0022-538X/79/02-0438/05$02.00/0
Recombination
of
Bacteriophage
pX174
by
the Red Function
of
Bacteriophage X
REIKICHIMUNEKIYO* ANDMUTSUO SEKIGUCHI
Department ofBiology, Faculty of Science, Kyushu University 33,Fukuoka812,Japan Received for publication1November 1978
Recombination ofbacteriophage 4X174 was effectively promoted when the Red function ofA wassupplied by either co-infection with A or induction ofA lysogens. Mutations in reda and redfl genesofAabolished recombinationnearly completely, whereasa mutation ingamgenereduced itonly slightly. The
Red-promoted recombination of 4X174 occurred in recA, recB, andpolAmutantsas
well as inwild-type strainsofEscherichia coli. Itwasfurtherstimulated when
OX174
mutantswereirradiated with UVlight before infection.Bacteriophage 4X174 and its
closely
relatedbacteriophage S13 are the smallest viruses to
undergo
genetic
recombination(11, 20),
andthemechanism has been analyzed extensively by
both genetic and physical methods. Recently,
"figure-8" molecules
consisting
oflinkedmono-mersof two double-stranded parental genomes
wereisolatedasrecombinationintermediates (4,
23). Most4X174 recombinantsareformed
by
amajor
pathway,
which requires the host recA+function (21, 22) but
apparently
does notneedanyof the known4X174geneproducts (3).
In the course of studies on genetic
recombi-nation of 4X174 under various
conditions,
wefound that recombination
frequencies
of4X174,
which are usually low compared to those of
other
organisms,
increasesignificantly
when thebacteria are co-infected with
bacteriophage
A.Subsequent
analysis
revealed thatrecombina-tion of 4X174 can be
promoted
by
the A Redfunction and that the contribution of the Red
pathway
in4X174recombination isgreaterthanthat of theRecpathway.
MATERIALS AND METHODS
Bacteria. The bacterial strains used in these ex-perimentsare listed inTable1. RY14was obtained by a crossofAB2470 (5) with KD15(+X')(10).
Bacteriophage. 4X174 wild type and its amber mutants, aml8, am86, am3, and am9, were obtained from R. L.Sinsheimer, and amNl was from M. Hay-ashi. aml8 and am86 carry an amber mutation in cistron A, am3 in cistron E, am9 in cistron G, and amNl incistron H (2).
AcI857bio69 and AcI857biol6 were provided by K. Shimada. The deletion of bio69 covers both int and redgenes, andthe deletion ofbiol6 covers only int gene(9).AcI857red314 (which is reda-),AcI857redll3 (redfl-),andAcI857gam210(gam-)were gifts from Y. Sakaki (14).
Media. Thecompositions of KC broth, starvation
buffer, bottom agar, and top agarwere asdescribed by Benbow et al. (2). To facilitate adsorption ofA, KC brothwassupplementedwith 0.01 MMgSO4and 0.2% maltose andwasdesignated KCA broth.
Crosses of 4X174mutants.Bacteria were grown inKCA brothto 108 cells per mlat370C with gentle aeration. The culture was made0.003 M KCN and aerated for 10 min. Two 0.5-ml samples of2 x 108 4X174 mutantphageper mlin KCA broth containing 0.003 M KCNweremixed inamating tube in an ice bath. Anappropriate concentration of Aphage suspen-sion(usually less than 0.1ml)wasaddedtothemating tube. A 0.2-ml amount of theKCN-treated bacterial culturewasaddedtoeachmatingtube and incubated at37°Cfor15min. Then the mixturewasdiluted 100-fold intoKCA broth and incubatedat370C with vig-orousaeration. After90min ofincubation,afewdrops of chloroform wereadded to theculture, and phage
titers were determined by platingwith Escherichia coli strain HF4714(fortotal number of
OX174
phage),strain C (for number of 4X174 wild-type recombi-nants), and strain C600 (for number of A phage). Though strains HF4714 and C were susceptible to both 4X174 andA, itwaspossibletotiter 4X174 in the presence of A since 4X174producedlargeplaqueseven afterashort timeof incubation(e.g.,3h),whereas A
didnot.InsomeexperimentsA-resistant derivatives of
strainsC and HF4714 isolated in thislaboratorywere
used,andessentially thesameresultswereobtained. Strain C600, a derivative ofK-12, was resistant to
4X174 infection and was used for titering A. Since C600 is deficientinrestrictionandmodification
func-tions,itwaspossibletotiterA grown inE.coliC. The recombination frequency between am mutants was definedasthe titer onstrainC divided by the titer on strain HF4714. Controlexperiments with one type of 4X174 mutant (selfimg) showed a frequency of less than lo-5wildtype per totalprogeny phage.
Crosses of4X174mutants in induced X lyso-gens. Thegrowth ofbacteria, treatment with KCN, and adsorption of4X174 were carried out at 330C,
whichprevented the induction of AcI857. Portions (0.1 ml) of themating mixture, without the addition of exogenousA,wereaddedto twotubes, each of which 438
on November 10, 2019 by guest
http://jvi.asm.org/
RED-PROMOTED RECOMBINATION OF OX174 439 TABLE 1. Bacterial strains
Strain Relevant characteristic Source and refer-ence C Wildtype, -X', Su- M. Hayashi HF4714 arg his leu thr pro -4X'Su' R. L. Sinsheimer
(3)
HF4712 recA13; isogenic to HF4714 R. L. Sinsheimer (3)
H560 polAI endoI thy str OX' R. L. Sinsheimer
Su- (3)
RY14 recB21 thr leu thi lac gal This laboratory aramtl pro his arg str
,XO Su+
C600 thr leu thi lac tonA oXr K.Matsubara Su+
contained 10ml of KCbroth;onetubewasincubated
at32°Candanother at40°C.Otherprocedureswere
essentiallythesame asdescribedabove.
Irradiation with UV. 4X174 phage were
sus-pended instarvation bufferat 109 phage perml and
irradiated with a 15 W Toshiba germicidal lamp to giveadose of 10J/m2 (about 10% survival).
RESULTS
Stimulation of 4X174 recombination by A co-infection. The growth of
OX174
is notsuppressedby co-infection withA.The burst size
of 4X174 in mixed infection with 4X174 and A
wasslightlyhigher than that in single infection, whereas the burst size of A was considerably reduced inthe mixedinfection. These situations allowedustoinvestigate theeffect ofAinfection
[image:2.505.255.448.78.194.2]on recombination of 4X174 in various host strains.
Table 2showsrecombination frequencies for severalambermutantscrossedpairwise inrecA+
and recA- cells withor without Ainfection. In eachcrosstherecombinationfrequencywas sig-nificantly lower in recA- cells than in recA+
cells,in agreementwithprevious results (21,22).
Infection withAcaused asignificant increase
in the recombination frequency in both recA+ and recA-cells. Almostthesamehigh levels of
recombinationwereattainedin thetwo typesof cells oninfection withA, and thusthe ratios of
recombination frequency in A-infected cells to that in noninfected cells were higher in recA-cells than inrecA+ cells.
The stimulation of
OX174
recombination by Ais furtherillustrated inFig.1.Withincreasing multiplicities of infection withA,recombinationfrequencies of 4X174 in recA, recB, andpolA
mutants as well as in the wild-type strain in-creased. Itseemsthatthehost functions suchas those controlled by the recA, recB, andpolA
genes are not required for the A-promoted
re-combination of
OX174.
However, there remains apossibility that exonucleaseV (the recB and -C gene product) and polymerase I (thepolAgene product) are concerned in the
recombina-TABLE 2. Stimulationof
OX174
recombination by A co-infectionaRecombinationfrequency (x10-4) on:
Crossb
recA+cells recA-cells
-A +A Ratio -A +A Ratio
am3(E)xam9(G) 4.2 57 13.6 0.56 41 73 am3(E)xamNl(H) 8.7 50 5.7 0.77 38 49 aml8(A)xam86(A) 12 -c - 0.84 59 70 am86(A)xamNI(H) 1.7 - - 0.12 10 83
am3(E)xaml8(A) 2.2 - - 0.24 17 71
'Crosses were performed as described in the text, using HF4714(recA+) and HF4712(recA-)as hosts. Forcoinfection, Aphagewasaddedtogiveamultiplicityof infection of5.
Inparentheses are the cistron for each mutant. -, Not tested.
100
50l f
0
O 5 10 15
MOI of A
FIG. 1. EffectofAinfectiononthe recombination
of fX174invarious hosts. Bacteriawereinfectedwith a mixture of4X174 mutants am3and am9 and
si-multaneously with Xc1857at various multiplicities.
Otherprocedures were asdescribed in the text. (0) HF4714(recA+);(0)HF4712(recA); (A)RY14(recB); (F) C(poLA,Su-);(U) H560(polA,Su-).
tionprocessin someindirect manner,since the
levels of recombination attained in X-infected
recB andpoLA strains are lower than those in
wild-type and recA strains.
Effects of A mutations on the 4X174
re-combination.
Phage
Xspecifies
twopathways
ofgenetic recombination: the Int
pathway
forsite-specific
recombination and the Redpath-way for
general
recombination(6, 17).
To seewhether
one or both of thesepathways
wereinvolved in the
promotion
of4X174recombina-tion, we
investigated
the effect of X mutationsonthe
4X174
recombination.First,
weexamined the effect of mutations ongeneral
recombina-tion. In thisexperiment
weusedXcI857,
whichproduces a
temperature-sensitive
repressor;VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
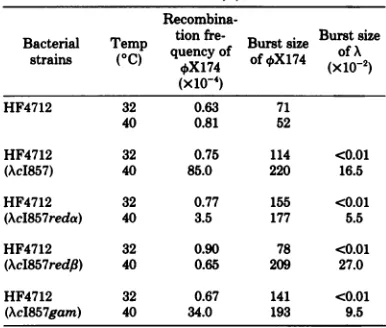
[image:2.505.266.436.200.431.2]thus, A gene functions are induced by shifting bacteria carrying AcI857tohigh temperature (1). The recombination of 4X174 was enhanced
when HF4712 (recA) cellslysogenic for AcI857
wereincubated at40°C (Table 3). No such en-hancement occurred whennonlysogenic bacteria
weretreated inasimilarmanner.Sincethe Red
pathway is controlled by three different genes,
reda,
redf3,
and perhapsgam (18), we nextex-amined which of the geneproductsare
respon-sible for the 4SX174 recombination. AAmutant havingamutation in theredfBgene wasunable
toenhance the 4SX174 recombination (Table 3). About 4 and 40% of the recombinationfrequency obtained by normal A was obtained with reda
andgam mutants,respectively. Second, we ex-amined the effectof absence oftheInt function. In thisexperiment, sinceaAInt-mutant cannot form a lysogen, A functions were provided by
infection. XcI857biol6, which deletesintbut car-ries redgenes, was able topromote 4X174
re-combination (Table 4), whereas AcI857bio69, which deletes both intand redgenes, wasnot.
These results suggest that recombination of
OX174
is carried out through the Red rather than theInt pathway.Effect of UV irradiation on the 4X174
recombination.Tessman(22) showedthat the
primary recombination pathway forS13,which isdependentontherecA+ function,iseffectively promoted by UV irradiation, whereas the recA-independent secondary pathway is not. It is of interest to see whether the Red-promoted
re-combination of 4X174 is stimulated by UV
ir-radiation.
[image:3.505.265.461.84.165.2]Table 5 shows the effect ofUV irradiationon the recombination frequencies of 4SX174under
TABLE 3. Effect oftheexpression ofXgenes onthe recombinationof4X174a
Recombina-Bacterial Temp tion fre- Burs Burst size
Bacterlluencofm Brt size
strains (°C) quencyof of tX174 of A (x 10-4)
HF4712 32 0.63 71
40 0.81 52
HF4712 32 0.75 114 <0.01
(AcI857) 40 85.0 220 16.5
HF4712 32 0.77 155 <0.01
(AcI857reda) 40 3.5 177 5.5
HF4712 32 0.90 78 <0.01
(AcI857red,B) 40 0.65 209 27.0
HF4712 32 0.67 141 <0.01
(AcI857gam) 40 34.0 193 9.5
aBacteria infected with am3and amNlwereincubatedat
32or400C.
TABLE 4. Recombination of4X174 in recA cells
infectedwith various A mutants' Recombination Burst size of Aphageb frequencyof
paX174
(x10-4)
"X174
None 0.53 325
AcI857 34 330
AcI857bio69A(int red) 0.63 330
AcI857biol6A(int) 57 300
aHF4712(recA)cellswereinfectedwith amixtureof0X174
mutants am3andamNl,togetherwithvarious A strains.The multiplicityofinfection forA was 5.
bA,Deletion.
TABLE 5. Effect ofUV irradiation on the three recombination mechanisms for Xl 74a
Recombination
fre-System quency(x10-4) Ratio +UV/ -UV
Bacteria A -UV +UV
HF4714(recA') - 8.8 39 4.4 HF4712(recA) - 0.52 0.49 0.94
HF4712 (recA) + 34 74 2.2
aBacteria were infected with a mixture of am3 and amN1 which either had or had not been irradiated with UV (10 J/
m2).
threedifferent
conditions,
eachof whichpermits
the
expression
ofoneof the three recombinationmechanisms.
Itwasfound that recombination of4X174
in recA+cells,
without Ainfection,
isstimulated
by
UVirradiation,
whereasnostim-ulationoccursin recA-
cells,
inagreementwiththe
result of
Tessman(22). The
Red-promoted
recombination
of4X174,
which could beseeninrecA-
cells
infected withA,
was stimulated byUV
irradiation, though
theextentof thestimu-lationwas
relatively
low. It has been shown thatUV irradiationincreases the recombination
fre-quencyfor
phage
A (7);thus,
theRed-promoted
recombination of
4X174
is similar to the Are-combination in thisrespectalso.
DISCUSSION
We haveshown that the Red function ofA can
promote the recombination of
4X174.
There-combinationfrequency of
4X174
increasedcon-siderably
afterinfection with A orinduction ofA
lysogenes,
and this increase was associated withexpression
of theARedfunction. TheRed-promoted
recombination of4X174
wasinde-pendent of recombinationby the hostRec
path-way, and the
efficiency
of the Red systemsur-passed
theefficiency
of the Rec system for4X174
recombination.Signer
and Weil(18)
investigated the effect ofred,int, andrecmutationsonrecombination of
Aandfound that the Redand theRecpathways
wereresponsibleforgeneral recombination ofX
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.268.458.230.297.2] [image:3.505.64.257.484.648.2]RED-PROMOTED RECOMBINATION OF
OX174
441whereas the Int pathway functioned for
site-specific
recombination. Table 6 summarizestheir
results
concerning the roles of the Red andthe Rec pathways in the X recombination and
also the present findings obtained with
cX174.
Actions of the two systems on A and
4SX174
recombination are essentially similar. It is of
interest that the ratio of the Red-promoted
re-combination to the Rec-promoted one is higher in
4X174
than inX.
Thisindicates that in spiteof theinherent low efficiency of the Rec system
for 4X174 recombination the Red system
pro-videshigh efficiency of recombination for both
4)X174
andX.
The A Red system has been shown to be
controlled
by three genes, reda,red,8,
andgam(17). The reda gene codes for X exonuclease,
which degrades DNA progressively from the 5'
end (8, 12), and the
redf3
genecodesfor,8
protein,which increases theaffinity ofthe exonuclease
for DNA (13). Thegam gene product has been
identified as aninhibitor of host RecBC enzyme,
whichis known to interfere with the A
recombi-nationreaction (14). The effect
of
thedeficiencyin these genes on A recombination has been
examined (16), and the result is
similar
to theresult of the present experiment with
4X174.
Inboth
phages,
reda and red/3 mutations abolishrecombinationcompletely or nearly completely,
whereasagam mutation reduces itonly slightly.
Thus, in thisrespect, too, the Red system acts
on
4X174
inasimilar mannerasonA.
4oX174
has asimple
genetic structure, and theentire nucleotide sequence of its DNA was
re-cently
determined(15).
This makes the virus anideal
organism
foranalysis
of the mechanisms ofrecombination and
replication
both in vivo andin vitro. The present
finding
mayprovide
an-other
useful
means forinvestigating
themolec-ular mechanism of the Red
recombination
path-wayand its interaction with other recombination
systems.
TABLE 6. Recombinationof
OX
74and Aby the Red andRecpathwaysRelative recombination fre-quency
Red Rec OX174a Ab
- - 1 <1
- + 7 36
+ - 72 83
+ + 100 100
aData taken fromTable2of thispaper:a crossof am3(E)andam9(G)mutants.
bDatataken fromSigner and Weil (18),a crossof
candRmutants.
ACKNOWLEDGMEENTIS
Wethank M. Hayashi, K.Matsubara,Y. Sakaki, K. Shi-mada,andR. L. Sinsheimerfor makingavailable to us the bacterialandphagestrainsused in this study. We also thank K.ShimadaandY. Sakaki for valuable suggestions.
LITERATURE CITED
1.Attardi, G.,S.Naono,J.Rouviere,F.Jacob,andF. Gros.1963.Productionof messenger RNAand regula-tion ofproteinsynthesis. Cold Spring Harbor Symp. Quant.Biol. 28:363-372.
2. Benbow,R.M., C. A. HutchisonIII,J. D.Fabricant, andR.L.Sinsheimer.1971.Genetic mapof bacterio-phage,X174.J.Virol.7:549-558.
3. Benbow,R.M.,A. J.Zuccarelli, G. C.Davis, andR. LSinsheimer.1974.Genetic recombinationin bacte-riophageOX174.J. Virol. 13:898-907.
4. Benbow,R.M.,A. J.Zuccarelli, and R. L. Sinshei-mer. 1975. Recombinant DNAmolecules of bacterio-phageOX174.Proc.Natl. Acad.Sci. U.S.A.72:235-239. 5. Clark,A.J., and A. D.MarguHies.1965.Isolation and characterization of recombination deficient mutants of EscherichiacoliK12. Proc.Natl. Acad. Sci. U.S.A.53: 451459.
6. Gottesman,M.E.,and R. A.Weisberg.1971.Prophage insertionandexcision, p. 113-138. In A. D.Hershey (ed.), Thebacteriophage lambda.ColdSpringHarbor Laboratory, ColdSpring Harbor,N.Y.
7. Jacob, F., andE.LWollman. 1955. Etudeg6n6tique d'unbacteriophage temper6d'Escherichia colim. Ef-fet durayonnement ultravioletsurla recombinaison g6netique.Ann.Inst.Pasteur 88:724-749.
8. Little,J. W. 1967. An exonuclease inducedby bacterio-phage A. II. Nature of the enzymicreaction. J. Biol. Chem.242:679-686.
9. Manly,K.F.,E. R.Signer,andC.M.Radding.1969. Nonessentialfunctions ofbacteriophage A. Virology37: 177-188.
10. Ono,M.,M.Kuwano,and T. Horiuchi. 1977. Genetic analysisof mutationsaffectingribonuclease fl in E8ch-erichiacoli.Mol. Gen. Genet. 153:1-4.
11.Pfeifer,D.1961.Genetic recombinationinbacteriophage *X174.Nature(London) 189:422-423.
12. Radding, C. M. 1966. Regulation of A exonuclease. I. Propertiesof Aexonucleasepurifiedfromlysogensof A
T,nandwild-type. J. Mol. Biol.18:235-250.
13. Radding,C.M., andD. M. Carter.1971. The role of exonuclease and,B proteinofphageA ingenetic recom-bination.mI.Bindingtodeoxyribonucleic acid.J.Biol. Chem. 246:2513-2518.
14. Sakaki, Y.,A. E.Karu,S.Linn,and H.Echols. 1973. Purification andpropertiesof they-proteinspecified by bacteriophageA:aninhibitorof the hostRecBC recom-bination enzyme. Proc. Natl. Acad. Sci. U.S.A. 70: 2215-2219.
15. Sanger,F.,G. M.Air,B.G.Barrell,N. LBrown,A. R.Coulson,J.C.Fiddes,C. A. HutchisonIm,P. M. Slocombe,and M.Smith.1977.Nucleotidesequence ofbacteriophage4X174DNA.Nature (London) 265: 687-695.
16.Shulman,M.J., L.M.Hallick,H.Echols,andE. R. Signer. 1970. Properties of recombination-deficient mutantsofbacteriophageA. J. Mol.Biol. 52:501-520. 17. Signer,E.1971.Generalrecombination,p.139-174.In A. D. Hershey, (ed.), The bacteriophage lambda. Cold SpringHarborLaboratory,ColdSpring Harbor,N.Y. 18. Signer,E.R.,and J. Weil. 1968.Site-specific recombi-nation inbacteriophageA. ColdSpringHarborSymp. Quant. Biol.33:715-719.
19. Sinsheimer, R.L. 1959.Purification andproperties of VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
bacteriophageOX174.J. Mol.Biol. 1:37-42.
20. Tessman, E.S., andI.Tessman. 1959. Genetic recom-bination inphage S13. Virology7:465467.
21. Tessman, I. 1966. Genetic recombination of phageS13 in
arecombination-deficientmutantofEscherichia coli K12.Biochem. Biophys. Res. Commun. 22:169-174. 22. Tessman, L. 1968. Selective stimulation ofone of the
mechanisms forgenetic recombination of bacteriophage S13. Science 161:481-482.
23. Thompson, B. J., C. Escarmis, B. Parker, W. C. Sla-ter, J. Doniger, I. Tessman, and R. C.Warner.
1975.Figure-8 configuration of dimersof S13 and 4X174
replicative form DNA. J. Mol. Biol.91:409419. 24. Zissler, J., E. Signer, and F. Schaefer.1971.Therole
ofrecombination in growth of bacteriophage lambda.I. Thegamma gene, p.455-468.InA.D. Hershey (ed.), Thebacteriophage lambda. Cold Spring Harbor
Labo-ratory,Cold Spring Harbor, N.Y.