
1990. Cis elements and trans-acting factors involved in the RNA dimeriza-tion of the human immunodeficiency virus HIV-1. J. Mol. Biol.216:689–699. 11.Darlix, J. L., M. Lapadat-Tapolsky, H. de Rocquigny, and B. P. Roques.
1995. First glimpses at structure-function relationships of the nucleocapsid protein of retroviruses. J. Mol. Biol.254:523–537.
12.De Rocquigny, H., C. Gabus, A. Vincent, M. C. Fournie-Zaluski, B. Roques, and J. L. Darlix.1992. Viral RNA annealing activities of human immuno-deficiency virus type 1 nucleocapsid protein require only peptide domains outside the zinc fingers. Proc. Natl. Acad. Sci. USA89:6472–6476. 13.De Tapia, M., V. Metzler, M. Mougel, B. Ehresmann, and C. Ehresmann.
1998. Dimerization of MoMuLV genomic RNA: redefinition of the role of the palindromic stem-loop H1 (278–303) and new roles for stem-loops H2 (310–352) and H3 (355–374). Biochemistry37:6077–6085.
14.Doria-Rose, N. A., and V. M. Vogt.1998. In vivo selection of Rous sarcoma virus mutants with randomized sequences in the packaging signal. J. Virol.
72:8073–8082.
15.Feng, Y. X., T. D. Copeland, L. E. Henderson, R. J. Gorelick, W. J. Bosche, J. G. Levin, and A. Rein.1996. HIV-1 nucleocapsid protein induces “matu-ration” of dimeric retroviral RNA in vitro. Proc. Natl. Acad. Sci. USA
93:7577–7581.
16.Fisher, J., and S. P. Goff.1998. Mutational analysis of stem-loops in the RNA packaging signal of the Moloney murine leukemia virus. Virology
244:133–145.
17.Fu, W., R. J. Gorelick, and A. Rein.1994. Characterization of human im-munodeficiency virus type 1 dimeric RNA from wild-type and protease-defective virions. J. Virol.68:5013–5018.
18.Girard, P. M., H. de Rocquigny, B. P. Roques, and J. Paoletti.1996. A model of PSI dimerization: destabilization of the C278-G303 stem-loop by the nucleocapsid protein (NCp10) of MoMuLV. Biochemistry35:8705–8714. 19.Gorelick, R. J., D. J. Chabot, D. E. Ott, T. D. Gagliardi, A. Rein, L. E.
Henderson, and L. O. Arthur.1996. Genetic analysis of the zinc finger in the Moloney murine leukemia virus nucleocapsid domain: replacement of zinc-coordinating residues with other zinc-zinc-coordinating residues yields noninfec-tious particles containing genomic RNA. J. Virol.70:2593–2597. 20.Hatzoglou, M., C. P. Hodgson, F. Mularo, and R. W. Hanson.1990. Efficient
packaging of a specific VL30 retroelement by psi 2 cells which produce MoMLV recombinant retroviruses. Hum. Gene Ther.1:385–397. 21.Hodgson, C. P., R. Z. Fisk, P. Arora, and M. Chotani.1990. Nucleotide
sequence of mouse virus-like (VL30) retrotransposon BVL-1. Nucleic Acids Res.18:673.
22.Katoh, I., T. Yasunaga, and Y. Yoshinaka.1993. Bovine leukemia virus RNA sequences involved in dimerization and specificgagprotein binding: close relation to the packaging sites of avian, murine, and human retroviruses. J. Virol.67:1830–1839.
23.Ly, H., D. P. Nierlich, J. C. Olsen, and A. H. Kaplan.2000. Functional characterization of the dimer linkage structure RNA of Moloney murine sarcoma virus. J. Virol.74:9937–9945.
on November 8, 2019 by guest
24.Marquet, R., F. Baudin, C. Gabus, J. L. Darlix, M. Mougel, C. Ehresmann, and B. Ehresmann.1991. Dimerization of human immunodeficiency virus (type 1) RNA: stimulation by cations and possible mechanism. Nucleic Acids Res.19:2349–2357.
25.McBride, M. S., and A. T. Panganiban.1996. The human immunodeficiency virus type 1 encapsidation site is a multipartite RNA element composed of functional hairpin structures. J. Virol.70:2963–2973.
26.McBride, M. S., M. D. Schwartz, and A. T. Panganiban.1997. Efficient encapsidation of human immunodeficiency virus type 1 vectors and further characterization ofciselements required for encapsidation. J. Virol.71:
4544–4554.
27.Mikkelsen, J. G., A. H. Lund, M. Duch, and F. S. Pedersen.2000. Mutations of the kissing-loop dimerization sequence influence the site specificity of murine leukemia virus recombination in vivo. J. Virol.74:600–610. 28.Mougel, M., and E. Barklis.1997. A role for two hairpin structures as a core
RNA encapsidation signal in murine leukemia virus virions. J. Virol.71:
8061–8065.
29.Mougel, M., Y. Zhang, and E. Barklis.1996.cis-active structural motifs involved in specific encapsidation of Moloney murine leukemia virus RNA. J. Virol.70:5043–5050.
30.Ory, D. S., B. A. Neugeboren, and R. C. Mulligan.1996. A stable human-derived packaging cell line for production of high titer retrovirus/vesicular stomatitis virus G pseudotypes. Proc. Natl. Acad. Sci. USA93:11400–11406. 31.Paillart, J. C., L. Berthoux, M. Ottmann, J. L. Darlix, R. Marquet, B. Ehresmann, and C. Ehresmann.1996. A dual role of the putative RNA dimerization initiation site of human immunodeficiency virus type 1 in genomic RNA packaging and proviral DNA synthesis. J. Virol.70:8348– 8354.
32.Poon, D. T., G. Li, and A. Aldovini.1998. Nucleocapsid and matrix protein contributions to selective human immunodeficiency virus type 1 genomic RNA packaging. J. Virol.72:1983–1993.
33.Poon, D. T., J. Wu, and A. Aldovini.1996. Charged amino acid residues of human immunodeficiency virus type 1 nucleocapsid p7 protein involved in RNA packaging and infectivity. J. Virol.70:6607–6616.
34.Prats, A. C., C. Roy, P. A. Wang, M. Erard, V. Housset, C. Gabus, C. Paoletti, and J. L. Darlix.1990.ciselements andtrans-acting factors involved in dimer formation of murine leukemia virus RNA. J. Virol.64:774–783.
35.Rein, A.1994. Retroviral RNA packaging: a review. Arch. Virol. Suppl.
9:513–522.
36.Rein, A., D. P. Harvin, J. Mirro, S. M. Ernst, and R. J. Gorelick.1994. Evidence that a central domain of nucleocapsid protein is required for RNA packaging in murine leukemia virus. J. Virol.68:6124–6129.
37.Riviere, I., K. Brose, and R. C. Mulligan.1995. Effects of retroviral vector design on expression of human adenosine deaminase in murine bone marrow transplant recipients engrafted with genetically modified cells. Proc. Natl. Acad. Sci. USA92:6733–6737.
38.Sakaguchi, K., N. Zambrano, E. T. Baldwin, B. A. Shapiro, J. W. Erickson, J. G. Omichinski, G. M. Clore, A. M. Gronenborn, and E. Appella.1993. Identification of a binding site for the human immunodeficiency virus type 1 nucleocapsid protein. Proc. Natl. Acad. Sci. USA90:5219–5223.
39.Shen, N., L. Jette, C. Liang, M. A. Wainberg, and M. Laughrea.2000. Impact of human immunodeficiency virus type 1 RNA dimerization on viral infec-tivity and of stem-loop B on RNA dimerization and reverse transcription and dissociation of dimerization from packaging. J. Virol.74:5729–5735. 40.Shen, N., L. Jette, M. A. Wainberg, and M. Laughrea.2001. Role of stem B,
loop B, and nucleotides next to the primer binding site and the kissing-loop domain in human immunodeficiency virus type 1 replication and genomic-RNA dimerization. J. Virol.75:10543–10549.
41.Torrent, C., C. Gabus, and J. Darlix.1994. A small and efficient dimeriza-tion/packaging signal of rat VL30 RNA and its use in murine leukemia virus-VL30-derived vectors for gene transfer. J. Virol.68:661–667. 42.Tounekti, N., M. Mougel, C. Roy, R. Marquet, J. L. Darlix, J. Paoletti, B.
Ehresmann, and C. Ehresmann.1992. Effect of dimerization on the confor-mation of the encapsidation Psi domain of Moloney murine leukemia virus RNA. J. Mol. Biol.223:205–220.
43.Van Beveren, C., J. Coffin, and S. Hughes. 1985. Nucleotide sequences complemented with functional and structural analysis, p. 900–911.InT. N. R. Weiss, H. Varmus, and J. Coffin (ed.), RNA tumor viruses, vol. 2. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
44.Van Beveren, C., F. van Straaten, J. A. Galleshaw, and I. M. Verma.1981. Nucleotide sequence of the genome of a murine sarcoma virus. Cell27:97– 108.
45.Wolgamot, G., L. Bonham, and A. D. Miller.1998. Sequence analysis ofMus
dunniendogenous virus reveals a hybrid VL30/gibbon ape leukemia
virus-like structure and a distinct envelope. J. Virol.72:7459–7466.