0022-538X/97/$04.0010
Copyright © 1997, American Society for Microbiology
Cell and Viral Regulatory Elements Enhance the Expression
and Function of a Human Immunodeficiency Virus
Inhibitory Gene
UDAYKUMAR RANGA,1CLIVE WOFFENDIN,1,2ZHI-YONG YANG,1,2LING XU,1SUNITA VERMA,1
DAN R. LITTMAN,2,3ANDGARY J. NABEL1,2*
Departments of Internal Medicine and Biological Chemistry1and Howard Hughes Medical Institute,2University of
Michigan Medical Center, Ann Arbor, Michigan 48109-0650, and Skirball Institute of Biomolecular Medicine, NYU Medical Center, New York, New York 100163
Received 21 January 1997/Accepted 4 June 1997
Regulated expression of recombinant genes in CD41cells is an important objective for gene therapy of AIDS,
as these cells represent the principal target for viral replication of human immunodeficiency virus (HIV). We report here that specific combinations of CD4 cell-specific and viral regulatory elements can enhance expres-sion of an antiviral gene product. Different viral regulatory elements were incorporated into a previously reported CD4 locus control region to increase the expression of reporter genes in T and monocytic cell lines. The CD4-specific regulatory elements were included to enhance expression in CD4 cells, and viral regulatory
regions, including the cytomegalovirus immediate-early (CMV IE) upstream enhancer, which contains thekB
and Ap1 regulatory elements and a Tat-responsive element of the HIV type 1 long terminal repeat, were used to increase gene expression and modulate its activity in response to viral infection. In transient transfection assays, this vector was 100- to 1,000-fold more active than the original CD4 regulatory elements alone. Expression of an inhibitory form of the Rev protein, Rev M10, was more effective than previously described
vectors and protected against productive viral replication in CD41peripheral blood mononuclear cells. The
combination of CD4 lineage-specific and viral regulatory elements will facilitate the development of more effective antiviral genetic strategies for AIDS.
The problems encountered in devising effective treatments for AIDS have emphasized the need to develop alternative strategies to treat this condition. One such approach is the transfer of genes which inhibit productive replication of human immunodeficiency virus (HIV) in its target cells. A genetic intervention strategy against HIV requires the identification of one or more gene products that are able to inhibit or restrict the replication of HIV. It is also important to regulate antiviral gene expression and minimize any adverse effects on normal cellular gene expression. A number of antiviral genes have been successfully identified in cell culture models. These in-clude Tat activation response (TAR) element and Rev re-sponse element (RRE) decoys (1, 5, 31, 34, 63), ribozymes (47, 55, 71, 73), toxic gene products (10, 13, 24), intracellular anti-bodies (15, 41, 43), antisense genes (67), and transdominant negative mutants of HIV structural (65) and regulatory (2, 4, 33, 38, 44, 48, 67) proteins. The Rev regulatory protein of HIV offers some attractive features that make it a viable candidate for genetic strategies. The expression of functional Rev is es-sential for the translocation of unspliced or singly spliced HIV mRNA from the nucleus to cytoplasm (18, 39, 60). More im-portantly, the presence of Rev above a threshold level is critical for its biological function (49). Rev M10, a transdominant negative mutant of Rev (36, 40), has been shown to effectively block the function of wild-type Rev in T cells and peripheral blood mononuclear cells (PBMC) (2, 4, 35, 38, 68).
Another factor which is likely to affect the success of gene
transfer strategies is the ability to express sufficient amounts of inhibitory gene products in the relevant cell type. Although the PBMC represents a relevant target cell of HIV infection, trans-fer of genes into PBMC by using viral vectors has become increasingly efficient but requires stimulation of cells for opti-mal gene transfer. This process also activates latent provirus in these cells (23, 72), but techniques have recently been used successfully to deliver antiviral genes without activating endog-enous HIV replication in vitro (19, 46). At least one nonviral vector, using particle-mediated gene transfer, has also been shown to provide effective gene transfer, improve safety, and confer antiviral effects (70). Monocytic cells are also readily infected by the virus, and for this reason, efforts have been made to express such genes in hematopoietic stem cells and early progenitors. Protocols for identification and isolation of the stem cells and the introduction of recombinant genes have been developed (29, 62). Independent of the method of gene transfer or the intended target cell population, cell-specific and regulated expression of the introduced genes is desirable for gene transfer strategies in AIDS. Most previous studies have made use of retroviral regulatory elements, such as those found in Moloney murine leukemia virus or Rous sarcoma virus (RSV), for this purpose. These regulatory elements do not confer cell-specific expression and are weakly active in PBMC. In addition, the expression pattern of these promoters varies among different cell types (52).
CD4, the principal cellular receptor for HIV (14), plays an important role in HIV infection and pathogenesis (64), is down-regulated on the cell surface following infection with this virus (25), and serves as a marker for the cell lineages which diminish progressively over the course of HIV infection (7). Several subsets of macrophages or dendritic cells which are believed to function as the reservoirs of HIV are also positive * Corresponding author. Mailing address: Departments of Internal
Medicine and Biological Chemistry, Howard Hughes Medical Insti-tute, University of Michigan Medical Center, 1150 W. Medical Center Dr., 4520 MSRB I, Ann Arbor, MI 48109-0650. Phone: (313) 647-4798. Fax: (313) 647-4730. E-mail: [email protected].
7020
on November 9, 2019 by guest
http://jvi.asm.org/
for CD4 expression (17). For these reasons, CD41T cells and macrophages or dendritic cells represent logical targets for anti-HIV gene therapy. A 339-bp regulatory element which controls CD4-restricted gene expression of reporter genes has been identified approximately 13 kb upstream of the murine CD4 promoter (56). When this regulatory element was placed upstream of the CD4 promoter, expression was restricted to CD41cells (56). The combination of the CD4 enhancer and promoter, however, is relatively weak in T cells, particularly in transiently transfected PBMC. In this study, we report modi-fications made to this vector that result in enhanced transcrip-tional activation in PBMC and responsiveness to viral transac-tivators, such as Tat, as well as cellular activators of HIV gene expression, including NF-kB and Ap1. Expression and efficacy of Rev M10 in these vectors were evaluated in an attempt to improve its antiviral effect and maintain expression in CD4 cells.
MATERIALS AND METHODS
Cell culture.All cell lines except HeLa were cultured in RPMI 1640 medium supplemented with 10% fetal calf serum penicillin (100 U/ml), streptomycin (100 mg/ml), and glutamine (Gibco BRL, Gaithersburg, Md.). 2-Mercaptoethanol (50 mM) was added to the THP-1 cultures. HeLa cells were grown in Dulbecco modified Eagle medium containing 10% fetal calf serum, glutamine, and anti-biotics. PBMC were cultured in X-Vivo 15 medium (BioWhittaker, Walkersville, Md.) supplemented with interleukin-2 (IL-2; 300 IU/ml; Cetus). Cells were incubated at 37°C in 5% CO2.
Vectors (i) pK7-CAT.The vector pK7-CAT was derived from the parental vector pBL(CD4)-CAT (56) by making two modifications. First, a uniqueBglII site was created upstream of theXbaI site at the 59end of the CD4 promoter. CD4 promoter fragments were amplified by using a common sense primer, and variable antisense primers containingBglII andXbaI restriction enzyme sites, respectively, replaced the original promoter. This created mutant promoters with progressive deletions at the transcription initiation site. The full version of the amplified promoter lacked the untranslated 70-bp fragment at the 39end of the original promoter. Second, a section of the HIV type 1 (HIV-1) genome (280 to 172) consisting of the three Sp1 sites, the TATA box, and TAR was amplified by PCR and cloned downstream of the CD4 promoter. Additional modifications were made to this basic vector, pK7-CAT. The cytomegalovirus (CMV) enhancer element (2583 to2224) (6), amplified with primers flanked withSphI sites, was cloned upstream of the CD4 regulatory element to derive pK7c-CAT. The CMV enhancer was also cloned into the parental vector to make the vector pBLc (CD4)-CAT.
(ii) pK7c Rev M10.The expression vector pK7c-Rev M10 was constructed by deleting the RSV promoter and the TAR sequence of the pRSV-TAR-Rev M10 vector byAatII andHindIII digestion. The pK7c cassette (CMV enhancer, CD4 regulatory element, CD4 promoter, and TAR) from pK7c-CAT was removed by HindIII andXhoI digestion and was blunt-end ligated to the backbone described above. The pK7c cassette was thus placed upstream of the Rev M10 gene. The bovine growth hormone poly(A) sequence present in these vectors was replaced by the poly(A) sequence derived from theb-globin gene (aBglII-PflMI fragment of 213 bp) (66). The vectors pRSV-Rev, pRSV-CAT-RRE, pRSV-Rev M10 (35), HIV-CAT (50), and pHD101 Tat (42) have been described previously.
(iii) pK7cI-Rev M10.The final version of the HIV-1 long terminal repeat (LTR) and the CD4 promoter chimeric vector, pK7cI-Rev M10, was constructed as follows. TheHindIII-SalI fragment of the CMV immediate-early (IE) intron was removed from the vector pUCint39and blunt-end ligated into theHindIII site (between TAR and Rev M10) of the vectors pK7c-Rev M10. This version of the CMV IE intron contains an internal deletion between theXcmI andHpaI sites. The control vector pK7cI-DRev M10 was constructed by using the same scheme. This control vector is identical to the expression vector except that there is a 2-bp deletion in the transcription initiation site. An additional control vector, pK7cI-DTARDRev M10, was constructed by deleting a 30-bp fragment of the TAR region (SacI toAflII) to disrupt the stem-loop structure.
Gene transfer.Suspension cultures were transfected by using a modification of the DEAE-dextran method (22). Briefly, 53106cells were transfected with 5 to 6mg of total DNA. The DNA concentrations were made up with pBluescript wherever necessary. Cells were washed twice with TS buffer (0.137 mM NaCl, 7 mM KCl, 0.3 mM Na2HPO4, 0.5 mM MgCl2, 0.7 mM CaCl2, 25 mM Tris base [pH 7.4]) and resuspended in 475ml of TS buffer containing DNA. DEAE-dextran (molecular weight, 500,000; Pharmacia) was added to the cell suspension at a final concentration of 0.5 mg/ml and then incubated for 15 min at room temperature. Five milliliters of RPMI complete medium supplemented with 1 mM chloroquine (Sigma, St. Louis, Mo.) was added to each tube, and the cells were incubated for 30 min at 37°C. The cells were washed once with complete medium and seeded in six-well tissue culture plates. Cells in parallel sets were stimulated with phorbol 12-myristate 13-acetate (PMA; 10 ng/ml) 24 h after
transfection and harvested after an additional 24 h of incubation. HeLa cells were transfected by the CaPO4method (12). PBMC were transfected by particle-mediated gene transfer as described previously (70).
Gene expression and viral challenge. Chloramphenicol acetyltransferase (CAT) assays were performed as described previously (59), with some modifi-cations. Cells were harvested, washed once with phosphate-buffered saline and resuspended in 0.25 M Tris (pH 7.8). They were lysed by three or four cycles of freezing and thawing, and clear lysates were collected after clearing the debris by microcentrifugation. Protein content of the cell extracts was determined by the dye-binding assay (Bio-Rad, Hercules, Calif.). The reaction mixture of 150ml consisted of measured amounts of protein, 20ml of acetyl coenzyme A (3.5 mg/ml in 0.25 M Tris), 2ml of [14
C]chloramphenicol, and 0.25 M Tris (pH 7.8). The amount of protein added and the length of incubation at 37°C varied depending on the transcriptional strength of the vectors used. Experiments were repeated several times. The reaction products were extracted with 0.5 ml of ethyl acetate. The organic phase was lyophilized, and the pellets were redissolved in 30 ml of ethyl acetate. The samples were spotted on a thin-layer silica gel plate (catalog no. 4410221; Whatman) and run in a glass chamber with 95:5 chloro-form-methanol mixture. The chromatograms were analyzed on ab-scope. Virus challenge experiments were performed as described previously (70).
RESULTS
Development of a CD4 promoter which is responsive to Tat.
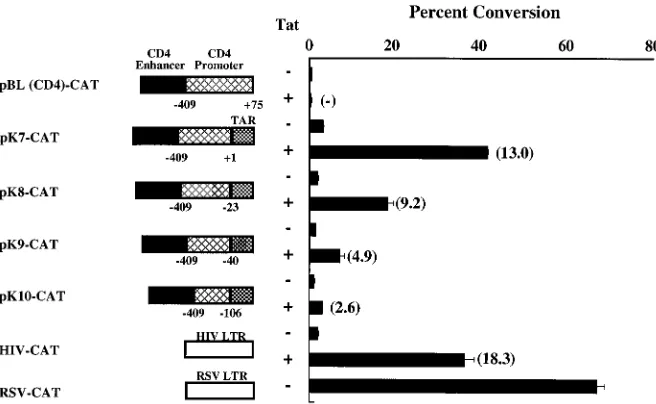
CD4 lineage-specific expression of a CAT reporter with regu-latory elements from the murine CD4 gene was reported pre-viously (56). A 339-bp regulatory fragment from a region;13 kb upstream of the murine CD4 gene was shown to confer CD4-specific expression of the CAT gene when used in com-bination with the CD4 promoter; however, this comcom-bination of CD4 regulatory elements was 10- to 20-fold less efficient than HIV or RSV promoters used previously to express recombi-nant genes in T cells (Fig. 1). To increase gene expression and maintain the CD4-specific regulatory elements, different com-binations of heterologous regulatory regions were analyzed. We first replaced 70 bp of untranslated sequence (at the 39end of the CD4 promoter) with a segment of the HIV-1 LTR consisting of the three Sp1 sites, the TATA box, and the TAR sequence. The presence of the Sp1 sites upstream of TAR is essential for responsiveness to Tat transactivation (3, 58, 61), as is the presence of TAR at the 59end of the mRNA (26, 45, 58). Deletion of this 70-bp 59 untranslated segment was thus required to make this hybrid promoter responsive to Tat (data not shown). Additional deletions were made at the 39end of the CD4 promoter to map sequences that might be dispensable for transcriptional function with the addition of the HIV pro-moter. Progressive deletions at the 39 end of the CD4 pro-moter were analyzed when linked to the HIV propro-moter placed immediately downstream of each of these deletions, and the transcriptional activity of each of these chimeric vectors was estimated in transient transfection assays of Jurkat cells in the presence or absence of Tat. The CD4-Sp1/TAR hybrid vectors were functional in the CAT assay at levels significantly higher than those generated from the parental vector pBL(CD4)-CAT. CAT expression was enhanced 2- to 15-fold in the pres-ence of Tat, indicating that the TAR sequpres-ence in these plas-mids was functional. The resultant hybrid vector, pK7-CAT, with the full-length CD4 promoter (2409 to11), the Sp1 sites, and TAR displayed higher CAT activity than other reporter plasmids. Hybrid vectors containing progressive deletions into the 39end of the promoter demonstrated proportionately de-creased CAT activity (Fig. 1). These results indicated that the full-length CD4 promoter without 59 untranslated RNA se-quences provided optimal gene expression from the chimeric vector in transient transfections. We also prepared chimeric vectors which lacked the Sp1 sites in the HIV-1 LTR, consist-ing of only the TATA box and the TAR sequence (247 to 72), but these vectors were not Tat responsive in transient trans-fection assays. In contrast, pK7-CAT was Tat responsive (Fig. 2A), confirming the functional importance of the Sp1 and TAR
VOL. 71, 1997 REGULATION OF ANTIVIRAL GENE EXPRESSION 7021
on November 9, 2019 by guest
http://jvi.asm.org/
sites (3, 61) in this promoter context. Based on these results, pK7-CAT was selected for further modification.
Addition of the CMV upstream enhancer to the CD4-TAR
promoter.A 360-bp CMV enhancer element was placed
up-stream of the CD4-specific enhancer element in pK7-CAT. This element contains at least sixkB regulatory elements and an additional Ap1 site. These sites are recognized by NF-kB and Fos-Jun, respectively, both of which are stimulated
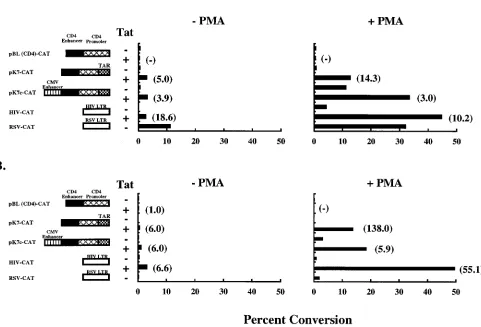
follow-ing T-cell activation (21, 28). The resultfollow-ing vector, pK7c-CAT, was tested in parallel with pK7-CAT for transcriptional activity in a transient transfection assay (Fig. 2B). In Jurkat cells, the levels of transcription in the absence of Tat transactivation were 8.6-fold higher when the CMV enhancer was present in the vector than in its absence. Although minimal increase in transcription was seen in unstimulated cells cotransfected with Tat, in most cell lines and in PBMC, the presence of the CMV enhancer in the vector resulted in increased CAT activity both in the presence and in the absence of Tat (see below). Addition of the CMV enhancer to the parental vector pBL(CD4)-CAT, on the other hand, had no effect on transcriptional activity, with CAT activity barely detectable (data not shown).
Expression of the CD4-TAR hybrid promoter in different
cell lines.Expression of the parental vector pBL(CD4)-CAT
[image:3.612.140.468.69.271.2]has been previously shown to be T-cell specific, due to the presence of its CD4-specific enhancer and promoter elements. To determine whether the pattern of gene expression of the altered promoter would differ from that of the parent, its expression patterns in T- and non-T-cell lines were analyzed. Transient transfection assays were performed, and cells were stimulated with PMA 24 h later. Cells were incubated for an additional 24 h, and CAT activity was determined. Then the vectors pBL(CD4)-CAT, pK7-CAT, pK7c-CAT, HIV-CAT, and RSV-CAT were compared in two T-cell leukemia lines in the absence or presence of PMA. In Jurkat cells, both pK7-CAT and pK7c-pK7-CAT showed enhanced pK7-CAT activity in the absence of PMA stimulation, and the level was four- to fivefold higher in the presence of Tat (Fig. 3A). When the cells were stimulated with PMA, these vectors displayed;3- to 14-fold-increased CAT activity (Fig. 3A). pK7c-CAT, for example, showed 10.3- and 40-fold-higher CAT activity in the presence of PMA alone and PMA and Tat together, respectively. In H9 cells, pK7c-CAT activity was 15.3- and 108.2-fold higher with PMA stimulation alone and PMA-Tat stimulation, respec-tively. The parental vector pBL(CD4)-CAT showed less than 0.1 to 0.3% of this CAT activity under the same conditions.
FIG. 1. Functional analysis of the CD4 promoter by progressive 39-end deletions. Deletions at the transcription initiation site in the CD4 promoter were performed by using PCR. Deletion mutants of the promoter were placed between the CD4 regulatory element upstream and the HIV-1 LTR consisting of the Sp1 sites, TATA box, and TAR sequence downstream, as shown. The length of the CD4 promoter in each construct, corresponding to the transcription initiation site, is shown. Transcriptional strength of these constructs was assessed in a transient transfection assay using Jurkat cells. CAT activity in the protein extracts was determined 48 h posttransfection. The values are shown as percent conversion of chloramphenicol to its acetylated derivatives. Each data point represents the mean6standard deviation of three replicate assays, verified in three independent transfections. Values in parentheses represent fold transactivation due to Tat.
FIG. 2. Effects of the Sp1 sites (A) and CMV enhancer (B) on the transcrip-tional function of the CD4-TAR hybrid vector. The HIV sequence in the pK7-CAT vector with or without the Sp1 sites ranges from280 to175 or247 to172, respectively. The CMV enhancer was placed upstream of the CD4 enhancer in the pK7 vector to generate the pK7c vector. Jurkat cells were transfected with either of these vectors with or without pHD101 Tat. The values are shown as percent conversion of chloramphenicol to its acetylated derivatives. Each data point represents the mean6standard deviation of three replicate assays. Values in parentheses represent fold transactivation due to Tat.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.60.289.461.650.2]Taken together, pK7c-CAT was significantly more active than the parental vector, with 52.3- and 141.5-fold-higher levels of CAT activity upon PMA stimulation and Tat transactivation in Jurkat and H9 cells, respectively.
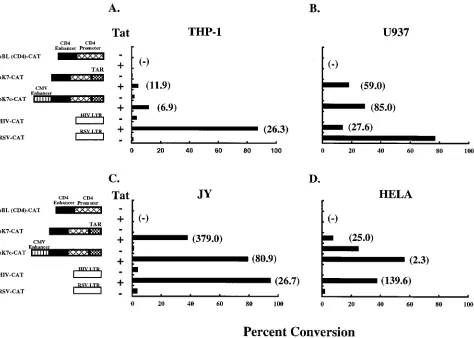
The transcriptional activity of pK7c-CAT was also deter-mined in non-T cells lines, including THP-1 (monocytic leuke-mia), U937 (premonocytic lymphoma), JY (B cells), and HeLa (cervical epithelioid carcinoma). The pattern of CAT activity in the presence of PMA (Fig. 4) was essentially the same as in unstimulated cells but was severalfold greater in magnitude (data not shown). The chimeric CD4 vectors were functional in each cell type. pK7c-CAT activity was again greater than pK7-CAT activity in all cells under all experimental conditions. As noted in T cells, Tat transactivation in all of these cells resulted in increased CAT activity in pK7c-CAT, ranging from 6.9-fold (THP-1) to 80.9-fold (JY) to 85.0-fold (U937). CAT expression from the parental vector pBL(CD4)-CAT remained undetect-able. With these additions, pK7c-CAT was found to be com-parable to HIV-CAT in several cell lines and often more active than the RSV LTR (Fig. 3 and 4). In summary, although viral regulatory elements in addition to the CD4 enhancer/promoter resulted in a broader range of cell-specific expression, it pro-vided for enhanced transcriptional activity. Unlike the viral LTRs or the parental CD4 locus control region, the CD4-TAR
hybrid promoter displayed higher transcriptional activity in all of the cell lines tested.
Gene expression in CD41PBMC.We tested the expression
pattern of pK7c-CAT in PBMC, as these cells present a rele-vant target for anti-HIV gene transfer. Importantly, in the absence of Tat, pK7c-CAT displayed transcriptional activity in PBMC from normal donors that was about 3.4-fold higher than that of RSV-CAT. Under the same conditions, pK7c-CAT demonstrated approximately 75.0- and 4.0-fold-higher CAT activity than HIV-CAT in the absence and presence of Tat (Fig. 5A). Inclusion of the CMV enhancer rendered this vector significantly more active in PBMC. pK7c-CAT displayed 18.8-and 27.2-fold-higher CAT activity compared to pK7-CAT in the absence and presence of Tat transactivation, respectively. Under the same conditions, the parental vector pBL(CD4)-CAT displayed no detectable pBL(CD4)-CAT activity.
[image:4.612.67.548.89.416.2]The results of transient transfection assays using PBMC demonstrated that the vector pK7c-CAT was several hundred-fold stronger than the parental vector pBL(CD4)-CAT. In an attempt to increase the transcriptional activity of this vector further, a 386-bp subfragment of CMV IE intron A was cloned into pK7c-CAT such that it was placed between the TAR sequence upstream and the CAT gene downstream. This in-tron has been shown previously to increase expression of the
FIG. 3. CD4-TAR hybrid promoter-driven CAT expression in T-cell lines. Jurkat (A) or H9 (B) cells were transfected with different CAT expression vectors by the DEAE-dextran method. Assays were performed with paired controls. Twenty-four hours posttransfection, PMA was added to one set at a final concentration of 10 ng/ml. Cultures were harvested after an additional incubation for 24 h. CAT activity in the protein extracts was determined. Results are representative of two independent experiments. Standard deviations for values within each experiment were generally#10%. The values are shown as percent conversion of chloramphenicol to its acetylated derivatives. Values in parentheses represent fold transactivation due to Tat.
VOL. 71, 1997 REGULATION OF ANTIVIRAL GENE EXPRESSION 7023
on November 9, 2019 by guest
http://jvi.asm.org/
CMV enhancer (11). Transcriptional activity of the new vector, pK7cI-CAT, was analyzed by transient transfection assay in PBMC and compared to activities of the other vectors (Fig. 5B). Addition of the intron A subfragment to pK7c-CAT in-creased transcriptional activity. Twenty-four hours after the transfection, pK7cI-CAT demonstrated 20.4- and 12.1-fold-stronger activity in the absence and presence of Tat transacti-vation, respectively. This vector also showed stronger activity than RSV-TAR-CAT, a prototype of the vector used in pre-vious clinical gene transfer studies (69), especially in the ab-sence of Tat transactivation, in which case it was 36.4-fold higher. Importantly, the CAT activity of pK7cI-CAT was ex-pressed at higher levels than those of the other vectors for longer time periods (Fig. 5B, right panel). Seventy-two hours after the transfection, pK7cI-CAT showed 10.9- and 9.5-fold-higher CAT activity compared to RSV-TAR-CAT in the ab-sence and preab-sence of Tat transactivation, respectively. CAT activity of the parental vector pBL(CD4)-CAT remained be-low detection in these experiments (data not shown).
Transcription start site in the optimized vector.In in vitro
transient transfection assays, the chimeric vector stimulates CAT reporter gene expression effectively. Because transcrip-tional initiation from this chimeric promoter could theoreti-cally be derived from two major transcription initiation sites, one from each promoter, we performed primer extension
anal-ysis on total RNA extracted form PBMC transduced with pK7c-Rev M10 to determine which sites were functional. DNA fragments of 68 or 148 bp were expected depending on whether the transcription initiation site was located in TAR or in the CD4 promoter, respectively. A single band at the 68-bp level was detected in a dose-dependent fashion (Fig. 6, lanes 3 and 4). No signal was seen with RNA isolated from untrans-duced PBMC (Fig. 6, lanes 1 and 2). No other extension product of higher molecular weight was found, indicating that the transcription start site from HIV but not the CD4 pro-moter is functional in this propro-moter. Identical results were obtained also in 293 cells transfected with CAT, pK7c-Rev M10, or pK7cI-pK7c-Rev M10 (data not shown).
Rev M10 function in chimeric CD4 expression vectors.
[image:5.612.69.543.71.409.2]These data indicated that the combination of the CD4 and viral regulatory elements in the pK7c-CAT vector significantly increased CAT reporter gene expression. To determine whether this vector could provide improved Rev M10 inhibi-tion of HIV replicainhibi-tion, the CAT gene in pK7c-CAT was replaced with the Rev M10 open reading frame to generate the pK7c-Rev M10 vector (Fig. 7A). The 386-bp CMV intron was subsequently cloned upstream of the Rev M10 gene to make pK7cI-Rev M10. A control vector with a deletion in the trans-lation initiation codon (pK7cI-DRev M10) was analyzed, al-though another control vector with an additional deletion in
FIG. 4. CD4-TAR hybrid promoter-driven CAT expression in non-T cells. THP-1 (A), U937 (B), or JY (C) cells were transfected with the CAT expression vectors with or without pHD101, using the DEAE-dextran method. HeLa cells (D) were transfected by the calcium phosphate method. Cultures were stimulated with PMA 24 h after transfection and harvested after incubation for an additional 24 h. CAT activity in the protein extracts was determined. The values are shown as percent conversion of chloramphenicol to its acetylated derivatives. Values in parentheses represent fold transactivation due to Tat. Results are representative of two independent experiments. Standard deviations for values within each experiment were generally#10%.
on November 9, 2019 by guest
http://jvi.asm.org/
the TAR stem-loop (pK7cI-DTAR-DRev M10) was tested to control for potential inhibition by the TAR RNA structure which can act as a decoy (63), with similar effects (data not shown). A cotransfection inhibition assay was performed as reported previously (35) to determine whether expression of Rev M10 from these promoters could inhibit Rev-dependent CAT activity. A vector expressing Rev M10 under the regula-tion of the RSV LTR and HIV TAR (pRSV-TAR-Rev M10) and its identical control vector (pRSV-TAR-DRev M10) were included for comparison. This vector had been used in previ-ous clinical studies (69). Coexpression of the reporter vector, pRSV-CAT-RRE, with increasing amounts of the transactivat-ing plasmid, pRSV-Rev, resulted in a linear increase in CAT activity (data not shown). To perform the inhibition assay, Jurkat cells were cotransfected with a constant concentration of pRSV-CAT-RRE and pRSV-Rev and increasing amounts of different Rev M10 or control expression plasmids both in the absence and in the presence of a Tat expression vector. Controls were included where the CAT expression vector was transfected in the absence of the Rev expression vector. The conversion levels obtained from such controls were substracted to account for background. These assays revealed that the optimized vector pK7cI-Rev M10 effectively inhibited Rev-dependent CAT expression in a dose-Rev-dependent manner both in the absence and in the presence of Tat expression (Fig. 7A and B, respectively) relative to the controls. In both cases,
pK7cI-Rev M10 showed stronger inhibition than pRSV-TAR-Rev M10. The experimental design is complex, as it requires four vectors to enter the same cell to show CAT reporter inhibition. Complete specific CAT inhibition is unlikely under these conditions. To overcome this limitation, we compared the vectors in the linear range of the assay. It is important to compare the vectors in the linear range rather than to look for complete inhibition at higher concentrations where such com-parisons are less sensitive. Such a comparison of pK7cI-Rev M10 and pRSV-TAR-Rev M10 showed the former to be an order of magnitude stronger than the latter. For example, approximately 50 ng of the optimized vector was required for half-maximal reduction in CAT conversion in the presence of Tat (Fig. 7B, right panel). To achieve the same levels of inhi-bition, 400 ng of the pRSV-TAR-Rev M10 was required under the same conditions.
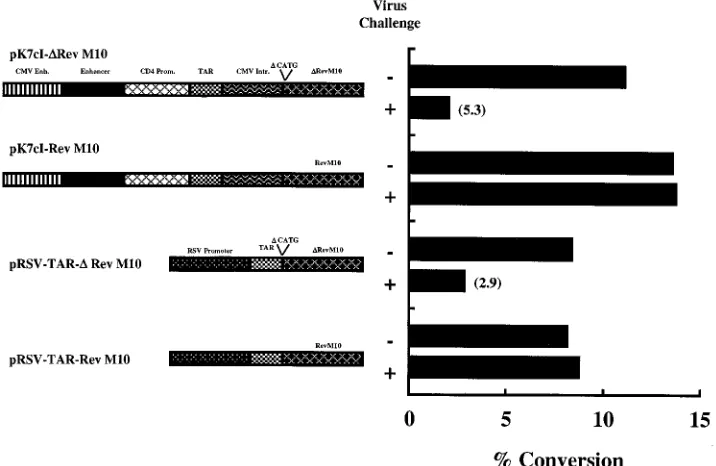
Protection against virus challenge. The potential of these
vectors to inhibit HIV-1 infection in PBMC was examined in virus challenge experiments. Freshly isolated PBMC were co-transduced with one of the Rev M10 orDRev M10 expression vectors and a CMV-CAT vector by particle-mediated gene transfer. CAT activity was used as a reporter to measure over-all cell viability after virus chover-allenge. The cells for each vector were divided into halves; one group was challenged with HIV-1Bru, while the other served as nonchallenged reference
con-trol. CAT activity was quantitated on days 2 and 3. The levels
FIG. 5. CD4-TAR hybrid promoter-driven CAT expression in peripheral blood lymphocytes. PBMC were freshly isolated by using Ficoll-Hypaque and stimulated with immobilized anti-CD3 antibody and IL-2 (50 U/ml) for 2.5 days. PBMC derived from different donors were used in these experiments. Cells (105
/assay) were transduced by particle-mediated gene transfer with 1mg of CAT expression vector and 0.3mg of pHD101 or pBluescript as described elsewhere (70). Following gene transfer, the cells were incubated in the presence of the anti-CD3 antibody for 24 h. (A) CAT activity in 20mg of the protein extract was determined in a 5-h incubation at 37°C; (B) 14mg of protein was used, and the reaction mixture was incubated for 1 to 5 h. The reaction conditions were optimized for the strongest vector used in the experiment such that the conversion would not exceed 60 to 70% level. The results are representative of at least four independent experiments, with standard deviation generally#10% for each determination in the CAT assay. The values are shown as percent conversion of chloramphenicol to its acetylated derivatives. Values in parentheses represent fold transactivation due to Tat.
VOL. 71, 1997 REGULATION OF ANTIVIRAL GENE EXPRESSION 7025
on November 9, 2019 by guest
http://jvi.asm.org/
of CAT activity in cells transfected with Rev M10 expression vectors as opposed to the controls reflected the protection afforded to the cells by Rev M10. In this analysis, the optimized vector pK7cI-Rev M10 provided effective antiviral protection in these experiments (Fig. 8). On day 3, no reduction in CAT activity was seen as a result of the viral challenge with the optimized vector, whereas a 5.3-fold reduction was observed in cells transfected with the control vector. Similar results were also observed with pRSV-TAR-Rev M10 and its control vector used previously in clinical studies (2.9-fold reduction), and identical results were observed on day 2 (data not shown). Protection against HIV-1IIIbinfection was also observed in 293
cells constitutively expressing the CD4 receptor (data not shown).
DISCUSSION
Because of its ability to inhibit productive HIV replication in several systems (2, 4, 35, 38, 68), the transdominant mutant of Rev, Rev M10, has been used as a model to develop intracel-lular immunization for AIDS. Genetic strategies which target Rev (1, 16) appear to be attractive since its biological function is required for HIV replication. When the levels of Rev within a cell exceed a certain threshold level, the virus switches from the early nonproductive to the late productive phase of repli-cation (49). Treatments which maintain Rev below the thresh-old level should therefore reduce the generation of infectious virus from the cell.
Recombinant viruses have traditionally been used for intro-ducing the inhibitory genes into PBMC. Although reasonably efficient, this gene delivery method has several potential dis-advantages. The possibility of infectious virus particles remain-ing in the sample below the detection level cannot be ruled out, raising potential safety concerns (23, 72). Gene expression regulated from viral LTRs is often not optimal in PBMC. Such viral vectors generally also lack tissue-regulated gene expres-sion (52). To overcome these problems, we sought to optimize regulated gene expression. Effective transfer of such vectors
into PBMC can be attained either by retroviral vectors (9, 51) or by particle-mediated transfer, as reported previously (70).
[image:7.612.318.553.71.304.2]The vector pBL(CD4)-CAT was previously shown to direct gene expression specifically in CD4-positive T cells in trans-genic mice (30, 56). The combination of a 339-bp CD4-specific enhancer element and the CD4 promoter in this vector is a relatively weak activator of gene expression but can maintain lineage-specific expression during development (30, 57). We were able to increase the transcriptional activity of the CD4 promoter in this vector by adding several heterologous ele-ments. The activity of each of these modified enhancers was compared with that of the original vector as well as the HIV-and RSV LTRs. Addition of the TAR sequence of the HIV LTR to the CD4 promoter made the latter responsive to Tat. As reported previously (35), this element can increase expres-sion of the inhibitory gene from this promoter in response to Tat synthesized after endogenous HIV gene expression and may provide a TAR decoy effect at lower concentrations of Tat (35, 63). Further addition of a CMV IE enhancer element upstream of the CD4 enhancer element, however, improved the gene expression 100-fold or more in several cell lines (Fig. 3 and 4) and PBMC (Fig. 5). The CMV IE enhancer is a strong transcriptional activator with ubiquitous gene expression prop-erties (6). The CMV IE enhancer consists of binding sites for several cellular transcription factors, including NF-kB and Ap1, both of which are inducible by mitogenic or antigenic activation (20, 32, 54). The presence of such inducible regula-tory elements in the vector could provide an advantage by
FIG. 6. Primer extension analysis of pK7cI-Rev M10 transcriptional initia-tion site. PBMC freshly isolated from a volunteer were stimulated with the immobilized OKT3 antibody for 4 days in AIM-V medium (Gibco BRL). The cells were transduced with pK7cI-Rev M10 vector by particle-mediated gene transfer as described previously (70). The cells were incubated for an additional 24 h in the presence of human IL-2 (50 U/ml) and harvested, and the total RNA was extracted with Trizol reagent (Gibco BRL); 20 (lanes 1 and 3) or 5 (lanes 2 and 4)mg of RNA extracted from untransduced (lanes 1 and 2) or transduced (lanes 3 and 4) cells was used in the assay. An oligonucleotide of 17 bp comple-mentary to the TAR sequence (bp 52 to 68) was end labeled and used in primer extension analysis (53). The extension product was resolved on a 12% acrylamide gel with 8 M urea. The 1-kb DNA ladder (Gibco BRL), end labeled, served as a size standard (lane 5).
FIG. 7. Competitive inhibition of Rev transactivation by Rev M10 expression vectors. Jurkat cells (53106
/assay) were cotransfected with pRSV-CAT-RRE and pRSV-Rev (1mg of each/assay) and different concentrations (1.0, 0.5, 0.1, 0.02, or 0mg/assay) of Rev M10 expression vectors. The experiments were performed in the absence (A) or presence (B) of Tat expression vector pHD101 Tat (0.5mg/assay). Cells were harvested 48 h after incubation, and CAT expres-sion levels were determined in the cell lysates. CAT expresexpres-sion in the absence of the Rev expression vector was used as a negative control. The conversion levels, which are usually 40 to 50 times lower in the absence of Rev, were subtracted as the background. In this experiment, the levels of percent conversion induced by pRSV-CAT-RRE were 0.6 and 34.4 in the absence and presence of Rev, respec-tively. These experiments were repeated at least three times under different conditions, and the data of a representative experiment are shown. Schematic diagrams of the Rev M10 andDRev M10 expression vectors are shown in Fig. 8.
on November 9, 2019 by guest
http://jvi.asm.org/
allowing increased production of the inhibitory protein in mi-togenic or antigen-activated cells. The continuous presence of HIV antigens or immune/inflammatory cytokines in the circu-lation of HIV-seropositive subjects is likely to stimulate the PBMC, thereby activating cellular transcription factors which stimulate CMV enhancer function, increasing expression of Rev M10; however, the presence of the CMV enhancer in this plasmid also resulted in gene expression in other lineages (Fig. 4). Such expression is not problematic for application in cur-rent gene therapy protocols, since CD41cells are preselected prior to gene transfer. It is anticipated that the inclusion of CD4 regulatory elements will be beneficial by maintaining long-term gene expression in vivo, as in transgenic mice (30), while viral regulatory elements increase expression and opti-mize the viral infection.
Addition of introns has been shown to enhance the tran-scriptional activity of the expression vectors and also stabilize the mRNA (8, 27). Previous reports (11) and preliminary work in our laboratory demonstrated that intron A of the CMV IE gene is a strong transcriptional activator. A search to identify binding motifs for cellular factors within this region revealed no such sequences other than NF-1. Deletion of a large inter-nal sequence, spanning fromXcmI toHpaI sites, did not alter the transcription-enhancing activity of this intron, although the deletion removed the NF-1 binding motif. To enhance the transcription activity of the CD4-TAR hybrid promoters, the 386-bp subfragment of CMV IE intron A was added to these expression vectors. This addition resulted in a stronger and prolonged expression of CAT by pK7cI-CAT vector compared to pK7c-CAT, pRSV-TAR-CAT, or HIV-CAT (Fig. 5).
The CD4 promoter in pK7c-CAT exhibited strong transcrip-tional activity in PBMC, which was 4.0- and 3.4-fold greater than those of HIV-CAT and RSV-CAT, respectively (Fig. 5).
[image:8.612.126.484.74.307.2]Tat transactivation increased this activity an additional 5.8-fold. The parental vector under the same conditions showed no detectable activity. The changes in the parental vector thus rendered it at least 2 logs more active in PBMC. The magni-tude of Tat inducibility of the optimized vector, pK7cI, is not as striking as that of the predecessor, pK7c (Fig. 5B). An inverse relationship between the strength of the promoter and levels of Tat transactivation has been reported previously, largely be-cause the denominator has been increased when calculating fold transactivation; however, TAR is still functional (Fig. 5B), and similar maximal levels are attained in the presence of Tat with cellular activation. Our previous studies suggest that higher basal levels of Rev M10 allow more effective inhibition of wild-type Rev, and the addition of TAR not only results in inducible gene expression but also functions as an RNA decoy (35). The data presented in Fig. 7 indicate that, especially at low levels of vector, pK7cI inhibits Rev function better than RSV-TAR in the presence or absence of Tat, confirming that CD4-CMV cassette has the advantage of improved potency for inhibition of Rev function. Most relevant to the antiviral strat-egy, expression of Rev M10 from CD4-TAR hybrid vectors was found to efficiently block wild-type Rev function. In an in vitro competition assay, Rev M10 strongly inhibited Rev-dependent CAT expression (Fig. 7). Expression of Rev M10 in PBMC also inhibited the significant reduction in the levels of CAT expres-sion after viral challenge (Fig. 8). These data indicate that pK7cI can also function in acute viral infection. The potential advantages of the optimized vector, pK7cI, over RSV-TAR are inferred from this assay and the in vitro studies; however, further studies in patients will be needed to address issues of potential clinical efficacy. This study, nevertheless, shows that specific features of HIV replication and cellular gene expres-sion can be exploited to develop potentially more effective
FIG. 8. Protection of PBMC transduced with Rev M10 expression vectors against virus challenge. CAT activity in PBMC transduced with Rev M10 expression vectors or control vectors by particle-mediated gene transfer and challenged with HIV. PBMC were isolated from a normal donor by centrifugation on a Ficoll gradient. The cells were stimulated with immobilized OKT3 antibody for 3 days. They were cotransduced by means of particle-mediated gene transfer with 1mg each of pCMV-CAT (CMV enhancer [Enh.]/promoter [Prom.] driving a CAT reporter) and the indicated Rev M10 orDRev M10 expression plasmid. The cells were incubated in the presence of recombinant human IL-2 (50 mg/ml) for 1 day. Cells in each group were divided equally. Each group was challenged with HIV-1Bruat a multiplicity of infection of 0.35, while the other served as an uninfected reference control. Cells were harvested after 3 days of incubation, and CAT activity in cell lysates was determined. Schematic diagrams of the Rev M10 andDRev M10 expression vectors are shown. Intr., intron.
VOL. 71, 1997 REGULATION OF ANTIVIRAL GENE EXPRESSION 7027
on November 9, 2019 by guest
http://jvi.asm.org/
genetic interventions for AIDS. The protective effects of this strategy could be attributed to at least three properties of the promoter. Rev M10 is believed to retain the RRE binding property; however, it is defective in Rev multimerization, a process that is critical for recruiting cellular factors to promote viral mRNA transport (37). The TAR sequence present in the promoter could enhance the basal levels of transcription a few-fold, thereby synthesizing more Rev M10. The TAR motif, on the other hand, may also sequester the available levels of Tat within the cell, thereby indirectly suppressing Tat transac-tivation of the HIV LTR. Such combinations of antiviral genes and appropriate regulatory elements may optimize the effects of gene products which may inhibit HIV replication and con-tribute to gene therapy for AIDS.
ACKNOWLEDGMENTS
We thank Donna Gschwend for manuscript preparation and Nancy Barrett for preparation of figures.
This work was supported in part by NIH grant AI36606 to G.J.N.
REFERENCES
1.Bahner, I., K. Kearns, Q. L. Hao, E. M. Smogorzewska, and D. B. Kohn.
1996. Transduction of human CD341hematopoietic progenitor cells by a retroviral vector expressing an RRE decoy inhibits human immunodeficiency virus type 1 replication in myelomonocytic cells produced in long-term cul-ture. J. Virol.70:4352–4360.
2.Bahner, I., C. Zhou, X. Yu, Q. Hao, J. C. Guatelli, and D. B. Kohn.1993. Comparison oftrans-dominant inhibitory mutant human immunodeficiency virus type 1 genes expressed by retroviral vectors in human T lymphocytes. J. Virol.67:3199–3207.
3.Berkhout, B., and K. Jeang.1992. Functional roles for the TATA promoter and enhancers in basal andtat-induced expression of the human immuno-deficiency virus type 1 long terminal repeat. J. Virol.66:139–149. 4.Bevec, D., M. Dobrovnik, J. Hauber, and E. Bohnlein.1992. Inhibition of
human immunodeficiency virus type 1 replication in human T cells by ret-roviral-mediated gene transfer of a dominant-negative Rev transactivator. Proc. Natl. Acad. Sci. USA89:9870–9874.
5.Bevec, D., B. Volc-Platzer, K. Zimmermann, M. Dobrovnik, J. Hauber, G. Veres, and E. Bohnlein.1994. Constitutive expression of chimeric neo-Rev response element transcripts suppresses HIV-1 replication in human CD41 T lymphocytes. Hum. Gene Ther.5:193–201.
6.Boshart, M., F. Weber, G. Jahn, K. Dorsch-Hasler, B. Fleckenstein, and W. Schaffner.1985. A very strong enhancer is located upstream of an immediate early gene of human cytomegalovirus. Cell41:521–530.
7.Bowen, D. L., H. C. Lane, and A. S. Fauci.1985. Immunopathogenesis of the acquired immunodeficiency syndrome. Ann. Intern. Med.103:704–709. 8.Buchman, A. R., and P. Berg.1988. Comparison of intron-dependent and
intron-independent gene expression. Mol. Cell. Biol.8:4395–4405. 9.Bunnell, B. A., L. M. Muul, R. E. Donahue, R. M. Blaese, and R. A. Morgan.
1995. High-efficiency retroviral-mediated gene transfer into human and non-human primate peripheral blood lymphocytes. Proc. Natl. Acad. Sci. USA
92:7739–7743.
10. Caruso, M., and D. Klatzmann.1992. Selective killing of CD41cells har-boring a human immunodeficiency virus-inducible suicide gene prevents viral spread in an infected cell population. Proc. Natl. Acad. Sci. USA
89:182–186.
11. Chapman, B. S., R. M. Thayer, K. A. Vincent, and N. L. Haigwood.1991. Effect of intron A from human cytomegalovirus (Towne) immediate-early gene on heterologous expression in mammalian cells. Nucleic Acids Res.
19:3979–3986.
12. Chen, C., and H. Okayama.1987. High-efficiency transformation of mam-malian cells by plasmid DNA. Mol. Cell. Biol.7:2745–2752.
13. Curiel, T. J., D. R. Cook, Y. Wang, B. H. Hahn, S. K. Ghosh, and G. S. Harrison.1993. Long-term inhibition of clinical and laboratory human im-munodeficiency virus strains in human T-cell lines containing an HIV-regu-lated diphtheria toxin A chain gene. Hum. Gene Ther.4:741–747. 14. Dalgleish, A. G., P. C. Beverley, P. R. Clapham, D. H. Crawford, M. F.
Greaves, and R. A. Weiss.1984. The CD4(T4) antigen is an essential com-ponent of the receptor for the AIDS retrovirus. Nature312:763–767. 15. Duan, L., O. Bagasra, M. A. Laughlin, J. W. Oakes, and R. J. Pomerantz.
1994. Potent inhibition of human immunodeficiency virus type 1 replication by an intracellular anti-Rev single-chain antibody. Proc. Natl. Acad. Sci. USA91:5075–5079.
16. Escaich, S., C. Kalfoglou, I. Plavec, S. Kaushal, J. D. Mosca, and E. Bohn-lein.1995. Rev M10-mediated inhibition of HIV-1 replication in chronically infected T cells. Hum. Gene Ther.6:625–634.
17. Fauci, A. S.1988. The human immunodeficiency virus: infectivity and
mech-anisms of pathogenesis. Science239:617–622.
18. Feinberg, M. B., R. F. Jarrett, A. Aldovini, R. C. Gallo, and F. Wong-Staal.
1986. HTLV-III expression and production involve complex regulation at the levels of splicing and translation of viral RNA. Cell46:807–817.
19. Fox, B. A., C. Woffendin, Z. Y. Yang, H. San, U. Ranga, D. Gordon, J. Osterholzer, and G. J. Nabel.1995. Genetic modification of human periph-eral blood lymphocytes with a transdominant negative form of Rev: safety and toxicity. Hum. Gene Ther.6:997–1004.
20. Ghazal, P., H. Lubon, B. Fleckenstein, and L. Hennighausen.1987. Binding of transcription factors and creation of a large nucleoprotein complex on the human cytomegalovirus enhancer. Proc. Natl. Acad. Sci. USA84:3658–3662. 21. Goodbourn, S.1994. T-cell activation: transcriptional regulation in activated
T cells. Curr. Biol.4:930–932.
22. Grosschedl, R., and D. Baltimore.1985. Cell-type specificity of immunoglob-ulin gene expression is regulated by at least three DNA sequence elements. Cell41:885–897.
23. Harada, S., Y. Koyanagi, H. Nakashima, N. Kobayashi, and N. Yamamoto.
1986. Tumor promoter, TPA, enhances replication of HTLV-III/LAV. Vi-rology154:249–258.
24. Harrison, G. S., C. J. Long, T. J. Curiel, F. Maxwell, and I. H. Maxwell.1992. Inhibition of human immunodeficiency virus-1 production resulting from transduction with a retrovirus containing an HIV-regulated diphtheria toxin A chain gene. Hum. Gene Ther.3:461–469.
25. Hoxie, J. A., J. D. Alpers, J. L. Rackowski, K. Huebner, B. S. Haggerty, A. J. Cedarbaum, and J. C. Reed.1986. Alterations in T4 (CD4) protein and mRNA synthesis in cells infected with HIV. Science234:1123–1127. 26. Huang, L. and K. Jeang.1993. Increased spacing between Sp1 and TATAA
renders human immunodeficiency virus type 1 replication defective: impli-cation for Tat function. J. Virol.67:6937–6944.
27. Huang, M. T., and C. M. Gorman.1990. Intervening sequences increase efficiency of RNA 39processing and accumulation of cytoplasmic RNA. Nucleic Acids Res.18:937–947.
28. Jamieson, C., P. G. McCaffrey, A. Rao, and R. Sen.1991. Physiologic acti-vation of T cells via the T cell receptor induces NF-kB. J. Immunol.147:
416–420.
29. Kessinger, A.1995. Circulating stem cells—waxing hematopoietic. N. Engl. J. Med.333:315–316.
30. Killeen, N., S. Sawada, and D. R. Littman.1993. Regulated expression of human CD4 rescues helper T cell development in mice lacking expression of endogenous CD4. EMBO J.12:1547–1553.
31. Lee, T. C., B. A. Sullenger, H. F. Gallardo, G. E. Ungers, and E. Gilboa.1992. Overexpression of RRE-derived sequences inhibits HIV-1 replication in CEM cells. New Biol.4:66–74.
32. Lemob, D., A. Angeretti, P. Foresta, G. Gribaudo, M. Gariglio, and S. Landolfo.1994. Trans-activation of the mouse cytomegalovirus immediate early gene enhancer by ras oncogenes. J. Gen. Virol.75:1685–1692. 33. Liem, S. E., A. Ramezani, X. Li, and S. Joshi.1993. The development and
testing of retroviral vectors expressing transdominant mutants of HIV-1 proteins to confer anti-HIV-1 resistance. Hum. Gene Ther.4:625–634. 34. Lisziewicz, J., D. Sun, J. Smythe, P. Lusso, F. Lori, A. Louie, P. Markham,
J. Rossi, M. Reitz, and R. C. Gallo.1993. Inhibition of human immunode-ficiency virus type 1 replication by regulated expression of a polymeric Tat activation response RNA decoy as a strategy for gene therapy in AIDS. Proc. Natl. Acad. Sci. USA90:8000–8004.
35. Liu, J., C. Woffendin, Z. Yang, and G. J. Nabel.1994. Regulated expression of a dominant negative form of Rev improves resistance to HIV replication in T cells. Gene Ther.1:32–37.
36. Malim, M. H., S. Bohnlein, J. Hauber, and B. R. Cullen.1989. Functional dissection of the HIV-1 Rev trans-activator—derivation of a trans-dominant repressor of Rev function. Cell58:205–214.
37. Malim, M. H., and B. R. Cullen.1991. HIV-1 structural gene expression requires the binding of multiple Rev monomers to the viral RRE: implica-tions for HIV-1 latency. Cell65:241–248.
38. Malim, M. H., W. W. Freimuth, J. Liu, T. J. Boyle, H. K. Lylerly, B. R. Cullen, and G. J. Nabel.1992. Stable expression of transdominant rev pro-tein in human T cells inhibits human immunodeficiency virus replication. J. Exp. Med.176:1197–1201.
39. Malim, M. H., J. Hauber, R. Fenrick, and B. R. Cullen.1988. Immunode-ficiency virus rev trans-activator modulates the expression of the viral regu-latory genes. Nature335:181–183.
40. Malim, M. H., D. F. McCarn, L. S. Tiley, and B. R. Cullen.1991. Mutational definition of the human immunodeficiency virus type 1 Rev activation do-main. J. Virol.65:4248–4254.
41. Marasco, W. A., W. A. Haseltine, and S. Y. Chen.1993. Design, intracellular expression, and activity of a human anti-human immunodeficiency virus type 1 gp120 single-chain antibody. Proc. Natl. Acad. Sci. USA90:7889–7893. 42. Markovitz, D. M., M. Hannibal, V. L. Perez, C. Gauntt, T. M. Folks, and
G. J. Nabel.1990. Differential regulation of human immunodeficiency vi-ruses: a novel HIV-2 regulatory element responds to stimulation of the T-cell antigen receptor. Proc. Natl. Acad. Sci. USA87:9098–9102.
43. Mhashilkar, A. M., J. Bagley, S. Y. Chen, A. M. Szilvay, D. G. Helland, and W. A. Marasco.1995. Inhibition of HIV-1 Tat-mediated LTR transactivation
on November 9, 2019 by guest
http://jvi.asm.org/
and HIV-1 infection by anti-Tat single chain intrabodies. EMBO J.14:1542– 1551.
44. Modesti, N., J. Garcia, C. Debouck, M. Peterlin, and R. Gaynor.1991. Trans-dominant Tat mutants with alterations in the basic domain inhibit HIV-1 gene expression. New Biol.3:759–768.
45. Muesing, M., D. H. Smith, and D. J. Capon.1987. Regulation of mRNA accumulation by a human immunodeficiency virus trans-activator protein. Cell48:691–701.
46. Nabel, G. J., B. A. Fox, L. Post, C. B. Thompson, and C. Woffendin.1994. Clinical protocol: a molecular genetic intervention for AIDS—effects of a transdominant negative form of Rev. Hum. Gene Ther.5:79–92. 47. Ojwang, J. O., A. Hampel, D. J. Looney, F. Wong-Staal, and J. Rappaport.
1992. Inhibition of human immunodeficiency virus type 1 expression by a hairpin ribozyme. Proc. Natl. Acad. Sci. USA89:10802–10806.
48. Pearson, L., J. Garcia, F. Wu, N. Modesti, J. Nelson, and R. Gaynor.1990. A transdominant tat mutant that inhibits tat-induced gene expression from the human immunodeficiency virus long terminal repeat. Proc. Natl. Acad. Sci. USA87:5079–5083.
49. Pomerantz, R. J., D. Trono, M. B. Feinberg, and D. Baltimore.1990. Cells nonproductively infected with HIV-1 exhibit an aberrant pattern of viral RNA expression: a molecular model for latency. Cell61:1271–1276. 50. Rosen, C. A., J. G. Sodroski, and W. A. Haseltine.1985. The location of
cis-acting regulatory sequences in the human T cell lymphotropic virus type III (HTLV-III/LAV) long terminal repeat. Cell41:813–823.
51. Rosenberg, S. A., P. Aebersold, K. Cornetta, A. Kasid, R. A. Morgan, R. Moen, E. M. Karson, M. T. Lotze, J. C. Yang, S. L. Topalian, M. J. Merino, K. Culver, A. D. Miller, R. M. Blaese, and W. F. Anderson.1990. Gene transfer into humans—immunotherapy of patients with advanced melanoma, using tumor-infiltrating lymphocytes modified by retroviral gene transduc-tion. N. Engl. J. Med.323:570–578.
52. Salmons, B., and W. H. Gunzburg.1993. Targeting of retroviral vectors for gene therapy. Hum. Gene Ther.4:129–141.
53. Sambrook, J., E. F. Fritch, and T. Maniatis.Molecular cloning: a laboratory manual, 2nd ed. 1994. Cold Spring Laboratory Harbor Press, Cold Spring Harbor, N.Y.
54. Sambucetti, L. C., J. M. Cherrington, G. W. G. Wilkinson, and E. S. Mo-carski.1989. NF-kappaB activation of the cytomegalovirus enhancer is me-diated by a viral transactivator and by T cell stimulation. EMBO J.8:4251– 4258.
55. Sarver, N., E. M. Cantin, P. S. Chang, J. A. Zaia, P. A. Ladne, D. A. Stephens, and J. J. Rossi.1990. Ribozymes as potential anti-HIV-1 thera-peutic agents. Science247:1222–1225.
56. Sawada, S., and D. R. Littman.1991. Identification and characterization of a T-cell-specific enhancer adjacent to the murine CD4 gene. Mol. Cell. Biol.
11:5506–5515.
57. Sawada, S., and D. R. Littman.1993. A heterodimer of HEB and an E12-related protein interacts with the CD4 enhancer and regulates its activity in T-cell lines. Mol. Cell. Biol.13:5620–5628.
58. Selby, M. J., E. S. Bain, P. A. Luciw, and B. M. Peterlin.1989. Structure, sequence, and position of the stem-loop in tar determine transcriptional
elongation by tat through the HIV-1 long terminal repeat. Genes Dev.
3:547–558.
59. Siu, G., A. L. Wurster, J. S. Lipsick, and S. M. Hedrick.1992. Expression of the CD4 gene requires a Myb transcription factor. Mol. Cell. Biol.12:1592– 1604.
60. Sodroski, J., W. C. Goh, C. Rosen, A. Dayton, E. Terwilliger, and W. Hasel-tine.1986. A second post-transcriptional trans-activator gene required for HTLV-III replication. Nature321:412–417.
61. Southgate, C. D., and M. R. Green.1991. The HIV-1 Tat protein activates transcription from an upstream DNA-binding site: implications for Tat func-tion. Genes Dev.5:2496–2507.
62. Spangrude, G. J., S. Heimfeld, and I. L. Weissman.1988. Purification and characterization of mouse hematopoietic stem cells. Science241:58–62. 63. Sullenger, B. A., H. F. Gallardo, G. E. Ungers, and E. Gilboa.1990.
Over-expression of TAR sequences renders cells resistant to human immunode-ficiency virus replication. Cell63:601–608.
64. Sweet, R. W., A. Truneh, and W. A. Hendrickson.1991. CD4: its structure, role in immune function and AIDS pathogenesis, and potential as a phar-macological target. Curr. Opin. Biotechnol.2:622–633.
65. Trono, D., M. Feinberg, and D. Baltimore.1989. HIV-1 gag mutants can dominantly interfere with the replication of the wild type virus. Cell59:113– 120.
66. Van Ooyen, A., J. Van Den Berg, N. Mantel, and C. Weissmann.1979. Comparison of total sequence of a cloned rabbitb-globin gene and its flanking regions with a homologous mouse sequence. Science206:337–344. 67. Vandendriessche, T., M. K. Chuah, L. Chiang, H. K. Chang, B. Ensoli, and R. A. Morgan.1995. Inhibition of clinical human immunodeficiency virus (HIV) type 1 isolates in primary CD41T lymphocytes by retroviral vectors expressing anti-HIV genes. J Virol69:4045–4052.
68. Venkatesh, L. K., and G. Chinnadurai.1990. Mutants in a conserved region near the carboxy-terminus of HIV-1 Rev identify functionally important residues and exhibit a dominant negative phenotype. Virology178:327–330. 69. Woffendin, C., U. Ranga, Z. Yang, L. Xu, and G. J. Nabel.1996. Expression of a protective gene prolongs survival of T cells in human immunodeficiency virus-infected patients. Proc. Natl. Acad. Sci. USA93:2889–2894. 70. Woffendin, C., Z. Yang, Udaykumar, L. Xu, N. Yang, M. J. Sheehy, and G. J.
Nabel.1994. Non-viral and viral delivery of a human immunodeficiency virus protective gene into primary human T cells. Proc. Natl. Acad. Sci. USA
91:11581–11585.
71. Yu, M., J. Ojwang, O. Yamada, A. Hampel, J. Rapapport, D. Looney, and F. Wong-Staal.1993. A hairpin ribozyme inhibits expression of diverse strains of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA90:
6340–6344.
72. Zagury, D., J. Bernard, R. Leonard, R. Cheynier, M. Fledman, P. S. Sarin, and R. C. Gallo.1986. Long-term cultures of HTLV-III-infected T cells: a model of cytopathology of T-cell depletion in AIDS. Science231:850–853. 73. Zhou, C., I. C. Bahner, G. P. Larso, J. A. Zaia, J. J. Rossi, and D. B. Kohn.
1994. Inhibition of HIV-1 in human T-lymphocytes by retrovirally trans-duced anti-tat and rev hammerhead ribozymes. Gene149:33–39.
VOL. 71, 1997 REGULATION OF ANTIVIRAL GENE EXPRESSION 7029