0022-538X/93/127684-06$02.00/0
Copyright © 1993, American SocietyforMicrobiology
Multiple,
Distinct trans-Activation Functions Are Encoded by
the Simian Virus 40 Large
T
and Small t Antigens, Only
Some of Which
Require the 82-Residue
Amino-Terminal Common Domain
MARY R. LOEKEN
Joslin DiabetesCenterand Harvard Medical School, OneJoslin Place, Boston, Massachusetts02215 Received24May1993/Accepted 15 September 1993
Simian virus 40 (SV40) small t and large Tantigens can each transactivate the adenovirus (Ad) E2A and
the Ad VA-I promoters. The first 82 amino acids of large T and small t are identical. However, this large
T-small t commondomain between residues 1 and 82 does not transactivate, suggesting that large T and small
teach encode separate trans-activation functions. To determine whether the large Torsmall tuniquedomains,
which are required for trans activation of the E2A promoter, are sufficientfor thisactivity, we have employed
expression plasmids separately encoding the common and unique domains of large T and small t.
Cotransfection ofa large Tunique domainexpression plasmid efficiently transactivated the E2A promoter.
Optimaltransactivationby large Trequiredthemotif thatbinds cellularproteinssuchastheretinoblastoma
gene product, which is located in the large Tunique domain, andadditional largeT structures outside this
motif.In contrast,the small tunique domain did not trans activate the E2A promoter.Experimentsutilizing
E2A promoter mutantscontaining only the ATF- or EIIF-binding sites demonstrated that transactivation by
small t involvesonly the EIIF transcription factor and that this function requiresboth thecommon (residues
1to82) and the small t uniquedomains expressed as a colinear protein. transactivation by large T, in contrast,
involves at least three mechanisms. There appear to be at least two mechanisms that involve the EIIF
transcription factor, at least oneof which does not require the common domain (residues 1 to 82) and one
mechanism thatinvolves the ATF factor and does require both the common and the large Tunique domains.
The simian virus 40 (SV40) genome encodes two early protein products, large T and small t antigens (large T and small t, respectively), that are generated from the same
primary transcript (32). As a result of differential splicing, these proteins contain identical 82-residue amino-terminal domains (which are referredto here asthe 1-82 or common
domain),whereas thecarboxy-terminal domainsareuniqueto
eitherlargeT orsmall t (theseare referred to here asunique domains) (Fig. 1). We have previously reported that during transient transfection ofplasmid DNA, SV40 largeTandsmall tantigenscaneach transactivatetheadenovirus(Ad)E2Aand
VA-I promoters (17). Structures that are unique to large T
(residues83 to 708
[T/u])
or tosmallt(residues83 to 192[t/u])are required for trans activation of the E2A promoter, since insertion ofastopcodon inplace of aminoacid83producesa
stable common-domain protein product that does not
stimu-late chloramphenicol acetyltransferase (CAT) activity
con-trolled by the E2A promoter
(17).
This suggests that transactivation by large Tand small t does not occur bya shared
mechanism activatedbytheircommonamino-terminal domain
but occurs by distinct mechanisms activated by structures
includingsequencesfrom withineach of their unique domains. We wished to determine the biochemical mechanism by
which small t activates transcription and to establish whether this activity can be distinguished from those of other trans
activators. We have previously found that the effects of smallt
can be distinguished from those oflargeT onthe SV40 late
promoter (5, 17) and ofthe Ad ElA 289- or 243-amino-acid
products on the Ad E2A promoter (16). Therefore, study of
transcription regulation bysmall t may elucidatenovel
regula-toryprocesses that would not be revealed withother systems.
Inaddition, itis not clear whether the amino-terminal domain
shared by both large T and small t is required for the trans-activation activities of either protein. Mutations within
thecommondomain have beengenerated whichdemonstrate
thatthisregionisimportant for largeTstabilityanditscellular
transformation activity (20). Forthe present study, we asked whether small t or large T structures that are necessary for trans activation are sufficient when expressed as separate
molecules. Furthermore,we employedreporterplasmids con-taining mutations ineither the ATFortheEIIF
transcription
factorbinding sitestodetermine howmanylargeT-and small
t-specific
trans-activation processesmight
be involved inregu-lationof the E2A promoter and whether anyofthese processes
mightbe sharedby large Tand small t.
Toaddressthesequestions,weemployed plasmidsencoding
large T cDNA (5), the 82-residue large T-small t common domain (5), or the large Tunique domain
(22)
all under the control of theRoussarcomavirus(RSV)
long terminalrepeat. Ithaspreviously been demonstrated that thecommondomain isexpressedas astableprotein product (17)and that thelargeTunique domain(T/u)appearstobeless stable thanwild-type large T (22). We also constructed anexpression plasmid that
would encode the small t unique domain by using the poly-merase chain reactionto
amplify
the small tunique
domain coding sequence. As shown in Fig. 2, by using amonoclonalantibodywhichrecognizesstructureswithin the smalltunique domain (21), a protein product with the molecular weight expected of the small t
unique
domain(t/u)
was detected byWestern blot analysis following transient transfection of
ex-pression plasmids into CV-1 cells. Expression of full-length
small tbythe plasmid containingthe smalltcDNA sequence
under the control of the
promoter-enhancer employed
forexpressionof the smallt
unique
domainwascomparabletothe 7684on November 9, 2019 by guest
http://jvi.asm.org/
T-unique (T/u)
.gm
174
u)
FIG. 1. Diagrams ofprotein products ofSV40large T andsmallt. As aresult ofdifferentialsplicing ofaprimary transcript, largeTand small thave anidentical 82-residue commonamino-terminal domain (solid area) andunique carboxy-terminal domains (largeTresidues83
to708 [diagonal striped area] and small tresidues 83 to 174
[cross-hatchedarea]).Theplasmids used in this study expressed only large T, the 1-82common domain, the largeTunique domain, smallt, orthe smalltunique domain.
Y v
-S
E
0 :
4-z
0*
14.3
level of small t expressed by the control plasmid, pRSV-T/t
(which contains the entireSV40 early coding region controlled
by the RSVlong terminal repeat [19]). That the small tunique
product does not appear tobe as abundant as thefull-length
small t protein may mean that the 1-82 common domain is
important for the stability of the small t unique domain just as
it isfor the largeTunique domain (20, 22).
trans activation of the E2A promoter by large T does not
require the commondomain, whereas trans activation by small
tdoes. The abilities of thelarge T or small t unique domains to
trans activatethe wild-typeAd E2A promoter linked to a CAT
reporter gene(18) were tested (Fig. 3A). As expected,
cotrans-fection ofcontrol plasmids encoding Ad EIA or SV40 large T
stimulated pE2A-w.t. CAT activity. Cotransfection of a
plas-mid encodingthe large Tuniquedomain(T/u)also stimulated pE2A-w.t. CAT activity. Since the large T unique domain, even
when expressed without the common domain, binds cellular
proteins such as the retinoblastoma gene product (RB) and
p107(22) thatarefound incomplexes with theEIIF
transcrip-tion factor (7, 10, 29),trans activation by the large T unique
domain could occur upon binding to any of these
EIIF-associated proteins. However, RB or p107 binding does not
appear to be the only mechanism by which large T trans
activates the E2A promoter, sinceaplasmid encodingalarge
T cDNAwith apoint mutation that destroys the binding site
for cellularproteins such as RB and p107 (T/Kl) can also trans
activate the E2A promoter. However,there is aslight
reduc-tion in the trans activation of the E2A promoter by the KI
mutant.Since this mutation doesnotaffect the stability oflarge
T (15), this suggests that the large T structure that binds
cellularproteins such asRB andp107 isnecessary forpartof
thetrans-activation function oflargeT, atleast withregard to
theE2A promoter. Thus, thelargeTunique domain, which is
necessaryfortransactivation, issufficient. Furthermore, there
appear to be structures both within the RB/p 107-binding
pocketandoutside it thatparticipatein transactivation of the
E2A promoter. Aspreviously reported (17),the 1-82common
domain could not trans activate the E2A promoter, while
6.2
B
_ 0)0 _
_ m
z w
'E
[image:2.612.58.297.77.182.2]C
FIG. 2. (A) Western blot analysis of products of plasmids encoding
small torthe small tunique domain. The plasmid encoding small t
(pSRaB-t/cDNA)wasconstructedbyLaurenSompayrac byinsertinga
small tcDNAsequence (kindly provided by Kathleen Rundell) into the plasmid pSRotB, which is a modified pSRo vector (31). The plasmid encoding the small t unique domain (pSRaB-t/u)was
con-structed by polymerase chain reaction amplification of the small t
unique domain with primersdesigned togeneratea translation
initi-ation site andto allow insertion into the pSRoaB expression vector.
Plasmid sequences were verified by dideoxy sequence analysis of double-stranded DNA (27). To examine steady-state levels of the products of these plasmidsupontransienttransfection, monkey CV-1 cells in 60-mmculturedishesweretransfectedwith 10 ,ug of plasmid
DNA and30,ug of calf thymus carrier DNA by the calcium phosphate coprecipitation method.Forty-eight hours after transfection, the cells
were harvested andanalyzedasdescribed elsewhere (6) with 2 ,ug of
smallt-specific Pab 280 (Oncogene Science) and horseradish peroxi-dase-coupled sheep anti-mouseimmunoglobulin G (Amersham). Asa
control for small t expression, a plasmid encoding the entire SV40 early region controlled by the RSV long terminal repeat(pRSV-T/t)
wasemployed. The migration positions of molecular weight markers areindicated.(B) Western blot analysis of the products of the plasmids
encodingthe smalltuniquedomainorwild-type smallt.The
concen-tration of the plasmid encoding the small t unique domain was
transfectedatahigher concentration than that of the plasmid encoding
wild-type small t to see whether increased levels of the t/u product
could be achieved. Cells were transfected and cell extracts were
assayed as described forpanel A,except that 40 ,ug of the plasmid encodingthe smalltunique domainor10,ugof theplasmid encoding wild-type small tplus 30 p.gof calfthymus carrier DNAwasused for
transfection.
1 82
1-82 LargeT
smallt
1 82
1-82 Vunique (Vt
A
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.70.283.288.688.2]A
4
*a
w-.- - X
-e
B
...
*----*-
040 1404
_ _ Tt~+
0
FIG. 3. CATactivities of pE2A-w.t. CAT.(A) CV-1 culture disheswere transfectedasdescribed elsewhere
of pE2A-w.t. CATalone (-) orwith2 p.gof plasmids
(13), large T cDNA(T), large Tuniquedomain (T/u), withapointmutationwithintheregionbetween residue (15), the large T-smalltcommondomain (1-82), small tunique domain (t/u), and carrier DNAto maintain a
DNA concentration of 15 jLg per plate. Forty-eigh
transfection, one-half of the cell extractwasassayed fo for I h with ['4C]chloramphenicol and acetyl coenzym
weretransfectedasdescribed for panel A,exceptthat 2(
plasmidencoding the small t uniquedomain wasempi
cells cotransfected with small t plus t/u plasmids (t transfected with 2jig of the small t and 1(1 ,ugof the ,
domainexpressionplasmids. In allcases,the total DNA
waskeptconstant at 15 jLgperplate with carrierDNA
full-length small tcould. However, unlike the lar
domain, the small tunique domain (t/u) did nott
the pE2A-w.t. CAT plasmid.
It was not totally unexpected that trans activ
small tuniquedomain might be reduced, since ita
less stable than wild-type small t.Wetestedwheth
theamountoftheplasmid encoding the smallt un
transfccted into cells might raise the cellular con(
the small tunique domaintoaconcentration suffic
in trans activation. As shown in Fig. 2B, transfecti
expression plasmid at a concentration four times that ofthe
wild-type small t plasmid generated a steady-state concentra-tion of the small t unique domain that was similar to that of
wild-type
small t.However,
even afivefold-higher
concentra-tion of the small tunique
domainplasmid
(10
compared
with2
pg)
wascompletely inactive with regard to transactivation ofthe
pE2A-w.t.
CATplasmid (Fig. 3B).
In contrast, transI
activationby
thelarge
Tunique
domain,
whosestability
is alsoreduced compared with full-length large T, did not appear to
be dramatically compromised. Thus, the common domain is
P _* required fortrans activation by small t but not by large T. We
considered that the common domain might be necessary for trans activation
by
small t either because it influences the * * conformation of the small t unique domain (but not of thelarge T unique domain) required for this activity or because
the common and unique domains together form a structure
that complexes with a cellular factor(s) involved in trans
activation of the E2A promoter. If a cellular factor(s) does
complex with both ofthe small t domains, it must do so only
when they are expressed as one contiguous molecule, since
cotransfection of plasmids encoding the common and unique
domains on separate plasmids did not stimulate pE2A-w.t.
CATactivity (Fig. 3B). Finally, cotransfection ofan excessof
plasmid encoding the small t unique domain did not inhibit
trans activation bysmall t, at least when theconcentrationsof
plasmids which we were able to transfect together were used
(Fig. 3B), suggesting that the small t unique domain without
the common domain does not titrate a limitingcellular factor
involved in trans activation.
Involvement of large T and small t domains inactivation of
specific E2A promoter-binding transcription factors. Full
ac-tivity of the Ad E2A promoter appears to beregulated within
a 55-nucleotide enhancerelement containing a single binding
site for the transcription factor ATF and two invertedbinding
sites for thetranscription factor EIIF(18). We havepreviously
determined that transactivation of the E2A promoterby small
t involves the binding sites only for EIIF (16). The results of
the experiment shown in Fig. 3A demonstrate that both the
cells in 35-mm common and the unique domains are required for trans
(18)with 2 jig activationbysmall t. Fromthis,weconclude thatwe can detect
encodingEIA one trans activation function of small t that requires an
large T cDNA interaction of the common and unique domains and affects the
es 105and 114 EIIF transcription factor. However, we have previously
deter-t,orthe small mined that trans activation bycoexpressed largeTand small t
constant total involves both the ATF and the EIIF transcription factors (18). it hours after This suggests that trans activation of the E2A promoter by
e
A.
(B) Cellslarge
T involves ATF and could involve EIIF. To determineor 10jlgof the which of the transcription factors that
regulate
the E2Aloyed, and the promoter are targeted by large T and which domains of
large
t+ t/u) were Tare responsible for activating these transcription factors,we
small t unique employed CAT plasmids containing mutations in the ATF or
concentration the EIIF site of the E2A promoter (18). As shown in Fig. 4,
CAT activity was stimulated upon cotransfection ofa plasmid
encoding only the large T unique domain (T/u) with the
plasmid containing mutations in the ATF site. Thus, the
rge T unique common domain is notrequired for activation of EIIF bylarge
trans activate T, whereas the common domain is required for activation of
EIIF by small t. As discussed above, trans activation of an
,ation by the EIIF-dependent promoter by the large T unique domain is not
appears to be unexpected, because the binding site forEIIF-associated
pro-erincreasing teins maps within the large Tunique domain. However, trans
iiquedomain activation of the EIIF-dependent promoter by the large T
centration of mutant (T/K1) that does not bind RB, p107, and related
,ientto result proteins is only slightly reduced when compared with that of
ion ofthe t/u wild-type large T, suggesting that additionaldomainsof large T
Adlabb,
IP
4
lie
*,
4.- 0 Z)
4.- coI Z)
v- -A-
1-
on November 9, 2019 by guest
http://jvi.asm.org/
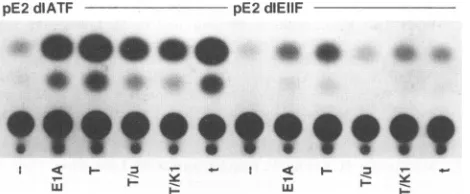
[image:3.612.66.301.75.455.2]pE2dIATF pE2dIEIIF
FIG. 4. CAT activity of E2 promoter mutants. Cells were
trans-fected as described in the legend to Fig. 3, except that the CAT plasmids employed contained mutationsatthe ATF site(and hence containedwild-typeEIIFsites)orthe EIIF sites(andhence contained
awild-type ATF site). Extracts ofcells transfected withpE2 dlATF
wereassayed for 2 h, and extracts of cells transfected with pE2dlEIIF
were assayed for4 h.
outside theRB/plO7-binding pocket participateinactivation of EIIF-dependenttranscription.
Study oftrans activation of the E2A promoter containing only the ATF site (pE2
dlEIIF)
isdifficult because of theweakstimulation of the ATF site. Whereas activation of the E2A
promoter containing only the EIIF sites by EIA, largeT, or
smallt is
consistently
10-to20-fold(Fig.
4) (18), activation ofthepromotercontaining only the ATF site is usuallynot more
than 5-fold
(18).
In theexperiment
shown in Fig. 4,cotrans-fection ofplasmids encoding EIAorlargeT stimulated CAT
activity
from pE2 dlEIIFeight-
and sevenfold, respectively.Thecommon domain appears toberequired for activation of
the ATF
site,
since cotransfection of theplasmid
encoding thelarge
Tunique
domain did not stimulate CAT activity from pE2 dlEIIEF. Interestingly, cotransfection with the plasmid encoding largeTwithamutationintheRB-binding site (T/K1) stimulated CATactivity only
3.8-fold(about
halfaseffectivelyas wild-type large
T),
suggesting that the same largeT struc-ture that is involved in activation of EIIF is also involved inactivation ofATF. Aspreviously reported (16),small tdidnot transactivateapromotercontaininganATFsite(only twofold activationwas observed in the experiment shown, which is in
the range of
experimental
variation normally observed).The results of these experiments suggest that there are at
leastfour trans-activation functions encoded in the SV40 early
genes that can be detected by assaying gene expression
di-rected by the Ad E2A promoter. There is one function
encodedby smalltthat
requires
both thecommonanduniquedomains and involves the EIIF transcription factor. Unlike large T, Ad EIA, or human papillomavirus E7, which are
thought
toactivateEIIF-dependent transcription
by bindingto RB or ap107-cyclin
A complex and thereby releasing free,activeEIIF
(2,
3, 7, 8,23),
small tdoesnotbindtoanyknownEIIF-associated factors. Thus, small t must employ a
mecha-nism distinct from that oflargeT toactivate EIIF-dependent
transcription.
It is conceivable that trans activation bysmall t occursuponbinding
toandinhibiting
the cellularenzyme, type 2Aphosphatase (PP2A) (24, 28, 33, 35), perhaps bysuppress-ing the dephosphorylation of cellular factors that associate
withEIIFonlyin theirunderphosphorylated state. Ithas been
suggested
that both the common and the small t unique domainsparticipate
in PP2Abinding
(25). Our results,whichindicate that these domainsarealsorequired fortrans
activa-tion,
areconsistent with amodel inwhich both of the small t domains arerequired
to bind to and inhibit PP2A activity,which results instimulation ofEIIF-dependent transcription.
Although
it has been difficult tostudy
thelinkage
betweenPP2A binding and trans activation because many small t
mutants are unstable (4, 14), study of stable small t mutants
will benecessary to test thismodel.
There appear to be at least three separate functions encoded
inlargeTthatwe candetectduringtransactivation of theE2A
promoter. One of these functions involves theATF
transcrip-tionfactor andrequiresthe common domain in addition to the
large T unique domain. This suggests either that a factor(s)
involved in activation of ATF binds to large T within a
structureformedby boththe commonand theuniquedomains
orthat the commondomaindirects a conformation of the large
T unique domain which activates ATF. Since activation of ATF-dependent transcription by large T is reduced upon
mutation of the RB/plO7-binding site within the unique
do-main, it is possible that large Tforms a structure inwhich a
factor(s) that regulates ATF contacts both the common
do-main and the RB-binding pocket. Very little is known about
regulation of ATF-dependent transcription to explain how it might be activated in response to large T. Since ATF is a
member of aleucinezipper family of proteins including cyclic
AMPresponseelement-binding (CREB) proteins whose
activ-ities are regulated by phosphorylation, it is possible that the
phosphorylated state of ATF affects its ability to direct
tran-scription. Phosphorylation of bacterially expressed ATF by
microtubule-associated kinase stimulates DNA binding by
ATF (1). It is possible that large T directly or indirectly regulatesakinase activity that phosphorylates ATF.
With regard to the EIIF sites, the common domain is not
required for trans activation by large T. Mutation of the
RB-p107-binding site reduced trans activation by large T but
didnoteliminate it. Since theKI mutantoflargeTappears to
be completely defective for binding ofRB, pIO7, and related proteins (9, 11, 15a), thissuggeststhat there are at least two waysforlargeT toactivate EIIF-dependenttranscription,one
of which involves the RB/p107 pocket and one of which involveslarge T structuresoutside the
RB/p1O7
pocket. EIA has been found to induce the phosphorylation of the RBprotein independently of its association with RB (34). Coex-pression of SV40 large T and small t also stimulated phos-phorylation of RB. Since EIIF appears to complex with
underphosphorylated formsof RB (7), large Tcould conceiv-ably activate EIIF-dependent transcription without bindingto RBby activating an RBkinase.
Itisinterestingto notethat when Zhu et al.tested the effects
ofseveral mutations in largeT on transactivationofthe RSV
andSV40 latepromoters,they also observedthat structures in
the commondomain and structureswithin the large Tunique
domain involved in DNAbindingorzinc bindingwere
neces-sary for optimal trans activation (36). However, the
RB-binding site was not necessary for trans activation of these
promoters. Thus, trans activation of the RSV and SV40 late
promotersby largeTmayinvolvesome common mechanisms
and some mechanisms distinct from those involved in trans
activation of the E2A promoter. Consistent with our results,
Srinivasanetal. foundthat atruncatedlarge Tcontainingthe
amino-terminal 121 residues, which retained the binding site
for RB and
p107,
could transactivate the E2A promoter (30).Nevertheless, this mutant did not trans activate the E2A
promoter aseffectivelyasdidwild-type largeT.However,since
thismutant also lost its nuclear translocation signal, it is not
clear from their experiments whether the decrease in trans
activation indicates that structures carboxy terminal to the
RB/plO7-binding site are involved in trans activation of the
E2A promoter or whether translocation of large T to the
nucleusis requiredforoptimaltrans activation.
Significance of multiple activation processes encoded by
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.62.297.73.170.2]large T and small t. It is not clear why there are multiple
pathways by which
SV40
early gene productsactivate the AdE2A
promoter.
Several viruses, including pseudorabies virus(13),
SV40
(19), polyomavirus (16), and papillomavirus (26)encode proteins that trans activate the E2A promoter. Thus,
even though there are no ATF- or EIIF-binding sites within
the
SV40
genome, productive SV40 infection may requireactivationof a
cellular
process, perhaps transcription ofcellu-lar genes containingATFandEIIFsites. These events may be
central to the subversion of host cell processes that lead to
cellular DNA synthesis during productive viral infection (see
reference 12 for a review). It is possiblethat several
indepen-dent pathways leading to activation of ATF- or
EIIF-depen-dent transcription must occur. By encoding large T and small
tproteinsthat each can interact with distinct pathways leading
to activation of EIIF- and ATF-dependent
transcription,
thevirus ensures that all essential pathways are active. Alterna-tively, there may be different pools of large T and small t that form complexes with various cellular proteins, each of which performs a specific function during regulation of viral
tran-scription,
DNA synthesis, and activation ofcellular processesneeded for viral assembly. Not allof these complexes may be
present simultaneously, but they each may form at different
times within the cell or lytic cycle. Further study of the
mechanisms of transcription regulation by small t and by large
T with structures outside the RB/p107pocket will be necessary
and will advance our understanding of the diverse biochemical
processes by which the
EIIF
and ATF transcription factors canbe regulated.
I am grateful to Sunanda Babu and Jeannine Richard for expert
technical assistance; to James DeCaprio, William Kaelin, Robin
Bachelder,
and Maria Bennet for helpful comments; and to Alan Smith for critical reading of the manuscript. Lauren Sompayrac kindlyprovided the pSRa.B and pSRetB-t/cDNA plasmids, and Ilan Bikel
provided the pRSV-T/u andpRSV-T/Kl plasmids.
This work was supported in part by grant MV-477 from the American Cancer Society and grant CA50599 from the National Cancer Institute.
REFERENCES
1. Abdel-Hafiz, H. A.-M., L. E. Heasley, J. M. Kyriakis, J. Avruch, D. J. Kroll,G. L. Johnson, and J. P. Hoeffler. 1992. Activating transcription factor-2 DNA-binding activity is stimulated by phos-phorylation catalyzed by p42 and p54 microtubule-associated protein kinases. Mol. Endocrinol. 6:20)79-2089.
2. Bagehi, S., R. Weinmann, and P. Raychaudhuri. 1991. The retinoblastoma protein copurifies with E2F-I, an EIA-regulated inhibitor of the transcription factor E2F. Cell 65:1063-1t)72.
3. Bandara, L. R., and N. B. La Thangue. 1991. Adenovirus E1A prevents the retinoblastomagene product from complexing with a cellular transcription factor. Nature (London) 351:494-497. 4. Bikel, I., and M. R. Loeken. Unpublished results.
5. Bikel, I., and M. R. Loeken. 1992. Involvement of simian virus40
(SV40) small t antigen in trans activation of SV40 early and late
promoters. J. Virol. 66:1489-1494.
6. Brady, J., J. B. Bolen, M. Radonovich, N. Salzman, and G. Khoury. 1984. Stimulation of simian virus 40 late gene expression by simian virus 40 tumor antigen. Proc. Natl. Acad. Sci. USA 81:2040-2044.
7. Chellappan, S. P., S. Hiebert, M. Mudryj, J. M. Horowitz, and J. R. Nevins. 1991. TheE2F transcription factor isacellulartarget for the RBprotein. Cell65:1053-10)61.
8. Chittenden, T., D. M. Livingston, and W.G.Kaelin, Jr. 1991. The
T/EIA-binding domain of theretinoblastomaproductcaninteract selectively with a sequence-specific DNA-binding protein. Cell 65:1073-1082.
9. DeCaprio, J. A., J. W. Ludlow, J. Figge, J.-Y. Shew, C. M.Huang, W.-H. Lee, E.Marsilio, E.Paucha,and D. M. Livingston. 1988. SV40 large tumor antigen forms a specific complex with the product oftheretinoblastoma susceptibilitygene.Cell 54:275-283. 10. Devoto, S. H., M. Mudryj,J. Pines, T.Hunter,andJ.R.Nevins. 1992. A cyclin A-protein kinase complex possesses sequence-specific DNA binding activity: p33cdk2 is a component of the E2F-cyclin A complex.Cell68:167-176.
11. Ewen, M. E.,J. W. Ludlow, E. Marsilio, J. A. DeCaprio, K. C. Millikan,S. H.Cheng,E.Paucha,and D. M.Livingston. 1989.An N-terminal transformation-governing sequence of SV40 large T antigen contributesto thebindingofbothpllORb andasecond cellular protein, p12(). Cell58:257-267.
12. Fanning,E. 1992. Simian virus 40largeTantigen: thepuzzle,the pieces, and theemerging picture.J. Virol. 66:1289-1293. 13. Imperiale,M. J.,L.T.Feldman,and J. R.Nevins.1983. Activation
of gene expressionbyadenovirus andherpesvirusregulatorygenes acting in trans and by acis-acting adenovirus enhancer element. Cell 35:127-136.
14. Jog, P., B. Joshik,V.Dhamankar,M. J.Imperiale, J.Rutila,and K. Rundell. 1990. Mutational analysis of simian virus 40 small-t antigen. J. Virol.64:2895-2900.
15. Kalderon, D., and A. E. Smith. 1984. in vitro mutagenesis ofa
putativeDNAbindingdomain of SV40large-T.Virology 139:109-137.
15a.Livingston, D.Personal communication.
16. Loeken, M. R. 1992. Simianvirus 40 smalltantigentransactivates the adenovirusE2A promoterbyusing mechanisms distinct from those used byadenovirus EIA. J. Virol. 66:2551-2555.
17. Loeken, M. R., I. Bikel, D. M. Livingston, and J. Brady. 1988. trans-activation ofRNApolymerase II andIII promotersby SV40
small t antigen. Cell 55:1171-1177.
18. Loeken, M. R., and J. Brady. 1989. The adenovirus EIIA
en-hancer: analysis of regulatory sequences and changes in binding activity ofATFand EIIFfollowing adenovirus infection. J. Biol. Chem. 264:6572-6579.
19. Loeken, M. R., G.Khoury, and J.Brady. 1986. Stimulation of the adenovirus E2 promoter by simian virus 40 T antigen or EIA occursbydifferent mechanisms. Mol. Cell. Biol. 6:2020-2026. 20. Marsilio, E., S. H.Cheng,B.Schaffbausen,E. Paucha,and D. M.
Livingston. 1991. TheT/t commonregion of simian virus40large T antigen containsa distinct transformation-governing sequence. J. Virol. 65:5647-5652.
21. Montano, X., and D. P. Lane. 1984.Monoclonal antibodytosimian virus 40small t. J.Virol. 51:760-767.
22. Montano, X., R.Millikan, J. M.Milhaven, D. A.Newsome, J.W. Ludlow,A. K.Arthur,E.Fanning, I.Bikel,and D. M.Livingston. 1990. Simianvirus40smalltumorantigen andanamino-terminal domain of large tumor antigen share a common transforming function. Proc. Natl. Acad. Sci. USA 87:7448-7452.
23. Mudryj, M., S. H. Devoto, S. W.Hiebert,T.Hunter,J.Pines,and J. R. Nevins. 1991. Cell cycle regulation of the E2Ftranscription factor involvesan interaction with cyclinA. Cell 65:1243-1253. 24. Pallas, D. C., L. K. Shahrik, B. L.Martin,S.Jaspers,T. B.Miller,
D. L. Brautigan, and T. M. Roberts. 1990. Polyoma small and middleTantigens andSV40 smalltantigenform stablecomplexes with proteinphosphatase 2A.Cell 60:167-176.
25. Pallas,D. C.,W.Weller,S.Jaspers, T. B.Miller,W. S.Lane,and T. M.Roberts. 1992. Thethird subunit ofprotein phosphatase2A (PP2A),a55-kilodaltonproteinwhich isapparentlysubstituted for by T antigens in complexes with the 36- and 63-kilodalton PP2A
subunits,bears little resemblance to Tantigens. J. Virol.
66:886-893.
26. Phelps,W. C., C. L. Yee, K.Munger,and P. M.Howley. 1988.The human papillomavirus type 16 E7 gene encodes transactivation and transformation functions similartothose ofadenovirus EIA. Cell 53:539-547.
27. Sanger,F., S.Nicklen,and A. R.Coulson. 1977. DNA sequencing
with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
28. Scheidtmann, K. H., M. C. Mumby, K. Rundell, and G. Walter. 1991. Dephosphorylation of simian virus 40 large-T antigen and p53 protein by protein phosphatase 2A: inhibition by small-t
on November 9, 2019 by guest
http://jvi.asm.org/
antigen. Mol.Cell. Biol. 11:1996-2003.
29. Shirodkar, S., M. Ewen, J. A. DeCaprio, J. Morgan, D. M. Livingston,andT. Chittenden.1992.The transcriptionfactorE2F interacts with the retinoblastoma product and a p107-cyclin A
complexinacellcycle-regulatedmanner.Cell 68:157-166. 30. Srinivasan, A., K. W.C.Peden,andJ. M. Pipas. 1989.The large
tumor antigen of simian virus 40 encodes at least two distinct transformingfunctions. J. Virol.63:5459-5463.
31. Takebe, Y., M.Seiki,J.-I.Fujisawa, P.Hoy, K. Yokota, K.-I. Aria, M.Yoshida,and N.Arai. 1988. SRcx promoter: an efficient and
versatile mammalian cDNAexpression system composed of the simian virus40early promoterand the R-U5 segmentof human T-cellleukemia virustype1long terminalrepeat.Mol.Cell. Biol.
8:466-472.
32. Tooze, J. (ed.). 1981. DNAtumorviruses: molecular biologyof
tumor viruses. Cold Spring Harbor Laboratory, Cold Spring
Harbor, N.Y.
33. Walter, G., R. Ruediger, C. Slaughter, and M. Mumby. 1990.
Association of protein phosphatase 2Awith polyoma virus
me-dium tumorantigen. Proc. Natl. Acad. Sci. USA 87:2521-2525. 34. Wang, H.-G. H., G. Draetta, and E.Moran. 1991. EIA induces
phosphorylation of the retinoblastoma protein independently of direct physical association between the ElA and retinoblastoma products. Mol. Cell. Biol. 11:4253-4265.
35. Yang, S.-I.,R.L. Lickteig, R.Estes, K. Rundell,G. Walter, and M.C.Mumby. 1991. Control of protein phosphatase 2A by simian virus 40 small-t antigen. Mol. Cell. Biol. 11:1988-1995.
36. Zhu, J., P. W. Rice, M. Chamberlain, and C. N. Cole. 1991. Mapping the transcriptional transactivation function of simian virus 40 large T antigen. J. Virol. 65:2778-2790.