0022-538X/97/$04.0010
Copyrightq1997, American Society for Microbiology
Internal Entry of Ribosomes Is Directed by the 5
9
Noncoding Region
of Classical Swine Fever Virus and Is Dependent on the Presence
of an RNA Pseudoknot Upstream of the Initiation Codon
RENE´ RIJNBRAND,1TAHARVAN DERSTRAATEN,1PIET A.VANRIJN,2WILLY J. M. SPAAN,1ANDPETER J. BREDENBEEK1*
Department of Virology, Leiden University, 2300 AH Leiden,1and Institute for Animal
Science and Health (ID-DLO), 8200AB Lelystad,2The Netherlands
Received 19 January 1996/Accepted 8 October 1996
Bicistronic RNAs containing the 373-nucleotide-long 5*nontranslated region (NTR) of the classical swine
fever virus (CSFV) genome as intercistronic spacer were used to show the presence of an internal ribosome
entry site (IRES) in the 5* end of the CSFV genome. By coexpression of the poliovirus 2A protease it was
demonstrated that the CSFV 5*NTR-driven translation is independent of the presence of functional eukaryotic
initiation factor eIF-4F. Deletion analysis indicated that the 5* border of the IRES is located between
nucleotides 28 and 66. The role of a proposed pseudoknot structure at the 3*end of the CSFV 5*NTR in
IRES-mediated translation was investigated by site-directed mutagenesis. Mutant RNAs that had lost the ability to base pair in stem II of the pseudoknot were translationally inactive. Translation to wild-type levels could be restored through the introduction of compensatory complementary base changes that repaired base pairing in stem II. In addition, we showed that the AUG codon, which is located 7 nucleotides upstream of the polyprotein initiation site and is conserved in pestiviruses, could not be used to initiate translation. Also, an AUG codon introduced downstream of the polyprotein initiation site was not recognized as an initiation site by ribosomes.
These data suggest that after internal entry on the CSFV 5*NTR, ribosomal scanning for the initiation codon
is limited to a small region.
The pestiviruses classical swine fever virus (CSFV), bovine viral diarrhea virus (BVDV), and border disease virus are grouped in the family of the Flaviviridae together with the flaviviruses and hepatitis C virus (HCV). Pestiviruses are the causative agents of several economically important animal dis-eases. The genome of pestiviruses consists of a positive-stranded RNA molecule approximately 12.5 kb in length en-coding a single polyprotein that is co- and posttranslationally processed (23, 25, 34, 35).
In contrast to the related flaviviruses, the 59 nontranslated region (NTR) of both pestiviruses and HCV is relatively long (CSFV, 373 nucleotides; BVDV, 385 nucleotides; HCV, 341 nucleotides) and contains multiple AUG codons upstream of the site used to initiate polyprotein synthesis. Several domains with a high nucleotide sequence identity have been identified within the 59NTR of the pestivirus and HCV genomic RNA (8). Moreover, similar secondary structure models have been proposed for the 59NTR (3, 13, 30), but only for HCV has the model been partially verified by biochemical analysis (3, 30).
The presence of elaborate RNA structures and AUG codons located in the 59NTR is reminiscent of the 59NTRs of picor-naviruses. For HCV (22, 28, 31) and more recently BVDV (20) it has been shown that as in picornaviruses (4, 9, 12, 19), the genome is translated in a cap-independent fashion. All these viruses use internal entry of ribosomes mediated by an internal ribosome entry site (IRES) element in their 59NTR to initiate viral protein synthesis. This is clearly different from the ribo-some scanning model (11) in which binding of the riboribo-somes to the RNA is cap dependent and scanning for the proper
trans-lation initiation codon starts at the extreme 59 end of an mRNA. The use of an IRES element to direct translation is not limited to picornaviruses and HCV, as there have been several reports on the presence of IRES elements in some eukaryotic cellular and viral mRNAs (1, 15, 18, 27).
We have previously demonstrated that with the exception of the most 59-located hairpin structure, each of the predicted hairpins of the HCV 59 NTR is essential for proper IRES functioning (22). Recently, a pseudoknot structure just up-stream of the initiator AUG has been shown to be a critical structural element of the HCV IRES (30). Such an RNA structure has also been predicted to be present at a similar position in the pestivirus 59NTR (13, 30).
In this paper we demonstrate that the 59 NTR of CSFV contains an IRES element and show that the predicted pseudoknot in the CSFV 59 NTR is an essential element for IRES function. In addition, we provide evidence indicating that the position of the translation initiation codon in the 59 NTR is of crucial importance. Ribosomes are unable to rec-ognize an AUG codon placed either 7 nucleotides or 19 nu-cleotides downstream of the inactivated authentic translation initiation codon.
MATERIALS AND METHODS
Cells and tissue culture.Hep2 and BT7-H (33) cells were grown in Dulbecco’s modified Eagle medium (Gibco BRL) supplemented with 10% fetal calf serum. BT7-H cells were cultured in the presence of 500mg (active compound) of Geneticin (Gibco BRL) per ml.
Enzymes, oligonucleotides, and bacterial strains.Recombinant DNA tech-niques were performed by standard procedures (24). Restriction endonucleases, DNA and RNA polymerases, and T4 DNA ligase were obtained from Pharma-cia, Promega, New England Biolabs, and Gibco BRL. Oligonucleotides were synthesized on a 391-DNA synthesizer (Applied Biosystems). Plasmids were transformed into Escherichia coli DH5aor MC1061.
Construction of plasmids for the expression of bicistronic mRNAs.DNA fragments containing the entire 59NTR and 14 nucleotides of the nonstructural
* Corresponding author. Mailing address: Department of Virology, Institute of Medical Microbiology, Leiden University, AZL, P.O. Box 320, 2300AH Leiden, The Netherlands. Phone: 715261652. Fax: 715266761. E-mail: [email protected].
451
on November 9, 2019 by guest
http://jvi.asm.org/
protein Nprocoding region of the CSFV Brescia (B) strain and the Chinese (C)
vaccine strain were amplified by PCR from plasmids p101-T7 (29) and pPRc129 (17), respectively. To facilitate cloning, the 59primer used in these PCRs con-tains EcoRI, BglII, and NsiI restriction sites upstream of the CSFV sequences, and the 39primer contains a KpnI site downstream of the CSFV sequences. After digestion with EcoRI and KpnI, the amplified DNA fragments were cloned in pBluescript SK(2) (Stratagene). The nucleotide sequences of the inserts were determined to verify that no unwanted mutations had been introduced during PCR.
The CSFV inserts were subsequently fused to a chloramphenicol acetyltrans-ferase (CAT) gene by replacing the HCV sequences in pWT-CAT (22) (for clarity this construct will be called pHCV-CAT) by the CSFV B and C strain
EcoRI-KpnI 59NTR/Nprofragment from the pBluescript derivatives, resulting in
pCSF/B-WTCAT and pCSF/C-WTCAT, respectively. For the construction of plasmids to express bicistronic mRNAs, pCSF/B-WTCAT and pCSF/C-WTCAT were digested with BglII and HindIII, and the CSFV 59NTR/Npro-CAT
frag-ments were cloned into BamHI-HindIII-digested pb-HCV-CAT/3 and pLuc-HCV-CAT (2). RNA transcription of the bicistronic mRNAs is under the control of a T7 RNA polymerase promoter. The resulting plasmids (pb -CSFV-B-CAT; pb-CSFV-C-CAT, pLuc-CSFV-B-CAT, and pLuc-CSFV-C-CAT) con-tain either theb-galactosidase or luciferase gene as the upstream cistron (Fig. 1). A similar set of plasmids for the expression of bicistronic RNAs was also created in the vector pET3a. Compared with the previous plasmids these constructs (Fig. 1, pET3aLuc-CSFV-C-CAT) contain a T7 transcription terminator sequence.
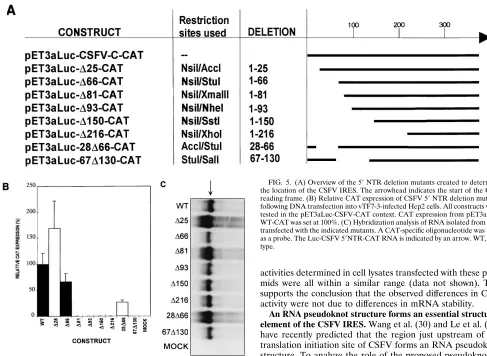
Several deletion mutants were constructed by digesting pCSF/C-WTCAT with
NsiI in combination with either AccI, XmaIII, NheI, SstI, or XhoI (Fig. 2). The
resulting ends were blunted with Klenow fragment prior to ligation. Site-directed mutagenesis was performed to introduce StuI and SalI restriction sites at posi-tions 66 and 130, respectively (Fig. 2). The StuI site was used in combination with
NsiI, AccI, and SalI as described above, which resulted in the deletion of the 59 66 nucleotides (D66) and two internal deletion mutants (28D66 and 67D130). All of the mutations described above were cloned into pET3aLuc-CSFV-C-CAT.
Plasmids pET3aLuc-Msc-CAT, pET3aLuc-DPst-CAT, and pET3aLuc-DPst/ Msc-CAT were created by PCR-based site-directed mutagenesis. The mutated sequences and their location are depicted in Fig. 6A. In plasmid pET3aLuc-DU375-CAT, the T residue at position 375 of the CSFV C strain 59NTR was deleted. Plasmid pET3aLuc-DU375/KpnAUG-CAT was created by replacing the
KpnI-BamHI CAT fragment from pET3ALuc-DU375-CAT by a KpnI-BamHI CAT fragment from pDAUGorf114AUG-CATT (21). In plasmid pDAUGorf1
14AUG-CATT, an AUG codon is inserted downstream of the KpnI site (see Fig. 7A). Polishing of the protruding ends of the KpnI site, at the border of the CSFV sequence and the CAT gene in pET3aLuc-CSFV-C-CAT, resulted in mutant pET3aLuc-DKpn-CAT.
Construction of plasmids expressing monocistronic RNAs containing the CSFV C strain 5*NTR.An EcoRI-XhoI fragment from plasmid pPRc129 (17) containing the complete 59end of the CSFV C strain genome fused to a T7 RNA polymerase promoter and a XhoI-BamHI fragment from pCSF/C-WTCAT were cloned into an EcoRI-BamHI-digested pHCV-CATT (21). The resulting plas-mid, pCSFV-CATT, has the complete CSFV C strain 59NTR/CAT engineered between a T7 transcription promoter and terminator (Fig. 1).
Transient-expression assay.Near-confluent monolayers of Hep2 cells, grown in 35-mm-diameter dishes, were infected at a multiplicity of infection of 10 with the vaccinia virus recombinant vTF7-3 (6) expressing T7 RNA polymerase. After an incubation at room temperature for 45 min, the inoculum was removed, and cells were transfected with 200ml of Dulbecco’s modified Eagle medium con-taining 5mg of CsCl gradient-purified plasmid DNA and 15ml of Lipofectin (Gibco BRL). Six hours posttransfection, cells were lysed and CAT expression was quantitated by a CAT enzyme-linked immunosorbent assay (Boehringer Mannheim). CAT values were adjusted for differences in protein concentration of the lysates. Protein concentration was determined by using a Lowry-based
protein quantification assay (Sigma). Luciferase activity was determined using a luciferase assay (Promega) to correct for differences in the transfection efficiency between different plasmids.
Poliovirus 2A protease cotransfection.BT7-H cells, an African green monkey kidney cell line expressing T7 RNA polymerase (33), were transfected as de-scribed previously (22) with pHCV-CAT (22), pRSV-CAT (7), or pCSFV-CATT and cotransfected with either pEP2A or pEP2A(H20N), which express active or inactive poliovirus 2A protease (36), respectively. At 36 h posttransfection, cell lysates were prepared and assayed for CAT activity as described above.
RNA analysis.Total RNA was isolated 8 h posttransfection by using RNAzol (Tel-Test, Inc.) as described by the supplier. RNA was separated in formalde-hyde-agarose gels (24). The gels were dried and subsequently hybridized with a CAT gene-specific 59-end-labelled oligonucleotide probe as described by Mein-koth and Wahl (16).
RESULTS
The 5*NTR of CSFV directs translation of the downstream
cistron in a bicistronic mRNA.To determine if the 59NTR of
CSFV possesses an IRES domain, we analyzed whether inser-tion of the 59 NTR of CSFV as intercistronic spacer induced translation of the downstream cistron. In this set of experi-ments we included the virulent CSFV B strain and the atten-uated CSFV C vaccine strain to analyze any differences that might be present in the efficiency of translation initiation be-tween virulent and attenuated CSFV strains. Plasmids express-ing bicistronic RNAs were constructed with either the 59NTR of the CSFV B or C strain as intercistronic spacer. In these constructs either the b-galactosidase or luciferase gene was used as the upstream cistron. The CAT gene, fused in frame with the 59end of the p23 coding region of CSFV, was used as the downstream cistron. (Fig. 1, CSFV-B-CAT, pLuc-CSFV-C-CAT, pb-CSFV-B-CAT, and pb-CSFV-C-CAT). Plasmid pLuc-ICS-CAT that contains an intercistronic spacer of similar length as the CSFV 59 NTR but lacks an IRES FIG. 1. Schematic representation of the expression vectors used to study
CSFV IRES activity. Shaded box, luciferase (Luc) orb-galactosidase (b-gal) reporter gene; open box, CSFV 59NTR sequences; solid box, 14 nucleotides of the CSFV capsid coding region; crosshatched box, CAT reporter gene; T7, T7 RNA polymerase promoter; Tf, T7 RNA polymerase transcription terminator.
BamHI, EcoRI, HindIII, KpnI, and NsiI are restriction sites used in the cloning
[image:2.612.333.532.413.666.2]procedures.
FIG. 2. Proposed RNA structure for CSFV (adapted from Brown et al. [3] and Wang et al. [30]). Components of the structure are labelled A through I. The position of the translation initiation codon of the CSFV polyprotein is indicated by AUG. Flags indicate the positions of upstream AUG codons in the CSFV C strain. Arrows indicate the positions of restriction enzyme sites used to create the deletion mutants. Sites created by site-directed mutagenesis are indicated in parentheses.
452 RIJNBRAND ET AL. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
element served as a control. Plasmid DNA was transfected into vTF7-3-infected Hep2 cells and analyzed for CAT expression. Transfection of pLuc-ICS-CAT did not result in any CAT expression above background levels (Fig. 3). Insertion of the CSFV B or C strain sequence resulted in a significant increase of CAT expression irrespective of the nature of the upstream cistron (Fig. 3, pb-CSFV-B-CAT, pLuc-CSFV-B-CAT, pb -CSFV-C-CAT, and pLuc-CSFV-C-CAT). From these experi-ments it can be concluded that the CSFV 59NTR is capable of directing translation initiation of the second reading frame in a bicistronic RNA. No significant difference in CAT expression has been observed between the levels of B and C strain 59 NTR-driven CAT expression. However, the first cistron ap-pears to have an effect on the amount of CAT protein pro-duced.
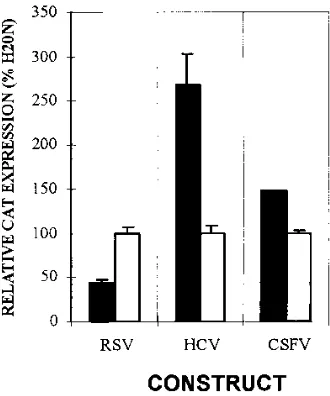
CSFV 5* NTR-driven translation is poliovirus 2A protease
resistant.The poliovirus 2A protease induces cleavage of the
p220 component of the eukaryotic translation initiation factor eIF-4F (14). Integrity of this initiation factor is essential for cap-dependent translation. To demonstrate that translation initiation driven by the CSFV 59 NTR is cap independent, CAT expression from monocistronic RNAs was tested in the presence of poliovirus 2A protease-mediated host shutoff. BT7-H cells were cotransfected with pCSFV-CATT that con-tained the CSFV C strain 59 NTR and a plasmid expressing either a functional (pEP2A) or inactivated [pEP2A(H20N)] poliovirus 2A protease. As controls, pRSV-CAT and pHCV-CAT were used in parallel transfections. The RNA encoded by pHCV-CAT contains the IRES domain of HCV and is resis-tant to translation inhibition induced by poliovirus 2A coex-pression (22). In contrast, the translation of RSV-CAT RNA, which is transcribed in the nucleus from the Rous sarcoma virus promoter, is cap dependent and therefore its translation is expected to be inhibited by coexpression of poliovirus 2A protease. The data presented in Fig. 4 demonstrate that the
uncapped CSFV-CAT and HCV-CAT RNAs are resistant to poliovirus 2A coexpression and are therefore independent of functional eIF-4F. Translation of both RNAs is in fact in-creased in the presence of poliovirus 2A coexpression. As expected, translation of RSV-CAT RNA decreased in the presence of the active 2A protease. These experiments and the studies with bicistronic RNAs clearly demonstrate the pres-ence of an IRES domain within the 59NTR of CSFV.
Hairpin B forms the 5*border of the CSFV IRES.Since for
HCV it has been shown that the IRES domain does not en-compass the entire 59NTR, we determined the 59border of the CSFV IRES domain by using several 59and internal deletion mutants. These mutants were based on restriction sites that were either present or introduced by site-directed mutagenesis in the CSFV C strain 59 NTR (Fig. 2 and 5A). Plasmids ex-pressing bicistronic RNAs were transfected into vTF7-3-in-fected Hep2 cells and analyzed for CAT expression. Deletion of the 5925 nucleotides, containing hairpin A, resulted in an increase in CAT expression compared with wild-type 59NTR (Fig. 5B). Extending this deletion to 66 nucleotides led to a significant decrease in CAT expression (Fig. 5B, D66), and when even more nucleotides were deleted, CAT expression was reduced to background levels (Fig. 5B). Deletion of hair-pin B (28D66) resulted in a 75% decrease in CAT expression (Fig. 5B), whereas deletion of hairpin C (67D130) resulted in no detectable CAT expression (Fig. 5B). These data strongly suggest that the 59 border of the CSFV IRES is located in hairpin B.
Steady-state levels of the bicistronic RNAs in vTF7-3-in-fected cells transvTF7-3-in-fected with the indicated plasmids were ana-lyzed to rule out significant differences in these RNAs that could account for the observed difference in CAT expression and thus translation initiation efficiency. RNA was isolated 8 h posttransfection and separated in denaturing formaldehyde-agarose gels and hybridized to an oligonucleotide probe di-rected against the CAT gene. As shown in Fig. 5C, cells trans-fected with the mutant CSFV 59 NTR-containing plasmids resulted in similar concentrations of CAT gene-containing RNA as cells transfected with the wild-type plasmid. These results demonstrate that the differences in CAT expression FIG. 3. Relative CAT expression in vTF7-3-infected Hep2 cells transfected
[image:3.612.353.518.69.267.2]with either pb-CSFV-C-CAT, pb-CSFV-B-CAT, CSFV-C-CAT, pLuc-CSFV-B-CAT, or pLuc-ICS-CAT. CAT expression of pLuc-CSFV-B-CAT was set at 100%.
FIG. 4. Relative CAT expression in BT7-H cells transfected with pRSV-CAT, pHCV-pRSV-CAT, or pCSFV-CAT and cotransfected with either pEP2A (black bars) or pEP2A(H20N) (white bars). For each construct the CAT expression with the cotransfection of pEP2A(H20N) was set at 100%.
on November 9, 2019 by guest
http://jvi.asm.org/
obtained with these constructs did not result from the lack of RNA or significant differences in RNA levels. The apparent higher steady-state level for some of the mutants was not reproducible and probably results from differences in transfec-tion efficiency or other experimental variables. The luciferase
activities determined in cell lysates transfected with these plas-mids were all within a similar range (data not shown). This supports the conclusion that the observed differences in CAT activity were not due to differences in mRNA stability.
An RNA pseudoknot structure forms an essential structural
element of the CSFV IRES.Wang et al. (30) and Le et al. (13)
have recently predicted that the region just upstream of the translation initiation site of CSFV forms an RNA pseudoknot structure. To analyze the role of the proposed pseudoknot in translation initiation, several mutations were introduced in ei-ther strand of stem II of this structure (Fig. 6A, constructs pET3aLuc-Msc-CAT and pET3aLuc-DPst-CAT). These muta-tions result in disruption of the predicted pseudoknot. No CAT protein could be detected upon transfection of pET3aLuc-DPst-CAT or pET3aLuc-Msc-CAT in vTF7-3-infected Hep2 cells (Fig. 6B). Next, we constructed a mutant in which the mutations of the previous two mutants were combined. T7 FIG. 5. (A) Overview of the 59NTR deletion mutants created to determine the location of the CSFV IRES. The arrowhead indicates the start of the CAT reading frame. (B) Relative CAT expression of CSFV 59NTR deletion mutants following DNA transfection into vTF7-3-infected Hep2 cells. All constructs were tested in the CSFV-CAT context. CAT expression from pET3aLuc-WT-CAT was set at 100%. (C) Hybridization analysis of RNA isolated from cells transfected with the indicated mutants. A CAT-specific oligonucleotide was used as a probe. The Luc-CSFV 59NTR-CAT RNA is indicated by an arrow. WT, wild type.
FIG. 6. (A) Mutagenesis of the predicted pseudoknot structure in the CSFV 59NTR. The proposed pseudoknot structures for CSFV C strain and the three engineered mutants are depicted. Altered nucleotides in the mutants are indicated. Nucleotides 323 and 361 are indicated for the wild type. (B) Effects of the mutations in the proposed pseudoknot structure on CAT production. CAT expression obtained for the wild-type (WT) construct was set at 100%.DPst, pET3aLuc-DPst-CAT; Msc, pET3aLuc-Msc-CAT;DPst/Msc, pET3aLuc-DPst/Msc-CAT.
454 RIJNBRAND ET AL. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.56.543.66.422.2] [image:4.612.63.562.537.687.2]RNA transcripts from this plasmid should again be able to form a similar pseudoknot structure albeit with a different nucleotide composition (Fig. 6A, pET3aLuc-DPst/Msc-CAT). Analysis of CAT expression after transfection of this plasmid showed that translation is restored to wild-type levels (Fig. 6B). RNA analysis did not reveal a significant difference in steady-state RNA levels in cells transfected with these mutant plas-mids (data not shown). These results support the presence of a pseudoknot structure and strongly suggest that the formation of stem II, but not its primary sequence, is essential for the CSFV IRES.
Translation initiation codon selection does not involve
exten-sive ribosomal scanning.In CSFV an AUG codon (AUG367)
is located 7 nucleotides upstream of the polyprotein initiation site (AUG374). This AUG codon is conserved in all
pestivi-ruses. To analyze if ribosomes are able to initiate at AUG367in
the context of the CSFV 59 NTR, the mutant pET3aLuc-DU375-CAT was constructed. In this plasmid the uridine res-idue of AUG374is deleted, which inactivates AUG374and puts
AUG367in frame with the CAT gene (Fig. 7A). CAT
expres-sion of this construct was at background levels (Fig. 7B). To exclude the possibility that AUG367is not recognized by
ribo-somes because the deleted U residue at position 375 is essen-tial for IRES activity, an additional mutant was made in which 4 nucleotides, at the 39border of the CSFV sequence and the CAT gene, are deleted (Fig. 7A,DKpn). In the resulting mu-tant, AUG367and AUG374are placed in and out of frame with
the CAT gene, respectively. CAT expression of this mutant RNA was at background levels, as was the case for the mutant pET3aLuc-DU375-CAT, indicating that AUG367 cannot be
recognized by ribosomes.
To determine whether an AUG codon at a position down-stream of AUG374can act as an initiation codon, the
KpnI-BamHI CAT fragment of pET3aLuc-DU375-CAT was re-placed by a KpnI-BamHI CAT fragment in which an AUG codon is present downstream of the KpnI site. In this plasmid (pET3aLuc-DU375/KpnAUG-CAT) the authentic initiation site is inactivated and a new AUG codon is present 20 nucle-otides downstream of it. Again no CAT expression was ob-served. From these data we conclude that if ribosomes do scan for the CSFV polyprotein initiation site then this is limited to
a region located 6 nucleotides upstream and 18 nucleotides downstream of it.
DISCUSSION
The data presented in this report show that the genome of CSFV is translated by a cap-independent and IRES-mediated mechanism. The 59387 nucleotides of both the CSFV B and C strains can direct translation of a second reading frame in a bicistronic context. Furthermore, translation of an uncapped monocistronic RNA containing nucleotides 1 to 387 of CSFV is independent of functional eIF-4F, as was shown by coex-pression of the poliovirus 2A protease.
The difference in CAT activity observed between bicistronic RNAs containing either theb-galactosidase or the luciferase gene as the upstream cistron has also been observed for the HCV 59NTR containing bicistronic RNAs (2). Possible expla-nations for this effect could be interference of theb -galacto-sidase coding region on folding of the RNA structures within the CSFV or HCV IRES domain or a difference in stability between theb-galactosidase and the luciferase gene containing RNAs.
Initially the 59NTRs of both the virulent CSFV B and the attenuated CSFV C strains were used to analyze whether the observed difference in virulence resulted from differences in translation initiation efficiency, as has been reported for the Sabin type 3 poliovirus vaccine strain (26). However, no dif-ferences in translation efficiency were observed between these CSFV strains, indicating that the attenuated phenotype of the C strain is not caused by inefficient translation initiation.
Our data clearly show that the 59border of the CSFV IRES is located between nucleotides 28 and 66. As it has been sug-gested (4) that this nucleotide sequence forms a stem-loop structure (hairpin B) (Fig. 2), it is very likely that this RNA structure is important for IRES activity. Surprisingly, deletion of both hairpin A and B had a less dramatic effect on transla-tion compared with deletransla-tion of only hairpin B. A possible explanation is that the negative effect of deleting hairpin B on IRES activity is partially neutralized by the enhancing effect of deleting hairpin A. An increase in IRES-mediated translation efficiency upon deleting hairpin A has also been observed for FIG. 7. (A) Mutants used to analyze AUG selection in CSFV. The CSFV polyprotein initiation codon and the upstream-located AUG codon are indicated as AUG374and AUG367, respectively. AUG codons in frame with the CAT gene are underlined. The locations of nucleotide deletions are indicated by dashes. (B) Relative
CAT expression obtained with wild-type (WT) and mutant constructs. CAT levels obtained for the wild type were set at 100%.
on November 9, 2019 by guest
http://jvi.asm.org/
HCV (22). As yet we have no explanation for this phenome-non. This conserved RNA structure at the extreme 59end of the RNAs of pestiviruses and HCV that is apparently not essential for IRES function can very well be required for viral RNA replication.
Recently Poole et al. (20) reported the presence of an IRES domain in BVDV, another pestivirus. They mapped the 59 border of the IRES between nucleotides 139 and 154 by hybrid arrest experiments in an in vitro translation assay. Their data showed that antisense oligonucleotides hybridizing with the BVDV analog of hairpin C did not affect translation, whereas oligonucleotides hybridizing with regions downstream of hair-pin C up to the AUG codon inhibited translation activity. Thus, although the predicted secondary structure model for CSFV and BVDV is nearly identical (4), there appear to be significant differences in the 59 RNA structures required for IRES activity. Alternatively, the observed discrepancies be-tween our data and those of Poole et al. (20) could be due to a decreased requirement for certain elements in either the IRES domain or the translation machinery in an in vitro trans-lation assay.
Downstream of hairpin C the predicted folding of the CSFV 59 NTR is very similar to the corresponding domain of the HCV 59 NTR (3, 13, 30) (Fig. 2). For HCV we have recently shown that all these structural elements are essential for proper IRES functioning (22). It is very likely that these struc-tures are also important for CSFV and BVDV. Poole et al. (20) showed that in BVDV deletion of hairpins D and E (our nomenclature) inactivated the IRES. A small but notable dif-ference in the predicted structure of the 59NTR of pestiviruses and HCV is the potential formation of an additional stem-loop structure just downstream of hairpin G in pestiviruses. Dele-tion of the sequence predicted to form this hairpin (named GII in Fig. 2) inactivates the CSFV IRES completely (data not shown), demonstrating that this predicted RNA structure or its sequence is essential for IRES activity.
Recently, evidence indicating the formation of an RNA pseudoknot just upstream of the polyprotein translation initi-ation site in pestiviruses and HCV has been presented (13, 30). Analysis of this pseudoknot in HCV revealed that both stems I and II of this structure are essential for IRES activity (30, 32). The importance of stem II in the proposed pseudoknot struc-ture in the CSFV 59 NTR was demonstrated in this study. Mutations that abolished the predicted base pairing for stem II of the pseudoknot inactivated IRES activity. However, IRES function could be restored to wild-type levels by the introduc-tion of mutaintroduc-tions that repaired base pairing in stem II. So, in conclusion, the formation of an RNA pseudoknot just up-stream of the polyprotein initiation site has been shown to be essential for IRES function in both HCV (30) and CSFV and is expected to be a critical IRES element in all the viruses closely related to HCV and pestiviruses.
The presence of an out-of-frame AUG codon located 7 nucleotides upstream of the polyprotein initiation site seems to be specific for pestiviruses since this AUG is absent from all the HCV isolates analyzed thus far. It has been suggested that this AUG codon could function as a negative regulatory ele-ment for the downstream-located polyprotein initiation site (5). The data presented in this study show that if this down regulation occurs at all, it is certainly not due to translation initiation by ribosomes at this AUG codon, since ribosomes are unable to initiate protein synthesis from this AUG codon in the context of a CSFV IRES. In addition, the ribosomes are also unable to initiate translation from an AUG codon located 19 nucleotides downstream of the inactivated, authentic trans-lation initiation site. Taken together these data suggest that in
the context of the CSFV IRES, scanning for AUG codons by ribosomes, if it occurs at all, appears to be limited to a small region surrounding the polyprotein initiation site. We have made similar observations for the selection of the translation initiation codon in HCV (21). These results indicate that this limited or even absent ribosomal scanning is an intrinsic prop-erty for the type of IRES found in HCV and pestiviruses. Recognition of the translation initiation site without scanning has also been proposed for encephalomyocarditis virus (10).
ACKNOWLEDGMENTS
We thank Adri Thomas, Gert Scheeper, and Caroline Brown for critical reading of the manuscript.
REFERENCES
1. Berlioz, C., C. Torrent, and J.-L. Darlix. 1995. An internal ribosomal entry signal in the rat VL30 region of the Harvey murine sarcoma virus leader and its use in dicistronic retroviral vectors. J. Virol. 69:6400–6407.
2. Bredenbeek, P. J., and R. Rijnbrand. 1996. Unpublished data.
3. Brown, E. A., H. Zhang, L. Ping, and S. M. Lemon. 1992. Secondary structure of the 59nontranslated regions of hepatitis C virus and pestiviruses genomic RNAs. Nucleic Acids Res. 20:5041–5045.
4. Brown, E. A., S. P. Day, R. W. Jansen, and S. M. Lemon. 1991. The 59 nontranslated region of hepatitis A virus RNA: secondary structure and elements required for translation in vitro. J. Virol. 65:5828–5838. 5. Deng, R., and K. V. Brock. 1993. 59and 39untranslated regions of pestivirus
genome: primary and secondary structure analyses. Nucleic Acids Res. 21: 1949–1957.
6. Fuerst, T. R., E. G. Niles, F. W. Studier, and B. Moss. 1986. Eukaryotic transient-expression system based on recombinant vaccinia virus that syn-thesizes bacteriophage T7 RNA polymerase. Proc. Natl. Acad. Sci. USA 83:8122–8126.
7. Gorman, C. M., G. T. Merlino, M. C. Williaghan, I. Pastan, and B. H. Howard.1982. The Rous sarcoma virus long terminal repeat is a strong promoter when introduced into a variety of eukaryotic cells by DNA medi-ated transfection. Proc. Natl. Acad. Sci. USA 79:6777–6781.
8. Han, J. V., V. Shyamala, K. M. Richman, M. J. Brauer, B. Irvine, M. S. Urdea, P. Tekam, P. Olson, G. Kuo, Q. Choo, and M. Houghton.1991. Characterization of the terminal regions of the hepatitis C viral RNA: iden-tification of conserved sequences in the 59untranslated region and poly(A) tails at the 39end. Proc. Natl. Acad. Sci. USA 88:1711–1715.
9. Jang, S. K., M. J. H. Krausslich, G. M. Nicklin, A. C. Duke, A. C. Palmen-berg, and E. Wimmer.1988. A segment of the 59nontranslated region of encephalomyocarditis virus RNA directs internal entry of ribosomes during in vitro translation. J. Virol. 62:2636–2643.
10. Kaminski, A., M. T. Howell, and R. J. Jackson. 1990. Initiation of encepha-lomyocarditis virus RNA translation: the authentic initiation site is not se-lected by a scanning mechanism. EMBO J. 9:3753–3759.
11. Kozak, M. 1989. The scanning model for translation: an update. J. Cell Biol. 108:229–241.
12. Kuhn, R., N. Luz, and E. Beck. 1990. Functional analysis of the internal translation initiation site of foot-and-mouth disease virus. J. Virol. 64:4625– 4631.
13. Le, S., N. Sonenberg, and J. V. Maizel, Jr. 1995. Unusual folding regions and ribosome landing pad within hepatitis C virus and pestivirus RNAs. Gene 154:137–143.
14. Lloyd, R. E., M. J. Grubman, and E. Ehrenfeld. 1988. Relationship of p220 cleavage during picornavirus infection to 2A proteinase sequencing. J. Virol. 62:4216–4223.
15. Macejak, D. G., and P. Sarnow. 1991. Internal initiation of translation me-diated by the 59leader of a cellular mRNA. Nature 353:90–94.
16. Meinkoth, J., and G. Wahl. 1984. Hybridization of nucleic acids immobilized on solid supports. Anal. Biochem. 138:267–284.
17. Moormann, R. J. M., H. G. P. van Gennip, G. K. W. Miedema, M. M. Hulst, and P. A. van Rijn.1996. Infectious RNA transcribed from an engineered full-length cDNA template of the genome of a pestivirus. J. Virol. 70:763– 770.
18. Oh, S.-K., M. P. Scott, and P. Sarnow. 1992. Homeotic gene Antennapedia mRNA contains 59-noncoding sequences that confer translational initiation by internal ribosome binding. Genes Dev. 6:1643–1653.
19. Pelletier, J., and N. Sonenberg. 1989. Internal binding of eukaryotic ribo-somes on poliovirus RNA: translation in HeLa cell extracts. J. Virol. 63:441– 444.
20. Poole, T. L., C. Wang, R. A. Popp, L. N. D. Potgieter, A. Siddiqui, and M. S. Collett.1995. Pestivirus translation initiation occurs by internal ribosome entry. Virology 206:750–754.
21. Rijnbrand, R., G. E. Abbink, P. C. J. Haasnoot, W. J. M. Spaan, and P. J. Bredenbeek.The influence of AUG codons in the hepatitis C virus 59
non-456 RIJNBRAND ET AL. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
translated region on translation and mapping of the translation initiation window. Virology, in press.
22. Rijnbrand, R., P. Bredenbeek, T. van der Straaten, L. Whetter, G. In-chauspe, S. Lemon, and W. Spaan.1995. Almost the entire 59non-translated region of hepatitis C virus is required for cap-independent translation. FEBS Lett. 365:115–119.
23. Rumenapf, T., G. Unger, J. H. Strauss, and H.-J. Thiel. 1993. Processing of the envelope glycoproteins of pestiviruses. J. Virol. 67:3288–3294. 24. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: a
laboratory manual, 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
25. Stark, R., G. Meyers, T. Rumenapf, and H.-J. Thiel. 1993. Processing of pestivirus glycoprotein: cleavage site between autoprotease and nucleocapsid protein of classical swine fever virus. J. Virol. 67:7088–7095.
26. Svitkin, Y. V., N. Cammack, P. D. Minor, and J. W. Almond. 1990. Trans-lation deficiency of the Sabin type 3 poliovirus genome: association with an attenuating mutation C4723U. Virology 175:103–109.
27. Thiel, V., and S. G. Siddell. 1994. Internal ribosome entry in the coding region of murine hepatitis virus mRNA 5. J. Gen. Virol. 75:3041–3046. 28. Tsukiyama-Kohara, K., N. Iizuka, M. Kohara, and A. Nomoto. 1992.
Inter-nal ribosome entry site within hepatitis C virus RNA. J. Virol. 66:1476–1483. 29. van Rijn, P. 1995. Unpublished data.
30. Wang, C., S. Le, N. Ali, and A. Siddiqui. 1995. An RNA pseudoknot is an essential structural element of the internal ribosome entry site located within the hepatitis C virus 59noncoding region. RNA 1:526–537.
31. Wang, C., P. Sarnow, and A. Siddiqui. 1993. Translation of human hepatitis C virus RNA in cultured cells is mediated by an internal ribosome binding mechanism. J. Virol. 67:3338–3344.
32. Wang, C., P. Sarnow, and A. Siddiqui. 1994. A conserved helical element is essential for internal initiation of translation of hepatitis C virus RNA. J. Vi-rol. 68:7301–7307.
33. Whetter, E. L., S. P. Day, O. Elroy-Stein, E. A. Brown, and S. M. Lemon. 1994. Low efficiency of the 59nontranslated region of hepatitis A virus RNA in directing cap-independent translation in permissive monkey kidney cells. J. Virol. 68:5253–5263.
34. Wiskerchen, M. A., S. K. Belzer, and M. S. Collett. 1991. Pestivirus gene expression: the first product of the bovine viral diarrhea virus large open reading frame, p20, possesses proteolytic activity. J. Virol 65:4508–4514. 35. Wiskerchen, M. A., and M. S. Collett. 1991. Pestivirus gene expression:
protein p80 of bovine viral diarrhea virus is a proteinase involved in polypro-tein processing. Virology 184:341–350.
36. Yu, S. F., and R. E. Lloyd. 1991. Identification of essential amino acid residues in the functional activity of poliovirus 2A protease. Virology 182: 615–625.
![FIG. 2. Proposed RNA structure for CSFV (adapted from Brown et al. [3]and Wang et al. [30])](https://thumb-us.123doks.com/thumbv2/123dok_us/1273149.79673/2.612.333.532.413.666/fig-proposed-rna-structure-csfv-adapted-brown-wang.webp)