0022-538X/06/$08.00⫹0 doi:10.1128/JVI.80.10.5074–5077.2006
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
NOTES
The High-Frequency Major Histocompatibility Complex Class I Allele
Mamu-B
*
17
Is Associated with Control of Simian Immunodeficiency
Virus SIVmac239 Replication
Levi J. Yant,
1,2Thomas C. Friedrich,
1Randall C. Johnson,
3Gemma E. May,
1Nicholas J. Maness,
1,2Alissa M. Enz,
1Jeffrey D. Lifson,
4David H. O’Connor,
1,2Mary Carrington,
3and David I. Watkins
1,2*
Wisconsin National Primate Research Center, University of Wisconsin—Madison, Madison, Wisconsin 537151; Department of
Pathology and Laboratory Medicine, University of Wisconsin—Madison, Madison, Wisconsin 537152; Basic Research Program,
Laboratory of Genomic Diversity, Science Applications International Corporation-Frederick, Inc., National Cancer Institute, Frederick, Maryland 217023; and AIDS Vaccine Program/Basic Research Program,
Science Applications International Corporation-Fredrick, Frederick, Maryland 210724
Received 8 November 2005/Accepted 1 February 2006
Particular HLA alleles are associated with reduced human immunodeficiency virus replication. It has been difficult, however, to characterize the immune correlates of viral control. An analysis of the influence of major histocompatibility complex class I alleles on viral control in 181 simian immunodeficiency virus SIVmac239-infected rhesus macaques revealed thatMamu-B*17
was associated with a 26-fold reduction in plasma virus concentrations (P< 0.001).Mamu-B*17
was also enriched 1,000-fold in a group of animals that controlled viral replication. Even after accounting for this group,Mamu-B*17
was associated with an eightfold reduction in plasma virus concentrations (P< 0.001).Mamu-B*
17-positive macaques could, therefore, facilitate our un-derstanding of the correlates of viral control.
Major histocompatibility complex (MHC) class I alleles have been associated with the control of human immunodeficiency virus (HIV) replication (1, 2, 3, 5, 13). The protective benefit of the MHC class I allelesHLA-B*27andHLA-B*57is well es-tablished, and cellular immune responses restricted by these alleles have been characterized (7, 13). Unfortunately, corre-lating particular responses with the control of viral replication has been difficult. The dual obstacles of HIV variability and allelic diversity of the human MHC class I loci confound in-terpretation of immune correlates in the rare patients who control the virus. An animal model for understanding the basis of this HLA class I-associated control of viral replication would facilitate an investigation of the underlying immune correlates of control.
The MHC-defined Indian rhesus macaque infected with sim-ian immunodeficiency virus (SIV) provides a tractable model system to discern the influence of MHC class I alleles on the replication of a pathogenic AIDS virus. Previous studies have reported associations between rhesus macaque MHC class I alleles and control of virus replication (9, 10, 11, 12). Here we investigate the effect of high-frequency MHC class I alleles in the Indian rhesus macaque on the control of viral replication after challenge with a clonal virus isolate.
High-frequency MHC class I allele associations with
chronic-phase viral control. To investigate whether particular high-frequency MHC class I alleles are linked to the control of pathogenic SIVmac239 replication, we measured postchal-lenge plasma virus concentrations in 181 Indian rhesus ma-caques from two independent cohorts. Multiple chronic-phase plasma virus time points were measured for each animal, from 10 weeks postchallenge through the time of sacrifice (range, 10 to 320 weeks postchallenge). All animals were infected with the clonal isolate SIVmac239.
Remarkably, 13 animals maintained chronic-phase plasma virus concentrations that were 1,000-fold lower than those of the cohort geometric mean. This group was significantly en-riched for four high-frequency MHC class I alleles: the closely linkedMamu-B*17andMamu-B*29(P⫽0.003),Mamu-A*02 (P⫽0.03), andMamu-A*11(P⫽0.03) (Table 1). Conversely, one allele,Mamu-B*01, was significantly underrepresented in this group. AlthoughMamu-B*01was present in 60 of the 181 animals in the infected cohort, it was not present in any of the 13 “elite controllers” (P⫽0.02) (Table 1). While 85% of the 181 animals had chronic-phase plasma virus concentrations within 2 log units of the cohort chronic-phase geometric mean of 457,000 viral RNA (vRNA) copies/ml, this group of elite controllers maintained plasma virus concentrations that were more than 3 log units lower (362 vRNA copies/ml) than the cohort chronic-phase geometric mean. In fact, the 13 elite controllers have maintained these plasma virus concentrations from week 10 postchallenge to the present (52 to 310 weeks postinfection). Interestingly, the percentage (7%) of macaques that controlled SIVmac239 replication to the lowest levels in this model system is very similar to the percentage (8%) of * Corresponding author. Mailing address: David Watkins,
Univer-sity of Wisconsin—Madison, 555 Science Drive, Madison, WI 53711. Phone: (608) 265-3380. Fax: (608) 265-8084. E-mail: watkins@primate .wisc.edu.
5074
on November 8, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 8, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 8, 2019 by guest
http://jvi.asm.org/
HIV-infected humans found to suppress their virus to compa-rable levels (6).
We performed independent statistical analyses for vacci-nated and nonvaccivacci-nated animals for all alleles and found no significant differences between plasma virus concentrations of vaccinated and nonvaccinated animals of every genotype (data not shown). While 97/181 (54%) of the overall cohort received an experimental vaccine, 6/13 (46%) of the elite controllers received an experimental vaccine.
The most dramatic enrichment of the elite controller group was for animals positive for bothMamu-B*17(Mamu-B*17⫹) and Mamu-B*29 (Mamu-B*29⫹). While 19% of the overall cohort wasMamu-B*17⫹/Mamu-B*29⫹, 69% of the elite con-trollers were positive for both of these alleles (P⫽0.003). While Mamu-B*17 binds at least 16 SIVmac239-derived peptides and
thus restricts various SIV-specific cellular immune responses (8), there is no evidence to date that the closely linked (linkage R square value, 0.87) Mamu-B*29allele encodes a gene product that binds any SIVmac239-derived peptides.
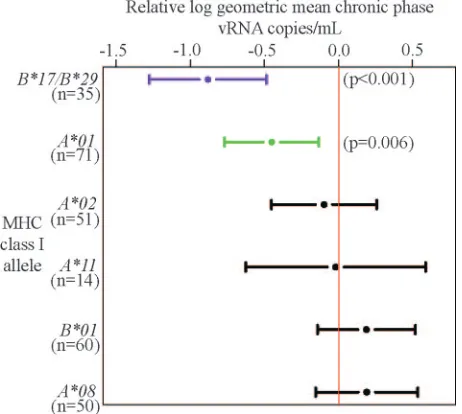
Mamu-B*17⫹/Mamu-B*29⫹animals exhibited a 26-fold de-crease in geometric mean chronic-phase plasma virus concen-trations, compared to the rest of the cohort (P⬍0.001). The
Mamu-B*17⫹/Mamu-B*29⫹animals consisted of two discrete
subgroups: the nineMamu-B*17⫹/Mamu-B*29⫹animals that controlled their chronic-phase plasma virus concentrations to below 1,000 vRNA copies/ml formed a separate distribution from the 25 Mamu-B*17⫹/Mamu-B*29⫹ animals with more typical chronic-phase plasma virus concentrations. We there-fore analyzed theMamu-B*17⫹/Mamu-B*29⫹animals, taking into account the influence of theMamu-B*17⫹/Mamu-B*29⫹ elite controller group (Fig. 1). When we accounted for these elite controllerMamu-B*17⫹/Mamu-B*29⫹ animals, we dis-covered that these two alleles were still associated with an eightfold reduction in geometric mean chronic-phase plasma virus concentrations (P⬍0.001).
Previous reports have indicated an association between the expression of Mamu-A*01 and a reduction in
chronic-phase plasma virus concentrations in animals infected with SIVmac239 (9) and SIVmac251 (10, 12) and lower lymph node-associated virus concentrations in SHIV89.6P infec-tion (14). Mamu-A*01 was associated with only slightly
[image:2.585.42.283.89.269.2]de-creased chronic-phase plasma virus concentrations in an expanded SIVmac239-infected cohort (11). Decreased chronic-phase plasma virus concentrations, however, were not ob-served inMamu-A*01⫹ cohorts infected with SIVsmE660, SHIV89.6P, or SHIVKU2 (12). In our further expanded cohorts, this allele was significantly associated with a three-fold reduction in geometric mean plasma virus concentra-tions after we took into account vaccination and elite con-troller status (P ⫽0.006) (Fig. 1).
[image:2.585.307.538.397.604.2]FIG. 1. Contribution of high-frequency MHC class I alleles to vari-ation in chronic-phase plasma virus concentrvari-ations in 181 Indian rhe-sus macaques. Changes in geometric mean (dots) chronic-phase (week 10 to present) plasma virus concentrations relative to the whole cohort (represented by the red line) are shown with 95% confidence intervals (bars). Purple,Pvalue of⬍0.001; green,Pvalue of⬍0.05 and⬎0.001. Determinations of allelic effects on chronic-phase plasma virus con-centration are given with mixed effects analyses, taking into account elite controller status, MHC class I genotype, and vaccination. All statistics were performed in R 2.2.0 with the lme4 package (R Foun-dation for Statistical Computing, Vienna, Austria). A total of 128 animals were infected at the WNPRC, and 53 animals were infected at the National Cancer Institute.
TABLE 1. Particular high-frequency MHC class I alleles are significantly overrepresented among elite controllersa
Animal or parameter
Presence of MHC class I allele or value for parameter
A*01 A*02 A*08 A*11 B*01 B*17 B*29
aj11 ⫺ ⫹ ⫺ ⫹ ⫺ ⫹ ⫹
00078 ⫺ ⫺ ⫹ ⫺ ⫺ ⫺ ⫹
01064 ⫺ ⫹ ⫺ ⫺ ⫺ ⫺ ⫺
95061b ⫹ ⫹ ⫺ ⫺ ⫺ ⫹ ⫹
95071 ⫺ ⫹ ⫺ ⫺ ⫺ ⫹ ⫹
95096c ⫹ ⫺ ⫺ ⫹ ⫺ ⫹ ⫹
96112d ⫺ ⫺ ⫹ ⫺ ⫺ ⫹ ⫹
98014e ⫺ ⫹ ⫺ ⫹ ⫺ ⫹ ⫹
98016 ⫺ ⫹ ⫺ ⫺ ⫺ ⫹ ⫹
98057 ⫺ ⫺ ⫺ ⫹ ⫺ ⫺ ⫺
99C051f ⫺ ⫹ ⫺ ⫺ ⫺ ⫹ ⫹
99x032 ⫺ ⫹ ⫹ ⫺ ⫺ ⫺ ⫺
96C114g ⫹ ⫺ ⫺ ⫺ ⫺ ⫹ ⫹
Frequency in elite controllers (%) (n⫽13)
23 62h 23 31h 0 69i 77i
Frequency in whole cohort (%) (n⫽181)
39 28 28 8 33 19 19
aAnimals that maintained chronic phase plasma virus concentrations below
1,000 vRNA copies/ml are shown. We genotyped all animals by sequence-specific PCR for seven high-frequency MHC class I alleles (4) (W. M. Rehraurer et al., unpublished data) and infected all of them with cloned SIVmac239. Significant enrichments of the elite controller group for a particular genotype are indicated in bold. Enrichment of this elite controller group for MHC class I alleles was determined with logistic regression in R 2.2.0 (R Foundation for Statistical Computing, Vienna, Austria). Animals were vaccine naive, except where indi-cated. We performed independent statistical analyses for vaccinated and nonva-ccinated animals for all alleles and found no significant differences between plasma virus concentrations of vaccinated and nonvaccinated animals of each genotype (data not shown).⫹, presence of indicated allele; ⫺, absence of indicated allele.
bDNA/MVA GagCM9 epitope only. cLipopeptide GagCM9 epitope only. dCpG in incomplete Freund’s adjuvant.
eIntramuscular injections of 107irradiated peripheral blood mononuclear
cells from animal 2055.
fSIVgagDNA5mgs/FL-ad5SIVgag. gMicrovesicles and inactivated SIV protein. hP⫽0.03.
iP⫽0.003.
VOL. 80, 2006 NOTES 5075
on November 8, 2019 by guest
http://jvi.asm.org/
It is possible that genes closely linked to Mamu-B*17and
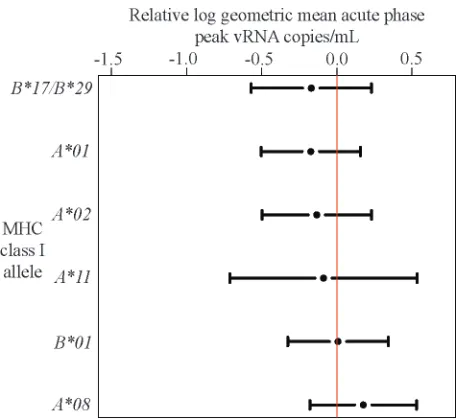
Mamu-A*01might influence the ability of the virus to replicate in vivo. There were, however, no significant differences in acute-phase peak plasma virus concentrations in animals of all genotypes before the onset of cellular immune responses (Fig. 2). This suggests that the accentuated decline from acute-phase peak to chronic-acute-phase plasma virus concentrations in
Mamu-B*17⫹ and Mamu-A*01⫹ animals was not due to a differential intrinsic susceptibility of their CD4⫹ T cells to infection with pathogenic SIVmac239 or to the enhanced effect of some non-MHC class I host restriction factor.
TheseMamu-B*17⫹animals promise to provide useful and clear-cut models for the understanding of the influence of particular MHC class I alleles on viral control. Because a comprehensive SIVmac239 epitope mapping study has been performed for Mamu-B*17 (8),Mamu-B*17⫹animals provide a unique opportunity to analyze particular CD8⫹T-cell spec-ificities linked to successful control of viral replication. For example, we can ablate the SIV-specific CD8⫹T-cell response in theseMamu-B*17⫹elite controllers with depleting antibod-ies. Additionally, since SIVmac239 has proven recalcitrant to effective neutralization by antibody responses, we can examine the effect of particular MHC class I-restricted cellular immune responses without the potentially confounding effects of neu-tralizing antibody.
A substantial proportion of captive Indian rhesus macaques express Mamu B*17. Of the Indian rhesus macaques at the
Wisconsin National Primate Research Center (WNPRC), ap-proximately 16% areMamu-B*17⫹. Given the influence of this allele on the outcome of SIV infection,Mamu-B*17⫹animals should be removed from vaccine cohorts to avoid the attribu-tion of Mamu-B*17-associated control of viral replication to
possible effects of vaccination. Simply balancing control and vaccine groups with these macaques is insufficient, because spontaneous control occurs in a sizeable subset of Mamu-B*17⫹animals. Until the precise determinants of this control are defined, the use of Mamu-B*17⫹ animals should be avoided except for analysis of the causes underlying their ex-ceptional viral control.
We gratefully acknowledge Eva Rakasz, Kirsten Bomblies, and Matt Reynolds for helpful discussions, Jess Maxwell, Crystal Glidden, and Tim Jacoby for MHC typing, and Ron Desrosiers for SIVmac239 inocula.
This work was supported by NIH grants R01 AI052056-03 and R24 RR016038-05; NIH contract HHSN26620040008C/N 01-AI-40088; the WNPRC base grant 5P51 RR 000167; federal funds from the National Cancer Institute, National Institutes of Health, under contract no. NO1-CO-12400; and by the Intramural Research Program of the NIH, National NO1-CO-12400, Cancer Institute, Center for Cancer Re-search.
The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organization imply endorsement by the U.S. Government.
REFERENCES
1.Carrington, M., and S. J. O’Brien.2003. The influence of HLA genotype on AIDS. Annu. Rev. Med.54:535–551.
2.Kaslow, R. A., M. Carrington, R. Apple, L. Park, A. Munoz, A. J. Saah, J. J. Goedert, C. Winkler, S. J. O’Brien, C. Rinaldo, R. Detels, W. Blattner, J. Phair, H. Erlich, and D. L. Mann.1996. Influence of combinations of human major histocompatibility complex genes on the course of HIV-1 infection. Nat. Med.2:405–411.
3.Kiepiela, P., A. J. Leslie, I. Honeyborne, D. Ramduth, C. Thobakgale, S. Chetty, P. Rathnavalu, C. Moore, K. J. Pfafferott, L. Hilton, P. Zimbwa, S. Moore, T. Allen, C. Brander, M. M. Addo, M. Altfeld, I. James, S. Mallal, M. Bunce, L. D. Barber, J. Szinger, C. Day, P. Klenerman, J. Mullins, B. Korber, H. M. Coovadia, B. D. Walker, and P. J. Goulder.2004. Dominant influence of HLA-B in mediating the potential co-evolution of HIV and HLA. Nature432:769–775.
4.Knapp, L. A., E. Lehmann, M. S. Piekarczyk, J. A. Urvater, and D. I. Watkins.1997. A high frequency of Mamu-A*01 in the rhesus macaque
detected by polymerase chain reaction with sequence-specific primers and direct sequencing. Tissue Antigens50:657–661.
5.McNeil, A. J., P. L. Yap, S. M. Gore, R. P. Brettle, M. McColl, R. Wyld, S. Davidson, R. Weightman, A. M. Richardson, and J. R. Robertson.1996. Association of HLA types A1-B8-DR3 and B27 with rapid and slow pro-gression of HIV disease. QJM89:177–185.
6.Mellors, J. W., A. Munoz, J. V. Giorgi, J. B. Margolick, C. J. Tassoni, P. Gupta, L. A. Kingsley, J. A. Todd, A. J. Saah, R. Detels, J. P. Phair, and C. R. J. Rinaldo.1997. Plasma viral load and CD4⫹lymphocytes as prog-nostic markers of HIV-1 infection. Ann. Intern. Med.126:946–954. 7.Migueles, S. A., M. S. Sabbaghian, W. L. Shupert, M. P. Bettinotti, F. M.
Marincola, L. Martino, C. W. Hallahan, S. M. Selig, D. Schwartz, J. Sullivan, and M. Connors.2000. HLA B*5701 is highly associated with restriction of
virus replication in a subgroup of HIV-infected long term nonprogressors. Proc. Natl. Acad. Sci. USA97:2709–2714.
8.Mothe, B. R., J. Sidney, J. L. Dzuris, M. E. Liebl, S. Fuenger, D. I. Watkins, and A. Sette. 2002. Characterization of the peptide-binding specificity of Mamu-B*17 and identification of Mamu-B*17-restricted
epitopes derived from simian immunodeficiency virus proteins. J. Immu-nol.169:210–219.
9.Mothe, B. R., J. Weinfurter, C. Wang, W. Rehrauer, N. Wilson, T. M. Allen, D. B. Allison, and D. I. Watkins.2003. Expression of the major histocompati-bility complex class I molecule Mamu-A*01 is associated with control of simian
immunodeficiency virus SIVmac239 replication. J. Virol.77:2736–2740.
10.Muhl, T., M. Krawczak, P. Ten Haaft, G. Hunsmann, and U. Sauermann.
2002. MHC class I alleles influence set-point viral load and survival time in simian immunodeficiency virus-infected rhesus monkeys. J. Immunol.169:
3438–3446.
[image:3.585.49.282.67.277.2]11.O’Connor, D. H., B. R. Mothe, J. T. Weinfurter, S. Fuenger, W. M. Rehrauer, P. Jing, R. R. Rudersdorf, M. E. Liebl, K. Krebs, J. Vasquez, E. Dodds, J. Loffredo, S. Martin, A. B. McDermott, T. M. Allen, C. Wang, G. G. Doxiadis, D. C. Montefiori, A. Hughes, D. R. Burton, D. B. Allison, S. M. Wolinsky, R. Bontrop, L. J. Picker, and D. I. Watkins.2003. Major histocompatibility complex class I alleles associated with slow simian immunodeficiency virus disease progression bind epitopes recognized by dominant acute-phase cy-totoxic-T-lymphocyte responses. J. Virol.77:9029–9040.
FIG. 2. The presence of particular high-frequency MHC class I alleles does not affect acute-phase peak plasma virus concentrations. Changes in geometric mean acute-phase peak (single highest value from weeks 1 to 3 for each animal) plasma virus concentrations relative to the whole cohort (represented by the red line) with 95% confidence intervals were estimated using linear regression. No significant associ-ations between acute-phase peak plasma virus concentrassoci-ations and MHC class I genotype were detected.
on November 8, 2019 by guest
http://jvi.asm.org/
12.Pal, R., D. Venzon, N. L. Letvin, S. Santra, D. C. Montefiori, N. R. Miller, E. Tryniszewska, M. G. Lewis, T. C. VanCott, V. Hirsch, R. Woodward, A. Gibson, M. Grace, E. Dobratz, P. D. Markham, Z. Hel, J. Nacsa, M. Klein, J. Tartaglia, and G. Franchini.2002. ALVAC-SIV-gag-pol-env-based vacci-nation and macaque major histocompatibility complex class I (A*01) delay
simian immunodeficiency virus SIVmac-induced immunodeficiency. J. Virol.
76:292–302.
13.Saah, A. J., D. R. Hoover, S. Weng, M. Carrington, J. Mellors, C. R. J. Rinaldo, D. Mann, R. Apple, J. P. Phair, R. Detels, S. O’Brien, C. Enger, P.
Johnson, and R. A. Kaslow.1998. Association of HLA profiles with early plasma viral load, CD4⫹cell count and rate of progression to AIDS follow-ing acute HIV-1 infection. Multicenter AIDS Cohort Study. AIDS12:2107– 2113.
14.Zhang, Z. Q., T. M. Fu, D. R. Casimiro, M. E. Davies, X. Liang, W. A. Schleif, L. Handt, L. Tussey, M. Chen, A. Tang, K. A. Wilson, W. L. Trigona, D. C. Freed, C. Y. Tan, M. Horton, E. A. Emini, and J. W. Shiver.2002.
Mamu-A*01allele-mediated attenuation of disease progression in
simian-human immunodeficiency virus infection. J. Virol.76:12845–12854.
VOL. 80, 2006 NOTES 5077
on November 8, 2019 by guest
http://jvi.asm.org/
0022-538X/06/$08.00⫹0 doi:10.1128/JVI.00943-06
ERRATUM
The High-Frequency Major Histocompatibility Complex (MHC)
Class I Allele
Mamu-B*17
Is Associated with Control of Simian
Immunodeficiency Virus SIVmac239 Replication
Levi J. Yant, Thomas C. Friedrich, Randall C. Johnson, Gemma E. May, Nicholas J. Maness,
Alissa M. Enz, Jeffrey D. Lifson, David H. O’Connor, Mary Carrington, and David I. Watkins
Wisconsin National Primate Research Center, University of Wisconsin—Madison, Madison, Wisconsin 53715; Department of Pathology and Laboratory Medicine, University of Wisconsin—Madison, Madison, Wisconsin 53715; Basic Research Program, Laboratory of Genomic Diversity, Science Applications
International Corporation-Frederick, Inc., National Cancer Institute, Frederick, Maryland 21702; AIDS Vaccine Program/Basic Research Program, Science Applications
International Corporation-Frederick, Frederick, Maryland 21072
Volume 80, No. 10, p. 5074–5077, 2006. Page 5074, abstract, lines 5 and 6: “Mamu-B*17was also enriched 1,000-fold in a group of animals that controlled viral replication.” should read “Mamu-B*17was also enriched in a group of animals that controlled viral replication by 1,000-fold.”