Copyright O 1975 AmericanSocietyforMicrobiology PrintedinU.S.A.
Characterization of T
Antigen
in Cells Infected
with
a
Temperature-Sensitive
Mutant
of Simian Virus
40
TAKAKO KUCHINO AND NOBUO YAMAGUCHI*
TheInstituteofMedicalScience,TheUniversity of Tokyo,.P.0.Takanawa, Tokyo, Japan 108 Received forpublication 2January 1975
T antigeninduced in Africangreenmonkeykidneycellsbya
temperature-sen-sitive mutant of simian virus 40, defective in a function required for cell transformation, was characterized. The number of T antigen-positive cells estimatedby animmunofluorescent techniques was almostequal atpermissive (32.5 C) andrestrictive (38.5 C) temperatures, butwasslightlyreducedwhenthe infected cells were incubated at a higher temperature (40.5 C). However, a
complementfixation test indicated that theamountofTantigeninducedby the
mutant is not significantly different from that induced by wild-type virus at
40.5 C. These results suggest that the Tantigen-inducing abilityof the mutantis not defective.Two distinct molecular species of Tantigen were induced by the
mutant at thepermissive temperature, whereas onlyone form wasobserved at
the restrictive temperature. Thelargermolecularform (14to15S)inducedby the
mutant atthepermissivetemperaturewasmoreheatlabilethanthatinducedby wild-type virus, suggestingthat the mutated geneproduct isacomponent ofthe larger molecular form.
T antigens of simian virus 40 (SV40) and
polyoma
virus are detectable notonly
in thenuclei
of tumors andtransformed
cells
initially
induced
by these viruses, but also
inacutely
infected
cells;
yetthey
are notstructural
pro-teins of the virions. Theviruses induceimmuno-logically distinct
Tantigens, and yet eachvirusinduces
antigenically identical
Tantigens
irre-spective of thespecies of cells used forinfection
and
transformation
(see
review21). The
induc-tion of the antigen after infectiondepends
onthe
expression
ofsomepart ofthe viral genome, since some temperature-sensitive (ts) mutants ofpolyoma
virusand
SV40 fail
toinduce
Tantigen
at restrictive temperature(15,
19, 20).The
sameconclusion
canbe drawn
from theobservations that the ability
ofpolyoma
virusorSV40
toinduce
T antigen is inactivatedby
gamma- or
UV-irradiation,
or bybeta-propi-olactone (2, 3, 5, 6, 30), and that a deletion mutant ofSV40, whose genome is
13%
shorter than that of the wild-type (WT) virus, fails to induce T antigen (31). However, none of these findings unravels whether T antigen is a virus-coded protein or a cellular protein specificallyderepressed
by the virus.The present study demonstrates that a mo-lecular form of T antigen induced by ts 900, a
transformation-defective
ts mutant of SV40 (29), is more heat labile than that induced byWT virus, indicating that the mutated gene
product may be T antigen or another compo-nentofthe molecular form of theantigen.
MATERIALS AND METHODS Virus. The WT SV40 was derived from the small-plaque 777 strain by cloning and a mutant, ts 900,was
derived from the WT virus mutagenized with UV irradiation(29).
Cell culture. The secondary cultures of African green monkey kidney (AGMK) cells were used to prepareT antigen. The GC7, a clone derived from a
AGMKcell, was used to prepare virus stocks and for plaque assay as described in the preceding paper (29). An SV40-transformed AGMK cell line, clone T22 which contains SV40 T antigen but not virion (V) antigen(24), wasprovidedbyK. Shiroki and usedto
prepareT antigen free from V antigen. The cells were cultivated in Eagle minimal essential medium sup-plemented with 10% calf serum.
Preparation of T antigen. T antigenwasprepared from confluent cultures of either T22 or virus-infected AGMK cells. Monolayer cultures of AGMK cellswereinfected withthevirus at amultiplicityof1 or10PFU/cell.After adsorption for 2 h at 37 C, and
then addition of maintainance medium (Eagle medium supplemented with 2% calf serum) with or without cytosine arabinoside (20
Ag/ml,
SigmaChem-ical Co.), the cultures were incubated at either 40.5Cfor 28hor at35Cfor 48 h. Themonolayercells were then washed twice with isotonic gelatin Vero-nal-NaCl buffer containing 0.15 mM Ca2+ and 0.5
1302
on November 10, 2019 by guest
http://jvi.asm.org/
mM Mg2+ (pH 7.5, ionic strength 0.147) (14),
scraped off witharubber policeman, andcentrifuged at 2,500 rpm for 10 min. The cell pellet was
sus-pended in the gelatin Veronal-NaCl buffer to give
about 20% (vol/vol) suspension, frozen and thawed
six times, and centrifugedat 2,500rpm for 10 min.
Thesupernatantfluidwasstored at-80C and used
asT antigen. T antigen from T22 cellswasextracted
similarly.
Complement fixation (CF) test. The
microcom-plement fixation tests for T and V antigens were
performed by the method of Sever (22) with minor modificationsaspreviously described (10). Briefly,one
drop ofsuitably diluted antiserum (2 CF units of T antibodyor4CFunitsof V antibody)andonedropof
guinea pigcomplementcontaining 5CH50unitswere
added inamicrotiter wellcontaining seriallydiluted
antigen. Themicroplatewasplacedovernightat4C. Onedrop of sheep erythrocytes sensitized with4units ofrabbithemolysin (2.5x101 cells/ml)wasadded and the plate was incubated at 37C for 1 h. After centrifugation, the amountofhemolysis wasgraded on a scale of one through five. The CF titers are
expressed as the reciprocal of the highest dilution
yieldingno more than 50%hemolysis. Diluent used wasthegelatin Veronal-NaCl buffer, pH7.5.
Antisera.Anti-Tsera wereobtained fromhamsters
bearingtumors induced by the SVH-93, a virus-free
clone ofSV40-induced hamster tumorcells. Each of the sera from individual animals was tested for
antibody against "T antigen" using 4 CF units of
solubleextractofWT virus-infected AGMK cells. To
testfor U antibody in thesera, the "T antigen"was
heated at 50C for 30 min and used as theantigen
inCFtestusing4CF units of theserum.Serawhich
demonstratedareduction of the "T antigen"titerby
more than 64-fold were considered to contain no
demonstrableamountofantibody against heat-stable Uantigen (12) andwereusedasspecific anti-Tsera.
None of theseracontained demonstrable V antibody
against4CF units ofpurified WT virions. The antisera against SV40Vantigenwerepreparedby inoculating rabbits with a purified "empty" band of WVT virus.
The antiserahad titers of 1:640or morewhen testedby
CF against 4 CF units of purified WT virions. A specific anti-V serum forCF test wasselected from
the antisera, which did not contain demonstrable
amountofTantibody against4CF units of T antigen from T22cells.
Immunofluorescence testsforT antigen.
Mono-layer cultureson coverslipswereinfected withserially
dilutedvirus, whichwasallowedtoadsorbat37 C for 2h. Thecellswerethen washed and incubatedatthe
indicated temperatures in medium (Eagle medium
supplementedwith 2% calfserum) containing cytosine arabinoside (15 pg/ml). The cultures were fixed
with cold acetone at 44 h (38.5 and 40.5 C) and 68h (32.5 and 35C) postinfection, and stained with
fluorescein-conjugated antibodies (23). The number of T antigen-positive nuclei were counted under a
fluorescentmicroscope.
Heat inactivationof Tantigen.Portionsof 0.2 ml oftheTantigenpreparationintesttubeswereheated
at 50 C ina waterbath for the times indicated, then cooled rapidly in an ice water bath, and tested for CF antigen titer.
Sucrosedensity gradientcentrifugation. Linear sucrose gradients were prepared with the aid of a mixing device from 5 and 20% (wt/wt) solutions of sucrose in 0.01 MTris-buffered saline (0.14 M NaCl, pH 7.4) containing 5 mM 2-mercaptoethanol (Wako Pure Chemical Co., Japan). The gradients were carefully overlaid with the T antigen preparations or with the marker SV40 DNA and centrifuged at 42,000 rpm at 4 C for 4 h in an SW50.1 rotor, or at 26,500 rpm at 4C for 24 h in an SW27 rotor. After centrifugation, fractions were collected from the bottom and each fraction was assayed for T antigen in CF test, or for radioactivity in a liquid scintillation spectrometer (Beckman LS-233) after dissolving it in triton X toluene scintillator.
Marker SV40 DNA. SV40 DNA was used as a
marker of knownsedimentationcoefficient, S,values. The WT virus was grown in the presence of 5MCiof
['Hlthymidine
per ml (25Ci/mmol)and waspurifiedby sedimentation onto a saturated KBr solution followed by two cycles of CsCl equilibrium density gradient centrifugationby the method of Uchida et al. (28). The virus DNAwas extracted from the band of complete virionsaccordingtoPagano (17).
RESULTS
T antigen in
infected cells.
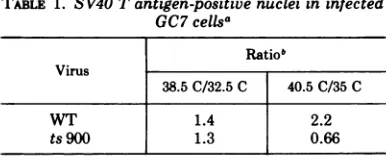
Table 1 shows that the number of Tantigen-positive nuclei in GC7 cells infected withWTvirus or ts 900 was almost equal at 38.5 and 32.5 C. Because the 38.5 C is the restrictivetemperature fortrans-formation or plaque formation by ts 900 (29), the results suggest that the mutation is not
expressed
atthestage of Tantigen
induction in thelytic cycle.However, the ratio of the number of Tantigen-positive
nuclei in ts 900-infected culture at 40.5 C to that at 35 C was abouttwo-thirds, whereas the ratio in WT-infected
[image:2.502.264.457.498.578.2]cultures was 2.2 (Table 1). In contrast to the
TABLE 1. SV40 Tantigen-positivenuclei ininfected
GC7 cellsa
Ratiob
Virus
38.5C/32.5 C 40.5C/35 C
WT 1.4 2.2
ts900 1.3 0.66
aSV40 T antigen-positive nuclei were scored by immunofluorescence at44h(38.5and 40.5C)andat
68 h (32.5 and 35C) postinfection as described in
Materials and Methods.
bThe number of T antigen-positive nuclei per culture was counted and theratio ofthe number of positive nuclei at the indicated temperatures was
calculated.
1303
on November 10, 2019 by guest
http://jvi.asm.org/
results obtained by the immunofluorescence test, no difference between the ratio of the amount of the T antigen induced at 40.5 C to that at 35
C
by WT virus and ts 900 wasdemonstrated
by
complement
fixation(Table
2).
Heat-inactivation of
Tantigen.
To investi-gate whetherthe
Tantigen induced
by
ts 900 wasidentical
in itsproperties
to that inducedby
WT virus, aheat-inactivation
testforthe T antigen wasperformed (Fig. 1).
The Tantigeninduced by
ts 900 at 35 C was inactivated much morerapidly
at 50C than
that inducedby
WT virus.The slopes
ofthe inactivation
curvesboth ofWT T
antigenand
of ts 900 were steep at thebeginning
of the inactivation. The rate of inacti-vation ofthe WT
Tantigen
slowed down after
5 min ofincubation
at 50C, whereas that
ofthe
T antigeninduced
by
ts 900proceeded
almostlinearly.
Theresults
suggest the presence of relativelyheat-stable
fraction inaddition
toheat-labile
fraction inthe
WT Tantigen prepa-ration,and the lack
ofthe
relatively heat-stable
fraction in the T
antigen
inducedby
ts900. The titer of V antigen inthe
T antigen preparation frommutant-infected cells
was notaffected by
the
heating
at 50C
for 30 min.Heat-inactivation of
Tantigen
fraction-ated
by
sucrosegradient
centrifugation.
The
twomain
molecular
weight classes
ofTantigen
reported previously (18)
have been observed
repeatedly when the
Tantigen induced
by WT
virus or ts 900 at 35
C
wascentrifuged
through
sucrose
gradient
andassayed by CF (Fig.
2,peaks
1and
2).However, the
larger molecular
weight class,
peak 2,which sediments with
anS
value
of 14 to 15S, was notdetected
inthe T
antigen extracted
fromcells infected
with
ts900TABLE 2. AmountsofSV40Tantigenininfected
AGMKcellsa
CF unitsc Ratio
Virus MOI' (40.5C/35C)
40.5C 35C (4.C/5)
WT 1 32 90.5 0.35
10 128 362 0.35
ts900 1 32 90.5 0.35
10 64 181 0.35
aInfected
cellswereharvestedafter 28 h of incuba-tionat 40.5Cand after 48 h at 35 C in the presence of cytosine arabinoside (20gg/ml).
The amount of T antigen in the cells was determined in CF test asdescribedin Materials and Methods.
bMOI, Multiplicityofinfection; PFU per cell.
cThe CF titers of the 20% cell extracts are the averages ofduplicate titrations
1/16
-1/64
1/256
5 10 20 30
INCUBATION TIME (min) FIG. 1. Heat inactivation of SV40 T antigen in-duced by WT virus and ts 900 at 35 C. Tantigen was extracted from AGMK cellsinfected with WT virus (0) or ts 900 (0) and incubated for 48h at 35C. Portions of 0.2 ml of the extract in test tubes were heated at 50 C for the times indicated and then cooled rapidly at 0C. Survival was assayed by CF test.
at 40.5
C (Fig. 2B). The
Tantigen species
ofpeaks
1 and 2 in extracts ofcells infected
withWT
virus or ts 900 at 35C
weretested for
heatlability
at 50C (Fig. 3).
Nodifference
inheat
stability
wasnoted
between the smaller
forms(peak
1) of Tantigen induced
by
WT virus or ts 900.However,
inthe
case ofthe
peak
2, T antigenobtained
fromcells
infectedwith
ts 900 wasconsiderably
more heatlabile than
thatobtained
from WT-infected cells.DISCUSSION
The present study was undertaken to identify
the
mutated gene product of an "early"SV40
mutant, ts 900. The results indicated that the antigen extracted from AGMK cells infected with ts 900 was more heat labile than that extracted fromthe cells infected with WT virus, as assayed by
CF
test using hamster antisera.SV40
can induce V, capsid (C), T, U, and surface antigens in infected AGMK cells (16,1304
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.502.257.444.62.360.2] [image:3.502.53.244.472.584.2]16- A
P2 P1
8
-4
2
~
5 10 15 20 25
UQ16- B 21S 16S
8
4
5 10 15 20 25
FRACTION NUMBER
FIG. 2. Ratezonal centrifugation ofSV40T anti-geninduced by WT virus and ts900. Tantigen was
extractedfromAGMK cells infected with WT virus (A) and ts 900(B) and incubatedfor28h at 40.5C (A) or for 48 h at 35C (0). The 4.5-mi sucrose gradients(5to
20%o),
each with 0.2 mloftheextract or markerSV40DNA, were centrifuged at42,000 rpmfor 4 h at 4C in an SW50.1 rotor. Fractions were
collectedfrom the bottom andassayed forTantigen orforradioactivity. Thepeaksoftheradioactivity of
the marker DNAareshown bythearrows.
21). The antisera
used in the
presentstudy
wereobtained
fromtumor-bearing
hamsters, which
had
been
transplanted with virus-free
tumorcells. The
seradid
notcontain
detectable
amount
of
antibody
against
Vantigen.
Anti-body
against
C
antigen
was notexamined. But
C
antibody
is
reported
tobe produced
in
ani-mals immunized with
disrupted
SV40
virions(16),
and
itis
unlikely,
therefore,
that the
seraused
inthis
study
contained
C
antibody.
The
presence
of
antibody
against
U antigen
wasexamined
by
using
asantigen in CF
teststhe
extract
of
virus-infected AGMK cells which had
been
heated
at 50C for
30 min.In
extractof
SV40-infected AGMK
cells,
U antigen can bedemonstrated
by
aT+U+ hamster
serumand
the
titer ofU
antigen
inthe
extract isusually
about one-fourth the
titer ofT
antigen
(12).
The
sera
used
inthis
study
werethose which
demon-strated the reduction
ofatleast 64-fold
inCF
antigen
titer afterthe
extracthad been heated.
Therefore,
it isunlikely
that the antiseracon-tained
antibody
against heat-stable U
antigen.
This evidence
mayprovide further
proof that theantisera used did
not containantibody
against heat-stable V and C antigens (1,
16).Antibody
against surface antigens is present inhamsters which have
rejected transplantation oftransformed
cells, or havebeen
immunized withSV40
virus,as well as in pregnant animals,but it
isabsent from
tumor-bearing animals (8,
26, 27).
Because
our antisera wereobtained
from
tumor-bearing hamsters, the
surface anti-gens areunlikely
tobe involved
in the CF reaction inthe
presentstudy. Based
onthese
observations,
we areconvinced that the
extracts ofSV40-infected
AGMK cells reacted with
antibody against T antigen alone.
Sedimentation
velocity
determinations
showed that the
extract fromAGMK cells
infected with WT
virus or ts 900 atthe
permis-sive temperature contain twomajor molecular
forms of
T
antigen(Fig.
2).This
result agreeswell with
that
reported by Potter
etal.
(18).However, the
extract fromAGMK cells infected
with
ts 900 atthe
restrictive temperaturedid
not contain
the
larger molecular
form ofT
antigen which sediments with
anS value
of 14 to15S
(Fig. 2B). Osborn and Weber have
reported similar result
using atsA
mutant ofSV40 (M. Osborn and K. Weber, Abstr.
ColdSpring
Harbor Tumor Virus Meet.
1973, p. 21).16
U
a"
<21
5 10 20
INCUBATION TIME min'
FIG. 3. Heat inactivation ofSV40 Tantigen
frac-tionatedbysucrosegradient centrifugation.Tantigen
was extracted from AGMK cells infected with WT
virusandts 900 at 35Casdescribed in thelegendof
Fig.2.The36-migradient (5to
20%),
eachwith1 mlof theextracts ormarkerSV40DNA,werecentrifuged at 26,500 rpm for 24 h at 4C in an SW27 rotor.
Fractions, 1mleach,werecollectedfrom the bottom and assayed for T antigen or for radioactivity. A
fractionfrom eachof peak1 (0,WTvirus;*,ts-900) andpeak2(A, WTvirus; A, ts-900) (seeFig. 2)was
heat inactivatedasdescribed in thelegend ofFig.1.
1305
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.502.58.247.51.317.2] [image:4.502.264.453.383.552.2]The 14to 15S form correspondstoanestimated molecular weight of 340,000 to 390,000 (13). Since SV40 DNA cannot code for a protein of molecular weight larger than 200,000 (21), this species of antigen may be an antigen complex which is composed ofanoligomer of the smaller molecular form, or an aggregate with another macromolecule(s). Recently Carrol et al. (4) showed the dissociation of larger molecular species of T antigen to the smallest molecular weight form in high salt concentrations. The 14
to 15S T antigen complex induced by ts 900 was more heat labile than that by WT virus (Fig. 3). Theresults suggestthe tsgene product is a component of the 14 to 15S T antigen complex and maybe best explainedasfollows: the mutated gene product affects the ability of the componentsofthe antigencomplextomake the more heat-stable, larger molecular form, thus the antigen complex induced by themutant at the permissive temperature is easily con-verted by heating to the more heat-labile, smaller form. In infected cells incubatedat the restrictive temperature, the ability is com-pletely destroyed and the 14 to 15S antigen complex cannotbe made. Theresults, however,
do notanswerthe question whether the tsgene
product is a proteinwhich determines the anti-genicity of T antigen oranother protein which may exist in the antigen complex and interact with the antigen-determining protein. Further characterization of individualcomponentsofthe T antigen complex will answerthis question.
The function ofTantigenseemstobeclosely correlated with its molecular forms, but not with its antigenicity because total amount of the T antigen induced by the mutant at the restrictive temperature was not significantly different from that induced at the permissive
temperature (Table 2), whereasthe 14to15ST
antigen complex wasnotformedat the restric-tive temperature. The defective function of ts 900 is not known. The function is required for
transformation of nonpermissive cells and for
viral replication in permissive cells (29). If ts
900 is a double-step mutant, the gene which affects the heatstabilityofthe T antigen could beadifferentgene fromthat which affectsboth for thetransformation andfortheviral replica-tion. But this is unlikely because T antigen induced by ts 904 and 907, which are grouped
intothesamecomplementation groupasts900
(29), was also more heat labile than that in-duced by WTvirus (datanotshown). Thegene product oftsA ofSV40, to which group ts 900
may also belong (29), is suggested to be an initiator molecule for viral DNA replication
itself or an essential component of the initiator (25). But the role of the gene
product
intransformation
has notyet been studiedexten-sively.
Atpresent, attempts to purify T antigen have not been very
successful
(7, 9, 11, 18).Since
the function of the T antigen inducedby
ts 900 islikely
tobe
temperaturesensitive,
it mayserveas a
good
materialtopurify
and toidentify
the function of T antigen.ACKNOWLEDGMENTS
We aregratefultoHirotoShimojo and Kinichiro Odafor their helpful discussions andcritical review ofthis manu-script. We would liketothank KazukoShirokiforsupplying usT22cells.
The work was partially supported by a grant from the MinistryofEducationofJapan.
LITERATURE CITED
1. Black, P. H., W. P. Rowe, H. C. Turner, and R. J. Huebner. 1963. Aspecific complement-fixing antigen present in SV40 tumor andtransformed cells. Proc. Natl. Acad. Sci. U.S.A. 50:1148-1156.
2. Brown, A., R. A. Consigli, J. Zabielski, and R. Weil. 1974.Effect of,B-propiolactone inactivation of polyoma virus onviral functions. J. Virol. 14:840-845.
3. Carp, R. 1., and R.V.Gilden. 1965.The inactivationof simian virus 40infectivity andantigen-inducing capac-ityby ultravioletlight. Virology 27:639-641. 4. Carroll, R. B.,L.Hager,andR. Dulbecco. 1974.Simian
virus40 Tantigen bindstoDNA.Proc.Natl. Acad.Sci. U.S.A. 71:3754-3757.
5. Coppey, J., and R. Wicker. 1968. Inactivation par les radiations U.V., X etgamma de deuxproprietes du virus SV-40: infectivete et induction de lantigene nucleaireprecoce.Ann.Inst.Pasteur 115:478-485. 6. Defendi, V., F.Jensen, and G. Sauer. 1967. Analysisof
someviral functions related toneoplastic transforma-tion, p. 645-662. In S. J. Colter and W.Paranchych, (ed.), The molecularbiologyof viruses.Academic Press Inc., New York.
7. DelVillano, B. C., and V. Defendi. 1973. Characteriza-tion oftheSV40 T antigen. Virology 51:34-46. 8. Duff, R., and F. Rapp. 1970. Reaction of serum from
pregnant hamsters with surfaceofcellstransformedby SV40.J.Immunol. 105:521-523.
9. Henderson, I. C., and D. M. Livingston. 1974. Partial purification and characterization ofthe SV40 T anti-gen. Cell 3:65-70.
10. Hino, S., N. Yamaguchi, and T. Yamamoto. 1972. Characteristics of a virusfoundin aRousmouse cell line,SR-C3H-2127. Jpn. J. Exp. Med. 42:43-52. 11. Kit,S., J. L. Melnick, M. Anken, D. R. Dubbs, R. A. de
Torres, and T. Kitahara. 1967. Nonidentity of some simian virus40-inducedenzymes with tumor antigen. J.Virol. 1:684-692.
12. Lewis, A. M., Jr., and W. P. Rowe. 1971. Studies on nondefective adenovirus-simian virus 40 hybrid vi-ruses.I.Anewlycharacterized simian virus 40 antigen inducedbytheAd2+ND,virus. J. Virol. 7:189-197. 13. Martin, R. G., and B. N. Ames. 1961. A method for
determining the sedimentation behavior of enzymes: Application to protein mixtures. J. Biol. Chem. 236:1372-1379.
14. Mayer, M. M. 1961. Complement and complement fixa-tion, p. 133-240. InKabatand Mayer'sexperimental
on November 10, 2019 by guest
http://jvi.asm.org/
immunochemistry. Charles C Thomas, Publisher, Springfield,Ill.
15. Oxman, M. N., K. K. Takemoto, and W. Eckhart. 1972. PolyomaT antigen synthesis bytemperature-sensitive
mutantsofpolyomavirus. Virology49:675-682.
16. Ozer, H. L., and P. Tegtmeyer. 1972. Synthesis and assemblyofsimian virus40.II.Synthesisofthemajor capsid protein anditsincorporationintoviral particles. J. Virol. 9:52-60.
17. Pagano, J.S.1969.Assay of infectious DNA, p.184-197. InK. Habel andN.P.Salzman, (ed.), Fundamental techniquesinvirology. AcademicPressInc.,NewYork. 18. Potter, C.W.,B. C. Mclaughlin, and J. S. Oxford. 1969. Simian virus 40-induced T and tumor antigens. J. Virol.4:574-579.
19. Robb,J. A.1973.Simian virus 40-host cell interactions.I. Temperature-sensitive regulation ofSV40 Tantigen in 3T3mouse cells transformed bythe ts* 101 tempera-ture-sensitive early mutant of SV40. J. Virol. 12:1187-1190.
20. Robb,J.A.,P.Tegtmeyer,A.Ishikawa,and H. L.Ozer. 1974. Antigenic phenotypes and complementation
groups of temperature-sensitive mutants of simian virus 40.J.Virol. 13:662-665.
21. Sambrook,J. 1972.Transformationby polyomavirusand simianvirus 40. Advan.Cancer Res. 16:141-180. 22. Sever,J. L. 1961.Applicationofamicrotechniquetoviral
serological investigations.J.Immunol.88:320-329. 23. Shimojo, H.,H.Yamamoto,and C. Abe.1967.
Differenti-ationofadenovirus12antigens in cultured cellswith immunofluorescent analysis. Virology31:748-752. 24. Shiroki, K., and H. Shimojo. 1971. Transformation of
greenmonkey kidney cells bySV40genome:the
estab-lishmentoftransformed cell lines and the replicationof human adenoviruses and SV40 in transformed cells. Virology45:163-171.
25. Tegtmeyer, P. 1972. Simian virus 40 deoxyribonucleic acidsynthesis: the viral replicon. J. Virol.10:591-598. 26. Tevethia, S. S., L. A. Couvillion, and F. Rapp. 1968.
Developmentinhamstersofantibodiesagainstsurface antigens present in cellstransformed by papovavirus SV40. J.Immunol.100:358-362.
27. Tevethia,S.S.,M.Katz,and F.Rapp.1965. New surface antigen in cells transformed by simian papovavirus SV40.Proc.Soc.Exp.Biol.Med. 119:896-901. 28. Uchida, S., K. Yoshiike, S. Watanabe, and A.Furuno.
1968.Antigen-forming defective viruses ofsimian virus 40.Virology34:1-8.
29. Yamaguchi, N., andT.Kuchino. 1975. Temperature-sen-sitivemutantsof simian virus 40selected by transform-ingability. J. Virol. 15:000-000.
30. Yamamoto, H., and H.Shimojo.1971. Inactivationof T antigen-forming capacities of simian virus 40 and adenovirus 12 by ultraviolet irradiation. J. Virol. 7:419-425.
31. Yoshiike, K.,A.Furuno, andK. Suzuki. 1972. Denatura-tion mapsofcomplete and defective simian virus 40 DNA molecules. J. Mol. Biol. 70:415-423.
VOL.15,1975