0022-538X/78/0025-0788$02.00/0
Copyright () 1978 AmericanSocietyforMicrobiology Printed in U.S.A.
Marker Rescue of Endogenous
Cellular Genetic
Information
Related
to
the Avian
Leukosis Virus Gene
Encoding
RNA-Directed DNA
Polymerase
GEOFFREY M. COOPER
SidneyFarberCancer Institute andDepartment of Pathology,Harvard MedicalSchool,Boston, Massachusetts 02115
Received for publication 7 November 1977
Endogenous cellular genetic information related to the avian leukosis virus
gene encoding RNA-directed DNA polymerase was studied, using a marker
rescueassay todetectbiological activityofsubgenomicfragmentsof virus-related DNAsofuninfected avian celLs.Recipientcultures of chickenembryofibroblasts weretreated with sonicated DNAfragments andwereinfected witha tempera-ture-sensitive mutant ofRoussarcomavirus that encodedathermolabile DNA
polymerase. Wild-type progeny viruses were isolated by marker rescue with
fragments of DNA of uninfected chicken, pheasant,quail, andturkeycells. The DNAs of theseuninfected aviancells,therefore, appearedtocontainendogenous
geneticinformation relatedtothe avianleukosis virus DNApolymerasegene.
Aviancellsappear to inheritendogenous ge-netic information related to three groups of avian retroviruses. The DNAs of uninfected chicken cells contain nucleotidesequences that
arehomologousto mostofthe RNAsequences
of Rous-associatedvirus0(RAV-0), an endoge-nousavianleukosisvirus (ALV) spontaneously produced bysomeinbred lines of chickens (20, 25, 26, 31, 37). Nucleotide sequences that are related to ALV RNA have also been detected in the DNAs of other avian species, including pheasant, quail, andturkey (20, 25, 31, 35). In
addition, avian DNAs contain nucleotide
se-quencesrelatedto somesequencesof the RNA of avianreticuloendotheliosis viruses,asecond group ofavian retroviruses which lack nucleic
acid homology with viruses ofthe ALV group
(20).Endogenous geneticinformation relatedto athirdgroupof avian retroviruses has also been identified in some species of Chrysolophus
pheasants (3, 9, 16).
Inmostcases, endogenous virus-related DNA sequences are not expressedtoyield infectious virus.Althoughapparently all chickenscontain similar amounts of RAV-0-related DNA se-quences (25, 26), onlyafew inbred linesofV+ chickensproduce RAV-0 (7,8, 29, 30, 37). Partial expression of the ALV-related DNAs of non-virus-producing (V-) chicken cells has been demonstratedbyimmunologicalassays of ALV-relatedproteins (2, 12,28) andby the abilityof someV- chickencellstocomplementreplication of Rous sarcoma virus (RSV) strainsdefective inenvelope glycoprotein synthesis(14, 39).
Sim-ilarly, somepheasant cellsproduce proteins re-latedtoALVinternal structuralproteins
(group-specific antigens) (9, 15) and some pheasant,
partridge, and Chinesequailcellsproduceviral
envelope glycoproteins orinfectiousvirus(3,9,
15, 16). The biological activity of endogenous genetic information related to ALV envelope glycoproteins has also been demonstrated by
recombination betweenexogenousALVand
en-dogenousenvelope glycoproteingenesofchicken
(17, 18, 31, 41) andpheasant (9, 15, 31, 32, 34)
cells.
Assays of DNA infectivity by transfection
were previously used to studythe endogenous
ALV-related genes ofuninfected chicken cells (6).In contrasttotheinfectivityof RAV-0 pro-viral DNAs of cells which were exogenously
infected with RAV-0, the endogenous RAV-0-related DNAs of uninfected V- andV+chicken celLs lacked detectableinfectivityintransfection assays (6). Therefore the endogenous RAV-0-related DNAs of uninfected chicken cells ap-pearedtodiffer fromtheRAV-0proviralDNAs of RAV-0-infected cells. Since uninfected chickencells did notcontaininfectious RAV-0 genomes, it was of interest to investigate the
biological activity ofsubgenomic fragments of
endogenous ALV-related DNAs.
We recently described a biological assay of
subgenomicfragments ofALV DNAbymarker rescue(4).Intheseexperiments, wild-type prog-enyRSVwereisolated aftertreatmentof recip-ient cells with subgenomic fragments ofALV DNA and infectionwithtemperature-sensitive 788
on November 10, 2019 by guest
http://jvi.asm.org/
RESCUE OF ENDOGENOUS ALV DNA POLYMERASE
mutants of RSV. We have now assayed DNAs of uninfectedavian cells forgenetic information relatedtotheALV gene forRNA-directed DNA polymerase (pol) by the marker-rescue tech-nique.Wild-typeprogenyRSVwereisolated by marker rescue ofpol' RSV withfragments of DNAofuninfectedchicken, pheasant, quail, and turkeycells.These results indicatedthat DNAs of these uninfected aviancells contained endog-enous genetic information related to the ALV polgene.
MATERIALS AND METHODS
Cells and viruses. Cells were grown in
Temin-modifiedEagleminimal essential medium containing
20% tryptosephosphatebroth(ETmedium) and
sup-plemented with calf and fetal bovine sera. Fertile
chicken eggswerepurchasedfromSPAFAS, Norwich,
Conn., and fromHyline,DaUasCenter,Iowa. Fertile
line 100 chicken eggs were a generous gift of L. B.
Crittenden,U.S.DepartmentofAgriculture,East
Lan-sing, Mich. FertileOrlopp turkeyeggswereprovided
by the Wilmar Poultry Co., Wilmar, Minn. Fertile
ringneck pheasanteggswere akindgiftof M.Smith, Apopka,Fla.FertileJapanesequaileggswereobtained fromLife Sciences, Inc., St. Petersburg, Fla. Fertile
Pekin duckeggs wereobtained fromalocal farmer.
MouseNIH 3T3cellswere akindgiftofR. A.
Wein-berg, Massachusetts Institute ofTechnology,
Cam-bridge.
SPAFAS chickenembryofibroblastswereC/E, V-,
chickenhelperfactor(chf) negative,and ALV
group-specific(gs)antigen negative (assayed by complement
fixation). HylineSC linechicken embryofibroblasts
wereC/O, V-,chfnegative,and ALV gsantigen
neg-ative. Line 100 chicken embryo fibroblasts were of
eitherV+C/BE or V+C/O phenotype. V+C/BE line
100cellsspontaneously producedlow titers ofRAV-0
and, since they were resistantto exogenous RAV-0
infection duetotheabsence of receptorsrequiredfor
RAV-0 penetration, presumably contained only
en-dogenousRAV-0-relatedgeneticinformation (6,8,33).
V+C/Oline 100 ceUsproduced hightiters ofRAV-0, presumably duetoexogenous infection with RAV-0
as aresult of exposuretotheRAV-0 thatthey
spon-taneously produced (6, 8, 33). Ringneck pheasant ceUs, turkeyceUs,Japanesequailcells,and Pekinduckcels
were virusnegative (assayed by sedimentable DNA
polymerase activity in culture fluids), helper factor
negative,and ALV gsantigen negative.
LA335(pol'),anearly temperature-sensitive
coor-dinatemutantofPragueRSVsubgroupC
(PR-RSV-C)withathermolabile DNApolymerase (21, 22,36),
was obtained from W. S. Mason, Philadelphia, Pa.
Schmidt-RuppinRSVsubgroupD(SR-RSV-D),B77
avian sarcoma virus (subgroup C), Rous-associated
virus 1 (RAV-1) (subgroup A), and Rous-associated
virus7(RAV-7) (subgroup C)wereobtained from H.
M.Temin, Madison,Wis. PR-RSV-A and PR-RSV-C
wereobtained from W.S. Mason.
Virusesweregrown andassayed by focus formation
as previously described (4). Viruses were cloned by
plating virus-infected chicken embryo fibroblasts in
softagarmedium(10),usingfeederlayersof
mitomy-cin C-treatedturkey embryo fibroblasts.
PreparationofDNA. DNA was extracted either
from cultured ceUs or from embryos as previously
described (5, 6). The average molecular weights of
DNAsextracted by this method were approximately
50 x 106 (5). Fragments of DNA for marker-rescue
experimentswereprepared bysonicationof the DNA
to average molecular weights of approximately 5 x
105(4).
Assayof infectious DNAbytransfection.
Cul-turesof C/Ochicken embryo fibroblasts were prepared
and treated withhigh-molecular-weight(unsonicated
DNA,using the calcium method of Graham and Van
der Eb(11)aspreviouslydescribed(6). DNA-treated
ceUswerecultured in ET mediumsupplementedwith
2% fetal bovine serum and polybrene (5pg/ml) for
approximately 4 weeks (four transfers) after DNA
treatment.Virusproductionwasdetennmined byassay
ofsedimentable DNApolymerase activityinculture
fluids (6). The assay was quantitated by end point
dilution of the donor DNA (5).
Assay of subgenomic DNA fragments by
marker rescue. Cultures of C/O or C/E chicken
embryo fibroblastsweretreatedwith sonicated DNA
fragmentsand wereinfected with LA335 (poll)
(ap-proximately0.1focus-formingunit[FFU]/cell)atthe
permissive temperature(35°C)aspreviouslydescribed
(4). WhenC/Ecells were used as recipients, turkey
cells (T/BD) were added after infection with LA335
to permit replication ofsubgroup E progeny RSV.
After6daysof incubationat35°C,supernatantmedia
wereharvested and assayed on freshC/0 orT/BD
cellsatthenonpermissivetemperature(41°C)toselect
for wild-type (pol+) progeny virus. AU 41°C assay
platespositivefortransformationwereharvested,and
the supernatant media were reassayed in a second
passage at 41°C to distinguish phenotypic leakiness
of the parentalLA335mutant from the presence of
wild-type progeny RSV (4).
Thermolability of virion DNA polymerases.
Virions were concentrated 100-fold from cell-free
su-pernatant media, suspended in Eagle medium, and
disrupted byaddition ofanequalvolume ofdisruption
buffer(0.05Msucrose-0.001 M EDTA-0.03 M
dithi-othreitol-0.01 M Tns-hydrochloride [pH 7.4]-0.25%
Nonidet P-40). Disrupted virions were incubated at
41°C, andsampleswithdrawnatdifferenttimeswere
keptat0°Cuntilallsampleswerecollected.Portions
ofdisruptedvirions(25pl)werethen addedto 100p1
of polymerase reaction mixture containing 0.02 M
Tris-hydrochloride (pH 8.0), 0.025 M KCI, 0.02 M
MgCl2,0.4mMEDTA,10mMdithiothreitol,0.1mM
dATP,0.075mMATP,2.5pgofphosphoenolpyruvate,
0.5U of pyruvatekinase,1.25pgofpoly(dA-dT),and
1.9 pCi of [3HJTTP (50 Ci/mmol, Schwarz/Mann,
Orangeburg, N.Y.). Sampleswere incubated at35°C
for 15, 30, and60minand25-plportionswereassayed
foracid-precipitable radioactivity.
Antibodyneutralizationof virion DNA polym-erases. DNA polymerase neutralizations were
per-formed as described by Mizutani and Temin (23).
Concentrated virions were disrupted with detergent
and incubatedwithantibodyat roomtemperature for
30min. DNApolymeraseactivityremainingwasthen
25, 789
on November 10, 2019 by guest
http://jvi.asm.org/
assayedat37°C. PreparationsofimmunoglobulinG
(IgG)from antiseraagainst RSV(RAV-0)DNA
polym-erase (IgG-RSV-RAV-0), spleennecrosis virus DNA
polymerase(IgG-SNV),chickencellDNApolymerase
a(IgG-a),and chickencellDNApolymerase ,B
(IgG-,f)(23, 24)weregenerously provided byH. M. Temin.
RESULTS
MarkerrescueofLA335 (polP) with DNA
fragments of uninfected avian cells. The biological activity of fragments of cellular DNAs
was tested in marker-rescue experiments with
LA335
(pol').
Recipient cultures ofchickenem-bryo fibroblastsweretreated withsalmonsperm
DNA or with sonicated fragments of cellular
DNAs and were infected with LA335at 350C. After 6 days of incubationat350C,supernatant
mediawere harvested, and 0.2-ml samples
(ap-proximately 5x 102FFUat350C)wereassayed onfreshcellsat410C.Foci oftransformedcells
(onetofour fociperplate)wereinducedat410C
by supernatant media from approximately 5% of the cultures treated with either salmonsperm orcellular DNAs. To determine whether these
fociwereduetophenotypic leakiness of LA335
ortopot'progenyRSV, thesupernatantmedia ofall410Cassayplates positive for
transforma-tionwere harvested andreassayed inasecond
passage at410C. Supernatant media harvested from transformation-positive 410Cassayplates
ofallcontrol cultures treated withsalmonsperm
DNA and from assay plates ofsome cultures
treated with fragments of cellular DNAs had titers of <20 FFU/mlin the second passage at
410C,indicating that focus formation in the first
assay at 410C had resulted from phenotypic leakiness of LA335.Incontrast,supernatant
me-dia harvested fromtransformation-positive410C
assayplates ofsomerecipient cultures treated
with cellular DNA fragments had titers of 102
to 2 x 103 FFU/ml in the second passage at
410C, indicating the presence ofpol' progeny
RSV.
None of 435 control cultures treated with
salmon sperm DNA and infected with LA335 producedprogenyRSV thatreplicatedat410C
(Table 1), indicating that reversion of LA335to
pol+ RSV didnotoccuratasignificant frequency
inthese experiments. Since approximately 5 x
102FFU ofpol'RSVwereassayedperculture,
the reversionfrequency of LA335 appeared to
be less than 5 x 10' (Table 1). As previously
reported (4), progeny RSV that replicated at
410C were isolated in marker-rescue
experi-ments with sonicated fragments of DNA of
RAV-0-infectedV+C/Oline 100cells(Table 1).
Progeny RSVthatreplicatedat410Cwerealso
produced by cells infected with LA335 and
treated with sonicated fragments of DNAs of
TABLE 1. Assay of subgenomic fragments of
cellular DNAs by marker rescue withLA335(polf9a
Fraction of pO- Frequencyof Donor DNAb itive recipient
Fprogeny"
culturesc
Salmon sperm 0/435 <5x 10-6
ChickenV+C/O 1/30 1x 10-4
Chicken V+C/BE 2/30 2x 10-4
Chicken V- 3/288 2X10-5
Pheasant 2/199 2X10-5
Quail 3/262 2x10-5
Turkey 3/138 4 x10-5
Duck 0/177 <1 x10-5
Mouse 0/208 <1 x 10-5
aCultures of chicken embryo fibroblasts were
treated with5
jig
ofsalmon sperm DNA or sonicatedfragments ofcellular DNAs and were infected with
LA335(polls) at350C. The formation ofpol+ progeny
RSV thatreplicatedat410Cwasassayed as described
inthe text.
b DNAs wereextracted from line 100V+C/O and
V+C/BE chicken embryo fibroblasts, SPAFAS
V-gs-chf chicken embryo fibroblasts, ringneck
pheas-ant embryo fibroblasts, Japanese quail embryos and
Japanesequailembryo fibroblasts, Orlopp turkey
em-bryofibroblasts, Pekin duckembryo fibroblasts, and mouseNIH 3T3cells. DNAsweresonicated to average
molecular weights of approximately5 x105.
cNumber of cultures that produced progeny RSV
thatreplicatedat41WCoverthe total number treated.
dRatio of the numberofpol'progenyRSV isolated
to the total FFU ofpolls RSV assayed from each
groupof DNA-treated cultures. Each sample of
super-natantmedium, assayedat41WCtodetect pol+
prog-enyRSV, contained approximately 5 x 102 FFU of
pollRSV(assayedat35°C).
uninfectedVIC/BEand V- chicken cells(Table
1). Inaddition,progenyRSV thatreplicatedat 410C were isolated in marker-rescue
experi-ments with sonicated fragments of DNAs of uninfectedpheasant,quail,andturkey cells, but notwithsonicatedfragmentsof DNAs of unin-fected duck ormouse cells (Table 1). The fre-quencies ofpol progenyRSVproducedby cul-turestreated withfragments ofchicken,
pheas-ant,quail,andturkeyDNAswere2 x 10-5to 2
X 10-4 (Table 1). Thus the frequencies of for-mationofpol+progenyRSVinculturestreated with these avian cell DNAswere atleast4-to 40-foldgreaterthanthereversion frequencyof LA335in cultures treated with salmon sperm DNA.
Progeny RSV that replicated at 410C were
produced by 1 to 5% of the recipient cultures treated with5,ug of eachdonorDNAactive in marker rescue (Table 1). The efficiency of marker rescuewith DNAs ofuninfectedavian cells, therefore, appeared to correspond to ap-proximately 1 to5infectious units (IU) per mg
on November 10, 2019 by guest
http://jvi.asm.org/
RESCUE OF ENDOGENOUS ALV DNA POLYMERASE
of DNA. This efficiency was two- to fivefold
lower than the efficiency of markerrescue
pre-viously reported with DNA of RSV-infected cells (4).
Transfection assaysof avian cellDNAs.
The isolation ofprogeny RSV that replicated
at 410C in marker-rescue experiments with LA335(poll) andsonicatedfragments ofDNA of uninfected chicken, pheasant, quail, and
tur-keycellsindicated that the DNAs of these
un-infected aviancellscontainedendogenous
ALV-relatedgenetic informationwith detectable bio-logical activity in marker-rescueassays.
There-fore, high-molecular-weight (not sonicated) DNAs of uninfected chicken, pheasant, quail, and turkey cells were assayed by transfection todetermine if they contained infectious
endog-enousviralgenomes.
As previously reported (6), infectious RAV-0 DNAwas obtained fromV+C/O line 100 cells exogenouslyinfected withRAV-0, but notfrom
uninfected V+C/BEorV- chickencells contain-ing only endogenous RAV-0-related DNA
se-quences (Table 2). In agreement withprevious results (6), the specific infectivity of DNA of
V+C/O cells, assayed by end point dilution of
the donorDNA,was approximately 1
IU/jg
ofDNA(datanotshown).Incontrast,thespecific infectivities of DNAs ofV+C/BE and V- cells
were less than 0.02 IU/,ug of DNA (Table 2).
Similarly, the DNAs of uninfected pheasant, quail, and turkey cellswerenotinfectious(<0.04
IU/,tgofDNA) (Table 2), indicatingthat
unin-fected cells of theseheterologous avianspecies
didnot contain infectiousendogenous proviral
DNAs.
[image:4.501.252.441.301.590.2]Since theDNAsof uninfectedchicken, pheas-ant,quail, and turkeycellswereactive inmarker
TABLE 2. Assayofinfectivity of donor DNAsby
transfectiona
DonorDNAb Fraction ofpositive recipientculturesc ChickenV+C/O ... 7/8
Chicken V+C/BE ... 0/25
Chicken V... 0/25
Pheasant ... 0/11
Quail 0/12
Turkey 0/12
aCultures of chicken embryo fibroblasts were
treated with 5,ugofhigh-molecular-weight
(unsoni-cated)DNAperculture.Virusproductionwasassayed
bysedimentable DNApolymerase activityinculture fluids.
bThe donor DNAs were asdescribed inTable 1.
Theaveragemolecularweightsof unsonicatedDNAs
wereapproximately50x 106.
cNumber ofcultures thatproduced virusoverthe
total numbertreated.
rescuebutnotin transfection, the marker-rescue assay appeared to detect endogenous virus-re-lated DNA sequences that were not part of an
infectious provirus.
Characterization of pol+ progeny RSV isolated by marker rescue. Progeny viruses
isolated in marker-rescue experiments with LA335
(pol')
were assayed at 35 and 41°C todeterminetheir efficienciesoftransformation at
thepermissiveandnonpermissivetemperatures.
In contrast to the parental RSVmutant, allof the progeny RSV isolates obtained in marker-rescueexperimentswith DNAfragments of un-infected chicken, pheasant, quail, and turkey cellstransformed with similarefficiencies at 35 and41°C (Table3).Theseresults indicated that theprogenyRSV isolated by selection for trans-formation andreplicationat410C(Table1) were p0l+.
Totestthegeneticstability of the pol+
prog-TABLE 3. Efficiencyoftransfornationofprogeny
viruses isolatedbymarkerrescuewith LA335(polfs)
Titer (FFU/ml) EOT(41/
35°C 410C 3500)b
Controls
LA335(polt) 3x104 <5 <2x10-4
PR-RSV-C(pol+) 9 x102 1x 102 0.1
B77V(pol+) 1x 107 1X 107 1.0
Marker-rescue prog-enyc
ChickenV+C/O
-1 4x102 3x101 0.1
ChickenV+C/BE
-1 1X 103 2x102 0.2
-2 4x102 4x101 0.1
Chicken
V--1 1x 103 2x 102 0.2
-2 5x 102 1x 102 0.2
-3 3x102 2x102 0.7
Pheasant
-1 6x 102 5x 102 0.8
-2 2x103 3x 10 1.5
Quail
-1 1 x102 1x 102 1.0
-2 6x 10 1X104 1.7
-3 3x 103 1X103 0.3
Turkey
-1 2x 103 4 x102 0.2
-2 2x103 5x102 0.3
-3 3 x 103 3 x 103 1.0
a
Vinrses
wereassayedfortransformationat35and41°C, and fociwerecounted4to6daysafterinfection.
bEOT,Efficiency of transformation: ratio of virus
titerat41°Ctothe titerat35°C.
cStocks of progeny viruses from marker-rescue
ex-periments(Table 1)wereharvested aftertwopassages at41°C.The progeny virus isolatesweredesignated
by thesourceofDNAfragmentsusedinmarkerrescue
followedbythe isolate numberof the progeny virus.
VOL. 25, 1978 791
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.501.45.241.465.581.2]enyRSV,representative isolates obtained with fragments of DNA of uninfected V- chicken cells, pheasantcells, andquailcells were cloned in softagar at 350C. Individual colonies of each
pot' progenyRSV isolatewerepickedand
pas-saged at350Ctogrow virusstocks,whichwere thenassayed fortransformationat35and410C. All clones of each of the polt progeny RSV isolates transformed with similar efficiencies at the two temperatures (Table 4).
Therefore,
the pol+ marker of the progeny RSV isolated in marker-rescueexperimentswasstabletocloningandpassageatthepermissivetemperature. Some of the pol progeny RSV previously
isolated in marker-rescue experiments with LA335 (poll, subgroup C) and fragments of DNA ofpot subgroup D RSV-infected cells acquired thesubgroupDenvelopespecificityas well as thepoltmarker of the RSV strain used
forpreparationofDNAfragments (4).Therefore
the subgroup ofeach of the progeny RSV
iso-lates obtained in the present experiments with fragments of DNA of uninfected cellswas
deter-mined. Progeny viruses were assayed on C/O
cells,
C/E
cells, T/BD cells, andC/E cells in-fected withRAV-1 (subgroup A)orwith RAV-7 (subgroup C) All of the pol progeny RSV isolates were infectious for C/0, C/E, T/BD, [image:5.501.67.256.398.570.2]and RAV-1-infected C/E cells, but were not infectious for RAV-7-infected C/E cells (data
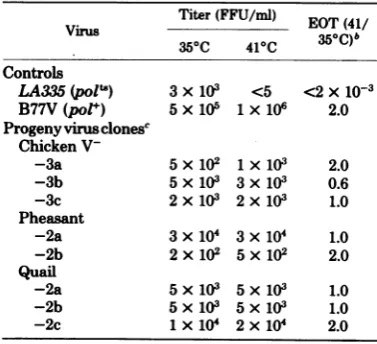
TABLE 4. Efficiency oftransformationofprogeny
virusisolatesclonedat350Ca
Titer (FFU/ml) EOT (41/
350C 410C )
Controls
LA335(polls) 3x103 <5 <2x10-3
B77V(pol+) 5x 105 1x 106 2.0
Progenyvirusclonese
Chicken
V--3a 5 x102 1x 103 2.0
-3b 5x103 3x 103 0.6
-3c 2x103 2x 103 1.0
Pheasant
-2a 3x104 3x 104 1.0
-2b 2x 102 5x102 2.0
Quail
-2a 5 x103 5 x 103 1.0
-2b 5x103 5 x 103 1.0
-2c 1 x104 2 x 104 2.0
a
Progeny
viruses isolated from marker-rescueex-periments(Table 1)werecloned in soft agarat350C,
passaged once at350C,andassayed for transformation at 35and 410C.
bEOT, Efficiencyoftransformation: ratio of virus
titerat410Ctothe titerat35°C.
'Progeny viruses from marker-rescue experiments
werenamedasdescribed in Table3.Clones of progeny
virusisolatesaredesignated byaletterfollowing the
isolate number.
obtained with representative isolates only are
presentedin Table5).Therefore, all of thepot
progenyRSVisolates obtainedbymarkerrescue
withfragmentsofDNA ofuninfectedavian cells
retained thesubgroupCenvelopespecificity of LA335.
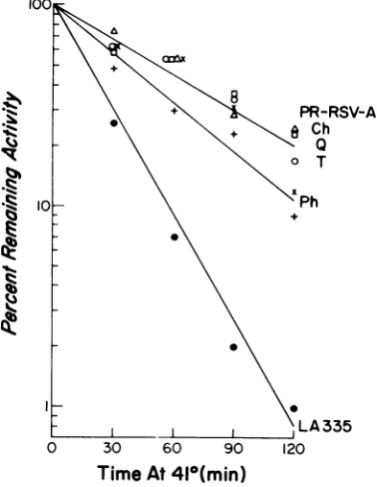
Tocharacterize the DNA polymerases ofpol progenyRSV, thethermolabilitiesof thevirion
DNApolymerasesofrepresentative pot
prog-enyRSV isolateswerestudied.Preparationsof
disrupted virions were incubated for different
times at 410C, and the amount of remaining
DNA polymerase activity was determined by
assay at 350C. The DNA polymerase of
dis-rupted virions ofwild-type PR-RSV-A (pot)
was inactivated at 410C with ahalf-life of
ap-proximately 50 min (Fig. 1). In contrast, the
DNApolymeraseofdisrupted virions ofLA335
(pol')
was inactivated threefold more rapidlyat410C,withahalf-lifeofapproximately17min
(Fig. 1). Theratesofinactivationat410C ofthe
DNA polymerases of representative progeny
RSV isolated in marker-rescueexperiments with
DNAfragmentsof V- chicken cells, quail cells,
andturkeycellswereindistinguishablefromthe
rate of inactivation of the PR-RSV-A (pot)
DNA polymerase, and the rate ofinactivation ofthe DNA polymerase of the representative progeny RSV isolate obtained with fragments
ofpheasant cell DNA was only slightly more
rapid (half-life, 36 min) than that of the
PR-RSV-A(pot) DNA polymerase(Fig. 1). Control experiments in whichdisrupted virions of LA335 weremixed withdisrupted virions of
PR-RSV-Aorof thepolt progeny RSV isolate obtained
withfragments of pheasant cellDNA indicated that the differences in the thermolabilities of the DNA polymerases of LA335 and pol RSV were notdue to diffusible factors in the virion preparations (data not shown). The thermola-bilities of the DNA polymerases of the pot progenyRSV isolated inmarker-rescue experi-ments, thus, appeared similar to that of the DNA polymerase of wild-type (polt) RSV.
Theantigenic specificities of the DNA polym-erasesofpol progeny RSV werecharacterized by neutralization tests of the DNA polymerases ofdisrupted virions. DNApolymerases of all of thepol progeny RSV isolates and of PR-RSV-A were neutralized (over 90%) by antibody against ALV DNA polymerase (50
#&g
ofIgG-RSV-RAV-0),but notbyantibodyagainst
retic-uloendotheliosis virus DNApolymerase (50 ,ug
ofIgG-SNV), chicken cell DNApolymerase a
(50
itg
ofIgG-a),orchickencellDNA polymer-ase 86 (200jug
ofIgG-,8)
(data not shown). Inaddition, the kinetics of neutralization of the
DNApolymerasesofpotl progeny RSVby IgG-RSV-RAV-0 did not differ significantly from VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
RESCUE OF ENDOGENOUS ALV DNA POLYMERASE
TABLE 5. Subgroupofprogenyvirusesisolated bymarkerrescuewithLA335(poltenvE)
Titer(FFU/ml)'
Virus
C/O C/E T/BD C/E(RAV-1) C/E(RAV-7)
Controls
PR-RSV-A 2 x104 4x 104 i xlo5 <5 1x 104
PR-RSV-C 5xi02 5 X102 i X103 5 x102 <5
SR-RSV-D 2x 105 2x105 <5 1xlo5 1 x lo5
Marker rescueprogenyb
ChickenV+C/BE-2 2x102 2x102 1x102 1x 102 <5
ChickenVV-l 5x 102 5x 102 1x 102 1x 102 <5
Pheasant-2 1x03 1X 103 2x 103 2x102 <5
Quail-3 4 x103 4 x103 4 x103 5x 102 <5
Turkey-1 3xiO2 3x102 1xiO2 1x1i2 <5
aViruses wereassayed for focus formation on cultures of C/O chicken embryo fibroblasts, C/E chicken
embryofibroblasts,T/BDturkeyembryofibroblasts,andC/Echickenembryofibroblasts whichwerepassaged
three times after infection with RAV-1[C/E (RAV-1) cells]orwith RAV-7[C/E (RAV-7)cells].
bProgeny viruses
from
marker-rescue experimentswereharvested anddesignatedasdescribed
in Table3.30 60 90
Time At41°(min)
FiG. 1. ThermolabilityofvirionDNApolymerases
ofprogeny RSV. Virions of LA335 (pol) (0),
PR-RSV-A @pot) (0), andrepresentativeprogenyvirs
isolatesfrom marker-rescue experiments with DNA
fragments ofV- chickenceUs (Ch, x), pheasant ceUs
(Ph, +), quail ceUs (Q, 0), and turkeycells (T, )
weredisrupted with Nonidet P-40 and incubated for
differenttimes at41°C. Theremaining DNA
polym-eraseactivitywasthenassayedat35°C. Theprogeny
virusesfrommarker-rescueexperimentswerethe
iso-lates designated chicken V-3, pheasant-2, quail-2,
andturkey-3inTabk3.The DNApolymerase
activ-itiesofeachofthedisruptedvirionsbefore incubation
at41°C(100%activity)werebetween5,000and20,000
cpmofpHJTMP incorporatedin60minat35°C.
0 2 3 4
IgG-RSV-RAV-O
(,u.g)
FIG. 2. Neutralization of virionDNA polymerase
activity ofprogeny RSV. Virionsof PR-RSV-A (0), pol+ progenyRSVisolatepheasant-I (0), andpoll
progenyRSVisolateturkey-I (x)weredisruptedwith
Nonidet P-40 andincubated with IgG-RSV-RAV-O
for 30min at room temperature. RemainingDNA
polymeraseactivitywasassayedat37°C. The 100%
activity for each ofthe viruses was between 1,500 and3,000cpmofpH]TMPincorporated in 30min at37°C.
neutralization of PR-RSV-A DNApolymerase.
(Representative data are presented in Fig. 2;
similar kinetics ofneutralization were also ob-served withpoP progeny RSV isolates
pheas-ant-2, quail-i, quail-3, turkey-2, and turkey-3.)
The DNA polymerases ofpot progeny RSV,
therefore,appeared to have antigenic
determi-nants similarto those ofwild-type RSV DNA polymerase.
DISCUSSION
Wild-type (pol) progeny RSV were isolated inmarker-rescueexperimentswithLA335(polts)
VOL. 25, 1978 793
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.50.239.272.516.2]andfragments of DNA ofuninfected VIC/BE chickencells,V-gs-chf chickencells, pheasant cells, quailcells, and turkey cells, butnotwith fragmentsof DNAof uninfected duckormouse cells. The pol+ progeny RSV isolates did not appear toberevertantsofLA335, sincenopot'
progenyRSVwereisolated frommorethan400 control cultures treated with salmon sperm DNA. Since approximately 5 x 102 FFU (at
350C)perculturewereassayedat41°Ctodetect
pol+progeny, the frequencyof pol+ revertants in the progeny of salmon sperm DNA-treated control cultureswasless than5x106. Asimilar reversion frequency of LA335 was previously notedby Linial and Mason (21), who found that none of25 LA335 clones that they isolated as
fociat41°C wereproduced bypol+revertants.
Since theefficiencyof focus formationby LA335
at 410C was approximately 5 x 10-4 (21), the
reversionfrequencyof LA335 in theexperiments
of Linial and Mason (21) appeared to be less than
10-'.
In contrast to the absence of pol+ revertantsintheprogenyofsalmonsperm DNA-treated control cultures, 14pol+ progeny RSV were isolated from cultures treated with frag-ments ofchicken, pheasant, quail, and turkey DNAs. Thefrequenciesofpol+ RSV in the prog-eny produced by cultures treated with these avian cell DNAs were 2 x 10' to 2 x 1O-4. Since these frequencies were atleast4- to 40-fold greater than the reversion frequency of LA335 incontrolcultures, itappearedthat for-mation ofpot'progenyRSVprobablyoccurred by recombination between LA335 and donorDNA fragments of the uninfected avian cells.
Inpreviousexperimentswithfragmentsof DNA
of RSV-infected cells,pol+ progeny RSV were obtained whenrecipientcellswereinfected with LA335 within 6 h after treatment with DNA fragments, butnotwhen the cellswereinfected with LA335 either24to48hbefore DNA
treat-ment or24 hafter DNA treatment (4;
unpub-lished observations). These observations are consistent with the possibility that recombina-tion inmarker-rescueexperiments might occur betweendonorDNAfragmentsandnewly syn-thesizedunintegratedDNAofmutantRSV.
Thepot'progenyviruses isolatedby marker rescue were genetically stable and had virion
DNApolymeraseswiththermolabilitiessimilar
tothatofwild-typeRSVDNApolymerase.The
pol+progenythusappearedtorepresent
genet-icallystable recombinantsofRSVwithcellular
genetic information homologoustothe ALVpol gene.Neutralization assaysof the DNA
polym-erase activities of disrupted virions indicated thatthe DNApolymerases ofpol+progenyRSV
wereserologically similartothe DNA
polymer-aseofwild-typeRSV. However,moresensitive
serological or biochemical characterizations of purified DNApolymerasesmightdetect altered properties of the enzymes, possibly related to cellulargeneticinformation.
The uninfected avian cells usedasdonors of DNA didnotcontain DNAprovirusesof
endog-enousavian retroviruses thatwereinfectious in transfection assays (6; Table 2). Therefore, the marker-rescue assay appearedtodetect nonin-fectious forms of endogenous virus-related
DNAs, indicating that uninfected chicken,
pheasant,quail, andturkey cells contained
en-dogenous DNAsequencessufficientlyrelatedto atleastaportionof the ALVpolgene toyield pot'recombinants. However,theactivityof
un-infected cell DNAs in marker-rescue
experi-ments does not necessarily indicate that the
endogenouspol-related geneticinformation
rep-resents acomplete polgene.
Although the endogenous RAV-0DNA of un-infected V+C/BE chicken cells was not infec-tious in transfection assays, the cells
sponta-neously produced a low titer of RAV-0 and
thereforeappearedtocontainacomplete endog-enous RAV-0 genome, including abiologically active ALV polgene (6, 8, 33). Therefore, the DNA fragments of V+C/BE cells active in markerrescuemayhave been derived from the polgeneof theendogenous RAV-0genome.
In contrast, uninfected V- chicken cells do not produce RAV-0, although they appear to contain DNAsequenceshomologousto mostof the nucleotide sequences of RAV-0 RNA (20, 25,26, 31). Some V- chickencellsproduce pro-teins relatedtoALVinternalstructural proteins and envelope glycoproteins (2, 12, 14, 28, 39),
buttheyapparentlydonotcontain aDNA
po-lymerasesimilartoALVDNA polymerase(42),
nor are uninfected V- chicken embryo fibro-blasts able to complement or recombine with mutantsofRSVdefective in pol (13, 40). How-ever, particles containing RNA-directed DNA
polymeraseactivitiesserologically distinctfrom
ALV DNA polymerase have been reported in chickenembryos (19) and in theallantoic fluid of uninfected chickeneggs(1). Inaddition, com-petition radioimmunoassays have detected an-tigenic material relatedtoALV DNA polymer-asein extractsof uninfected V- chickenembryo fibroblasts (27) and weak serological relation-ships between ALV DNA polymerase and chicken cell a and ,6 DNA polymerases have
beenreported(24). Theisolation
ofpolt
progenyRSV by marker rescue with fragments of V-chicken cell DNAindicates thatV- chickencells
containgeneticinformation relatedto atleasta portion of the ALVpolgene. Thelack of
on November 10, 2019 by guest
http://jvi.asm.org/
RESCUE OF ENDOGENOUS ALV DNA POLYMERASE
mation of pot recombinants after infection of chicken embryo fibroblasts with pot (13) or
poll (40) RSVmayindicate that cell-virus re-combination in virus-infected cells is dependent ontranscription of the endogenous virus-related genes (40, 41). The absence of complete tran-scripts of the ALV pol gene in the RNA of uninfectedV-gs'chf+ chickencells (38) is con-sistent with this hypothesis.
Uninfected pheasant, quail, and turkey cells appear tocontain DNA sequences homologous to afraction of the nucleotide sequences of RAV-0RNA, whereas the DNAs of uninfected duck and mouse cells lack detectable sequence
ho-mologywith RAV-0 (20, 25, 31). The DNA of
pheasantcellsappears to hybridize to 10 to 20% of the RNA sequences of RAV-0 (20, 25, 31). Kang and Temin (20) reported that DNAs of uninfected quail and turkey cells hybridized,
respectively, to 15and 10%of the sequences of
RAV-0 RNA, although others have reported that the DNAs ofuninfectedquail (25, 31) and
turkey (31) cells hybridized to less than 5% of
theRNAofRAV-0.Uninfectedpheasant, quail, andturkeycells,but notduck cells,also contain DNA sequences homologous to approximately 10% of the RNA of reticuloendotheliosis viruses (20). Biological activity of endogenous
retrovi-rus-relatedgenetic information in pheasantcells,
partridge cells,andChinesequailcellshas been
demonstratedby complementation and
recom-binationbetweenALV andendogenousenvelope
glycoprotein genes (3, 9, 15, 34). In addition,
infectiousendogenous viruses have been isolated fromcells of severalspecies of pheasants (3, 15)
and from Chinese quail (3). The endogenous
viruses isolated from Chrysolophus pheasants represent a third group of avian retroviruses unrelated by immunological cross-reactivity or nucleic acid hybridizationtothe avian leukosis
orreticuloendotheliosis virus groups (16). The
endogenouspol-related genetic information we have detected in marker-rescue experiments with the DNAs of heterologous avian species could correspond to endogenous ALV-related DNAsequences detectedby nucleicacid hybrid-ization, to endogenous genetic information re-latedtothereticuloendotheliosisvirusor
Chry-solophus pheasant virus groups, or to
genetic
information related to cellular DNA
polymer-ases. Furthercharacterization of thepot prog-eny virus isolatesmay elucidate the nature of theendogenouspol-related genetic information inuninfectedaviancells.
ACKNOWLED)GMENTS
Ithank L B.Crittenden,W. S.Mason,M.Smith,H. M.
Temin, and R.A.Weinbergforgenerously supplying experi-mentalmaterials.I amgratefultoS.Castellot,K.MacCarthy,
S.Okenquist,andL.Silverman for technical assistance and toN.CopelandandD. M.Livingstonforhelpfulcomments
onthemanuscript.
ThisinvestigationwassupportedbyPublicHealth Service
grantCA18689, awardedbytheNationalCancer Institute.
LITERATURE CITED
1. Bauer,G.,and P. H. Hofschneider. 1976. An RNA-dependentDNApolymerase, differentfrom the known viral reversetranscriptases, in thechickensystem.Proc. Natl.Acad.Sci.U.S.A.73:3025-3029.
2. Chen, J.H.,W. S.Hayward, andH.HanaL 1974. Avian tumor vrs proteins and RNAin uninfected& chickenembryocells. J.Virol. 14:1419-1429.
3. Chen,Y.C., andP. K.Vogt.1977.Endogenousleukosis viruses in the avian family Phastanidae. Virology 76:740-750.
4. Cooper, G. M., and S. B. Castellot. 1977. Assay of noninfectiousfragmentsof DNA ofavian leukosis virus-infectedcellsby marker rescue. J. Virol. 22:300-307.
5. Cooper,G.M., and H.AL Temin.1974.Infectious Rous sarcoma virusandreticuloendotheliosisvirus DNAs. J. Virol. 14:1132-1141.
6. Cooper, G.M., andH.MLTemin. 1976.Lackof infectiv-ity of theendogenousavianleukosis virus-relatedgenes in the DNA of uninfected chicken cells. J. Virol. 17:422-430.
7.Crittenden, L B.,J. V.Motta, andE. J.Smith.1977. Genetic control ofRAV-0production inchickens. Vi-rology76:90-97.
8. Crittenden,LB.,E. J.Smith,R. A.Weiss,and P.S. Sarma. 1974. Host genecontrolofendogenousavian leukosis virusproduction. Virology 57:128-138.
9. Fujita,D.J.,Y.C.Chen,R. R.Friis, andP. K.VogL 1974.RNA tumorvirumse ofpheasants:characterization of avian leukosisvirussubgroups F and G.Virology 60:558-571.
10. Graf,T. 1973.Twotypesof targetcellsfor transformation with avian myelocytomatosis virus. Virology 64: 398413.
11. Graham,F.L, and A. J. Vander Eb. 1973. A new
techniqueforthe assay ofinfectivityof human adeno-virus 5 DNA.Virology52:456-467.
12. Halpern,MLS.,D. P.Bolognesi,R. R.Friis,and W. S. Mason.1975.Expressionof themajorviral glycopro-tein of avian tumorvirus in cells ofchf(+) chicken embryos.J.Virol.15:1131-1140.
13. Hanafusa, H., andT. Hanafusa. 1971. Noninfectious
RSV deficient in DNA polymerase. Virology 43: 313-316.
14.Hanafusa,H.,T.Miyamoto,and T.Hanafusa. 1970.
Acell-associatedfactor essentialforformation of an infectiousforn of Roussarcomavirus. Proc. Natl. Acad. Sci.U.S.A.66:314-321.
15.Hanafusa, T.,and H.Hanafusa.1973.Isolation of leu-kosis-typevirusfrompheasant embryo cells:possible presenceof viral genes incells.Virology51:247-251.
16.Hanafusa, T., H. Hanafusa, C. E. Metroka, W. S. Hayward, C.W.Rettenmier,R.C.Sawyer,R. M. Dougherty, and H. S. DiStefano. 1976. Pheasant virus: newclass ofribodeoxyvirus. Proc. Natl. Acad. Sci. U.S.A.73:1333-1337.
17.Hanafusa,T.,H.Hanafusa,and T.Miyamoto. 1970.
Recoveryof anewvirus fromapparently normal chick cells byinfection with aviantumorvinuses.Proc.Natl. Acad. Sci.U.S.A.67:1797-1803.
18. Hayward,W.S.,and H.Hanafus 1975. Recombina-tionbetweenendogenousandexogenousRNA tumor virusgenes asanalyzed by nucleic acid hybridization. J. Virol.15:1367-1377.
19. Kang,C.-Y.,and H. M. Temin.1972.Endogenous
RNA-795
VOL. 25, 1978
on November 10, 2019 by guest
http://jvi.asm.org/
directed DNA polymerase activity in uninfected chicken embryos. Proc. Natl. Acad. Sci. U.S.A. 69:1550-1554.
20. Kang,C.-Y., and H. M. Temin. 1974. Reticuloendothe-liosis virusnucleicacid sequences incellularDNA. J. Virol. 14:1179-1188.
21. Linial, M.,andW. S. Mason. 1973.Characterization of twoconditionalearlymutantsofRoussarcomavirus. Virology53:258-273.
22. Mason,W.S.,R. R.Friis,M. Linial, and P. K.Vogt. 1974. Determinationof the defective function in two mutants ofRous sarcoma virus. Virology 61:559-574. 23. Mizutani,S., andH. M.Temin. 1973.Lackof serological relationshipamong DNApolymerasesofavian leukosis-sarcoma viruses, reticuloendotheliosis viruses, and chicken cells. J. Virol. 12:440-448.
24. Mizutani, S., and H. M.Temin. 1974. Specific serological relationshipsamongpartiallypurified DNA polymer-asesof avian leukosis-sarcoma viruses, reticuloendothe-liosisviruses andavian cells. J. Virol. 13:1020-1029. 25.Neiman,P. E. 1973.Measurement ofendogenousleukosis
virusnucleotidesequences in the DNA of normal avian embryos by RNA-DNA hybridization. Virology 53:196-204.
26. Neiman,P.E., H.G.Purchase,andW.Okazaki.1975. Chicken leukosis virus genome sequences in DNA from normal chickcelUsand virus-induced bursal lymphomas. Cell 4:311-319.
27. Panet,A.,D.Baltimore,and T.Hanafusa.1975. Quan-titation of avian RNA tumor virus reversetranscriptase byradioimmunoassay.J.Virol. 16:146-152.
28. Payne,L.N.,and R.C.Chubb. 1968. Studies on the nature and genetic control of an antigen in normal chickembryoswhichreactsinthe COFAL test. J. Gen. Virol. 3:379-391.
29. Robinson,H. L 1976. Intracellular restrictionon the growth of inducedsubgroupEavian type C viruses in chickencells.J. Virol.18:856-866.
30. Robinson,H.L, C.A.Swanson,J.F.Hruska,and L B.Crittenden. 1976.Production ofunique C-type virusesby chickencellsgrown inbromodeoxyuridine. Virology69:63-74.
31. Shoyab, M.,and M. A.Baluda.1975.Homologybetween
avianoncornavirus RNAs and DNA from several avian species. J.Virol. 16:1492-1502.
32. Shoyab, M., and M. A. Baluda. 1976. Ribonucleotide sequencehomologyamong avianoncornaviruses. J. Vi-rol. 17:106-113.
33. Smith,E.J., L B.Crittenden,and T. H.Brinsfield, Jr. 1974. Status of theendogenousavian leukosis virus in resistant cells from a producing line. Virology 61:594-596.
34. Temin, H. M., and V. K. Kassner. 1976. Avian leukosis viruses ofdifferentsubgroupsand typesisolated after passage of Roussarcomavirus-Rous-associated virus-0 incells fromdifferentring-necked pheasant embryos. J.Virol.19:302-312.
35. Tereba, A., L Skoog, and P. K. Vogt. 1975. RNA tumorvirusspecific sequences in nuclear DNA of sev-eral avianspecies.Virology65:524-534.
36. Verma, I. M., W. S. Mason, S. D. Drost, and D. Baltimore. 1974. DNApolymerase activityfromtwo
temperature-sensitivemutantsof Roussarcomavirus isthermolabile.Nature(London)251:27-31. 37. Vogt, P. K.,and R. R.Friis. 1971. Anavianleukosis
virus related to RSV(0): propertiesand evidence for helperactivity.Virology43:223-234.
38. Wang,S.Y.,W. S.Hayward,and H.Hanafusa.1977. Geneticvariation inthe RNAtranscriptsofendogenous virus genes in uninfected chicken cells. J. Virol. 24:64-73.
39. Weiss, R.A. 1969. The host range ofBryanstrain of Roussarcomavirussynthesizedinthe absence ofhelper virus. J. Gen. Virol. 5:511-528.
40. Weiss, R. A. 1973. Transmission of cellulargenetic ele-mentsbyRNAtumorviruses,p. 130-141. InL. Silvestri (ed.),Possibleepisomesineukaryotes.NorthHolland, Amsterdam.
41.Weiss, R.A., W. S. Mason,and P. K. Vogt. 1973. Geneticrecombinants andheterozygotesderived from endogenousand exogenous avian RNAtumorviruses. Virology62:535-552.
42.Weissbach, A.,A.Bolden,R.Muller,H.Hanafusa,
andT.Hanafusa.1972.Deoxyribonucleicacid polym-erase activities in normal and leukovirus-infected chickenembryocells. J.Virol. 10:321-327.
on November 10, 2019 by guest
http://jvi.asm.org/