0022-538X/11/$12.00 doi:10.1128/JVI.00178-11
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
The L-Coding Region of the DA Strain of Theiler’s Murine
Encephalomyelitis Virus Causes Dysfunction and Death
of Myelin-Synthesizing Cells
䌤
G. D. Ghadge,
1R. Wollmann,
2G. Baida,
1M. Traka,
1,3and R. P. Roos
1,3*
Departments of Neurology1and Pathology2and University of Chicago Center for Peripheral Neuropathy,3The University of
Chicago Medical Center, 5841 S. Maryland Ave., Chicago, Illinois 60637
Received 26 January 2011/Accepted 1 July 2011
The DA strain and other members of the TO subgroup of Theiler’s murine encephalomyelitis virus (TMEV) induce an early transient subclinical neuronal disease followed by a chronic progressive inflammatory demy-elination, with persistence of the virus in the central nervous system (CNS) for the life of the mouse. Although TMEV-induced demyelinating disease (TMEV-IDD) is thought to be immune mediated, there is also evidence that supports a role for the virus in directly inducing demyelination. In order to clarify the function of DA virus genes, we generated a transgenic mouse that had tamoxifen-inducible expression of the DA L-coding region in oligodendrocytes (and Schwann cells), a cell type in which the virus is known to persist. Tamoxifen-treated young transgenic mice usually developed an acute progressive fatal paralysis, with abnormalities of the oligodendrocytes and Schwann cells and demyelination, but without significant lymphocytic infiltration; later treatment led to transient weakness with demyelination and persistent expression of the recombined transgene. These findings demonstrate that a high level of expression of DA L can cause the death of myelin-synthesizing cells and death of the mouse, while a lower level of L expression (which can persist) can lead to cellular dysfunction with survival. The results suggest that expression of DA L plays an important role in the pathogenesis of TMEV-IDD. Virus-induced infection and death of oligodendrocytes may play a part in the demyelination of other diseases in which an immune-mediated mechanism has been stressed, including multiple sclerosis.
Theiler’s murine encephalomyelitis virus (TMEV) is a mem-ber of theTheilovirusspecies of theCardiovirusgenus of the family ofPicornaviridae(reviewed in reference 20). The En-cephalomyocarditis virus species of the Cardiovirus genus in-cludes the closely related encephalomyocarditis virus (EMCV) and mengovirus (MV). TMEV strains can be divided into two subgroups on the basis of their differing biologic properties. The GDVII strain and other members of the GDVII subgroup are highly virulent and produce a rapidly fatal neuronal infec-tion in mice, with no persistence of the virus. In contrast, DA, BeAn, and other members of the less virulent TO subgroup induce an early transient subclinical neuronal disease followed by a chronic progressive inflammatory demyelination, with per-sistence of the virus in the central nervous system (CNS) for the life of the mouse. During TMEV-induced demyelinating disease (TMEV-IDD), relatively large amounts of the TMEV genome can be detected in oligodendrocytes and microglia, with little evidence of viral antigen or infectious virus, i.e., there is a restricted expression of DA viral proteins. TMEV-IDD is an excellent model of multiple sclerosis (MS) because of the similarity in the demyelinating pathologies and because the immune system appears to contribute to pathology in both disorders. Although TMEV-IDD is thought to be immune mediated, there is also evidence that supports a key role for the
virus infection in demyelination: the virus persists throughout the demyelinating disease, primarily in oligodendrocytes and microglia; DA virus antigen and virions have been identified in oligodendrocytes (the CNS cells that make myelin), which have a “dying back” pathology (25, 26); and DA virus is known to cause demyelination in infected nude mice (29, 30).
In order to clarify the functions of selected DA virus genes, we previously generated a mouse that expresses a subgenomic region of the virus as a transgene in myelin-synthesizing cells– oligodendrocytes and Schwann cells. We hypothesized that this transgenic mouse might show a phenotype of interest, thereby clarifying the functions of one or more of the expressed viral genes within myelinating cells. In order to control the time of transgene expression and to avoid tolerance in future immu-nological studies, we made the expression inducible. To achieve the intended site-specific and inducible expression, we prepared double transgenic mice, called DA/Cre mice, which carried a tamoxifen-inducible Cre recombinase protein under the control of the myelin proteolipid protein (PLP) transcrip-tional control element along with a subgenomic region of DA virus. The subgenomic region, which was preceded by a floxed transcription stop, included the DA 5⬘ untranslated region (5⬘-UTR), the leader (L) protein-coding region, a region that encodes an out-of-frame protein (L*) important for TMEV-IDD (5, 34), and P1 (the capsid-coding region) (1). Interest-ingly, tamoxifen-treated young DA/Cre mice developed an acute progressive fatal paralysis with abnormalities of the oligodendrocytes and Schwann cells and demyelination but with-out significant lymphocytic infiltration. This result suggested
* Corresponding author. Mailing address: Department of Neurol-ogy, The University of Chicago Medical Center, 5841 S. Maryland Ave., MC2030, Chicago, IL 60637. Phone: (773) 702-5659. Fax: (773) 834-9889. E-mail: [email protected].
䌤Published ahead of print on 13 July 2011.
9377
on November 7, 2019 by guest
http://jvi.asm.org/
that a gene(s) in this DA subgenomic region was capable of causing death of myelin-synthesizing cells without a clear contribution from the immune system. Additionally of note, treatment of DA/Cre mice at a later age led to transient weakness with persistent expression of the recombined transgene in the CNS, suggesting that this gene could persist in myelin-synthesizing cells, causing dysfunction without cell death, resembling the situation in DA virus-infected demy-elinated mice.
We hypothesized that the pathogenic segment of the DA subgenomic transgene was the DA L-coding region because the L protein of cardioviruses is multifunctional, with patho-genic properties. L protein of cardioviruses has been shown to inhibit type I interferon (IFN) gene transcription (11, 35, 41), inhibit nucleocytoplasmic transport and host cell trans-lation (6, 23), and regulate cell apoptosis (9, 27, 36).
In order to clarify whether the DA L-coding region was the region within the DA subgenomic transgene that caused the white matter disease in DA/Cre mice, we generated another transgenic mouse that only expressed DA L downstream of the floxed transcription stop, i.e., the transgene lacked the DA 5⬘-UTR and P1. As in the case of the previous DA subgenomic region, DA L was inducibly expressed in myelin-synthesizing cells after tamoxifen treatment. We now report that this trans-genic mouse, called DAL/Cre, showed a clinical and patholog-ical picture similar to that for DA/Cre mice following early induction of DA L with tamoxifen, i.e., an acute progressive fatal paralysis with demyelination and without significant lym-phocytic infiltration, indicating that the pathogenic region of the transgene in DA/Cre mice was the DA L-coding region. Of interest, there was evidence of death of myelin-synthesizing cells and apoptosis of cells in the peripheral nervous system (PNS) and CNS. Administration of a lower dose of tamoxifen caused dysfunction of myelin-synthesizing cells without cell death, similar to that seen with tamoxifen-treated older DA/ Cre mice. These results suggest that the expression of L plays a major role in the pathogenesis of TMEV-IDD. A virus in-fection and gene directly causing the death or dysfunction of oligodendrocytes may also underlie other virus-induced demy-elinating diseases in which an immune-mediated mechanism has been stressed (19, 43), as well as immune-mediated dis-eases in which virus involvement is not certain, such as MS (2, 10, 18).
MATERIALS AND METHODS
Transgenic mice.A DAL transgene (Fig. 1A) that contains the chicken-actin promoter followed by a floxed transcription terminator, then followed by the DA L-coding region, and finally followed by the globin poly(A) signal, was
con-structed in the following way. (i) The plasmid pBS246loxP, which contained an
efficient transcription terminator surrounded byloxPsites, was engineered as
previously described (1). (ii) The DA L-coding region fragment was amplified from a full-length infectious clone of DA, pDAFL3 (28), using the following primers, which contained NheI enzyme cleavage sites as well as an overlapping
Kozak sequence: forward, 5⬘-AGCAACCGCTAGCCACCATGGCTTGCAAA
CATGGATAC-3⬘; reverse, 5⬘-AGCAACCGCTAGCTTACTGGGGTTCCAT
GACAATATCG-3⬘. The PCR product was digested with NheI and cloned into
SpeI-digested pBS246loxP⬃30 nucleotides downstream from the 3⬘-flanking
loxP site. (iii) The transcription terminator sequence and DA L-coding region
fragment within pBS246loxP were removed from the resulting plasmid with
NotI and inserted into a derivative of pCAGGS that contained a NotI site (1). The resultant DAL transgene was linearized, purified by agarose gel electropho-resis, and used for pronuclear injection. Transgenic mice were produced by the
University of Chicago Transgenic Mouse Core as described previously (1). At 15 to 18 days, a tail biopsy specimen was taken to determine the genotype of the pups by
using the following primers in a PCR: forward, 5⬘-ATGTGTGCCCTATTTGCAC
AGC-3⬘; reverse, 5⬘-ACGAGCGGTAAGTCAGTCCATT-3⬘. Transgene-positive
pups (founders) were saved and bred to make lines.
Transgenic mice carrying DA L were crossed with PLP/CreERT
mice (7), which have the proteolipid protein (PLP) transcriptional control element driving expression in myelin-synthesizing cells of Cre recombinase protein fused to a mutated ligand-binding domain of the human estrogen receptor, thereby
con-ferring tamoxifen inducibility. The presence of the PLP/CreERT
transgene was detected as previously described (7), and the DAL transgene was detected by PCR using the above primers. Double transgenic mice, called DAL/Cre mice, were inoculated intraperitoneally with tamoxifen each day for 5 days for a total
dose of tamoxifen that varied in different experiments between 15 and 50g/g
body weight. The tamoxifen solution was prepared by dissolving 50 mg of 4-hy-droxytamoxifen (Sigma, St. Louis, MO) into 0.2 ml dimethyl sulfoxide (DMSO) and 0.3 ml ethanol (EtOH) and then adding 4.5 ml sunflower seed oil (Sigma) followed by a 20-min sonication. At the time of sacrifice, mice were anesthetized and in some cases perfused with phosphate-buffered saline (PBS) followed by 4% paraformaldehyde (pH 7.3).
RNA analysis.RNA was isolated from various tissues by extraction with TRIzol (Invitrogen, Carlsbad, CA), followed by purification using RNeasy (Qiagen, Valen-cia, CA). The RNA was subjected to reverse transcriptase PCR (RT-PCR) using random hexamers and Superscript III Platinum (Invitrogen) to generate cDNA. The cDNA was used in a PCR (Eppendorf Mastermix) with the same primers shown above, and the amplified products were analyzed by agarose gel electrophoresis.
Teased fiber preparation, histopathology, and electron microscopy.Following sacrifice, the brain, optic nerves, spinal cord, and sciatic nerve were collected, processed for light and electron microscopic examination with paraffin or Epon embedding, and sectioned using standard procedures. In some cases, the sciatic nerves were dissected and prepared for teased fiber analysis, as described pre-viously (1).
Immunohistochemical analysis.At various times after tamoxifen treatment, animals were sacrificed and tissues collected and processed for terminal
deoxy-FIG. 1. The DA L transgene and its spatial and temporal expres-sion. (A) Schematic representation (not drawn to scale) of the DALflox construct (top) showing theloxPsites (which flank an upstream tran-scription stop) as triangles and the recombined DAL (bottom) after Cre-induced recombination. (B) RT-PCR analysis of RNA isolated from spinal cords (sc) and lungs of representative DAL/Cre and lit-termate control mice. Twelve- to 16-week-old mice were treated for 5 days with a total dose of 25g/g (lanes 1 to 6) or 15g/g (lanes 7 to 12) of tamoxifen (Tmx) or no drug (lanes 13 and 14). The presence of the PLP/CreERTand L transgenes is noted above the gel. 1862 and 1864 mice were sacrificed when moribund⬃20 days after the last day of tamoxifen treatment, while 1830 and 1842 were sacrificed⬃100 days after tamoxifen. 1849, which carried only the PLP/CreERTtransgene, and 1870, an untreated double transgenic mouse, were sacrificed when
⬃150 days old. The predicted ⬃180-bp product of L mRNA was detected in the spinal cords (lanes 3, 5, 9, and 11), but not lungs from tamoxifen-treated double transgenic mice and not from lungs or sc from untreated nontransgenic or treated single transgenic mice.
on November 7, 2019 by guest
http://jvi.asm.org/
nucleotidyltransferase-mediated dUTP-biotin nick end labeling (TUNEL) assay (Millipore Corp., Billerica, MA) and immunohistochemical staining using rabbit cleaved caspase-3 antibody (Cell Signaling Technology, Danvers, MA), anti-Iba1 antibody (Wako Pure Chemical Industries, Osaka, Japan), and anti-CD3 rat monoclonal antibody (17A2, sc-18843; Santa Cruz Biotechnology, Santa Cruz, CA) with an ABC Vectastain kit (Vector Laboratories, Burlingame, CA).
RESULTS
Double transgenic DAL/Cre mice were prepared as de-scribed in Materials and Methods. DAL/Cre mice contained the PLP/CreERTtransgene along with the DA L-coding region under the control of the chicken-actin promoter and down-stream of a floxed transcription stop site. Cre is predicted to enter the nuclei of myelinating cells of tamoxifen-treated dou-ble transgenic mice and recombine out the transcription ter-minator upstream of the DA L-coding region, thereby allowing spatially and temporally controlled expression of DA L in myelin-synthesizing cells (Fig. 1A).
Agarose gel electrophoresis of the amplified product of an RT-PCR of total RNA from the spinal cord of DAL/Cre mice treated at age 15 weeks with 25g/g tamoxifen and sacrificed⬃20 days later showed a band of the size predicted following Cre-induced recombination (Fig. 1B, lanes 3 and 5); no band was detected in RNA from nonneural tissue (Fig. 1B, lanes 4 and 6). As in the case with DA/Cre mice, CNS tissues from surviving DAL/Cre mice that had been treated at age 12 or 16 weeks with 15g/g tamoxifen had evidence of persistent expression of the recombined DA segment (Fig. 1B, lanes 9 and 11) for as long as 3 months (the longest time sampled), demonstrating that some cells expressing DA L had not died. In contrast, RT-PCR of lungs from these mice as well as CNS tissue from noninduced double transgenic mice and from tamoxifen-treated single transgenic mice showed no evidence of recombination after 30 cycles of amplification (Fig. 1B).
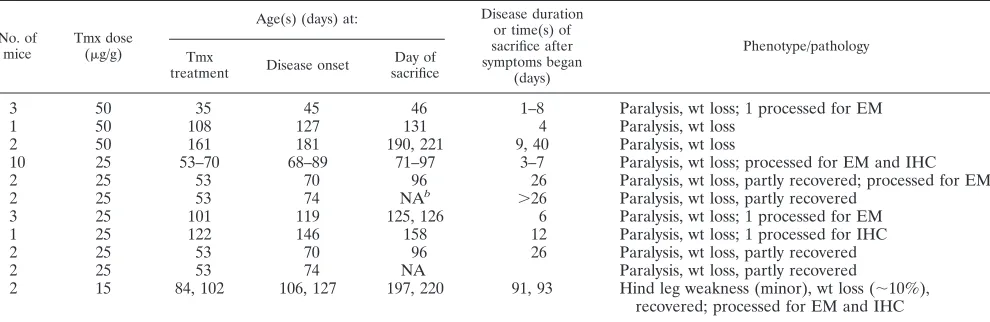
Tamoxifen-treated double transgenic DAL/Cre mice develop weakness.The tamoxifen-treated DAL/Cre mice (Table 1) had a clinical and pathological phenotype similar to that of tamox-ifen-treated DA/Cre mice (1). DAL/Cre mice that were 8 to 10
weeks of age and treated with 25 to 50 g/g of tamoxifen typically began to show weakness of their hind limbs 1 to 2 weeks after the last tamoxifen injection. This weakness pro-gressed rapidly to involve the forelimbs. Animals with severe weakness had shallow respirations and usually became mori-bund within a week after the onset of signs, at which time they were euthanized.
In order to facilitate the survival of mice, we supported mice during their time of maximal weakness and also treated older DAL/Cre mice with a lower dose of tamoxifen. DAL/Cre mice that were 12 to 16 weeks of age and treated with 15g/g of tamoxifen remained unaffected or tended to develop subtle weakness and weight loss, which generally appeared within 2 to 3 weeks of the last tamoxifen injection, and recovered over time. No weakness was observed in DAL/Cre mice that were not tamoxifen treated or in single transgenic mice that were tamoxifen treated.
Tamoxifen-treated double transgenic DAL/Cre mice that be-come paralyzed and die show demyelination with dysfunction or death of oligodendrocytes and Schwann cells. Moribund paralyzed tamoxifen-treated DAL/Cre mice were euthanized, and the sciatic nerves, optic nerves, brains, and spinal cords were processed for histological examination. Individual my-elinated axons teased from the sciatic nerve showed prominent segmental demyelination (Fig. 2A). Epon-embedded sections of the sciatic nerve (Fig. 2B) confirmed the presence of demy-elination of peripheral nerve axons seen in the teased fiber preparations: there was an increase in unmyelinated axons compared to results for tamoxifen-treated control mice, as well as abnormally thin myelinated axons (indicating remyelina-tion) and degenerating Schwann cells (Fig. 2B). Frequently surrounding the thinly myelinated axons were fully myelinated axons and normal-appearing Schwann cells, presumably be-cause the recombination mediated by Cre is incomplete and random in removing DNA segments betweenloxPsites. Ab-normalities of CNS myelin and oligodendrocytes were easiest to detect in the optic nerves. The optic nerves tended to have clusters of unmyelinated axons, abnormally thin myelinated sheaths of axons, and degenerating oligodendrocytes with
ev-TABLE 1. Tamoxifen-treated DAL/Cre micea
No. of mice
Tmx dose
(g/g)
Age(s) (days) at: Disease duration
or time(s) of sacrifice after symptoms began
(days)
Phenotype/pathology Tmx
treatment Disease onset
Day of sacrifice
3 50 35 45 46 1–8 Paralysis, wt loss; 1 processed for EM
1 50 108 127 131 4 Paralysis, wt loss
2 50 161 181 190, 221 9, 40 Paralysis, wt loss
10 25 53–70 68–89 71–97 3–7 Paralysis, wt loss; processed for EM and IHC
2 25 53 70 96 26 Paralysis, wt loss, partly recovered; processed for EM
2 25 53 74 NAb ⬎26 Paralysis, wt loss, partly recovered
3 25 101 119 125, 126 6 Paralysis, wt loss; 1 processed for EM
1 25 122 146 158 12 Paralysis, wt loss; 1 processed for IHC
2 25 53 70 96 26 Paralysis, wt loss, partly recovered
2 25 53 74 NA Paralysis, wt loss, partly recovered
2 15 84, 102 106, 127 197, 220 91, 93 Hind leg weakness (minor), wt loss (⬃10%), recovered; processed for EM and IHC
a
For each group of mice, littermates of DAL/Cre mice (not shown) were used as controls and either treated with tamoxifen (Tmx) or not treated. EM, electron microscopic evidence of findings detailed in the text; IHC, immunohistochemistry with staining for TUNEL, microglia, and caspase-3.
b
NA, not applicable (mice still alive).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.44.539.82.243.2]idence of myelin debris (Fig. 2C and D). Immunohistochemical studies showed a proliferation and infiltration of microglial cells in the sciatic nerves (Fig. 2E), spinal cord white matter (Fig. 2G), and optic nerves (Fig. 2I) of DAL/Cre mice com-pared to controls (Fig. 2F, H, and J), with no evidence of T cell infiltration (data not shown).
Because L of BeAn, a TO subgroup strain closely related to DA, induces apoptosis of cultured BHK-21 cells (9), we TUNEL-stained PNS and CNS tissues for evidence of apop-tosis. There was TUNEL staining of cells in the sciatic nerves (Fig. 3A), spinal cords (Fig. 3B), and optic nerves (Fig. 3C) of weak DAL/Cre mice following tamoxifen treatment. Sections of these same tissues as well as nerve roots had evidence of activated caspase-3 immunostaining (Fig. 3D to G), confirming the apoptosis. Of interest, the activated caspase-3 staining of nerves and nerve roots (Fig. 3E) was more prominent than the staining of oligodendrocytes in the CNS white matter (Fig. 3F and G). In addition, the activated caspase-3 staining was more prominent than TUNEL staining. No TUNEL staining or ac-tivated caspase-3 immunostaining was seen in tissues from littermate controls (data not shown).
Surviving tamoxifen-treated DAL/Cre mice exhibit demye-lination and microgliosis.At times, DAL/Cre mice that were treated with tamoxifen became weak and survived, especially when they were supported during the time of maximal weak-ness. These mice tended to have prominent abnormalities of the Schwann cells and myelin of the sciatic nerve (data not shown) that resembled findings in paralyzed moribund tamox-ifen-treated mice (Fig. 2A and B). At times, robust white matter abnormalities were also present in the CNS. Figures 4C and D, 5C and D, and 6 show sections from CNS tissues of two DAL/Cre mice that were treated with tamoxifen when 53 days of age, became paralyzed at 70 days of age, and were sacrificed when partly recovered at 96 days of age. There were prominent white matter abnormalities in the corpus callosum (Fig. 4C, 5C, and 6A) and optic nerve (Fig. 4D, 5D, and 6B and D), with demyelinated or thinly myelinated axons and degenerating oli-godendrocytes. In addition, there were swollen dystrophic ax-ons filled with mitochondria and lysosomes and, at times, sur-rounded by thin myelin (Fig. 4C and 6A). The white matter abnormalities in the corpus callosum and optic nerve were more extensive than in the spinal cords (data not shown) of these mice. In addition, the CNS pathology in some of the mice that partly recovered from their initial paralysis was more prominent than the CNS pathology in mice that were treated with high doses of tamoxifen at a young age and sacrificed when paralyzed and moribund (Fig. 2C and D). The changes that were seen in these tamoxifen-treated DAL/Cre mice were not apparent in littermate controls (Fig. 4A and B and 5A and B).
DAL/Cre mice that were treated with a lower dose of ta-moxifen (15g/g) developed no disease or had subtle transient weakness. When these mice were sacrificed 3 months after tamoxifen treatment, Epon-embedded sections of the sciatic nerves showed no obvious pathology. Abnormalities of the optic nerve, however, were apparent, with an increase in non-myelinated axons and abnormally thin myelin sheaths (Fig. 7A and B). Iba1 immunohistochemical staining demonstrated that the optic nerve had an increase in activated microglia and macrophages (Fig. 7C) compared to the optic nerves of litter-mate controls (Fig. 7D). There was no TUNEL staining or activated caspase-3 staining of the sciatic nerves, spinal cords, or optic nerves (data not shown) of these tamoxifen-treated or control mice at this time.
FIG. 2. Pathology of representative paralyzed tamoxifen-treated DAL/Cre mice. DAL/Cre mice were treated with 25 to 50g/g tamox-ifen at 5 to 17 weeks of age, became weak 5 to 19 days later, and were sacrificed when moribund 1 to 12 days later. Littermate mice that contained only one or no transgene were similarly treated with tamox-ifen and sacrificed at similar times and were used as controls. (A) Teased fiber of sciatic nerve of tamoxifen-treated DAL/Cre mouse shows demyelinated segments (arrows). (B) Electron micrograph of sciatic nerve of tamoxifen-treated DAL/Cre mouse shows thinly my-elinated axons (some with asterisks) and a Schwann cell with myelin debris (arrows). (C) Electron micrograph of optic nerve from tamox-ifen-treated DAL/Cre mouse showing a cluster of demyelinated and thinly myelinated axons (some with asterisks). (D) Electron micro-graph of optic nerve from tamoxifen-treated DAL/Cre mouse shows degenerating oligodendrocyte with myelin debris (arrows). Immuno-histochemical Iba1 staining of macrophages and microglia of sciatic nerve (E), spinal cord posterior column (G, arrows), and optic nerve (I) from a tamoxifen-treated DAL/Cre mouse. Similar staining was also performed on a sciatic nerve (F), spinal cord (H), and optic nerve (J) from a littermate control mouse.
on November 7, 2019 by guest
http://jvi.asm.org/
DISCUSSION
The DA strain and other members of the TO subgroup of TMEV produce a chronic inflammatory CNS demyelinating disease that is thought to be immune mediated; however, ev-idence also supports a key role for the virus in carrying out the demyelination (reviewed in reference 20). The relative contri-bution in TMEV-IDD of a direct virus-mediated oligodendro-cyte dysfunction/lysis vis-a`-vis immune-mediated demyelina-tion remains unclear, as is the case with other demyelinating diseases (19, 43), including MS (2, 10, 18). In order to clarify the functions of selected DA virus genes, we made use of the Cre/loxP system to provide inducible and cell-type-specific
ex-pression of DA L in oligodendrocytes (and Schwann cells), a cell type in which DA virus is known to persist.
[image:5.585.113.473.71.236.2]Our previous studies involved inducible expression of a DA subgenomic region (DA 5⬘UTRLL*P1) as a transgene in DA/Cre mice (1). Induction of the transgene in myelin-synthesizing cells of young mice led to rapidly fatal paralysis with demyelination and abnormalities of oligodendrocytes and Schwann cells, but without significant T cell infiltrates. Mice that survived or mice that were treated with tamoxifen at a later age also had evidence of demyelination and remy-elination when sacrificed. In order to delineate the patho-genic region within the DA5⬘UTRLL*P1 subgenomic seg-ment, we prepared DAL/Cre mice, which express only the
FIG. 3. Apoptosis in tissues of tamoxifen-treated DAL/Cre mice. DAL/Cre mice were treated with 25g/g tamoxifen at 8 to 10 weeks of age, became weak⬃10 days later, and were sacrificed when moribund 7 to 12 days later. Shown is TUNEL staining (A to C) and activated caspase-3 immunohistochemical staining (D to G) of the sciatic nerves (A and D), nerve roots (E, arrowheads), spinal cord posterior columns (B and F, arrows), and optic nerves (C and G) of paralyzed moribund tamoxifen-treated DAL/Cre mice.
[image:5.585.44.284.460.641.2]FIG. 4. CNS white matter pathology in tamoxifen-treated DAL/ Cre mice that partly recovered from paralysis. DAL/Cre mice were treated with 25g/g tamoxifen at 53 days of age, became weak at 70 days of age, and were sacrificed when partly recovered at 96 days of age. Electron micrographs of the corpus callosum (A and C) and optic nerve (B and D) in tamoxifen-treated DAL/Cre mice (C and D) and a littermate control (A and B). Demyelinated and thinly myelinated axons and dystrophic axons (marked by X) are shown in panels C and D (for higher magnification, see Fig. 5 and 6).
FIG. 5. CNS white matter pathology in tamoxifen-treated DAL/Cre mice that partly recovered from paralysis. Demyelinated and thinly my-elinated axons (some designated with asterisks) are present in tamoxifen-treated DAL/Cre mice (C and D) that were tamoxifen-treated as described in Fig. 4 but not in a littermate control (A and B). Panel C shows a degenerating oligodendrocyte (surrounded by arrows), which is shown at a lower mag-nification in Fig. 4C, containing an abnormal myelin membrane (arrow-heads). (A and C) Corpus callosum; (B and D), optic nerve.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.302.542.468.651.2]DA L-coding region as a transgene. We found that the clinical and pathological findings for DAL/Cre mice were very similar to those for DA/Cre mice, indicating that the DA L-coding region was the pathogenic segment in the DA subgenomic transgene in DA/Cre mice. The findings empha-size the potential importance of DA L-induced dysfunction and death of oligodendrocytes in TMEV-IDD.
Recent studies have demonstrated that cardiovirus L protein has a number of activities that are likely to contribute to dis-ease: inhibition of type I IFN gene transcription (11, 35, 41), inhibition of cellular mRNA export from the nucleus leading to interference with translation (6, 23), and regulation of apop-tosis (9, 27, 36). Recentin vitrostudies have shown that trans-fection of an L expression construct of BeAn into BHK-21 cells leads to cell death and apoptosis (9) and that DA L is apoptotic in both BHK-21 and HeLa cells (36). Apoptosis has been identified in varied neural cells in the CNS after TMEV infec-tion of mice, both early after infecinfec-tion and during TMEV-IDD (3, 4, 33, 39); however, the TMEV gene responsible for the apoptosis in infected neural cells has not been identified. The present study is the first to demonstrate that expression of DA L causes apoptosis of cells in the PNS and CNS white matter of mice. We found greater caspase-3 staining than TUNEL staining of these cells, perhaps reflecting a failure to imple-ment the full apoptotic machinery, as is the case in certain cultured cells infected with TMEV (36). We presume that at least some of the apoptotic cells are Schwann cells and oligo-dendrocytes since L is expressed only in these cells and since there was electron microscopic evidence of degeneration of these cells (Fig. 2B and 6B to D) with what appeared to be apoptotic changes of the nucleus (Fig. 6C); however, we were unsuccessful in carrying out double label immunohistochemi-cal studies, perhaps due to the inhibition of translation of cell type-specific markers caused by L.
It is of interest that animals supported during their acute
weakness as well as older animals treated with a low dose of tamoxifen survived despite the continuing expression of DA L. We suspect that these mice survived because Cre recombinase-mediated recombination occurred in a more limited number of myelin-synthesizing cells within an individual animal. Also of interest was the prolonged survival of myelin-synthesizing cells despite the continuing expression of L. We presume that these cells did not die despite continuing expression of L because only some of the multiple copies of the floxed transgene within an individual cell were recombined, resulting in lower levels of L expression in some cells, i.e., low levels of expression of DA L allowed a cell to survive (while high levels killed the cell).
Mice that became paralyzed following tamoxifen treatment but survived had evidence of demyelination of the corpus cal-losum and optic nerve with microgliosis (Fig. 4 to 6 and 7C). Interestingly, despite the spatial restriction of DA L to myelin-synthesizing cells, tamoxifen-treated DAL/Cre mice had abun-dant evidence of swollen axons with dystrophic neurites filled with mitochondria and lysosomes (Fig. 4C and 6A). Our data, as well as those of the two recent studies (22, 38) that used transgenic mice identical or similar to the present one demon-strate that axonal damage can occur following a restricted oligodendrocyte injury in the absence of T cells. Axonal dam-age has been described in TMEV-IDD and MS, including the presence of dystrophic axons (15, 31). The DAL/Cre mouse model may be of value in studies of demyelination and
remy-FIG. 7. Pathology in optic nerves of DAL/Cre mice 3 months after tamoxifen treatment. (A) Electron micrograph of the optic nerve of a DAL/Cre mouse that was treated with 15g/g tamoxifen at 12 weeks of age, became weak 17 days after the last tamoxifen injection, and was sacrificed 3 months later. The optic nerve has a large cluster of unmy-elinated axons (asterisks mark some of them). (B) Electron micro-graph of the optic nerve of a DAL/Cre mouse that was treated with 15
[image:6.585.299.539.69.289.2]g/g tamoxifen at 16 weeks of age, became weak 20 days after last tamoxifen injection, and was sacrificed 3 months later. The optic nerve has a cluster of unmyelinated and thinly myelinated axons (asterisks). (C) Immunohistochemical Iba1 staining of the optic nerve of the same mouse as in Fig. 4A. (D) Immunohistochemical Iba1 staining of the optic nerve of a littermate control mouse treated similarly to the mouse in Fig. 4C.
FIG. 6. CNS white matter pathology in tamoxifen-treated DAL/ Cre mice that partly recovered from paralysis. Dystrophic axons (marked by X) in the corpus callosum (A) and degenerating oligoden-drocytes in the optic nerve (B and D) and spinal cord (C) are present in tamoxifen-treated DAL/Cre mice (that were treated as described for Fig. 4). The nucleus in panel C is densely stained and appears apop-totic.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.43.283.70.252.2]elination caused by a single viral gene targeting myelin-synthe-sizing cells.
The phenotype of DAL/Cre (and DA/Cre) mice (i.e., CNS demyelination with persistent expression of DA L in oligoden-drocytes) resembled that seen in TMEV-IDD, in which demy-elination is associated with persistence of the virus genome in oligodendrocytes (and microglia) and little production of in-fectious virus. The similar phenotypes indicate the potential importance of DA L to TMEV-IDD pathogenesis. Myelin protein synthesis may be disturbed during TMEV-IDD be-cause of an inhibition by L of mRNA exit (6, 23) from the nuclei of oligodendrocytes and also because of suppression of host cell protein synthesis by type I IFN, which is present at a low level in the CNS of DA virus-infected chronically demy-elinated mice (35). There is presumably a balance between L and type I IFN expression, so that type I IFN inhibits the synthesis of viral proteins, including L, but type I IFN tran-scription is inhibited by DA L (35). Of note, abnormalities in translation underlie nonviral causes of demyelination, such as those involving ER stress (which can lead to a block in trans-lation) in inherited demyelinating diseases (17, 32, 40). Myelin-synthesizing cells may be especially vulnerable to translational abnormalities because of the large amount of membrane pro-tein that needs to be synthesized by these cells.
It is of interest that DAL/Cre mice had prominent periph-eral nerve demyelination with Schwann cell degeneration, pre-sumably because DA L was expressed in Schwann cells (as well as oligodendrocytes). These abnormalities may have led to the paralysis and subsequent death of mice treated early with ta-moxifen. Although peripheral nerve pathology is not a feature of mice infected intracerebrally with DA virus, intrasciatic nerve injection with DA virus is known to cause peripheral nerve demyelination (8). The peripheral nerve disease in ta-moxifen-treated DAL/Cre (and DA/Cre) transgenic mice tended to be more prominent than the CNS disease; for ex-ample, there were more cells that had prominent activated caspase-3 antibody staining in the nerve roots (Fig. 3E) than in the spinal cords of moribund tamoxifen-treated DAL/Cre mice (Fig. 3F). The finding of greater pathology in the PNS than the CNS contrasts with findings in two recent studies in mice that carried a transgene similar or identical to PLP/CreERT (22, 38). In these studies, which involved inducible and myelin-synthesizing cell-specific expression of the diphtheria toxin A subunit, disease of the white matter of the CNS was prominent, with little or no mention of pathology in the PNS. The greater pathology in the PNS seen in DAL/Cre mice may relate to the use in our study of a lower dosage of tamoxifen (allowing for greater recombination in the periphery). It may also be that DAL/Cre mice died before the bulk of CNS white matter pathology became apparent since the reaction to injury by peripheral nerves is greatly accelerated compared to that found in oligodendrocytes (42). This possibility garners sup-port from our finding of prominent CNS demyelination in tamoxifen-treated mice that partly recovered and were sacri-ficed 26 days following the onset of significant weakness (Fig. 4 to 6).
Although some tamoxifen-treated DAL/Cre mice had prom-inent CNS demyelination (Fig. 4 to 6), the demyelination dif-fered from that seen in TMEV-IDD. Tamoxifen-treated DAL/ Cre mice had scattered demyelinated axons, presumably
because the Cre-induced recombination randomly occurred in myelin-synthesizing cells. In contrast, demyelinated areas in mice with TMEV-IDD are well-demarcated regions, perhaps related to the spread of virus. Tamoxifen-treated DAL/Cre mice with prominent CNS demyelination had more demyeli-nation in the optic nerves and brain than the spinal cord, presumably because the activity of the PLP gene transcription control region driving Cre varies in location depending on the age of the mouse (16). In contrast, the brunt of the white matter disease in the case of TMEV-IDD is in the spinal cord, presumably as a result of the intracerebral inoculation of virus followed by trafficking of the virus. It may also be that the demyelination in tamoxifen-treated DAL/Cre mice differs from TMEV-IDD because T cells are present in the lesions in TMEV-IDD, but not in DAL/Cre mice. Abnormalities of T cells, including T regulatory cells (24) and Th17 cells (14), play a role in TMEV persistence and disease pathogenesis. In ad-dition, a persistent TMEV infection of microglia is important in initiating of immune responses, including a myelin-specific immune response (21), and in generating a proinflammatory immune response (13). Furthermore, L* protein, which is ini-tiated in the L-coding region and is out of frame with the polyprotein, is important for efficient growth of TMEV in microglia (37) and in antagonizing the antivirus immune re-sponse (reviewed in reference 20).
Our findings bring up the possibility that other demyelinat-ing diseases, and perhaps MS, may also be associated with direct oligodendrocyte dysfunction from a pathogen. Of note, oligodendrocyte death and apoptosis in the absence of T cell infiltration have been reported to be an early feature of MS (12). Furthermore, a recent genomics study of MS identified the apoptotic regulator geneBCL2as important for suscepti-bility (44).
ACKNOWLEDGMENT
This work was partly supported by R01 NS037958 (R.P.R.).
REFERENCES
1.Baida, G., et al.2008. A subgenomic segment of Theiler’s murine
encepha-lomyelitis virus RNA causes demyelination. J. Virol.82:5879–5886.
2.Brahic, M.2010. Multiple sclerosis and viruses. Ann. Neurol.68:6–8. 3.Buenz, E. J., et al.2009. Apoptosis of hippocampal pyramidal neurons is
virus independent in a mouse model of acute neurovirulent picornavirus
infection. Am. J. Pathol.175:668–684.
4.Carlson, N. G., K. E. Hill, I. Tsunoda, R. S. Fujinami, and J. W. Rose.2006. The pathologic role for COX-2 in apoptotic oligodendrocytes in virus in-duced demyelinating disease: implications for multiple sclerosis. J.
Neuro-immunol174:21–31.
5.Chen, H. H., W. P. Kong, L. Zhang, P. L. Ward, and R. P. Roos.1995. A picornaviral protein synthesized out of frame with the polyprotein plays a key role in a virus-induced immune-mediated demyelinating disease. Nat. Med.
1:927–931.
6.Delhaye, S., V. van Pesch, and T. Michiels.2004. The leader protein of Theiler’s virus interferes with nucleocytoplasmic trafficking of cellular
pro-teins. J. Virol.78:4357–4362.
7.Doerflinger, N. H., W. B. Macklin, and B. Popko.2003. Inducible site-specific
recombination in myelinating cells. Genesis35:63–72.
8.Drescher, K. M., and S. M. Tracy.2007. Injection of the sciatic nerve with
TMEV: a new model for peripheral nerve demyelination. Virology359:233–
242.
9.Fan, J., K. N. Son, S. Y. Arslan, Z. Liang, and H. L. Lipton.2009. Theiler’s murine encephalomyelitis virus leader protein is the only nonstructural pro-tein tested that induces apoptosis when transfected into mammalian cells.
J. Virol.83:6546–6553.
10.Gilden, D. H.2005. Infectious causes of multiple sclerosis. Lancet Neurol.
4:195–202.
11.Hato, S. V., et al.2007. The mengovirus leader protein blocks interferon-alpha/beta gene transcription and inhibits activation of interferon regulatory
factor 3. Cell. Microbiol.9:2921–2930.
on November 7, 2019 by guest
http://jvi.asm.org/
12.Henderson, A. P., M. H. Barnett, J. D. Parratt, and J. W. Prineas.2009. Multiple sclerosis: distribution of inflammatory cells in newly forming
le-sions. Ann. Neurol.66:739–753.
13.Himeda, T., T. Okuwa, Y. Muraki, and Y. Ohara.2010. Cytokine/chemokine profile in J774 macrophage cells persistently infected with DA strain of
Theiler’s murine encephalomyelitis virus (TMEV). J. Neurovirol.16:219–
229.
14.Hou, W., H. S. Kang, and B. S. Kim.2009. Th17 cells enhance viral persis-tence and inhibit T cell cytotoxicity in a model of chronic virus infection. J.
Exp. Med.206:313–328.
15.Kornek, B., et al.2000. Multiple sclerosis and chronic autoimmune enceph-alomyelitis: a comparative quantitative study of axonal injury in active,
in-active, and remyelinated lesions. Am. J. Pathol.157:267–276.
16.Leone, D. P., et al. 2003. Tamoxifen-inducible glia-specific Cre mice for somatic mutagenesis in oligodendrocytes and Schwann cells. Mol. Cell.
Neu-rosci.22:430–440.
17.Lin, W., et al.2007. The integrated stress response prevents demyelination by protecting oligodendrocytes against immune-mediated damage. J. Clin.
Invest.117:448–456.
18.Lipton, H. L., Z. Liang, S. Hertzler, and K. N. Son.2007. A specific viral
cause of multiple sclerosis: one virus, one disease. Ann. Neurol.61:514–523.
19.Matthews, A. E., E. Lavi, S. R. Weiss, and Y. Paterson.2002. Neither B cells nor T cells are required for CNS demyelination in mice persistently infected
with MHV-A59. J. Neurovirol.8:257–264.
20.Michiels, T., and R. P. Roos.2010. Theiler’s virus central nervous system
infection, p. 411–428.InE. Ehrenfeld, E. Domingo, and R. P. Roos (ed.),
The picornaviruses. ASM Press, Washington, DC.
21.Olson, J. K., and S. D. Miller.2004. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs. J. Immunol.
173:3916–3924.
22.Pohl, H. B. F., et al.2011. Genetically induced adult oligodendrocyte cell death is associated with poor myelin clearance, reduced remyelination, and
axonal damage. J. Neurosci.31:1069–1080.
23.Porter, F. W., Y. A. Bochkov, A. J. Albee, C. Wiese, and A. C. Palmenberg.
2006. A picornavirus protein interacts with Ran-GTPase and disrupts
nucle-ocytoplasmic transport. Proc. Natl. Acad. Sci. U. S. A.103:12417–12422.
24.Richards, M. H., et al.2011. Virus expanded regulatory T cells control disease severity in the Theiler’s virus mouse model of MS. J. Autoimmun.
36:142–154.
25.Rodriguez, M.1985. Virus-induced demyelination in mice: “dying back” of
oligodendrocytes. Mayo Clinic Proc.60:433–438.
26.Rodriguez, M., J. L. Leibowitz, and P. W. Lampert.1983. Persistent infection of oligodendrocytes in Theiler’s virus-induced encephalomyelitis. Ann.
Neu-rol.13:426–433.
27.Romanova, L. I., et al.2009. Antiapoptotic activity of the cardiovirus leader
protein, a viral “security” protein. J. Virol.83:7273–7284.
28.Roos, R. P., et al.1990. Infectious Theiler’s virus cDNA clones: intratypic
recombinants and viral polyprotein processing, p. 128–134.InM. A. Brinton
and F. X. Heinz (ed.), New aspects of positive-strand RNA viruses. Amer-ican Society for Microbiology, Washington, DC.
29.Roos, R. P., and R. Wollmann.1984. DA strain of Theiler’s murine
enceph-alomyelitis virus induces demyelination in nude mice. Ann. Neurol.15:494–
499.
30.Rosenthal, A., R. S. Fujinami, and P. W. Lampert.1986. Mechanism of
Theiler’s virus-induced demyelination in nude mice. Lab. Invest.54:515–522.
31.Sathornsumetee, S., D. B. McGavern, D. R. Ure, and M. Rodriguez.2000. Quantitative ultrastructural analysis of a single spinal cord demyelinated lesion predicts total lesion load, axonal loss, and neurological dysfunction in
a murine model of multiple sclerosis. Am. J. Pathol.157:1365–1376.
32.Scheper, G. C., M. S. van der Knaap, and C. G. Proud.2007. Translation matters: protein synthesis defects in inherited disease. Nat. Rev. Genet.
8:711–723.
33.Schlitt, B. P., M. Felrice, M. L. Jelachich, and H. L. Lipton.2003. Apoptotic cells, including macrophages, are prominent in Theiler’s virus-induced
in-flammatory, demyelinating lesions. J. Virol.77:4383–4388.
34.Stavrou, S., et al.2010. Theiler’s murine encephalomyelitis virus L* amino acid position 93 is important for virus persistence and virus-induced
demy-elination. J. Virol.84:1348–1354.
35.Stavrou, S., Z. Feng, S. M. Lemon, and R. P. Roos.2010. Different strains of Theiler’s murine encephalomyelitis virus antagonize different sites in the
type I interferon pathway. J. Virol.84:9181–9189.
36.Stavrou, S., G. Ghadge, and R. P. Roos.2011. Apoptotic and antiapoptotic activity of L protein of Theiler’s murine encephalomyelitis virus. J. Virol.
85:7177–7185.
37.Takata, H., et al.1998. L* protein of the DA strain of Theiler’s murine encephalomyelitis virus is important for virus growth in a murine
macro-phage-like cell line. J. Virol.72:4950–4955.
38.Traka, M., et al.2010. A genetic mouse model of adult-onset, pervasive
central nervous system demyelination with robust remyelination. Brain133:
3017–3029.
39.Tsunoda, I., C. I. Kurtz, and R. S. Fujinami.1997. Apoptosis in acute and chronic central nervous system disease induced by Theiler’s murine
enceph-alomyelitis virus. Virology228:388–393.
40.van der Knaap, M. S., J. C. Pronk, and G. C. Scheper.2006. Vanishing white
matter disease. Lancet Neurol.5:413–423.
41.van Pesch, V., O. van Eyll, and T. Michiels.2001. The leader protein of Theiler’s virus inhibits immediate-early alpha/beta interferon production.
J. Virol.75:7811–7817.
42.Vargas, M. E., and B. A. Barres.2007. Why is Wallerian degeneration in the
CNS so slow? Annu. Rev. Neurosci.30:153–179.
43.Wu, G. F., A. A. Dandekar, L. Pewe, and S. Perlman.2000. CD4 and CD8 T cells have redundant but not identical roles in virus-induced demyelination.
J. Immunol.165:2278–2286.
44.Zuvich, R. L., et al.2010. Genetic variation in the IL7RA/IL7 pathway
increases multiple sclerosis susceptibility. Hum. Genet.127:525–535.