Nine years of change in the flora of Ellerburn Bank, a limestone grassland in the North York Moors
Full text
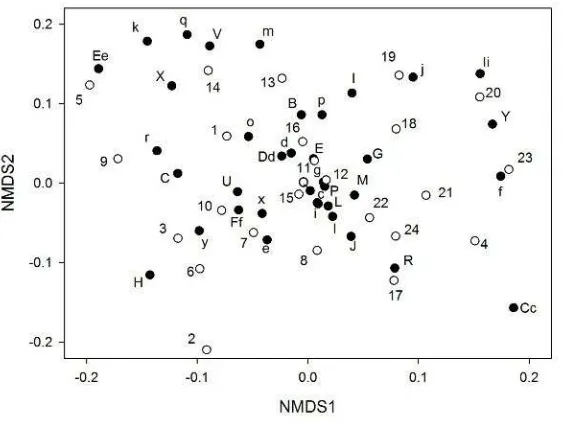
Figure




Related documents
The paper is discussed for various techniques for sensor localization and various interpolation methods for variety of prediction methods used by various applications
Field experiments were conducted at Ebonyi State University Research Farm during 2009 and 2010 farming seasons to evaluate the effect of intercropping maize with
Although formal hunting status is found significant when individually entered, the variation it explains is not significant enough (or unique enough) to be included in the
Conclusion: Our study demonstrates that midge diversity of a swamp forest can be so high that it questions global species diversity estimates for Chironomidae, which are an
It was decided that with the presence of such significant red flag signs that she should undergo advanced imaging, in this case an MRI, that revealed an underlying malignancy, which
The paper assessed the challenges facing the successful operations of Public Procurement Act 2007 and the result showed that the size and complexity of public procurement,
Мөн БЗДүүргийн нохойн уушгины жижиг гуурсанцрын хучуур эсийн болон гөлгөр булчингийн ширхгийн гиперплази (4-р зураг), Чингэлтэй дүүргийн нохойн уушгинд том
19% serve a county. Fourteen per cent of the centers provide service for adjoining states in addition to the states in which they are located; usually these adjoining states have
