Copyright01976 American Society forMicrobiology Printed in U.S.A.
Effect
of the "RNA Control" Locus in Escherichia coli on RNA
Bacteriophage R23 Replication
JAN ERNBERG* AND OLA
SKOLD
Department of Microbiology, BiomedicalCenter, University of Uppsala, Uppsala, Sweden
Received for publication 7 August 1975
The effect of the rel gene of Escherichia coli on the
RNA synthesis inducedby
phage R23 was studied. This RNA phage has the property of inhibiting ribosomal
RNA
formation and completely dominating the RNA synthesis of the host.Phage-specific RNA
formation was found to be dependent on the allelic state ofthe
rel
gene. Determinations of RNA synthesis were made by both cumulativeand
short-term
incorporations of uracil and adenine. Variations in labeling of nucleotide pools were compensated for by determining specific activities of ATPand UTP and using these values
toobtain
true,relative
rates of RNA synthesis.Rapid synthesis
of RNA inbacteria
isde-pendent
onthe
presence of aminoacids. Amino
acid
regulation of RNA synthesis
is awell-established phenomenon and has been
shown tobe
governed by the rel locus
ofEscherichia coli
(for review see reference 5).
Full synthesis
ofRNA is
dependent
notonly
onthe
presence offreeamino
acids but also
on afull
complementof
aminoacyl-tRNA
(15). During amino acid (oraminoacyl-tRNA)
deprivation in arel+
strain, rRNAsynthesis
israpidly curtailed
and acon-comitant
accumulation
ofthe
two guanosinenucleotides
ppGpp and
pppGpp
isobserved
(11).
Under similar conditions
in areh
strain,
rRNA
synthesis
continues
unabated,
andneither
ppGpp
norpppGpp
isformed.
The mechanism
ofaminoacid regulation
as itworks
atthe
transcriptional level
in vivo isunknown,
although
adirect
effect on theactiv-ity of the
DNA-dependent RNA polymerase
would
be conceivable.
To testsuch
anidea
itwould
be
of interest to investigatewhether
therel
gene,which
isdirectly linked
toppGpp
synthesis
inE.
coli, also
governsthe
RNAsynthesis that
iseffected
by
bacteriophage-specific,
RNA-dependent RNA
polymerase.
Anideal
tool forthis investigation would be
anRNA
phage with the
ability
to turn offribo-somal
RNA
synthesis, because
insuch
asystemphage-specific
RNAsynthesis
couldeasily
beseparated
fromhost-specific
RNAsynthesis.
Phage
R23, which resembles f2 inchemical,
physical,
andimmunological characteristics,
hasbeenreported to dominate the RNA
synthe-sis ofthe host cell
completely (22).
In the present
investigation
phage
R23 wasshown
by electrophoretic
analysis
tobe able toturn off
completely the
rRNAsynthesis
ofthehost. The
formation of R23 phage RNA wasfurthermore found
tobe dependent
onthe
presence of free amino
acids.
This stringentregulation
ofphage
RNA synthesis
wasdepend-ent on
the allelic
stateofthe
rel locus
ofthe
hostand
wasshown
tobe absent
in arel
strain.MATERIALS AND METHODS
Phage and bacteria. The strains of E. coli K-12, Cp78
(rel+)
and Cp83(rel ), were obtained from N. Fiil.They were originally derived from E. coli K-12 W677, are F-, require arginine, histidine, leucine, threonine, and thiamine, and are isogenic except at therel locus (6). The described strains were made F+ andsusceptibletoRNAphage infectionby incubation with aprototrophic male strain of E. coli K-12. The maledonorwasmixedwith theauxotrophic F- strain inequal proportions at a total cell concentration of about0.5 x 109/ml in arich medium (supplemented LBofNordstrometal. [17 ]). After incubationat37C for 60 min without shaking, the cell mixture was plated and the small coloniesofauxotrophic bacteria werepicked and testedforthe male propertybyRNA phagesusceptibility. The relaxed and stringent RNA synthesis patternsforthe male strainsconstructedin this wayare shown in Fig. 1, where the rep pheno-typeisclearly discernibleby the cumulative incorpo-ration ofradioactive adenineintoacid-insoluble ma-terial. E. coliQ13(10)wasusedroutinelyforthe assay ofphage. RNA bacteriophageR23 (22) was supplied byJ. T.August. Phagestocks wereprepared accord-ingtoWatanabeand August (21).Chemicals. [2-'4C]uracil(60or22mCi/mmol)and
[8-14C
]adenine
(60 mCi/mmol) were purchasedfromThe Radiochemical Centre, Amersham, England, or New England Nuclear Chemicals, ]Dreieichenhain, Germany.
[5-3H]uracil
(20Ci/mmol) and[2-3Hlade-nine (18 Ci/mmol) were obtained from The
Radio-chemicalCentre. [3PTjorthophosphate waspurchased 307
on November 10, 2019 by guest
http://jvi.asm.org/
308 ERNBERG AND SKOLD
40
30
m
2J
i-)
20
0
^ -ao +aa
aeration
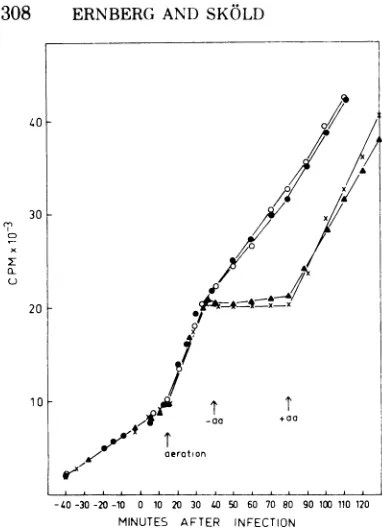
-40-30-20 -10 0 10 20 30 40 50 60 70 80 90 100 110 120 MINUTES AFTER INFECTION
FIG. 1. Incorporation of [14C]adenine into
acid-insoluble product underamino acidstarvationofthe used F+ strains with andwithoutphageR23infection. At -60min, cellsweredilutedintothelow-phosphate mineral salts medium supplemented with
["4Clade-nine (0.2
ACi/ml,
6 vg/ml) and incubated at 37C without aeration for 60 min. At time zero, viablecount determination (1.5 x 108 cells/ml) andphage
infection took place. Aeration wasstartedat15min.
Amino acidswereremovedby filtrationat40minand added back at 80 min. Samples of 0.1 ml were
withdrawn and precipitated onfilterpaper disks as
described in the text. Values along the ordinate
represent countsperminute incorporatedper0.1-ml
sample. Symbols: x, Cp78F+; A, Cp78F+ + R23; 0, Cp83F+;0,Cp83F++R23.
from AB Atomenergi, Studsvik, Sweden. Amino acids, thiamine, uracil, and adenine were obtained
from Nutritional Biochemicals Corp., Cleveland, Ohio. Polyethylene glycol (A 6000) and sodium
dex-tran sulfate (500) for phage purification were
pur-chased from Union Carbide Corp., New York, and Pharmacia, Uppsala, Sweden, respectively. Acrylam-ide (catalogue no. 5521, for electrophoresis), N,N'-methylenebisacrylamide, and NN,N',N'-tetramethyl-ethylenediamine were the products of Eastman
Kodak Co., Rochester, N.Y. Agarose was purchased
from Miles-Seravac, Maidenhead, Berks., England. Cellulose thin-layer plates (cellulose polyethylenei-mine MNPolygram cellulose300PEI) wereobtained
fromMachery Nagel and Co.,Buren, West Germany. The plateswere soaked for30 min indistilled water
and dried beforeuse.
Media. Most experiments were carried out in a
low-phosphate mineral saltsmedium ofthefollowing composition (per liter): 0.5 g of NHC1, 0.3 g of (NH4)2SO4, 5.0 g of NaCl, 0.5 g ofKCI, 0.203 g of
MgCl2*6H20, 14.7 mg of CaCl2 2H20, 0.178 g of
Na2HPO4 2H2O, and 6.25gof Tris. pHwasadjusted to 8.0 with 1 M HCl and, after autoclaving. the medium was supplemented with 0.9 ml of 1 M
CaClI2/liter, glucoseto5g/liter, and
FeCl,
to 10-5M.Supplements were aminoacids (50gg/ml) and thia-mine (1
pg/ml).
MediaforplatingofphageR23were asdescribed by Watanabe and August (21).Infected cells. Anovernight culturewasdiluted to about1.5 x 101cells/mlinthelow-phosphate mineral saltsmediumsupplementedasnecessary.The culture was then left for 60 min at 37C without aeration before the addition of phage. This procedure was found to give good and reproducible phage produc-tion. At time zero phage was added to 20 to 30 PFU/bacterium. Aeration wasstartedat 15 min after infection. Amino acid starvation was initiated by filtration on a membrane filter (Gelman, Metricel GA-6) and resuspension in amino acid-deficient me-dium. Phage production was determined routinely and amounted to 1,200 to 1,800 phages/bacterium. This burstsize concurswith that reported byLodish and Zinder(14) forphagef2 inE. coli K-12.
Determination of RNA synthesis. The synthesis ofRNA was measured by thecumulative incorpora-tion of [14C]uracilor
[14C]adenine
intoacid-insoluble product. Determination of RNA synthesis was per-formed either by precipitating a 1-mlsample ofthe labeled culture in5 ml ofcold5%trichloroaceticacid and collecting theprecipitate on aglassfilter(What-man GF/C) or by pipetting a 0.1-ml sample onto a
2-cm-diameter filterpaperdisk, whichwas immedi-ately immersed intocold 5% trichloroacetic acid (1). Filters were dried and then counted in a Packard scintillation counter.
Determination of the relative rate of RNA synthesis. Relative rates of RNA synthesis were determined mainly as described by Winslow and Lazzarini(24). Tomeasuretherelativespecific activ-ity of theATPand UTP pools, bacteriawere labeled for several generations with ["P ]orthophosphate (10 to 15
ACi/ml,
10 to 15Ci/Mmol).Labeled bacteriawere diluted into low-phosphate mineral salts medium (containing([32P]orthophosphateatthesame specific activity [10 to 15pCi/,pmoll)
and incubatedat 37 C for60min without aeration, to yield about 1.5 x 108cells/ml (cf. infected cells above andFig. 1).Thecells
were theninfected with R23, and aeration was started 15minlater. Atindicated intervals
500-gl
samples of the culture were withdrawn and added to vials containing either [2-_H]adenine (final concentration 20uCi/ml, 20mCi/pmol)
or [5-3H]uracil(20pCi/ml,
20mCi/Amol).
After 60 s of incubation at 37 C under aeration, a
100-gA sample waswithdrawn and pipetted into 50
pl
of4 M formic acid in an ice bath. After 30 min of extraction at 0C,samples were centrifuged for 15min at 10,000 x g, and 10
Al
of the supernatant was applied to a polyethyleneimine cellulose thin-layer plate.Asreferences, UTP and ATP(0.1 mol of each) TJ. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.491.44.235.48.313.2]AMINO ACID REGULATION OF PHAGE SYNTHESIS were added. The chromatograms were run up twice
with water before development in 0.75 M KH2PO4, pH3.4(3).
The ATP and UTP spots were located under UV or byautoradiography, cut out, and placed in a scintilla-tion vial. Thenucleotide was then extracted with 250 Mlof 2 M NH40Hfor 20minand finally counted in5 ml ofAquasol (New England Nuclear Corp., Boston, Mass.) scintillation liquid containing 5% acetic acid and 8% water to decrease chemiluminescence. The settings ofthe scintillation spectrometer used gave 7% spillover of 32P counts into the 3H channel.
Determinations of guanosine tetraphosphate were performed in asimilar way and mainly as described earlier (3).
The rate of incorporation of [3H
]adenine
or [3H uracil into RNA was determined in cultures treated exactly asdescribed above except that they were not labeled with 32P. At intervals of 15 s, 50-ul samples were removed from the 500-julculture sam-ples and pipetted into 2.0 ml of 0.3 M NaOH in an ice bath. One halfofeach sample wasthen neutralized with 0.3 M HCl and precipitated with 5% trichloro-acetic acid. The other half of each sample was incubated at 37C for 12h, cooled, neutralized with 0.3MHCl, and finally precipitated with 5% acid. The precipitates were collected onmembrane filters (Sar-torius, Gdttingen, Germany), which were dried and then counted inthe scintillation counter. Thecounts remaining after alkaline hydrolysis were subtracted from total trichloroaceticacid-precipitable countsto give incorporation into RNA. This value was then dividedbytherelative specific activity(3H/32P ratio), at the corresponding time interval, ofthe ATP and UTP pools, respectively, to give the relative rate of RNAsynthesis.Extraction of RNA. RNA for electrophoretic anal-ysiswasextractedfrom5-mlsamples withdrawnfrom the aerated, phage-infected cultures. The infected bacteria were rapidly centrifuged, resuspended in 3 ml of electrophoresis buffer, quickly frozen, and
disintegrated in a high-pressure cell (X-press, AB
Biotech, Stockholm, Sweden). To the disintegrated materialwas added0.1 mlof abentonite suspension
(30mg/ml), fractionated according to
Fraenkel-Con-rat et al. (8). Afterthawing, the suspension was cen-trifuged for 15 min at 0C and 20,000 x g. To the supernatantwasadded0.1mlofbentonitesuspension
(30 mg/ml) and 0.07 ml ofcarrier RNA solution (2
mg/ml) prepared from uninfected E. coli Cp78 F-.
RNA wasthen extracted by the method ofPeacock andDingman (18)forcytoplasmicRNA.After treat-ment with 0.1% sodium dodecyl sulfatefor 4 min at room temperature, the supernatantwas mixedfor 15 minwithanequal volumeofwater-saturated phenol. Sodium acetate was added to the mixture to a final concentration of 0.3 M. Theseparated phenolphase was reextracted for 5 min with 0.5 ml of electro-phoresis buffer. The combined water phaseswere re-extracted once with 0.5volume ofphenolfor 5 min, and RNAwasfinally precipitatedfromthe
separated
waterphase with2volumesofcold ethanol at -20C overnight. Aftercentrifugation, the
precipitated
RNAwas washed as described by Peacock and Dingman (19) and finally dissolved in 0.5 ml ofelectrophoresis buffer. RNA concentration was determined by UV absorption, using an extinction coefficient at 260 nm of0.24 for a 0.001% RNA concentration.
Acrylamide gel electrophoresis. The procedure forthe preparation of the gels and theelectrophoretic techniques were mainly those of Peacock and Ding-man (18). The electrophoresis apparatus used was that originally described by Davis (4). A composite gel of 0.5% agarose and 2.5% acrylamide was used throughout. The electrophoresis buffer was Tris-chlo-ride (0.025 M), pH8.0-MgCl2 (0.001 M). All gels were prerun for1h at 9mA/tube, the gels were cooled, and the buffer was changed before application of the samples, which consisted of 10MlofRNA(about 0.5 mg/ml) and20Mlof 24%glycerol, with added bromo-phenol blue as a dye marker. All samples were electrophoresed for 2 h at 9mA/tube. After electro-phoresis the gelsweresoakedin1 M acetic acid for15 min and thenstained for20 min in 0.2% methylene blue in 0.4 M sodium acetate buffer, pH 4.7. The excess stain was removed by rinsing in water. To measure radioactivity, the gels were cut into slices, 1.40 mm thick, each ofwhich was transferred to a scintillation vial and treated with 0.5 ml of0.5 M NHOH for2h at 80C. The samples were then dried andfinally counted inthescintillationcounter after theadditionof 10mlofnaphthalene dioxane scintilla-tionfluid.
RESULTS
Growth characteristics
forphage
R23 inthe bacterial strains used. Production
ofR23in
the
stringent male strainCp78
F+ isdemon-strated in
Fig.
2A. At 37 C and in thelow-phos-phate
mineralsalts
medium used, mature phage began to appear intracellularly at 50 to 60minafter infection. Aburst size of 1,800
phages/
bacterium
was reached within 120 min. Theinability
oftheoriginal
female strain Cp78 F- tosupport phage growth is demonstrated in the
same
figure
forcomparison. Phage
R23 had theability
tokill its host. This is shown inFig. 2B,where the
exponential killing
of the male strainsof
Cp78
andCp83
is demonstrated. Theper-centage ofsurviving bacteriawas 50% at 25 to30
min after infection and decreased to 5% at 120
min. It is shown forcomparison that the female
strain of
Cp78
wascompletely
unaffectedby
phage.
Interference
of R23
withhost
RNA
synthe-sis. The interference of
phage
R23 with hostgrowth
isalso reflected in the curvesrepresent-ing RNA
synthesis (Fig. 3).
The upper curvedemonstratesRNA
synthesis
afterthe infectionof
Cp78
F-with R23. The cumulativeincorpora-tion of
[4C
]uracil
increasedexponentially
inthe
growing,
phage-insensitive
cells. In thesensitive
Cp78
F+cells, however,
RNAsynthe-309
VOL.17, 19X6on November 10, 2019 by guest
http://jvi.asm.org/
310
ERNBERG ANDSKOLD
U
:1
/;
m 1400
-5-.
o -1000
cr ~~~0.505,
2 100010 20
0 0~~~~~~1
600~~~~~~~~~~~~00
20 40 60 80 100 120 20 40 60 80 100 120
MINUTES AFTER INFECTION
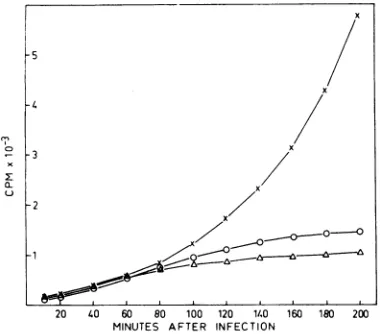
FIG. 2. Growth characteristicsofR23 inCp78F+, Cp78 F-,andCp83 F+. (A)Cellsweregrownandinfected at 0.5x 106 cells/ml in thelow-phosphatemineral salts mediumasdescribed in thetext.At the timesindicated,
0.5-misampleswerewithdrawn andlysedasdescribed(22).Intracellularphage production expressedasphages
perbacteriumwasplottedversustime.Symbols:0,Cp78 F+;0,Cp78 F-. (B) Cellsweregrown andinfected as in (A) butat2.5 x 108cells/ml. At timesindicated, samples werewithdrawn and titrated by viable count of bacteria. Thefraction ofsurvivingbacteria isplottedversustimeon asemilog scale. Symbols:0, Cp78 F+, x, Cp78 F-; A, Cp83 F.
x
-5
-4
-3
-2
1
-8-
- '1..
20 4I 0 8 0 2 4 6 8 0
20 40 60 80 100 120 ll.0 160 1B0 200
MINUTES AFTER INFECTION
FIG. 3. Synthesis of RNA in Cp78 F+ after R23 phage infection. Cellsweregrownandinfectedat0.5 x 108 cells/ml in the low-phosphate mineral salts medium supplemented with ["4CJuracil (0.1
IACi/ml,
10 ug/ml). Samples of0.1 ml were withdrawn andprecipitatedonfilterpaperdisks asdescribedinthe
text. Values along the ordinaterepresent countsper
minute incorporatedper0.1-ml sample.Symbols: x,
Cp78F-;0,Cp78F+;A,Cp78F++ chloramphenicol
(20
,ug/ml)
addedat5minafterinfection.sisdiminishedbutnotuntil about80minafter
infection, i.e., ata time when80 to 90% ofthe
cellswerealready killed by phage (cf. Fig. 2B).
Thelowercurvefor comparison shows the RNA
synthesis in R23-infected Cp78 F+ cells, in
which protein synthesis was inhibited by the
addition
ofchloramphenicol
at 5 minafter
infection and
the
phage could thus
notestab-lish its
RNA synthesis. The middle
curve,demonstrating RNA
synthesis after normal
phage
infection,then
seems toshow that
what-ever amountof
host-specific RNA
synthesis
wasturned
offby phage, during the first
80min ofinfection,
wassubstituted
by
phage-specific
RNA
synthesis,
because
the cumulative
[I4C]uracil
incorporation
inthat
timeperiod
was
identical
inthe F- and F+ strains.
The
fraction
ofribosomal and phage-specific
RNA
synthesis, respectively,
wasdetermined
by electrophoretic analysis
onpolyacrylamide
gels. The RNA synthesis
patternobtained
inthis
way forF+
and
F-bacteria
isshown
inFig.
4.
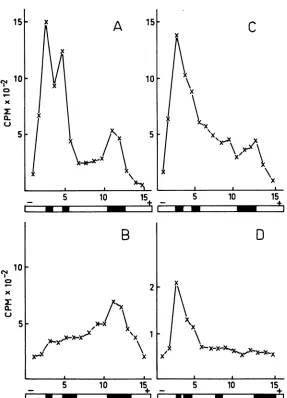
When
[04C]uracil
wasadded
toCP78 F- cells
at 40 min after
the addition
ofphage, the
incorporation
ofradioactivity between
40and
60min was
predominantly,
intoribosomal 16S and
23S
RNA (Fig. 4A). In thecorrespondingexperi-ment
with the
F+ strain, the typical rRNAlabeling
pattern wasabolished
(Fig. 4B), andinstead
a more diffuse labeling pattern oc-curred.A new
peak,
running in front ofthe
23S
marker,
wasobserved
when the[04C
Juracil
incorporation took
place between
30and 90 minafter infection
(Fig. 4C) and ought
torepresentR23
phage-specific RNA,
since it partlycoin-cided with labeled
RNA extracted frompuri-fied,
maturephage (Fig. 4D).
Effect
of
the rel locus of the host onR23-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.491.102.388.66.239.2] [image:4.491.49.239.319.486.2]AMINOACID REGULATION OF RNAPHAGE SYNTHESIS
A
15F
C
10oF
5 10
15+
5 10 15.
=_ nmow-
I I5 10
15
I
5
-M
10 15
[image:5.491.110.397.62.460.2]~~FI
FIG. 4. Electrophoretic analysis of hostandphageRNAsynthesisonpolyacrylamide gels. Cp78F- andCp78 F+ cellsweregrownand infected at2.5 x 108 cells/mlin the low-phosphate mineral saltsmedium to which uracil(2
,ug/ml)
wasaddedat20minand [14C]uracil(0.1O Ci/ml) wasaddedatdifferenttimesafter infection. Samples of5 ml werewithdrawn, RNA extracted, andsubjected toelectrophoresisasdescribed in the text.LabeledR23 RNAwasprepared from phagegrownin E. coli K38(21)inthepresenceof[14C]uracil(0.1 iCi/ml, 2 ig/ml)andpurifiedasdescribed. Thelabeledphage suspensionwasmixed with unlabeledphage beforeRNA
extraction. Thefinal activitywasabout 500counts/minper101IPFUof phage.Extraction andelectrophoresis ofRNAwasperformedasdescribedfortheinfectedcells. The valuesalongthe ordinatesrepresentradioactivity
pergel slice,the numbersofwhichareplotted alongtheabscissae. Below eachframethepositions ofthe stained
bandsareshown.GelsA, B,andCwere runfor2h,andgelDwas runfor4h.(A) Cp78 F-,labeledfor40to60 min after infection; (B) Cp78 F+,labeledfor40to60minafter infection; (C) Cp78 F',labeledfor30to90min after infection; (D)R23RNA,runtogetherwith unlabeled RNAfrom uninfected Cp78F+.
specific RNA synthesis. The effect of amino
acid starvation on the RNA synthesis in Cp78 F+ and Cp83 F+ infected with phage R23 is shown in Fig. 5. In the stringent strain,
[I4C]uracil,
when added at 3 min after amino acid removal (40 min after infection), was tosome extent incorporated into acid-insoluble material. Thisincorporationtookplaceatarate thatwasonlyabout 2.5% of that obtained after readdition of amino acids. When the identical
experiment was performed with [14CJadenine
as the radioactive precursor, the rate of in-"IC-,
x
v 10
CD
x
CL
B
2 X
W-. . .
- - - - +
__
_
I
I
311 VOL. 17,1976
5
[
on November 10, 2019 by guest
http://jvi.asm.org/
312
ERNBERG AND SKOLD11
-10
T7
y6
0-u
40 50 60 70 80 90 100 110 MINUTES AFTER INFECTION
FIG. 5. Incorporationof
[14C]uracil
or['4C]adenine
into the RNA ofstringent and relaxed host strains after R23phageinfection. CellsofCp78 F+, Cp78
F-,
andCp83F+ weregrownandinfectedasdescribed in Fig. 1. Amino acidswereremoved byfiltrationat40 min after infection, and 3 min later [14C]uracil or
[14C]adenine
(0.1 uCi/ml, 4 Ag/ml) was added. RNA was determinedin 1-ml samples as described in the text. Amino acids were added back at80 min after infection.Sym bols: x,Cp78F+ +R23,[I4C
juracil;
A, Cp78F+uninfected, [14C Jadenine;*,Cp78 F'+R23,[l4Cladenine;
I,
Cp83' + R23,[14Cjuracil;
0,Cp83F-l +R23, [14Cadenine.
corporation without amino acids was about
[image:6.491.47.239.51.328.2]13%
ofthat
seen after the end of starvation.Figure 5 also
describes
similar experiments withCp83 F+, where
noeffect ofamino acidstarva-tion on either uracil or adenine
incorporation
wasobservable. The experiments of Fig. 5 then
seemtoshowthat at 40 minafter R23infection,
whenaccording to Fig. 4 most RNA synthesis is
phage specific, amino acid deprivation of the
stringent strain severelyrestricted the
incorpo-rationofbothuracil andadenine into RNA. The
demonstrateddifference in uptake between
ura-cil and adenine agrees with
observations
byLazzarini and Dahlberg (13), who observed the
uptake of adenine into ATP to be much less affected than that of uracil into UTP under
stringent conditions. The restriction of
radioac-tive precursor incorporation into R23 RNA
shown in Fig. 5
could be
partly
explained by
apossible
labilization
ofphage
RNA in theab-sence of coat protein
synthesis during
aminoacid
starvation. This wouldresult
inreplenish-ing of
nucleotide
pools
fromphage RNA
turn-over
and
a consequentblock
in exogenousuptake in
the absence
ofnucleotide
pool
expan-sion(16). To testthis
possibility,
anexperiment
was performed with starvation only for
histi-dine. This amino acid is
absent
from R23 coatprotein, the
synthesis
ofwhich will thusproceed
inits
absence
(22). Starvation foronlyhistidine,
starting at 40 min after infection, almost
com-pletely
inhibited
incorporation ofradioactive
uracil
intophage
RNA, with
a pattern verysimilar
tothat
obtainedwith
starvation forother amino acids
(Fig.
6).Determinations
ofguanosine
tetraphosphate
in
the
stringent strainCp78
F+with and
with-out
phage
infectionand with
aminoacid
starva-tion asdescribed
in Fig. 5 showed arapid
increase in
ppGpp
in both cases. The rise was,however, repeatedly found to be somewhat
slower and to reach a slightly lower maximum
level in the
infected
than in theuninfected
bacteria.
In oneexperiment theppGpp
concen-tration in
uninfected
Cp78
F+ thusincreased
from 50 to 600
pmol
per 108 cellsduring
the first5min ofaminoacid starvationto
reach
afigureof733 after
about
20min andthen
decreased
to-2
AA
20 30 40 50 60 70 90 90 100 110 120 130
MINUTES AFTER INFECTION
FIG. 6. Incorporation of ['4C]uracil into RNA of R23-infected stringentandrelaxedhoststrainsduring histidine starvation. Cells ofCp78 F+, Cp78 F-, and Cp83F+ weregrown and infectedat 0.5 x 108/mlin the low-phosphate mineral salts medium. Histidine wasremoved byfiltration at 40 min afterinfection, and 3 min later [14C]uracil(0.1
ACi/ml,
2jg/ml)
was added. RNA was determined in 0.1-ml samples as described in the text. Histidine was added back at 80 minafter infection. Values along theordinatedenote counts per minute per 0.1-ml sample. Symbols: 0,Cp78F+; A, Cp83F", x, Cp78F-.
J VIROL
&I
T
x:
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.491.256.442.381.535.2]AMINOACID REGULATION OF RNAPHAGE SYNTHESIS 313
717at40min. In R23-infected Cp78F+,ppGpp
rosefrom 50to 417 pmol per 108cellsin 5min
and to550 after 20
min,
and finally reached avalue of 617 after40min ofaminoacid
starva-tion. The dramatic changes induced by amino
acid starvation in theratesofRNAsynthesisas
measured in the
described
experimentscould bedue tothe primary restriction of the uptake of
uracil and adenine, respectively, intoprecursor
nucleotide pools. To investigate this possibility,
the relative specific activity of the UTP and
ATP pools was
determined
under differentla-beling
conditions.
Cells were exposed to[32P]phosphate for several generations and then
labeled during short times with [3(H adenineor
[3H
]uracil. In Table1areshownthe fractions ofthe total UTP and ATP pools labeled during
1-min pulses of theexogenous precursors, given
atdifferent timesafter aminoacidremoval from
the stringent Cp78 F+, with and without phage
infection. In an
uninfected
culture theentry ofexogenous
[3H
Juracil
intotheintracellular UTPpool wasreduced from 23% toabout 1% during
amino acid starvation. After readdition of
amino acids the uptake rapidly rose
several-fold, although the
prestarvation
level wasap-proached only slowly. A similar pattern was
seen in an infected culture where the uptake
decreased somewhatmore slowly from 20.5%to
about 1.5% under amino acidstarvation. When
the above experiments were repeated with
[3'H]adenine as a precursor, in an uninfected
culture the adenine uptake into the ATP pool
decreased from 50%toabout 20%during
starva-tionand thenrapidly returnedtothe
prestarva-tion value at readdition of amino acids (Table
1). In the infected culture the [3H
]adenine
uptake was lessrestricted during starvation. It
decreased from 63% toabout35%. Table 1thus
demonstrates that the incorporation of both
uraciland adenine into theirrespective
nucleo-tide pools is restricted during amino acid
star-vation, butslightly lesssoinR23-infectedthan
inuninfected host bacteria, and much lesssofor
adenine thanforuracil. Thepool labeling
frac-tions of Table 1 could then be used tofurther
test the dramatic changes in RNA synthesis
observed in cumulative incorporation
experi-mentslikethosedescribedinFig.5 and 6. To do
this, rates of RNA
synthesis
were measuredunderdifferent conditions by short-term
incor-porations of radioactiveprecursor. Theeffectof
aminoacid starvationon 75-sincorporationsof
[3H
]adenine into RNA of the stringent strainCp78F+ isshown inFig.7A.During starvation,
the labeling rapidly decreased with time after
amino acid removal.The same general pattern
TABLE 1. Uptake of[3H]uraciland
[3HJadenine
into the UTP and ATP, respectively, of Cp78 F+ during amino acid starvation and with and without phageinfectiona
['HJuracil [3H]adenine (pmolof pmolof
Cells ['HJUTP)/ ['H]ATP/
Min" (pmolof Min (pmolof
[3P]UTP) [32P]ATP)
x100C x10
Uninfected 30 23 30 50
43 1.9 43 38
58 0.5 58 17
76 0.9 76 19.5
88 7.7 88 54
110 17.7 110 53
Infected 30 20.5 30 63
43 5.4 43 45
58 1.5 58 33
76 1.2 76 36
88 7.7 88 42
110 13.6 110 49
aCellswere grown and infectedinthe presenceof
[3"P]-phosphate
asdescribedinthetext.°Labeling with [3H
Juracil
or['H
]adenine
tookplace during 1-min periods starting at the indicated times. Amino acids were removed at 40 min after infection (or start ofexperiment) and added back at 80 min.
cMolar amounts of 32p- and
'H-labeled
nucleotidcswere calculated from samples of known concentra-tions added tothin-layer plates asdescribed in the text.
was seen after
R23
phage
infection(Fig. 7B),
although
inthis
casethe decrease
was lesspronounced. In the relaxed
strainCp83
F+,
infected with R23, the short-term
incorpora-tions were
completely
unaffected
by
amino
acids starvation
(Fig. 7C).
The data of
Fig.
7 werecombined
with the
specific activities,
presented
inTable
1 forthe
ATP
pool
atcorresponding
times,
togive the
relative
rates ofRNA
synthesis.
The
resulting,
corrected
rates ofRNA
synthesis
areshown
inFig.
8forCp78
F+during amino acid starvation
and with and without R23
phage
infection. The
infected
and noninfected cultures,
respectively,
ofthe stringent
strainshowed
a verysimilar,
stringent response to amino acid
starvation,
which
indicatesthat
thephage-specific
RNA-synthesis
was also under thecontrol
ofthe rel gene of the host.DISCUSSION
The aim of these
experiments
was tostudy
the effect of the relgene on anRNA
synthesis
VOL.
17,
1976on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.491.253.449.110.306.2]314
ERNBERG AND SKOLDA B
40 e 40
30
2
S20
/
30L
u20
30
10
-15 30 45 60 75
LABELLING TINME (SECONDS)
1o~~~~~~~~~~-~~~
15 30 45 60 75
LABELLING TIME (SECONDS)
10
5 30 45 60 75
LABELLING TIME (SECONDS)
FIG. 7. Rates of RNA synthesis during amino acid starvation ofR23-infected stringent and relaxed host bacteria. At timesindicated oneach curve, 500-,ul samples weretransferred to tubes containing [3H]adenine
andincorporationintoalkali-labileproduct wasmeasuredasdescribed in the text. Ineachexperiment, amino
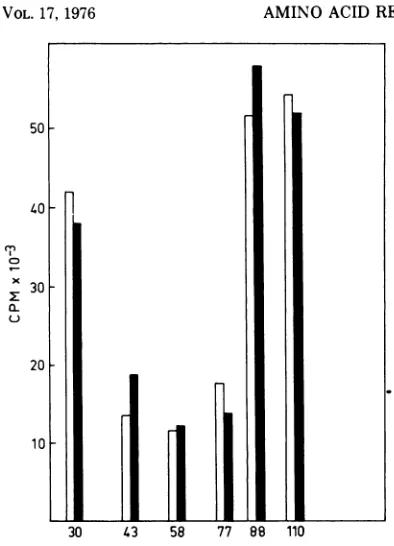
acids wereremoved at40minafterinfection(or startofexperiment) and added backat80min. Valuesalong the ordinate denotecountsper minuteincorporated per 50-Mul sample. (A) Uninfected Cp78 F+; (B) R23-infected Cp78 F+;(C)R23-infected Cp83F'.Symbols: (a) [3H] added 30 min after infection; (x) 43 min after infection; (%) 58 minafter infection; (A) 77min after infection; (0) 88 min after infection; and (U) 110 min after infection.
that is independent of the DNA-directed RNA
polymerase of E. coli. The RNA-dependent
RNA synthesis of phage R23 was chosen for
examination because of itsremarkableproperty
of
completely
dominating host RNA formationafterinfection.
Earlier studies on the stringent regulation of
phage RNA synthesis yielded conflicting
re-sults. Thus the replication of phage f2 was
observed to be dependent on the presence of
amino acids in a system where host RNA
synthesis
was inhibited by rifampin (9). Incontrast,theformation of RNAby the
serologi-cally relatedphage R17inasimilar
rifampicin-inhibited system wasfound not to be governed
by the rel geneofthe host (23). Finally,
deter-mination ofinfectiousQfB RNA formation by a
spheroplast
assay indicatedindependence
ofamino acid starvation in both stringent and
relaxed hosts (20).
In the present work the domination of R23
RNAsynthesisintheinfected cells obviated the
use of rifampin, and the careful correction for
variations in precursor pool uptake was
at-tempted.
The reported inhibition of host RNA
synthe-sis by R23 (22), which is important for the
approach
used, waschecked byelectrophoreticanalysis. The phagewasclearlyseentointerfere
with theribosomallabelingpattern. The
label-ing of boththe 16S and the 23S RNA peakswas
abolished at 40minafterphage infection. Very
little radioactivity, and
only
in adiffuse
pat-tern, was
incorporated
inthe
areacorrespond-ing to these
peaks
whenlabel
wassupplied
between
40and
60min afterinfection.
At
a timeperiod after
infection whenphage
interference
with
rRNAsynthesis
waswell
es-tablished and
phage
RNAsynthesis
ought
to havedominated,
the
cumulative
incorporation
of
both adenine and uracil
wasseverely
re-stricted
by
aminoacid
starvation ofthe
strin-gent strain,
whereas the
relaxed
strainshowed
unaffected incorporation.
Thisinhibition
ofin-corporation
wasnotdue
tolabilization
ofphage
RNA
inthe absence
of coatprotein
formation,since
the
pattern was verysimilar during
latestarvation
foronly histidine, which
is notpart ofthe coat
protein. Furthermore,
theunaffected
incorporation during
starvation ofrelaxed cells
argues
against such
aninterpretation.
Earlierreports
demonstrated
that theuptakeofuracil into the UTP
pool
ofstringentbacteria
wasseverely inhibited during amino acid
star-vation(24), andalso that the uptake of
adenine
into ATP was much less
affected under
similar
conditions (13). In the present
investigation
adenine was used in short-term
incorporations
toexamine further thedramatic effect of
starva-tion on
the
RNA synthesis of R23-infectedbacteria. The
inhibition
ofadenine
uptake intothe ATP
pool
wascorrected
forby
determiningthe
specific
activities ofthe
ATP toobtain
trueratesofRNA
synthesis
atdifferenttimes duringJ. VIROL.
m
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.491.52.440.56.233.2]50F
40-x
0L
x
30
20F
10
30 43 58 77 88 llU
FIG. 8. Relativeratesof RNA synthesis in Cp78 F+ with and without R23 phage infection. The corrected
ratesof RNA synthesiswereobtained by dividing the
radioactivity incorporated from
[3Hladenine
into RNA in 1 min (cf. Fig. 7A and B) by the specificactivity of the ATP poolateachtime of sampling (cf. Table 1).Amino acidswereremovedat40minafter infection (orstart of experiment) and addedbackat
80 min. Values below bars denote time from phage infection (0 min). Symbols: Filled bars, infected Cp78 F+; unfilled bars, uninfected Cp78F+.
amino acid starvation. These rates in
phage-infected bacteria under starvation decreased
similarly
to those of uninfectedbacteria,
indi-cating
astringent regulation
also ofphage-specific
RNAsynthesis.
This is in concurrencewith the formation of
ppGpp
in stringentR23-infected
bacteriaduring
starvation and the recentfinding
that the initiation ofQ,3
RNAsynthesis
in vitro is inhibitedby ppGpp (12).
ACKNOWLEDGMENTS
Thisinvestigationwassupported byagranttoO.S. from the Swedish Medical Research Council. J.E. is much
in-debtedtoThe SwedishAcademyofPharmaceutical Sciences forafellowship.
LITERATURE CITED
1. Bollum, F. J. 1968. Filter paper disk techniques for
assayingradioactive macromolecules, p. 169. In S. P.
Colowick and N.0.Kaplan (ed.),Methods in
enzymol-ogy,vol. 12B. Academic PressInc.,NewYork.
2. Cashel, M., and B. Kalbacher. 1970. The control of
ribonucleic acidsynthesisin Escherichia coli. J. Biol.
Chem.245:2309-2318.
3. Cashel, M., R. A. Lazzarini, and B. Kalbacher.1969.An
improved method for thin-layer chromatography of
nucleotide mixtures containing "P-labelled orthophos-phate. J. Chromatogr. 40:103-109.
4. Davis, B. J. 1964. Disc electrophoresis. II. Method and application to human serum proteins. Ann. N.Y.Acad.
Sci. 121:404-427.
5. Edlin, G., and P. Broda. 1968. Physiology and genetics of the"ribonucleic acid control" locus in Escherichia coli. Bacteriol. Rev. 32:206-226.
6. Fiil, N., and J. D. Friesen. 1968. Isolationof"relaxed" mutants of Escherichia coli. J. Bacteriol. 95:729-731. 7. Fiil, N., K. von Mevenburg, and J. D. Friesen. 1972.
Accumulation and turnover of guanosine tetraphos-phate in Escherichia coli. J. Mol. Biol.71:769-783.
8. Fraenkel-Conrat, H., B. Singer, and A.Tsugita. 1961.
Purification of viral RNA by means of bentonite. Virology 14:54-58.
9. Friesen, J. D. 1969. Dependence of f2 Bacteriophage RNA replicationonaminoacids.J. Mol. Biol. 46:349-353. 10. Gesteland,R. F. 1966. Isolation and characterization of
ribonuclease I mutants of Escherichia coli. J. Mol. Biol. 16:67-84.
11. Kjeldgaard, N., and K. Gausing. 1974. Regulation of
biosynthesisofribosomes. p. 369-392. In M.Nomura, A. Tissieres, and P. Lengyel (ed.), Ribosomes. Cold SpringHarborLaboratory, Cold Spring Harbor, N.Y. 12. Landers, T. A., T. Blumenthal, and K. Weber. 1974.
Function and structure in ribonucleic acid phage QB ribonucleic acid replicase. J. Biol. Chem. 249:5801-5808.
13. Lazzarini, R. A., and A. E. Dahlberg. 1971. The control of ribonucleic acid synthesis during amino acid depriva-tion inEscherichia coli. J. Biol. Chem. 246:420-429. 14. Lodish, H. F., and D. N. Zinder. 1966. Replication of
RNAofbacteriophagef2.Science 152:372-377.
15. Neidhardt, F. C. 1966. Roles of amino acid activating enzymes in cellular physiology. Bacteriol. Rev. 30:701-719.
16. Nierlich, D. P. 1967. Radioisotope uptake as a measure of synthesisofmessengerRNA. Science 158:1186-1188.
17. Nordstrom, K., K. G. Eriksson-Grennberg, and H. G. Boman. 1968. Resistance ofEscherichia coli to penicil-lins.Genet. Res. Comb. 12:157-168.
18. Peacock, A.C., and C. W. Dingman. 1967. Resolution of multipleribonucleic acid species by polyacrylamide gel electrophoresis. Biochemistry6: 1818-1827.
19. Peacock, A. C., and C. W. Dingman. 1968. Molecular weight estimation and separation of ribonucleic acid by electrophoresis in agarose-acrylamide composite gels. Biochemistry 7:669-674.
20. Siegel, J., and N. 0. Kjeldgaard. 1971. Effect of the rel locusonQB RNA synthesis.J. Mol. Biol. 57:147-151.
21. Watanabe, M., and J. Th. August. 1967. Methods for selecting RNA bacteriophage, p. 337. In K. Maramo-rosch and H. Koprowski (ed.), Methods in virology, vol.
III. Academic PressInc., New York.
22. Watanabe, M., H. Watanabe, and J. Th. August. 1968. Replication ofRNA bacteriophage R23. J. Mol. Biol.
33:1-20.
23. Watson, R., and H.Yamazaki.1972. Expressionof the rel
gene during R17 phage infection. Biochemistry 11:611-614.
24. Winslow, R. M., and R.A.Lazzarini.1969.Therates of
synthesis and chain elongation ofribonucleic acid in
Escherichia coli.J.Biol.Chem. 244:1128-1137.
25. Winslow, R. M., and R.A.Lazzarini. 1969. Amino acid
regulationofthe rates ofsynthesisandchainelongation
ofribonucleic acid in Escherichia coli. J. Biol.Chem.
244:3387-3392.
iin -M
315 VOL.17, 1976
![FIG.5.Fig.afterandintominwastext.infection.Cp78Cp83F-l[l4Cladenine;[14C]adenine Incorporation of [14C]uracil or ['4C]adenine the RNA of stringent and relaxed host strains R23 phage infection](https://thumb-us.123doks.com/thumbv2/123dok_us/1563210.108924/6.491.47.239.51.328/afterandintominwastext-infection-cladenine-incorporation-adenine-stringent-strains-infection.webp)
![TABLE 1.amino Uptake of [3H]uracil and [3HJadenine intothe UTP and ATP, respectively, of Cp78 F+ during acid starvation and with and without phageinfectiona](https://thumb-us.123doks.com/thumbv2/123dok_us/1563210.108924/7.491.253.449.110.306/table-uptake-uracil-hjadenine-intothe-respectively-starvation-phageinfectiona.webp)
![FIG. 7.andacidsthebacteria.Cp78(%) Rates of RNA synthesis during amino acid starvation of R23-infected stringent and relaxed host At times indicated on each curve, 500-,ul samples were transferred to tubes containing [3H]adenine incorporation into alkali-l](https://thumb-us.123doks.com/thumbv2/123dok_us/1563210.108924/8.491.52.440.56.233/andacidsthebacteria-synthesis-starvation-stringent-indicated-transferred-containing-incorporation.webp)