JOURNALOFVIROLOGY, Sept. 1978,p.483489
0022-538X/78/0027-0483$02.00/0
Copyright X1978 AmericanSocietyforMicrobiology
Vol. 27, No.3
PrintedinU.S.A.
Bacteriophage SPOl Development:
Defects in
a
Gene
31
Mutant
ALBERTO N. SARACHU, MARIA C.ANON, ANDOSCARGRAU*
FacultaddeCiencias Exactas,UniversidadNacional de LaPlata,1900 LaPlata, Argentina
Received forpublication21December 1977
SPOl temperature-sensitive mutant ts14-1, located in cistron 31, has a DD
(DNAsynthesis-delayed) phenotypeat370C and producesprogenyin astretched
program. At 440C it behaves as a DO (DNA synthesis-defective) mutant and
shuts off the viral RNA synthesis about 10 min after infection. The thermal
sensitivity of thismutantis duetotheinactivity ofgp-31(the productof gene31)
at440C. However, gp-31 issynthesized at that temperatureandpartlyrecovers
itsactivityat370C. Only5min atthepermissivetemperatureisenoughtotrigger
thecontinuation of thephage program and to produce progeny. Thepartial defect
at37°Candthe expansion ofthemiddleprogramtogether with thepleiotropic
defectsatthe nonpermissive temperaturecould besuitableforthe study of the
controlsinvolvedinbacteriophage development.
SPO1 is a large bacteriophage that infects
Bacillus subtilis. Itstranscriptionprogram has
beendescribed indetail (9, 10). Alimitedsetof
controlsseems toregulate its genetic expression
(8). At5minafter infection the bacterial
speci-ficityof thetranscriptiveapparatusis alteredby
virus-coded positiveregulators of transcription
(1, 12). This action is mediatedbyaproteinof
28,000molecularweightcodedbygene 28(24,
6, 7). A similarsituation, though less clarified
due to the lack of appropriate mutants, was
found for the related bacteriophage SP82 (19).
About 10min after infectionat
370C
severalimportanteventsin the viraldevelopmental
se-quence takeplace. The synthesisoftwoclasses
ofviral RNA is shut
off,
athird class isamplified,andafourth class is turnedon. DNA synthesis
also starts at that time. Thisstage of the
pro-gram is less understood, but some knowledge
has been gained through the study of several
conditional lethalmutants. Nine cistrons have
beenfoundtobeessentialfor DNAreplication
(16), and twowereclassifiedasbeingmaturation
defective (8). Thesetwogenes, 33and 34, were
showntocode fortwopolypeptidesthat bindto
RNA polymerase (7), enabling thatenzyme to
performlatetranscription (5, 20).
In thispaperwepresentthe invivostudyof
apleiotropic temperature-sensitive (ts) mutant
in SPOl gene 31. Under nonpermissive
condi-tions,this mutant is defective in DNA synthesis,
and its transcription is shut off about 10 min afterinfection duetothelack ofatrigger func-tion mediated by this gene. This phenotype is also found in two other gene 31-suppressible mutants, susFl and susHA32.
MATERIALS AND METHODS
Bacteria andbacteriophage.Twostrains of B.
subtiliswere used, 168M and HA-101-B. The latter carries a strong suppressor (15, 18) and was kindly
supplied byS.Okubo.
Wild-type SPOl anditssuppressiblemutantswere the same as inreferences14and 16.
CHT medium(9)wasused for the growth of 168M, and L medium(16)wasused forHA-101-Bgrowth.
The bacteriawerealwaysinfected attheindicated
multiplicity ofinfectionwhen the optical densityat 500nmreached 0.5. Thedoublingtimefor B.subtiliu
168M was 30 to 32 min at either 37 or440C.
Experimental procedures. Labeled RNA was purified from cells thatwerequicklychilled, washed, lysed with lysozymeandsodium dodecylsulfate,and
hot-phenol extracted,asinFujitaetal.(8).
DNA was prepared from step gradient-purified SPOl phage byphenolextractionaccordingto
Man-dell andHershey(13).
DNA-RNA hybridizations were performed in 2x SSC(0.3MNaClplus0.03M sodiumcitrate)at700C for 3 h in a final volume of 200
pl.
DNase-free pan-creaticRNase was added to a final concentration of 12.5 pg/ml andincubated for 15 min at 37°C. The mixture was diluted in 0.01 M Tris-chloride (pH 7.5)-0.5M KCIandcollectedonnitrocellulose mem-brane filters. The filters were slowly washed three times with 10 mlof thesamebuffer, dried, and countedin aliquidscintillation counter with
Omnifluor-Tolu-ene(New England Nuclear).
RESULTS
Characterization of the SPOl mutant
tsl4-1. Mutant ts14-1 wasisolated from a
mu-tagenized growthofwild-typeSPO-1 in the
pres-ence of 10 yg of
N-methyl-N'-nitro-N-nitroso-guanidine per ml(the isolation wascarriedout 483
on November 10, 2019 by guest
http://jvi.asm.org/
484 SARACHU, AISIN, AND GRAU
in thelaboratory ofE.P.Geiduschekby I. Lahti
and 0. Grau (1). It is a temperature-sensitive
mutant because its burst sizes at 37 and 44°C
were about 30 and0.2 phages perbacteria,
re-spectively. Itwasbackcrossedtowild-type SPOl three times, giving reversionratesbetween 10-6
and 10-7,whichweretakenasanindication ofa
single mutation.
The DNA-synthesizing ability of tsl4-1-in-fectedB.subtilisatnonpermissivetemperature
was measured in comparison with that ofone
knownDO (DNAsynthesis-defective)mutantin
gene22andwild-type SPOl.Theincorporation of[3H]adenine intoalkali-stable, acid-precipita-blematerialwasnegligible comparedtothe wild
type (Fig. 1). The residual labeling observed for
tsl4-1was nothigherthan that of the DO
mu-tant susF30 (10). From this data it was
con-cluded that ts14-1 is aDO mutant and also is
abletoshutoff the host DNAsynthesis. To determine whether thismutant belonged tooneof thealready knownDO cistrons(16), it
wastestedbyspotcomplementation against
rep-resentatives ofallDO cistrons.Itcomplemented
susF2 (cistron 21); susF30 (cistron 22); susF20 and susO5 (cistron 23); susHA20 (cistron 27); susF21, susO36,susO45,and susO54(cistron 28); susF13 (cistron 29);susF26 (cistron 30); susF38
3
WI x
"I
I'_ 12
2
10 20
Time after infection (min)
FIG. 1. Incorporation of [3HJadenineinto DNAof wild-type-, susF30-, and ts14-1-infected B. subtilis 168Mat44°C. Culturesofwild type(0),susF30(0),
and tsl4-1 (V) SPOl-infected cells (multiplicity of infection, 10)weremadeupto0.5,uCiand 16pgof [3H]adenineperml2.5 min after infection. Atthe indicatedtimes, 0.5-ml samplesweretakenand hy-drolyzedwith 1 mlof1MKOHat45°C for3 h. Each
samplewasprecipitatedwith trichloroacetic acid(5% final concentration),collectedonnitrocellulose
mem-brane filters, washed with 5% trichloroacetic acid, dried,andcountedasdescribed in thetext.
(cistron 32). It did not complement susFl,
susF39, susHA32, and susO8 (cistron 31). In
accordance with these results, ts14-1 was
as-signedtocistron31.
tsl4-1-coded transcription. The ability of tsl4-1todirectthesynthesisof RNAduring its development in nonpermissive conditions was
tested bypulse-labeling B. subtilis-infectedcells
with[5,6-3H]uracilatdifferent times after
infec-tion. The labeled RNAs were purified and
hy-bridizedtoalargeexcessof SPOl DNA. These
data (Fig. 2) give us ameasurementof the
par-titionofRNAsynthesis between B. subtilis and phage DNA templates at different times after
infection (this is notcorrected for the
hybridi-z
0
4.
a.
4 ._
L._
:-
._-co
10 20
Time after infection(min) FIG. 2. Viral transcription level in SPOI
wild-type- and tsl4-1-infected B. subtilis 168M at 44°C.
RNAwaslabeledatspecified periods after infection
by2-minpulses of[3H]uracil (20yICi/ml and 0.26-pg/mlfinalconcentration)andimmediatelyisolated andpurifiedaccordingtothetext.From1.5 to2.5 pg
ofeach RNA per mlwashybridizedto50pgofSPOl
DNA per ml as in the text. Symbols: Wild type
(- - );tsl4-1 (-). Multiplicity of infection, 10
bacte-riophages per bacterium.
Backgroundwithout .
.o
. DNA Inputradioactivity['H]RNA
la- DNA (cpm) beling period (cpm)(min) Wild tsl4-1 Wild tsl4-1
type type
1-3 10 11 2,000 8,870
3-5 10 16 1,820 4,800
5-7 12 17 1,750 2,370
8-10 13 15 3,240 2,600
15-17 11 16 2,500 7,380
25-27 12 8,800
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.501.62.438.245.626.2]SPO1 GENE 31 485
zationefficiency of fully
complementary
RNA inthesameconditions, which is about50%). From
hereon, wewill refertothismeasurement asthe "level of transcription."
Atthebeginning oftheinfectionat440C,the
level of tsl4-1-coded transcriptionis somewhat
lower thanthat of thewild type,but itfallsto
less than 10% ofthewild type at 15 min after
infection and remains at that level thereafter.
Thisshutoffis common toothermutantsinthis
gene. Atnonpermissive conditions susHA32 has
levels oftranscription of12.5and2.8% between
5 to 7and 15 to 17 minafter infection,
respec-tively. Another representative of this gene,
susFl, alsohasaverylowlevel oftranscription
at 25to 27minafter infection(2.2%). This abrupt
shutoff of the phage RNA synthesis must
in-volve the limitation of the synthesis of three
RNAclasses:
m1L
m21,and1(9).Thelate level of transcriptionints14-1
infec-tionis proportional to thenumber ofinfecting
viral genomes. In Table 1 it is shown that a
variationin themultiplicity of infectionfrom4
to 16bacteriophagesperbacteriumgiveslevels
of viral
transcription
ranging from 0.8 to 4%.Eventhe highest valueismuchlower than the
30 to 40% usually obtained with the wild type
(multiplicityofinfection, 10).
In vivo
study
ofts14-1
mutation.Temper-ature-sensitive phage mutants can be used to
analyze: (i) thenatureof the mutation
itself;
(ii)the type offunction ofthealteredgene; (iii)the
timespaninwhich the functionisrequired for
the development of the phage (17).The
follow-ingexperimentsweredesignedtoelucidate these
characteristicsinmutantts14-1.
Phenotype
of tsl4-1 at 370C. The ts14-1burstsize at
370C
is aboutone-half that of the [image:3.501.259.450.360.519.2]wild-type SPOl.Inspiteofthisdifference,
370C
TABLE 1. Lateviral transcriptionlevel atseveral multiplicities of infection'
Multiplicity Input radio_ Background % Input
re-of infection Inu ai- without %Intre (phage/ activity DNA tainedon
fil-bacterium) (cpm)
(cpm)
ter4 13,125 20 0.8
4 7,400 7 0.9
8 6,080 6 2.5
10 5,154 3 2.5
16 4,075 6 4.0
16 1,993 5 3.7
aTwo-minute
pulse-labeled
(25 to 27 min afterin-fection)RNAswerepreparedfromtsl4-1-infectedB.
8ubtilis168M, usingthelabelingconditions inFig. 2. From1.5 to2.5 yg of each RNA per ml washybridized to10jg ofSPO1 DNA per ml(all other conditions andtreatmentasinMaterials and Methods).
wasinitially taken as the permissive
tempera-tureduetothe largevariation in phage
produc-tionamongindividual experiments.
Forabetterunderstanding of the shift exper-iments thatfollow,a moredetailed study ofthe
ts14-1 mutant phenotype at 370C was
per-formed.The kinetics ofDNAsynthesis of
ts14-1-infected B. subtili at 370C were measured.
The results (Fig. 3) indicatethat even at370C
this mutant behaves differently from the wild type. ItsDNAsynthesisstartsveryslowlyand
picks up speed about 20 min after infection,
whereas thewildtypestarts atabout 10 min and
keepsitsslopemore orlessconstantthereafter.
Second, the timecourse ofphage production
isalsodifferent(Fig. 3). The
intracellular
phage curves aresimilar,
but the one for tsl4-1 isdelayed for about 10 min, as it is the time of
appearanceofoneintracellularphageequivalent
andbacteriallysis.
Theseresultsindicatethat370Cis a
semiper-missive temperature for the tsl4-1 mutant.
Therefore,thefollowingexperimentsshould be
analyzedinaccordancewith theparticular time
scaleresulting from Fig.3.
Shis fromthepermissivetothe
nonper-missivetemperature.Inanattempt to
estab-0 0
'U
6,~~~~~~~~~~~~~~~~C<
0502
4006
[image:3.501.55.248.500.626.2]Time after infection(min)
FIG. 3. Phage production and DNA synthesis in SPOIwild-type-andtsl4-1-infectedB.subtilis 168M
at370C.Intracellularphages weremeasured at dif-ferenttimesafter infectionaccordingtoGeiduschek
andSklar(11).The resultsareexpressedas percent-ageofthe burst size. Inthreeexperimentsthese burst sizes were36, 82, and50bacteriophagesperinfected bacteriumforthewildtype(0) and60, 9, and24for
ts14-1 (0). The backgrounds (unadsorbed phages)
weresubtracted,theirmaximal valuesbeing0.8and 0.5%forwildtype andts14-1, respectively.
Incorpo-rationof/6HJadenineinto DNAwasdetermined as inFig. 1, except that
rHJadenine
(2.7 yCiand17-pg/mlfinal concentration) was added 6 min after infection and incubation was performed at 370C. Symbols: Wild type(A);ts14-1(A).
VOL. 27,1978
on November 10, 2019 by guest
http://jvi.asm.org/
486 SARACHU, AISON, AND GRAU
lish theperiodinwhich thefunctionof gene 31
is required, weperformedaseries ofexperiments
involving sudden changes from the permissive
tothenonpermissive temperature.
At different times after infection, B. subtilis
cells infected with ts14-1 at 37°C were
trans-ferred to440Corto amediumcontaining
chlor-amphenicolat370C. The yieldinboth cases was
measured60 minlater. The internalphages at
the time ofshiftwerealso determined (Fig. 4).
Phage production began to be higher than
thatofthe control (internal phages) whenthe
infected bacteriawereshiftedtothe
nonpermis-sive temperature after 20 min postinfection.
Whenthe shiftswereperformed later, the
pro-ductionincreasedconcomitantlytoaverage
val-uesup to 100times thatof thecontrol;thephage
yield obtained 60 min after the shift-up was
always higherthanthe internalphagespresent
atthe momentof shift. Theoppositewasfound
whencells, infected with either wild-type (data
100
0-IV
m
c
4,
4-n
._
mO
0
'I
5
40-C %NU
J. VIROL.
notshown) orts14-1 phage,weretransferredat
the time of shiftto medium containing
chlor-amphenicol; the amount of progeny was never
higher than the internalphages alreadyformed within thecells.
For comparison, several experiments with
wild-type SPOl were performed in the same
conditions.Theyshowedapartial sensitivityof
thewild-typephagetothe temperatureshift-up
from 0 to 25 min after infection. The average value resulted in 26% (standard error of the mean, 4) of the phage yield without shift-up
(average of13 experiments).When the transfer wasdonebetween 25 and35minafterinfection,
thevaluesincreased,although they showed large
variationbetweenindividual experiments.After
35minall the assays (5) gavemaximum yields
(thecellsarelysed), equaltothe ones obtained
when the cells were kept for 60 min at
370C
(value takenas 100%).
Shifts from the
nonpermissive
totheper-20
40 [image:4.501.116.405.301.594.2]Time
at
37'C(min)
FIG. 4. Shift-up experiments. B.subtilis 168Mwasinfectedwith ts14-1 at37°C (multiplicity of infection, 0.04). Samples ofthe culture werediluted at various times into bubblers at44°C, where incubation was
continuedfor60 min. At the timeof shift-up, othersampleswerediluted into medium containing chloram-phenicol (0.1 mg/ml). Internalphages present at time of shift were determined by lysis, according to Geiduschek andSklar(11). The results areexpressedaspercentageoftheyieldwhen theculturewasnot shiftedup(burstsize at37°C).Infour experimentsburst sizeswere60,24,5,and 9bacteriophagesperinfected
bacterium, respectively. Thehighest experimental background, given bytheextracellularphages,was0.5%of
the burstsize at37°C. Symbols: Shift-up (0); chloramphenicol shift (V);internalphages (0).
Q0
/
0
0
0~~~~~~~
000 0 0 v
0
- - 0-o_4-&--°--°80°f
v88A
#V-A a A A
on November 10, 2019 by guest
http://jvi.asm.org/
SPOl GENE 31 487
missive temperature. Atdifferent times after
infectiontsl4-1-infectedB. subtilis were
trans-ferred from 44 to 37°C, and the progeny was
determined 60 min later. The culture can be kept at440Cfor15 min withoutanyimpainnent of the phage yield (Fig. 5). However, more ex-tended periods caused an abrupt decrease in phage production: at 20 min it was less than 50% anddecayedrapidly thereafter. The wild-type-infected cells gave an average of 60 (standard errorofthe mean, 4)phages per bacterium in 25
experiments in which the shift was performed
between 0and 30 min after infection. They did notshow any trend towards a decrease in the
yield at the later times; for example, when
shifted down 30 min after infection, the yield
was 63phagesperbacterium.
Experimentsinvolvingacombination of
shifts.Ananalysisof theabove-mentioned
re-sults led to the conclusion that there was a
favorable window between min 15 and30 after infection to furtherstudythe nature ofthe tsl4-1mutation and theaction of the gene 31 product. The experiments performed involvedthe com-bination ofshift-ups and shift-downs with the addition and removal of chloramphenicol. To establish the feasibility ofthe experiments, it was necessary tomake surethat theextra
ma-*
100
S~~~~S
L I
U,,\
.o
~
~~~
0L
\
6) ~ ~ r-~
20 40 60
[image:5.501.51.246.376.558.2]Time at 44'C (min)
FIG. 5. Shift-down experiments. B. subtilis 168M
wasinfectedwith tsl4-1at44°C(multiplicity of infec-tion, 0.22). Sanprles ofthe culture were diluted at various times into bubblersat37°C,where incubation
wascontinuedfor60 min. Theresultsareexpressed aspercentages, in reference to theyieldwhen the culturewasshifteddown between 5and 15 minafter
infection.In twoexperiments,100%was30(standard
error, 2) and 69(standarderror, 4) bacteriophages
perinfectedbacterium, respectively. Thehighest
ex-perimental background was 0.8% (0.24
bacterio-phagesperinfected bacterium).
nipulations involvedwouldnotalter the behav-iorofthe system.
The positive control, that is, the growth of
tsl4-1at
370C,
was notaffectedbythedilutions and transfersrequiredby the experimentalde-signand gave 60phages per infected bacterium (Fig. 6,line 7). The experimental conditions were
adjustedtolower the backgroundgivenby the
unadsorbed phages; the value of 0.02 to 0.04 phages per infected bacterium obtained was sat-isfactory. Chloramphenicol added 15 min after infection gave a highlevel of inhibition, confirm-ing previousresults,and when it was diluted off at 30minafterinfection,about 10% of the phage
yield of the controlwasobtained(Fig. 6, lines8
and 9).
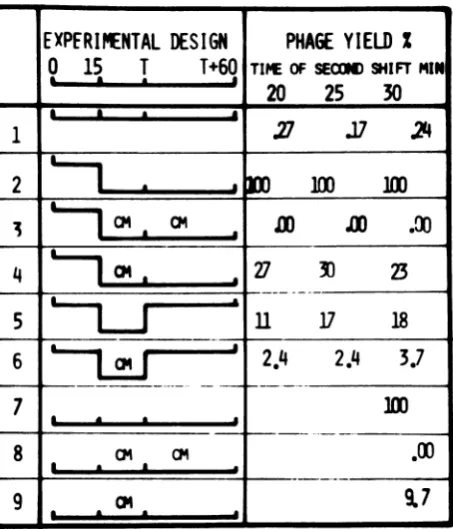
Fortheanalysisof theexperiments, theyield
of a shift-down at 15 min was taken as 100%
(Fig.6,line 2), and the unadsorbedphageswere
determined and subtracted in all cases before
the ratioswere calculated. The combination of shift-down and chloramphenicol gave the
ex-pected results. When chloramphenicol was
added atthe time of shift-down and remained for therestof theexperiment (line3), therewas
no phage production. When chloramphenicol
wasdiluted 5, 10,or15minafter theshift-down,
thephageproduction recovereduptoabout 25%
(line 4). The experiment shown in line 5 gave
about a 15% yield,
indicating
that 5 min wasenoughtosynthesize and/or permitthe action
of gp-31 to produce progeny. The
experiment
shown inline6gaveabouta2.5%yield,
which is10times higherthan the controlat 44°C (line
1). Thisproduction indicates that gene31 had been activebefore min15,producing gp-31,that
the product was
partly
reactivable at theper-missive temperature, and that5minofactivity
(atmost) wasenough toproducesomephages.
DISCUSSION
TheSPOl temperature-sensitivemutant
tsl4-1isarepresentativeof cistron31.In accordance
with thephenotypeof thiscistron,it isnotable
toreplicateits DNA(16).Atnonpermissive
con-ditions tsl4-1 as well as susHA32 and susFl
transcription is severely impaired 15 min after
infection. This effect is similarto the one ob-served insus mutantsF30, F4,and F14 (8) but
is more intense. In this case the low level of
transcription is proportional to the number of
infecting genomes. This isclearly not the only factorinvolvedinthe shutoff ofthesynthesis of
RNA, because even at large multiplicities of infectionthelevel of viraltranscriptiondoesnot
exceed 10% of that of the wild type.
Thetemporal parameters of the lytic cycle of the tsl4-1 mutant at
370C
are different fromVOL. 27,1978
on November 10, 2019 by guest
http://jvi.asm.org/
488 SARACHU, ANMSN, AND GRAU
EXPERIMENTAL
DESIGN
PHAGE YIELD S
°
15
TT+60
TIM OFSECN
SHIFT MIN_
20
25
30
1
.
,
.
W
.
27
17
.24
2
I DO
liO
100
_
cm cm.0o
4
27
0
2
5
II
17
18
6
2.4
2.4
3.7
7 100
8 * c c
.__
FIG. 6. Experimentscombining temperatureandchloramphenicol(CM) shifts. B. subtilis 168M wasinfected
withtsl4-1at440C (linesIto6;multiplicity ofinfection,0.39) or370C(lines 7 to 9;multiplicityofinfection,
0.33); 15min laterthecultures were diluted 10 times into thefollowingmedia: CHT at 44°C (line 1); CHT at
370C (lines2,5,and 7), andCHTat370Ccontaining 0.1 mg ofchloramphenicolperml (lines 3,4,6,8, and 9).
Five, 10, or 15min later(Tminutesafter infection),the cultures wereagaindiluted: 10 times into CHT at
44°C (line1);100times into CHTat37°C (lines2, 4, 7,and9);10times into CHT at37°C containing 0.1 mg
ofchloramphenicolperml(lines3and 8); and 100 times into CHT at44°C (lines5and 6). Totalphages were
determined60min later, lysingthe cells as described by Geiduschek andSklar(11).Resultsinlines 1 to 6
correspondtoexperimentsperformed simultaneously. Theyareexpressedaspercentageoftheyieldin line 2 (71 bacteriophagesperinfected bacterium). Results in lines 7 to 9correspondtoexperimentsperformedin parallel. Theyareexpressedaspercentageoftheyieldin line 7(60bacteriophagesperinfected bacterium). Theexperimental background (extracellularbacteriophages)was 0.04and0.02bacteriophagesperinfected bacteriumforlines 1 to 6 and 7 to9,respectively.
those ofthe wildtype. Theeclipse,the onset of
replication, and lysis take place about 10 min
laterthaninSPOlwild-type infection.
These results could be interpreted as ifthe
ts14-1 program at 370C were stretched out.
There isnoindication ofasubstantial alteration
in the known sequence of events leading to
phage production.
In theshift-up experiments the effect of the
gene 31function,in tsl4-1, beginstobe
notice-able atabout 20min afterinfection. After this
time it could be interpreted that gp-31 is
re-quired all alongthephage cycletoobtain
maxi-mumyield. However, this could not be
deter-mined definitelybecause the sensitivity ofthe
wild type to shifts-up up to 25 min could be
extendedtolongertimesbythetsl4-1 stretching of theprogram.
Itwassurprisingtofind thatts14-1 and
wild-typeSPOLare notassembledwithout
concomi-tantproteinsynthesis,asitwasobservedduring
thebacteriophageT4lyticcycle (17). The
pres-enceof
chloramphenicol
inthemedium reducesthe finalphage
yield
tothe amountofinternalphages present at the time of addition of the drug and sometimestovalues lower than those. This is not due to a direct effect of
chloram-phenicolupon thephage,becausematurephages
arenotaffectedby thedrugatthe concentration
used.
The cell machinery,
partially
controlled by the viralgenome,canspare itscapacitytopro-duce phage in the absence of gp-31 for some
time.
Forexample,afullamountofphageswasobtained when tsl4-1-infected B. subtiliswere
kept at thenonpermissivetemperature for upto 15 min and then shifted down to
370C.
After thattimethesystem isnotstable,
andat30min J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.144.371.79.344.2]SPOl GENE 31 489
it cannotrecoveritsabilitytoproduceprogeny.
This time interval, 15 to 30 min, was used in
studying thenatureof the ts14-1 mutation and thetime of action and synthesis of thegp-31.
The window experiments (Fig.6)indicatethat gp-31is inactivatedat440C butcanbe
synthe-sizedatthattemperatureand that its activity is atleast partly recovered when it is transferred to370C. Part of this synthesis takes place before
15min at440C, and its action between 15 and
20min is enoughtoproducesomephages. This
phage production takes place after thewindow
is closed(see Fig.4and line6ofFig. 6), indicat-ing that the action of this gene generates
pre-cursors orotherwise triggers intermediate
con-trols that will allow itssynthesis.
The data in Fig.6, although showingasmall
correlation between the length of the window and the phage yield, are not conclusive. How-ever, the amounts of phage DNA and RNA
synthesized are proportional to the time the window is open (Sarachu, submitted for
publi-cation).
The results of shifts and window experiments just mentioned, and the peculiar behavior of
ts14-1 at370C,suggestthatgp-31 issynthesized early in infection (before 15 min) but that its action isspecially delayed. This could arise from
an abnormally long processing time (if any is
involved) or fromthe existence ofa threshold
for its activity, which is attained after a long
time due to poor specific activity ofgp-31 or
some othermorecomplex (or indirect)
mecha-nism.
It istobenotedthatpartial damage ofgp-31
activity of tsl4-1 leads at 370C to aparticular
DD (DNA synthesis-delayed) phenotype (Fig. 3), which,incontrast tothe known DDmutants (16), is abletoproduceprogeny.
Thissituation, together with the already men-tionedstretchingof the program,could be
par-ticularlysuitable forstudyofthe multiple
con-trols involved in SPOl middledevelopment.
ACKNOWLEDGMENTS
Wewarmlythank E. P. Geiduschek and G. Favelukes for advice,support, andencouragement.
This researchwassupported by grantsfrom theConsejo NacionaldeInvestigacionesCientfficasyTecnicasde la Re-publica Argentina,Comisi6n deInvestigacionesCientificasde
laProvinciade BuenosAires,and Universidad Nacional de LaPlata.
A.N.Sarachu and M. C.Afi6nwerefellows ofthe Comi-si6n deInvestigacionesCientificas de la UniversidadNacional deLaPlata andConsejoNacional deInvestigaciones
Cienti-ficasy Tecnicas de la Republica Argentina. 0. Grau isa
recipientofaresearch careeraward from the Comisi6n de Investigaciones Cientificas de laProvincia de Buenos Aires.
LITERATURE CITD
1. Afi6n, M. C., and0. Grau. 1973.Transcripci6n en el crecimientoyla diferenciaci6n de SPO1, p. 109-112.In F. BarbieriandA. Legname (ed.), Progresos en biolo-gia.Tucuman, Argentina.
2. Duffy,J. J.,andE. P.Geiduwchek.1973. Transcription specificity of an RNA polymerase fraction from bacte-riophageSPO 1 infected Bacillus subtilis. FEBS Lett. 34:172-174.
3. Duffy,J.J., and E. P.Geiduschek.1975.RNA polym-erase fromphage SPO 1 infected and uninfected Bacil-lus subtilis. J.Biol.Chem. 250:4530-4541.
4. Duffy,J.J., R.L.Petrusek, and E. P.Geiduwchek. 1975. Conversion of Bacillus subtilis RNA polymerase activity in vitro by a protein induced by phage SPO 1. Proc. Natl. Acad.Sci. U.S.A.72:2366-2370.
5. Fox, T. D. 1976.Identificationofphage SPO 1 proteins codedby regulatorygenes 33and 34. Nature (London) 262:748-753.
6. Fox, T. D., R. Losick, andJ. Pero. 1976. Regulatory gene 28 ofbacteriophage SPO 1 codes for phage-induced subunit of RNApolymerase.J.Mol. Biol. 101:427433. 7. Fox,T.D., andJ. Pero. 1974.New phage SPO1 induced polypeptides associated with Bacillus subtilis RNA polymerase.Proc.Natl.Acad. Sci. U.S.A.71:2761-2765. 8.Fujita,D.J.,B. M.Ohlsson-Wilhelm,andE. P. Gei-duschek. 1971. Transcription during bacteriophage SPO 1development:mutationsaffecting theprogram of viraltranscription.J.Mol. Biol. 57:301-317. 9. Gage,LP., andE. P.Geiduschek.1971.RNAsynthesis
during bacteriophage SPO1development:sixclassesof SPO1RNA.J. Mol.Biol.57:279-300.
10. Gage,LP., andE. P.Geiduschek.1971.RNAsynthesis during bacteriphage SPO1development.II.Some mod-ulations andprerequisitesofthetranscriptionprogram. Virology44:200-210.
11. Geiduschek,E. P.,andJ. Sklar. 1969.Continual re-quirementfor a host RNApolymerasecomponent in a bacteriophage development. Nature (London) 221: 833-836.
12. Grau, O.,B.M.Ohlsson-Wilhelm,and E. P. Geidus-chek. 1970.Transcription specificityinbacteriophage SPO1development.ColdSpringHarborSymp. Quant. Biol.35:221-226.
13.Mandel,J.D.,and A. D.Hershey.1960.Afractionating column foranalysis ofnucleic acids. Anal. Biochem. 1:66-77.
14. Okubo, S.,M.Stodolsky, and B. Strauss.1964.The possiblerole ofrecombinationintheinfection of com-petent Bacillus subtilis by bacteriophage deoxyribo-nucleic acid.Virology24:552-562.
15. Okubo, S., and T. Yanagida.1968.Isolationof suppres-sor mutant in Bacillus subtilis. J. Bacteriol. 95:1187-1188.
16. Okubo, S.,T.Yanagida,D.LFujita,and B. M. Ohls-son-Wilhelm. 1972. The genetics ofbacteriophage SPO1.BikenJ.15:81-97.
17. Pulitzer,J. F. 1970.Functionof T 4gene 55. I. Charac-terization of temperature-sensitive mutations in the "maturation"gene 55. J.Mol. Biol. 49:473-488. 18. Shub,D. A. 1975.Nature of the suppressor of Bacillus
subtilis HA101B. J.Bacteriol. 122:788-790.
19.Spiegelnan,G.B.,andH. R.Whiteley. 1974. In vivo andin vitrotranscription byribonucleic acid polymer-asefrom SP82infected Bacillussubtilis.J. Biol. Chem. 249:1483-1489.
20. Tjian,R., andJ.Pero. 1976.Bacteriophage SPO 1 reg-ulatory proteins directing late gene transcription in vitro.Nature(London) 262:753-757.
VOL. 27,1978
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG.1.[3H]adeninefinalanddrolyzedsampleinfection,wild-type-,indicateddried,168Mbrane Incorporation of[3HJadenine into DNA of susF30-, and ts14-1-infected B](https://thumb-us.123doks.com/thumbv2/123dok_us/1525668.105142/2.501.62.438.245.626/fig-adeninefinalanddrolyzedsampleinfection-wild-indicateddried-mbrane-incorporation-hjadenine-infected.webp)