0022-538X/79/06-0692/09$02.00/0
Effect of
Temperature-Sensitive
Mutation on
Activity
of
the
RNA
Transcriptase
of
Vesicular Stomatitis Virus
New Jersey
J. F. SZILAGYI* AND C. R. PRINGLE
Medical Research CouncilVirologyUnit,Instituteof Virology, University of Glasgow, Glasgow Gll 5JR, Scotland
Received forpublication23January 1979
The virion-associated RNA transcriptase activity ofvesicularstomatitis virus New Jersey temperature-sensitive (ts) mutants was assayed in vitro at the permissive (3100) andrestrictive (3900) temperatures. RNA synthesis at390C
bytheRNA-negativets Al andtheRNA-positive tsCl andtsDl mutants was
similar to that ofwild-typevirus. The RNA-negativetsBlsynthesized only small amountsof RNA in vitro at390C.The three mutants ofcomplementationgroup E were dissimilar in the amounts of RNA they synthesized at 390C: ts El synthesized very little RNA, ts E2 synthesized moderate amounts, and RNA synthesis by ts E3 was not inhibited. The two mutants of group F were also
dissimilar, since ts Fl synthesized very little RNA at 390C, whereas ts F2
synthesized asmuch RNA aswild-typevirus. The revertant clones ts Bl/Rl, ts El/Rl, and tsFl/Rlsynthesized RNA at
390C
in amounts comparable towild-typevirus, indicatingthat the heatsensitivity of thetranscriptaseactivity of the
mutants ts Bl, ts El, and ts Fl was associated with temperature sensitivity.
Similarheat sensitivities were observed when transcribing nucleoprotein
com-plexeswereused in the assays,showingthat the mutatedpolypeptideswerepart
of the viral core. The heatstability of the mutant ts Bl was similar to that of wild-type virus, and in vitro RNA synthesis wasfullyrestored when the temper-aturewasloweredto 310Cafter 30 min ofpreincubation at390C, showing that the inhibition was due to reversible configurational change of the mutated
polypeptide.When virions ofthemutant tsElwereheated for5hat
390C,
theirinfectivity and transcriptase activity were as stable as those of the wild-type virus, whereastranscriptaseactivitybecame very heat labile afterdisruption of the viral coatwith a neutraldetergent. This suggests an interaction between the mutated
polypeptideanda coatpolypeptidewhich stabilizes theactivityof the
transcrip-tase. The RNAtranscriptase activity of themutant tsFl was also heatlabile,
althoughto alesserextentthanthat of tsEl.Thus, the defects intranscriptase
activityof groupsB,E, and F suggest thatall threepolypeptidesofthe virus core,
polypeptides L, N, and NS, are involved in the transcription. In addition, we
postulatethat themutated gene products of groups E and Faremultifunctional,
being requiredboth intranscriptionandreplication,and that the geneproductof
group E mayalso be involved insomelate stage of virusdevelopment. Temperature-sensitive (ts) mutants of
vesic-ular stomatitis virus (VSV) Indiana have been classified into six non-overlapping
complemen-tation groups (3, 4, 13, 19). At the restrictive temperature
(390C),
mutants ofcomplementa-tion groups I and IV do not synthesizevirion
RNA, whereasmutantsofgroups IIIandV do (15). GroupHisuniqueinthat it contains mu-tants of both RNA-negativeand RNA-positive phenotypes (15). Theviriontranscriptase
activ-ity of several tsmutants was comparedin vitro
at 31 and
390C
by Szilagyi and Pringle, whofound thatmutants ofgroupsII, III,and IVall synthesized RNA at
390C,
indicating that inthese groups the RNA transcriptase was not
affectedby the mutation (20). However, several
mutantsin group I did notsynthesize RNA in
vitroat390Cbecause of the heatlability oftheir
transcriptase (20).Studies ofprimaryRNA
tran-scriptionininfected cellsconfirmed thatin
com-plementation group Ithe mutation altered the
activity of the RNAtranscriptase (18).
Dissocia-tion and reconstitution experiments indicated
thatpolypeptideL wasthe siteofthe mutational lesion(2, 5, 6).Recentlyweexaminedmutant ts
045 from the RNA-positive complementation
group V (17,24) and mutant ts M602from the
RNA-negativegroupVI(7,8,19)and foundthat
692
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 30, 1979
bothmutantssynthesizedRNA in vitro at390C, indicating that the transcriptase activityin these
groupsis not affected by themutation (unpub-lished data).
ts mutants isolated from VSV New Jersey have also been classifiedinto six non-overlap-ping complementationgroups by Pringle etal. (16). Complementation groupsA, B, and F did not synthesize virion RNA at the restrictive temperature(3900),whereasmutantsofgroups
C and D did (16). Complementation group E
contained both RNA-negative and RNA-posi-tive mutants, since two of themutants (tsEl and ts E3) did not synthesize virion RNA at
39°C whereas the third mutant (ts E2) did (16). Lesnaw and Reichmann usedone mutantfrom each of the sixcomplementationgroups toshow that ininfected cellstwo of the RNA-negative
mutants, tsAlandtsEl,wereableto carry out
primary transcription but not replication of vir-ionRNA at39.50C (10). The RNA-positive
mu-tants tsCl andtsDl synthesized virion RNAat
the restrictive temperature, whereas mutants from the othertwoRNA-negative
complemen-tationgroups(tsBlandtsFl) exhibited neither transcription nor replication (10). Recently
Les-naw and Dicksonhave carried out in vitro dis-sociation and reconstitution experiments which indicate that the mutational lesion in ts Bl involves the L or NS polypeptides (9).
Conditional temperature-dependent (td CE) mutantshave been isolated which grow both at 31and 390C in BHK cells but only at 310C in secondary chicken embryo cells (14, 21). Some of thetd CE mutantsof VSV New Jersey syn-thesize RNAinvitro at390C similarly to wild-typevirus, while others do not (21). One mutant of the latter category, td CE3, was studied in
somedetail, and it was shown that the activity
ofitstranscriptase was fully and instantaneously reversible bylowering the temperature from 39
to310C (21). Thus, the mode of inhibition of the
transcriptase of this mutant differs from that
observed in some of the group I mutants of VSV
Indiana, where thermal inactivation was
irre-versible (20). Dissociation and reconstitution ex-periments showed that the td CE3 mutation affectedpolypeptide L(22).
Inthiscommunication we examine the
kinet-ics of in vitroRNAsynthesis by purified virions
of the conventional ts mutants of VSV New Jerseytodetermine whether virion transcriptase activity was defective in any of the six
comple-mentationgroups.
MATERIALS AND METHODS
Mutants. The tsmutantsof VSVNewJersey de-scribedby Pringleetal.(16)wereused. We choseone
mutantfrom groupsA, B,and C (ts Al,tsBH,andts C1), the onlymutant of group D (ts D1), allthree mutantsof group E(tsEl,tsE2,andtsE3),and both mutantsof group F (ts Fl andtsF2). We also isolated revertantclones frommutants tsBi,tsEl,andtsFl. These revertants(tsBl/Rl,tsEl/Ri,andtsFl/Ri) were isolated from BS-C-1 monolayersincubatedat 390C.
Growthandpurificationof virus.BHK-21clone 13monolayersin 6to 10Burrler bottleswereinfected withlow multiplicity of themutantsand incubatedat
310C. Growth andpurification followed the method describedby SzilagyiandPringle (20). Thepurified virus wassuspendedin 20 mMTris-hydrochloricacid buffer (pH 8.0), the amount of the protein in this purified preparationwasdeterminedbythe method of Lowry etal. (11), and the concentration ofproteinin eachsuspensionwasadjustedto1.3mg/ml. Infectivity assayswerecarriedoutunder agaroverlay using BS-C-1 cellmonolayers.
Thermalinactivation ofinfectivity.Purified vi-rus preparations (0.005ml)weresuspendedin1mlof Eagle mediumcontaining 10% fetal calfserum and wereincubatedat390Cfor 5 h. The residualinfectivity of thesesuspensionswastitrated at310ConBS-C-1 cell monolayers, andresults areexpressed as a per-centageof the infectivityofidenticalsampleswhich werekept at00Cfor thesamelengthoftime.
TNP preparations. Transcribing nucleoprotein (TNP)complexeswerepreparedfrompurifiedvirions bythe method ofSzilagyiandUryvayev (23) usingthe modifications described bySzilhgyiandPringle (21). Assay of theRNAtranscriptase.Thepreviously described RNAtranscriptaseassays(1, 12, 20, 21, 23) were modified. Incubation mixtures (0.3 ml) were made upat00Cbyaddingthefollowing ingredients: calculated volume of 20 mMTris-hydrochloridebuffer (pH 8.0) tomakethe final volume0.3ml,0.03mlof TNP or purifiedvirus suspension, 0.015 ml of 0.8% Triton-N 101 (added when virus was used but not when TNPwasused),0.015mlof70mM dithiothrei-tol,0.006ml ofSearle "ribonuclease inhibitor" prepa-ration (50units/ml),0.018ml of 32 mMATP,0.0075 ml of1 mMS-adenosyl methionine, and 0.075 ml of "reagent mixture" (containing Tris-hydrochloride buffer, pH8.0,NaCl, actinomycinD,ATP, CTP,GTP, and3H-labeled UTP) (22). The incubation mixtures werethen placed in 31 or390Cwaterbaths for 1 min, and RNA synthesis was started by the addition of 0.0075ml of 220 mMMgCl2. Samples (0.02ml) were taken eitherbefore the addition ofMgCl2 (zero-time sample) or at intervals duringincubation and were placed on Whatman DE-81 paper disks. The disks werewashedatroomtemperaturefivetimeswith5% sodiumpyrophosphate, twice with water, twice with ethanol, and twice withether,andradioactivitywas measured by liquid scintillation spectrophotometry. Thermal inactivation of transcriptase. Two identical incubation mixtures containingallthe ingre-dients exceptMgCl2wereprepared for eachmutant. One incubation mixture of eachpair was incubated at
390C for30minwhile the otherwaskept inan ice-waterbath.Thetranscriptase activity of both mixtures wasassayed at310Cafter the additionofMgCl2, and theamountofRNAsynthesized by the heat-treated
on November 10, 2019 by guest
http://jvi.asm.org/
incubation mixture during 3 h was expressed as a percentage of theamountsynthesized by itscontrol.
RESULTS
Temperature sensitivity of the mutants
of VSV NewJersey. Throughout the
experi-mentsdescribed in thiscommunication,weused purifiedpreparations of the varioustsmutants
of VSV NewJersey. Inpreliminary experiments wenoticed that somepreparationsof mutant ts E1, and to a lesser extent ts D1, appeared to
have ahigh frequency ofrevertantvirus. This
difficultywasovercomebyrecloningthese
mu-tants. The purified preparations of the ts mu-tants used in the experiments exhibited a ts
phenotype,and the revertant clones showed ef-ficiencies ofplatingat39°Csimilartowild-type virus.
Invitro RNAsynthesisby thets mutants. Todetermine the temperaturesensitivityof the
transcriptaseactivityof the varioustsmutants
ofVSV New Jersey, we assayed the virion-as-sociated RNA transcriptase in vitro at 31 and
390Candcomparedthe resultstothose obtained
with wild-type virus. These experiments were
repeatedthreetimes,and the results ofasingle
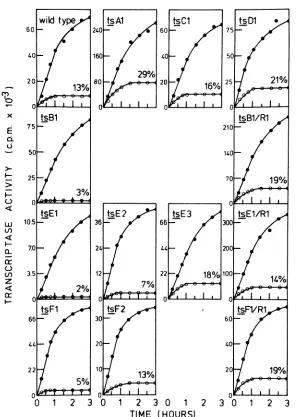
experiment are shown in Fig. 1. The results
obtained in.the othertwoexperimentswere in verygood agreement with thosegiveninFig.1.
Wild-typevirussynthesized RNAat310C
lin-earlyfor1h,and RNAsynthesiscontinuedat a
reduced ratethroughout the 3 h of the experi-ment. At390Ctheinitialrateof RNAsynthesis wasabouthalf of therate at
310C,
and aplateauwasreachedbyabout60min, the total amount of RNAproducedbeingabout13 to16% of that synthesized at310C.
Mutant ts Al consistently synthesized more RNAat390Cthan thewild-typevirus. Mutants
tsC1 andtsDlsynthesizedRNAat390C
simi-larlyorslightlybetter thanwild-typevirus.
In the case of ts B1, the mutation almost
certainlyaffected apolypeptideinvolved in the
transcriptionprocess,since thismutant
synthe-sizedverylittle RNA at390C (only 2 to4%of
thatsynthesizedat
310C),
whereas itsrevertant,tsBl/Rl, synthesizedRNA at
390C
atleastaswellaswild-type virus.
Likewise, mutant ts El synthesized verysmall
amounts ofRNAat
390C,
whereas theamountofRNAsynthesizedat390Cby its revertant,ts
El/Ri,
was comparable to that produced bywild-typevirus. This indicates that themutated
polypeptideofts Elisinvolvedintranscription.
However, mutant ts E3 synthesized RNA nor-mallyat390C, and tsE2synthesized RNA at a
somewhat reduced rate.
Therewaslimited RNA synthesisat
390C
bymutant tsFl, about 5% of the amount synthe-sized at310C. Since the revertant clone (tsFl/ Rl) of thismutant synthesized RNA as well as or better than the wild-type virus, it is likely that the mutated polypeptide of ts Fl is also involved inthe transcription process. However, mutant ts F2synthesized RNA at390C similarly towild-type virus.
These experiments were repeated using TNP complexes of the tsmutants. These TNP prep-arations are ribonucleoprotein complexes iso-lated from purified virions (23). They contain only threepolypeptides(L, N, and NS) in close association with the virion RNA and retain the transcriptaseactivity of the virion (23). Results with the TNPs of the ts mutants were very similar to those given in Fig. 1, indicating that the temperature-sensitivepolypeptides of ts Bl, tsEl,andtsFlarepart of theviral core (Table 1).
Thermalstabilityofthetranscriptase
ac-tivity of the ts mutants.The mode of
inhibi-tion of RNAsynthesisat390Cwasinvestigated by examining the heatstabilityofthe
transcrip-taseactivityof thetsmutants.Identical reaction
mixtures, containing all ingredients except MgCl2, were incubated for 30 min at either 39 or 00C, and their residual transcriptase activities were assayed after the addition of
MgOl2
at310C.
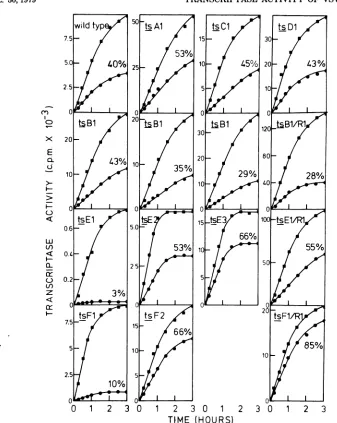
Figure 2 shows thatwild-type virus retained
approximately 40% of itstranscriptase activity
after heattreatment at390C. Similar heat
sta-bilitieswereobserved in thecaseofmutants ts
Al, tsC1, andtsDl (Fig.2).
Sincepreliminary experimentsindicated that
thetranscriptase activityoftsBl washeat
sta-ble, we assayed the heat stability oftwo
addi-tionalpreparations of thismutant. In all three
preparations, the heat stabilities of the
tran-scriptaseactivityweresimilarto oronly slightly
lessthan that ofwild-typevirusorthe revertant clone tsBl/Rl (Fig. 2).Thus, theinhibition of
RNAsynthesisat390C bythemutant tsB1was
probably the result of a reversible
configura-tionalchangeof the mutatedpolypeptide. Thetranscriptase activity of mutant ts El was very heat labile at390C. However, the revertant clone tsEl/Rl,aswell as the mutants ts E2 and
ts E3, had more stable transcriptase activities
thanwild-typevirus(Fig. 2). Thus, theinability
oftsEl tosynthesize RNA in vitro at
390C
isduetothermal inactivationofthetranscription process.
Mutant ts Fl retained only 10% ofits
tran-scriptaseactivity after heat treatment, showing
thatthe transcriptase activity of thismutant is
slightlyless heat labilethanthat oftsEl. The
on November 10, 2019 by guest
http://jvi.asm.org/
m
0 x
E
cr 0
u UC C)
w
L(
z
4r
ora±LLT r r I I I I.ll OXl l l l l I
0 1 2 3 0 1 2 3 0 1 2 3 0 1 2 3
[image:4.505.108.406.66.483.2]TIME
(HOURS)
FIG. 1. Assayofthe virion-associated RNA transcriptaseofthe ts mutantsofVSVNew Jerseyat 31and 39°C.Thevirion-associatedRNAtranscriptaseinpurifiedpreparationsofthets mutants wasassayedeither at31°C(@)or at39°C(0) asdescribed in the text.
transcriptase activity ofts F2 wasconsistently
more heatstable than that ofwild-type virus, and thetranscriptase activityoftherevertant ts
Fl/Rlwasthemostheatstable ofallthe clones
tested (Fig. 2). Thus, heat lability ofthe
tran-scriptase activityof the mutant tsFl probably
accountsfor itsinability tosynthesize RNA in vitroat390C.
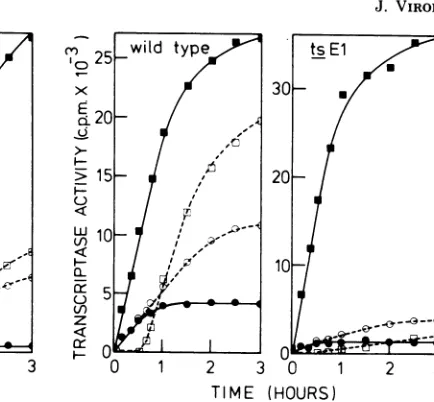
In vitro temperature-shift
experiments
withtsBi.
The mode of inhibition of the tran-scriptaseactivity of the mutant tsBi
was further studied by temperature-shift experiments, whereareaction mixture was incubated at390C for30minbefore transfer to 31°C (Fig. 3). Theamountof RNA
synthesized
after thetempera-ture was lowered to 31°C was
approximately
25% of that synthesized by a reaction mixturethatwas incubatedat310C throughout the
ex-periment. Another reaction mixture
containing
alltheingredients except
MgCO2
wasincubated for 30 min at 39°C, and the residual enzymeactivitywasassayedat31°C after the addition
ofMgCl2.Theamountsof RNAsynthesizedby
thetwopreparationswereverysimilar, showing
that enzyme activity was fully restored after
temperatureshift from39 to
310C.
These results suggestthatareversibleconfigurational change of the mutatedpolypeptideisresponsiblefor theon November 10, 2019 by guest
http://jvi.asm.org/
696 SZILAGYI AND PRINGLE
TABLE 1. RNAsynthesizedbyTNPpreparationsof the ts mutantsof VSV New Jerseya
Mutant % RNA synthesized at
Wild type ... 16
tsAl ... 26
tsCi ... 20
tsDl ... 12
tsB1 ... 4
ts BR/Ri ... 22
tsEl ... 3
tsE2 .... 8
tsE3 .... 15
ts El/Ri .... 19
tsFl... 3
tsF2 ... 12
tsFl/Rl .... 42
aUsing TNP preparations of thets mutantsof VSV
NewJersey,complementary RNAsweresynthesized at31and390Cunder conditions otherwiseidenticalto thosedescribed inFig. 1. Theamountsof RNA syn-thesizedat390Cafter 3 h of incubationareexpressed
asthe percentage of theamountsof RNAsynthesized
at 310C after thesame length ofincubation bythe sameTNPpreparation,asfollows: percent RNA syn-thesized at390C = (counts per minute after3 hat 39°C/counts per minuteafter3hat3100) x 100.
inhibition of the transcriptase activity of the mutant tsB1.
Heat sensitivity of the infectivity of the
tsmutants.Purifiedpreparationsof thets
mu-tantswereincubated for5hat
390C,
andtheir residualinfectivitywascompared with the infec-tivity of identical suspensions kept at 00C for the samelengthoftime.Inthe case ofwild-type New Jersey, around 5% of the infectivity sur-vived this heattreatment.Considering the vari-ationsinherent in suchexperiments,theresults indicate that the mutants and the revertant clones under investigation showed similar heat stabilities tothewild-typevirus. The only pos-sible exception is tsFl,where only 0.7% of theinfectivity wasrecovered, suggesting that this
mutantmay be heatlabile.
Since it hasbeenshownthat mutant ts El has
avery heat-labile transcriptase activity (Fig. 1 and2), it wassurprising that the proportion of infectious virussurviving the heat treatment at
390C
for 5 h was similar to that ofwild-type virus. For thisreasonwe retested the heatsta-bilityof theinfectivity of this mutant using two
furtherindependentlygrown and purified prep-arations (Table 2). The resultsconfirmthat the heat stability of the mutant ts El issimilarto that of the wild-type virus, since in both cases 10% oftheinfectivity was recovered after
treat-ment for5 h at
390C.
Webelieve that the low survival in the case oftsEl/Rl is erroneouslylow, since in another experiment the heat stabil-ity of this viruswas significantly higher (13.5%). Comparison of the heat lability of the transcriptase activity of disrupted and in-tact virions of mutant ta El. Two further experiments were conducted to determine how mutant ts El retains itsinfectivity after a rela-tively long incubation at 390C, even though the activity of its transcriptase is heat labile.
In thefirst experiment, a purified preparation of the mutant ts E1 was incubated for 5 h at
390C,and then itsresidual transcriptase activity
was assayed at310C. The results showed that, like wild-type virus, mutant ts El retained ap-proximately 8 to 10% of its RNA transcriptase activity. Therefore it appears that, when the undisrupted virions of ts El are heated, both the mutant's infectivity and transcriptase activity areasstableasthose ofwild-typevirus.
Next, the heat stability of the in vitro tran-scriptase activity of ts El was reexamined using aTNPpreparation(Fig. 4). There was very little 1 173RNA synthesis at 310C after an initial incubaRNA synthesis at 310C after an initial incubation of the reaction mixture at 390Cfor
30min.Similarly, almost all transcriptase activ-ity was lost when a control reaction mixture
containing alltheingredients exceptMgCl2was
incubated at 390C for 30 min and then the residual transcriptase activity was assayed at
310C.These results show thattranscriptase
ac-tivityofthe viralcore, unlike theactivityof the
undisrupted virion, is heat labile, presumably
duetothe irreversible denaturation of the
mu-tatedpolypeptide.
DISCUSSION
Inthis paperweattempttoidentifythe
num-ber of viral polypeptides involved in the
tran-scriptionofthe genome of VSV NewJersey by
comparing the in vitro activity of the
virion-associated RNA transcriptase of the tempera-ture-sensitive (ts )mutantsof all six complemen-tation groups. Similar experiments made with the ts mutants of VSV Indiana (20) and the temperature-dependenthostrange (td CE)
mu-tants (21)provided thefirstevidence thatsome mutants possess a temperature-sensitive
tran-scriptase.
The RNA transcriptase of the mutants of
complementationgroups C andDisunlikelyto
beaffectedbythe mutationsincemutants tsCl
and ts D1 synthesized RNA in vitro at 390C
similarlytowild-typevirus.Theseresultsagree
with thefinding that bothprimary transcription
andreplicationof the viralgenomeoccurincells
infected with either of these mutants (10, 16). Since the mutations didnotappeartoaffectany
J.
on November 10, 2019 by guest
http://jvi.asm.org/
Cr)0 ~~~~~00 0
30-X 20
-E
~~~~~3%1-20
8010
~
1029%
28%75L 7 S10 1.0
0 1 0 0
< tsEl
PsE
15otE3
100 ttsEl/R0 6 5.0
w
~~~~~~~~66%
(1)
~~~~~~~~~53%
10 50.4/.
H 50~~~~~~~
)0.2
25-o
0.
z 3%
H tsFl tsF2
20tsFl/R
7.5
~~~15-5- 10-
6510
10
2.5
5-10%
0 0 0
TIME (HOURS)
FIG. 2. Heatstabilityofthe virion-associated RNA transcriptase activity ofthets mutantsduring incu-bationat39°C for30min.Identical reaction mixturescontainingallingredientsexceptMgCl2wereincubated at39°C(0)and0°C(a)for30min,and their residualtranscriptase activitywasassayedat31°C afterthe additionofMgCl2asdescribed in thetext. Themutants werethesame asdescribed inFig.1exceptthattwo
furtherindependentlygrownandpurified preparations ofthemutanttsBlwereadded.
stageof RNAsynthesis in either of thesegroups,
it islikelythat thepolypeptides M and G of the
virus coat areaffected by these mutations. Wun-ner and Pringle found, paradoxically, that,
al-thoughinmutant tsDltheelectrophoretic
mo-bility of both polypeptides Gand Nwas altered, the tsphenotypeof thismutant appeared to be
due to mutation in the Npolypeptide because
reversion oftemperature sensitivity was
accom-panied by reversion of the N polypeptide to
nornal mobility(25).However, the gene
assign-mentof thismutantisunderreinvestigation by extensive biochemical andgenetic analysis.
In vitro RNAsynthesisat39°C bytsAl was even more pronounced than that by wild-type virus. This makes itveryunlikelythat
transcrip-tion in group A isaffectedbythemutation,and
suggests that some function in the RNA
repli-cation is defective. This is in good agreement with the results obtained with infected cells,
where replicationwas restricted whileprimary
transcriptionwasunaffected(10, 16).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.70.407.65.488.2]LU wild type tsBl
>- 80
50-H
~~~~25-z t 3,
0
0nube 1t3 r2 30 2 3
[image:7.505.63.251.62.260.2]TIME (HOURS)
FIG. 3. Reversibility of the activity ofthe RNA
transcriptase ofthemutant tsBJ. Thetranscriptase activity of purified preparations ofwild-type VSV NewJerseyand themutant tsBJwasassayedeither
at31°C(U)or390C(0).Afurtherreaction mixture was incubated at39tC for30mi and then
trans-ferred to 31'C (0). As control, a reaction mixture which contained all theingredientsexcept
MgCl2
wasincubated at390Cfor30min, and then
MgCl2
wasadded and the residual transcriptase activity was assayed at 31'C (El). The conditions ofassay are
described in thetext.
TABLE 2. Stability oftheinfectivity ofpurified group E mutantpreparationsduringincubation in
vitroat390Ca PFU/ml
% Sur-Mutants After
0°C
for After39°C
vivalb5h for5h
tsEl(2nd 6x 1010 6x 109 10
prepn)
tsEl(3rd 6.3x 10'0 6.3 x109 10 prepn)
tsE2 1.2x 10"1 1.7x10'0 14
tsE3 1.4x 1010 1.2 x109 9
tsEl/Ri 2.7x109 4.1x 107 1.5 a The purified virus suspensionswereincubated for 5h at39°C, and the residualinfectivitywastitratedat 31°C as described in the text. Two independently
grownand purifiedpreparations of themutant tsEl wereused.The preparations of themutantstsE2 and tsE3 andthe revertant tsEl/Rlwere thesame as
describedinFig.1.
'Percentsurvivalisexpressedas(PFU after39°C
for5h/PFU after0°Cfor5h)x 100.
Mutants of the other three RNA-negative complementation groups (B, E, and F) are the mostlikelytobetemperaturesensitive for
tran-scription,and the in vitroexperimentsindicated
0 1 2 3 0 1 2 3
TIME (HOURS)
FIG. 4. Reversibility oftheRNAtranscriptase ac-tivity ofthemutant tsEl.Wild-typeVSVNewJersey
andthemutanttsElwereused. Transcriptase
activ-itywasassayedat31°C(U),at39°C(0),andat31°C
after 30 min ofincubation at 39°C (0). Another reactionmixturecontainingalltheingredients except MgCl2 wasincubatedat39°Cfor30min, andthen
MgCl2 was added and the residual transcriptase activitywasassayedat31°C(O). Conditionsof
en-zyme assayaredescribed in thetext.
that therepresentativemutantsofthesegroups, mutant ts B1, ts E1, and ts F1, did possess
temperature-sensitive transcriptase activity.
Therefore, the investigation was extended to
includeallthreemutantsofgroupE andboth of groupF.
Therewasverylittle RNAsynthesisat39°C
inthecaseofmutanttsBi,whereasRNA
syn-thesiswasnotreducedin thecaseofthe revert-antclonetsBl/Rl,strongly indicating thatthe mutated gene product is involved in the
tran-scriptionprocess.Thisis ingoodagreementwith
the resultsobtainedwithtsBl-infectedcells(10,
16). Temperature-shift experiments withts Bi
showed that inhibition is a reversible process
presumablybrought about bya configurational
change of the mutated polypeptide. Thus, the
mode ofinhibitionin thismutantisverysimilar
tothat of thetemperature-dependenthostrange mutanttdCE3 (21,22).
Mutantsof complementationgroupEshowed differencesin the amounts ofRNAthey
synthe-sized invitroattherestrictivetemperature.The
involvementof the mutatedpolypeptideof tsEl in the transcriptionprocessis suggestedby the
fact that this mutant synthesized very little RNAat39°C,whereas its revertant clonetsEl/ Rl synthesized normal amounts of RNA at
39°C. On the otherhand, mutants tsE2 and ts
E3 bothsynthesized RNA at39°C. Since
repli-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.234.451.64.265.2]cationof viralRNAis observed at the restricted temperatureincellsinfected withtsE2(16), the mutated polypeptide in group E is almost
cer-tainly multifunctional. Thus, experiments
in-volving ts Elindicate that the polypeptide takes part intranscription, whereas those involving ts E3 suggestthat thepolypeptideisalso involved in replication, and thoseinvolving ts E2 indicate that thepolypeptide isdefectiveeither in some later stage ofreplicationorin somestageof virus
development following replication.
Alterna-tively,itispossible thatthetranscription
prod-uctssynthesizedat
390C
bytsE2and ts E3 arenotfunctional.
The mode of inhibition of the transcriptase
activity ofts El is irreversible thermal
inacti-vation of the mutated polypeptide. However, whenundisrupted virusparticles aresubjected
to heat, both the infectivity and transcriptase
activityof this mutant and ofwild-typevirus are
verysimilar.This maybe theresult of an
inter-action between the mutated polypeptide and
either the M or,lesslikely, the Gpolypeptide,
resulting in the suppression of the heatlability of the mutated polypeptide. Like interactions,
possibly involving somehost polypeptide, may
accountfor the primary transcription that takes place incellsinfectedwith tsEl (10).
The two group F mutants are alsodissimilar inthe amount ofRNA they synthesize in vitro
at
390C.
One of them, ts Fl, synthesizes verylittle RNA at
390C,
and the other, ts F2,syn-thesizes nornal amounts of RNA at this
tem-perature. Since the revertant clone, ts Fl/Rl,
synthesizesRNA at
390C,
weconclude that themutatedpolypeptideoftsFlis also involved in
the transcription process. This conclusion
ac-cords well with the finding that
primary
tran-scription in infected cells was inhibited at the restrictive temperature (10). The heat lability of the mutatedpolypeptide appears to be respon-sible for the inhibition of transcriptase activity. The fact that tsFl and ts F2 synthesizeddiffer-ent amountsof RNA in vitroat390C suggests that the polypeptide is
multifunctional,
being involved in both transcription and replication.However,it is alsopossible that the RNA species
synthesizedat390C bytsF2 are notfunctional.
Thus invitro experiments indicate that mu-tation affected transcription in group B, repli-cation in group A, and both transcription and
replication in groups E and F. The mutated
polypeptideingroupE may also be involved in
somelatestage of virus development. Itis hardly surprising that some of the polypeptides should be multifunctional, since there are only five of them, although some host polypeptide may also beinvolvedinsome stages of viral RNA
synthe-S1S.
Becausetherearesixcomplementationgroups
andonly fiveviralpolypeptides,it isdifficultto
make gene assignments. It ispossiblethat VSV codes for a sixth, so far unrecognized poly-peptide,althoughthepossibilitythat one of the six groups represents an extreme caseof intra-cistroniccomplementationhastobe considered
also.Sincepurifiedcoresisolated frommutants
tsBl, tsEl,andtsFlexhibit thesame temper-ature sensitivity as their virions, it is possible
thatthecorepolypeptides L, N,and NSarethe
sitesof themutationallesion in groupsB, E,and
F.
We propose that the mutated polypeptide in group B is polypeptide L, since the mode of inhibition in mutant ts B1 issimilartothat in td CE3, wherepolypeptideLwasshown to be the mutatedpolypeptide(21, 22). Thispossibilityis
strengthenedbya recentreport ofdissociation
and reconstitution experiments in which it was
foundthat the mutatedpolypeptideoftsBl was
either L orpossibly NS (9). If in group B the mutatedpolypeptide is L, then polypeptides N
and NSare the likelyones in groups E and F.
ACKNOWLEDGMENTS
We thank J. H.Subak-Sharpefor criticalreadingof the manuscript.We also thank C.Cunningham and P. Malloy for their excellent technical assistance, and J. T. Poyner for helping with the preparation of the manuscript.
LITERATURE CITED
1. Baltimore, D.,A. S.Huang,and M.Stampfer. 1970. Ribonucleic acidsynthesisof vesicular stomatitis virus. fl.AnRNApolymerasein the virion. Proc. Natl. Acad. Sci. U.S.A. 66:572-576.
2. Emerson, S.U., and R. R. Wagner. 1973. Lprotein requirement for in vitro RNA synthesis byvesicular stomatitis virus. J. Virol. 12:1325-1335.
3. Flamand, A. 1970. Etudegenetiquedu virusde la sto-matitev6siculaire:classement demutants thermosen-sibles spontanes en groupes de complementation. J. Gen.Virol. 8:187-195.
4.Flamand, A.,andC.R.Pringle.1971.Thehomologies
of spontaneousand inducedtemperature-sensitive mu-tantsofvesicular stomatitis virusisolatedinchick em-bryo and BHK-21 cells. J. Gen.Virol.11:81-85. 5. Hunt, D.M., S.U.Emerson,and R. R.Wagner.1976.
RNA-temperature-sensitivemutantsof vesicular sto-matitis virus:L-proteinthermosensitivityaccountsfor transcriptaserestriction of group Imutants.J. Virol. 18:596-603.
6. Hunt,D.M.,and R. R.Wagner.1974.Location of the transcription defect in group I temperature-sensitive mutantsof vesicular stomatitis virus. J. Virol. 13:28-35. 7. Knipe,D.M.,D.Baltimore, and H. F. Lodish. Matu-ration ofviralproteinsincellsinfected with tempera-ture-sensitive mutants of vesicular stomatitis virus. J. Virol. 21:1149-1158.
8. Knipe,D.M., H. F. Lodish, and D. Baltimore. 1977. Analysisof the defectsoftemperature-sensitive mu-tantsof vesicular stomatitis virus:intracellular degra-dation ofspecific viral proteins. J. Virol. 21:1140-1148. 9. Lesnaw,J.A.,and L. R. Dickson.1978.In vitro func-tional analysis of a temperature-sensitive mutant of vesicularstomatitisvirus, New Jersey serotype, defec-tive intranscription. Virology 91:51-59.
30,
on November 10, 2019 by guest
http://jvi.asm.org/
10. Lesnaw, J. A., and M. E. Reichmann. 1975. RNA synthesis by temperature-sensitive mutants of vesicular stomatitisvirus,NewJerseyserotype.Virology 63:492-504.
11. Lowry, 0. H., N. J.Rosebrough,A.L.Farr,and R. J. Randall. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193:265-275.
12. Preston, C. M., and J. F.Szilagyi.1977.Cell-free trans-lation of RNAsynthesized in vitro by atranscribing nucleoproteincomplex prepared from purified vesicular stomatitis virus. J. Virol. 21:1002-1009.
13. Pringle, C. R. 1970. Genetic characteristics of conditional lethalmutants of vesicular stomatitis virus induced by 5-fluorouracil,5-azacytidine,andethyl methane sulfo-nate. J. Virol. 5:559-567.
14. Pringle, C. R. 1978. The td CE and hr CEphenotypes: host range mutants of vesicular stomatitis virus in whichpolymerasefunction is affected.Cell 15:597-606. 15. Pringle, C. R., and I. B. Duncan. 1971. Preliminary physiological characterization of temperature-sensitive mutants of vesicular stomatitis virus. J.Virol. 8:56-61. 16. Pringle,C.R., I. B.Duncan,and M.Stevenson. 1971. Isolation and characterization of temperature-sensitive mutantsof vesicular stomatitisvirus,NewJersey sero-type. J. Virol. 8:836-841.
17. Printz, P., and R. R.Wagner.1971. Temperature-sen-sitive mutants of vesicular stomatitis virus:synthesisof virus-specific proteins. J. Virol. 7:651-662.
18. Repik, P., A. Flamand, and D. H. L. Bishop. 1976. Synthesis of RNA bymutantsofvesicular stomatitis
virus (Indianaserotype) and the ability ofwild-type VSV NewJersey tocomplement the VSV Indiana ts GI-114transcription defect. J. Virol. 20:157-169. 19. Rettenmier, C. W., R. Dumont, and D. Baltimore.
1975. Screening procedure for complementation-de-pendentmutantsofvesicular stomatitis virus. J. Virol. 15:41-49.
20. Szilagyi,J.F.,and C. R.Pringle. 1972. Effect of tem-perature-sensitive mutations on thevirion-associated RNAtranscriptaseofvesicular stomatitis virus. J. Mol. Biol. 71:281-291.
21. Sziliagyi,J.F.,and C. R.Pringle. 1975. Virion transcrip-taseactivity differences in host range mutants of vesic-ular stomatitis virus. J. Virol. 16:927-936.
22. Szilagyi,J.F.,C. R.Pringle,and T. M.Macpherson. 1977.Temperature-dependenthost rangemutation in vesicular stomatitis virus affecting polypeptide L. J. Virol. 22:381-388.
23. Szilagyi,J.F.,and L.Uryvayev. 1973. Isolation of an infectious ribonucleoprotein from vesicular stomatitis viruscontaining an active RNA transcriptase. J. Virol. 11:279-286.
24. Wunner,W.H.,andC. R. Pringle. 1972. Protein syn-thesis in BHK-21 cells infected withvesicular stomatitis virus. I. Tsmutants of the Indiana serotype. Virology 48:104-111.
25. Wunner, W.H., and C. R. Pringle. 1974. A temperature-sensitive mutant of vesicularstomatitis virus with two abnormal virion proteins. J. Gen. Virol. 23:97-106.
on November 10, 2019 by guest
http://jvi.asm.org/