Copyrighti 1973 American SocietyforMicrobiology Printed inU.S.A.
Molecular
Weight
Determination
of
Sendai
RNA
by
Dimethyl
Sulfoxide
Gradient
Sedimentation
DANIEL KOLAKOFSKY AND ANDRtE BRUSCHI
Department of Molecular Biology, UniversityofGeneva, Geneva, Switzerland
Received for publication 14 December 1972
The molecularweight of the largeRNA of Sendai virus has beendetermined by sedimentation analysis in sucrose gradients containing 99% dimethyl
sulfoxide
(DMSO)
tobe 2.3 x 106.Sendai RNA recovered from 99% DMSOwasfound tocosediment with nondenatured Sendai RNA at 46 to 48s in ordinary sucrose gradients. The molecular weight value of 2.3 x 106 is considerably
smaller than the estimates of 6 x 106to7 x 106 determined under nondenatur-ingconditions, suggesting auniquestructurefor Sendai RNA.
Sendai virions contain
alarge
single-stranded RNA
(3,
4,18)
which isencapsulated
in a
nucleocapsid
structuremorphologically
similar
totobacco mosaic virus
(TMV)
butapproximately three times
aslong
(14,15). The
molecular
weight
ofparainfluenza viral RNA
(Sendai, Newcastle disease virus
[NDV],
sim-ian virus
5[SV5])
has
been estimated
by
sucrose
gradients under
nondenaturing
condi-tions to
be
approximately
6 x 106 to 7 x 106(3,
9,
11). We have
recently measured the size of
this RNA
under
completely
denaturing
condi-tions,
99%dimethyl
sulfoxide
(DMSO) (28),
and have found the molecular weight
tobe
2.3x
106,
avalue
considerably
smaller than
those
obtained from sedimentation
analyses under
nondenaturing conditions.
Since
Sendai RNA,
which
wasrecovered
from aDMSO
gradientby
ethanol precipitation,
wasfound
tocosediment
in
ordinary
sucrosegradients with
non-denatured Sendai RNA,
it isunlikely
that thislower-molecular-weight determination
wasdueto
fragmentation
ofthe
Sendai
genome (seeDiscussion). The implications
of thisdifference
are
discussed.
MATERIALS AND METHODS
Viral RNA.
3"P-Sendai
virus(Harris
strain,ob-tained fromR. D. Barry,Cambridge, England) and
32P-NDV(Californiastrain, obtained from B.Colby,
Storrs, Conn.) were grown in 9-day-old embryonated
chickeneggs labeled with 0.5mCiof 32p,per egg (at
timeofinfection) for 72 h at 34 C and 48 h at 39 C,
respectively. -HSendai virus was labeled by
prein-jectingthe eggs one day before infection with 0.3 mCi
of 3H-uridine (20 Ci/mmol). Virus was purified as
previously described (19), except that the viruswas
bandedonce in 25to65%sucrosegradients (4 h at
26,000 rpm,7C, in theSpinco SW27rotor) instead of
banding intartrategradients.
RNAwasisolated from the virionsasfollows.Virus
suspensions, at approximately 5 mg/ml in TNE
buffer (19), were made 0.2% in sodium dodecyl
sulfate(SDS)and 100Mg/mlin ProteinaseK(Merck)
and incubated for 20 min at25C. The solutionwas
then twice extracted withanequalvolume ofphenol
(saturated with TNE) at25C, and thewaterphase
was made 0.2 M inNaOAc, pH 5.3. The RNAwas
precipitated with two volumes of ethanol and left
overnight at -20C.The RNAwasthen collectedby
centrifugation, dissolvedindistilledwater, and
cen-trifuged in sucrose gradients as described below.
Fractionscontaining the fastsedimentingRNApeak
(cf.Fig.1A)werepooled, andthe RNAwasrecovered
from the gradient by ethanol precipitation. S2p_
labeled Sendai and NDV RNA, on isolation,
con-tained approximately20,000 countsper min per
jg,
'H-labeledSendaiRNA,3,300countsper min per
jg.
Other RNAs. Radioactive cellular RNAs were
obtained by labeling nonconfluent uninfected
cul-turesofmousekidney cells with4
MCi
of"C-uridine(60mCi/mmol) permlorchickenembryofibroblast
cultures with 25uCi of 3H-uridine (20Ci/mmol) per
ml for 27 h. RNA was extracted as described by
Achesonetal.(1). `C-RNA contained 10,000 counts
perminper
gg,
and3H-RNA, 150,000 countsper minper
Ag.
'H-labeled 45s rRNA precursor (220 counts per
min perMg), the kind gift of M.-E. Mirault and K.
Scherrer,wasisolated from Hela nucleoli labeled for
15 min with methionine-methyl-3H CT,, as
previ-ouslydescribed (22).
Sucrosegradient sedimentation. Linear sucrose
gradients (5-23%) contained 0.1 M LiCl, 10 mM Tris-hydrochloride (pH 7.4), 4 mM EDTA, and 0.1%
615
on November 10, 2019 by guest
http://jvi.asm.org/
KOLAKOFSKY ANDBRUSCHI
SDS. LiCl was used in place of NaCl to aid the
solubility of SDS at lower temperatures.
Centrifuga-tions were carried out for 105 min at 50,000 rpm, at 7
C, in the Spinco SW56 rotor. Gradients were
frac-tionatedbypuncturing thebottom of the tube,and
drops were collected on paper discs. The paper discs
were then dried, washed first in 6% trichloroacetic
acid and then in ethanol, dried, and counted by liquid scintillation.
DMSO gradient sedimentation. DMSO
gradi-ents were prepared, and sample application was
carried out as described by Acheson et al. (1).
Because the absolutesedimentation rate of RNA was
found to varyconsiderably in different preparations
of the DMSO solutions (presumably due to the
hygroscopic nature of the solvent),DMSOsolutions
were prepared in batchquantities, divided into equal
parts, and kept sealed in a desiccator to ensure
reproducibility from runto run. Gradients reported
here were centrifugedfrom 3 to 4.5 h at 48,000 rpm,
26 C, in the Spinco SW56 rotor. Gradients were
fractionated and countedas described above.
RESULTS
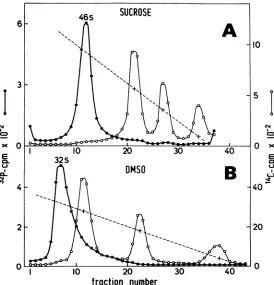
In sucrose gradients
containing 0.1 M NaCl,
the sedimentation
value of Sendai RNA has
46s
SUCF
6-3
/
EI 10 20
32S
8
^
DM
c
W
been
measured
as 57srelative
toTMV RNA
(3), and 50s relative
to28sand
18srRNA
(18).
The RNA of
twoother
type IImyxoviruses,
NDV and SV5,
have both been measured
as50s
(9, 16).
Under the conditions
wehave used
(0.1M
LiCl,
seeMaterials and
Methods), the large
RNA of four different
preparations ofSendai
virus
wasfound
tosediment
asasharp peak
at46 to 48s
relative
to 28sand 1&s rRNA
(Fig. 1A). However,when the
same RNA was run ingradients containing
99%DMSO, Sendai RNA
was
found
tosediment just ahead
of28s
rRNA,at 32s.
(All
sedimentation values
inDMSO
reported
here
arerelative
to theribosomal
RNAs
taken
as 18sand
28s, inDMSO).
Be-cause
of
this unexpected finding, the
DMSO
gradients
werecalibrated with
anRNA marker
expected
tosediment faster than
Sendai
RNA
in
DMSO,
namely,
the 45s ribosomalRNA
precursor (29).
As
canbe
seen in Fig. 2, the 45srRNA
precursor (46s under our conditions on
sucrose) was found to sediment at 43s in
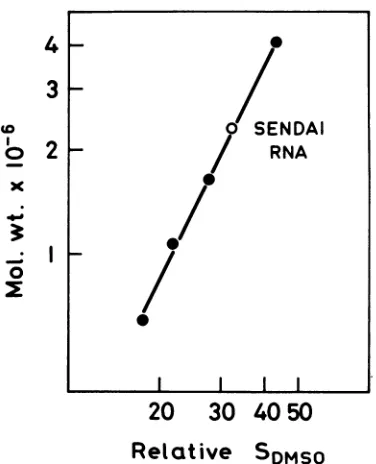
DMSO. The molecular weight of Sendai RNA
using the
abovementioned markers (and R17
)SE
A
10
30 40 0
E
B
20
fraction number
FIG. 1. Comparisonof32P-Sendai RNA and14C-cellularRNA
(4s,
18s,and28sRNA)onordinary
sucrose(A)andDMSOgradients
(B).
Gradient(A)
contained0.6sg of
32P-Sendai RNA(3,200
counts/min)and 0.5ygof"IC-cellular RNA. Gradient
(B)
contained 0.6ygof
82P-SendaiRNA and 2.0ggof "4C-cellular RNA.Con-ditionsofcentrifugationand
counting
aredescribed in Materials and Methods.616
J.VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.493.121.395.336.621.2]VOL. 11, 1973
MOLECULAR WEIGHT
RNA
xe
E
I
E
2-2
10 20 30 40
[image:3.493.47.232.60.399.2]fraction number
FIG. 2. Comparison of 'H-45s rRNA precursor,
"4C-cellular
RNA,
and32P-Sendai RNAonordinary
sucrose (A) and DMSOgradients (B)
and(C).
Gradient (A) contained2.9
Mg of
3H-45s rRNA and 2.0Mg
of "4C-cellularRNA. Gradient(B)
contained2.9 ug of 3H-45s rRNA and 2.0
Mg of
"4C-cellularRNA. Gradient(C) contained 2.9
Mg of
'H-45srRNAand 2.0Mg of32P-Sendai RNA (4,100 counts/min).
RNA, data
notshown)
wasestimated
tobe
2.3x
106
(Fig. 3).
The
possibility that,
asfor
Rous
sarcomavirus
(RSV)
RNA
(10, 12,
23),
the
large
differ-ence
insedimentation
rateof
Sendai RNA
inDMSO and sucrose
gradients
might be due to
disaggregation
of
asegmented
genome
inDMSO
wasnext
examined.
"P-labeled Sendai
RNA
wasprepared
by
first
sedimenting
it ona
DMSO gradient, and the RNA sedimenting at
32SDMSO
was recovered from the
gradient
by
ethanol
precipitation after
theprofile of the
gradient
hadbeen determined by
Cerenkov
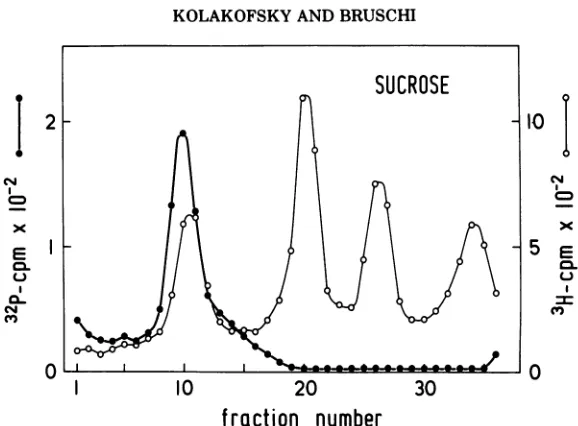
counting (cf. Fig. 1B). This Sendai RNA was
then
mixed withnondenatured,
8H-labeled
Sendai
RNAand cell
RNAmarkers and
resedi-mented
inan
ordinary sucrose gradient. The
results, shown
inFig. 4,
demonstrate that
Sendai
RNAsediments
at thesame
position, 46
to
48s, even
after recovery from the DMSO
gradient.
Thus, unlike RSV RNA, the
sedimen-tation
behavior of
Sendai.
RNA in sucrose
gradients is unchanged
by DMSO treatment,
and 46 to 48s
Sendai RNA appears to be a
single, continuous RNA chain of
2.3x
101(see
Discussion).
Although the molecular weight of Sendai
RNA has
not previously
been measured under
denaturing
conditions, Duesberg
(10) has
measured the molecular
weight of the
large
RNA of
NDV, another type II
myxovirus, in
DMSO gradients as 6.3 x 10' daltons. We have
therefore
compared
the
large
RNAs of Sendai
and NDV on both
ordinary
sucrose
and
DMSO
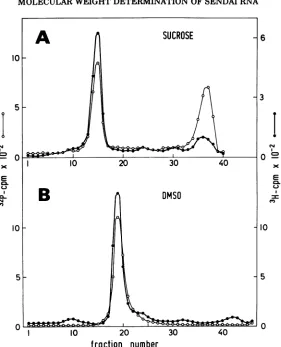
gradients. We find that Sendai and NDV RNA
are
indistinguishable
in
sedimentation
ratein
both
ordinary
sucrose(reference
4,
Fig. 5A)
and
DMSO
gradients
(Fig.
5B),
and therefore
con-clude that the
large
RNA of
NDV,
like that of
Sendai
virus,
has
amolecular
weight
of
2.3x
106.
DISCUSSION
As
previously
mentioned, Duesberg (10)
has
reported the molecular
weight
ofNDV RNA in
4
3
I
0
x
3:
0
2
2
20
30 40
50
Relative
SDMSO
FIG. 3. Determination of the molecular weight of
Sendai RNA bysedimentation inDMSOgradients.
RNAs used as reference markers were: 18s rRNA,
0.65 x 10' (20); R17RNA, 1.05 x 10' (12); 28s rRNA,
1.65 x
106
(20); 45s rRNA precursor, 4.1 x106
(27).All
SDS,O
arerelative to 28s and 18s rRNA taken as28& and 18s in DMSO. The molecular weight of
Sendai RNA, represented by an open circle, is
determined as 2.3 x 10 .
617
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.248.436.357.589.2]KOLAKOFSKY AND BRUSCHI
2
0
10 20 30
I0
C-)
5
E6.)
0
fraction
number
FIG. 4. Comparison of DMSO-denatured and nondenaturedSendai RNAon sucrosegradients. 32P-Sendai
RNA (3.5,g,10,400counts/min)wasfirst sedimentedinaDMSO gradient (cf. Fig.1B),and theprofileof the
gradientwasdetermined by Cerenkovcounting. RNApresentinthe threepeak tubeswasrecoveredfrom the
gradient by ethanol precipitation and dissolved in distilled water. A sample of this material (1,200
counts/min)wasmixedwith 1.1 Mgof nondenatured sH-Sendai RNA and0.09ugof 3H-cellular RNA andthen
sedimented inanordinarysucrosegradient.
DMSO gradients to be 6.3 x 106, whereas we
find both NDV and Sendai RNA to be 2.3 x
106, using similar methods. The molecular weight value of 6.3 x 106l, however, was
ob-tained by means of the empirical equation
determined by Strauss et al. (28) for the absolute sedimentation rate of RNA in the analytical ultracentrifuge and not by
extrapo-lation fromthesedimentation rateofRNAsof known molecularweight determined under si-milar conditions, i.e., sucrose gradients in DMSO. Because of the large effects ofsucrose on theviscosity of solutions, it isunlikely that
the empirical equation determined by Strauss
etal. wasapplicablein this case.
The large difference in molecular weight between thatexpectedfora46to48sRNA(27) and that measured under denaturing
condi-tions can be accounted for if Sendai RNA
contained a great deal more secondary
struc-ture than the ribosomal RNAs. Sendai RNA
has recently been shown to self-anneal, to
variableextents(16-60%,references24,25)ina
concentration-dependent fashion. However, as
Portner and Kingsbury (24) havepointed out,
the fact that the RNA had been degraded to
approximately 10s during the annealing
reac-tiondidnot allow themtodistinguishwhether
thisannealingwasintra-orintermolecular.We
have also found that Sendai RNAself-anneals
(15-25%, datanotshown),and if thisannealing
is in fact intramolecular, it could explain the
abovementioned discrepancy in sedimentation
rateunderdenaturing and nondenaturing
con-ditions.
There is, however, another possibility to
explain this discrepancy. Ourconclusions that the 46to48sRNA iscomposed ofasingle chain
of 2.3 x 106 molecular weight is basedon the
unaltered sedimentation rate of Sendai RNA
on sucrose gradients after
DMSO
treatment(cf. Fig. 4). However, if Sendai RNA were
capable of rapid andquantitative reassociation (e.g., byannealing ofcomplementary ends) due
tothe increase inconcentration when theRNA is removed from the denaturing solvent by ethanol precipitation and subsequent solution
in water, then the 46 to 48s RNA could be
composed ofmorethanoneRNAchain of2.3 x 106 molecular weight. Although this latter possibility seems unlikely, there is some
evi-dence in its favor. In thevirion, parainfluenza virus RNAoccursintheformofanucleocapsid
which is remarkably similar in structure to
TMV(6), but
approximately
3.3 timesaslong(6-8, 14, 15, 17). These nucleocapsids have
beenshowntocontainfrom3.7to4.1% RNA(7, 14), only slightly less than the 5.1% RNA
determined for TMV (19), whose RNA has a
molecular weight of 2.1 x 106 (5). These
nucleocapsids should therefore contain 5.0 x
106 to 5.6 x 106 daltons of RNA. Since the
molecular weight of Sendai RNA as
deter-mined by DMSO gradient sedimentation is 0~
L)
C.) x
E
618 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.111.400.60.273.2]I0
I4
N
x
C.
Ci4
C,)
6
3
N
I
°x
cL u
I
10
5
O
I
10
20
30
40
[image:5.493.101.382.64.411.2]frraction number
FIG. 5. Comparison of32P-NDV RNA and
3H-Sendai
RNA onordinary
sucrose(A)
andDMSO
gradients
(B).Gradient
(A)
contained38,000 counts/minof
32P-NDV virions and 0.8Mtg of
3H-Sendai RNA. Thesamplesweresuspended in 75
Mliters
ofwater,made1%inSDS,and thensedimentedon anordinary
sucrosegradient.
Gradient(B) contained0.4
Mgg
of32P-NDV RNA (4,600counts/min)
and1.6Mg of
'H-SendaiRNA.well
below this
value, Sendai
nucleocapsids
should therefore contain
morethan
onechain
of
RNA. It is
possible
that the
structurerespon-sible
forthese
multiple
chains
ofRNA in the
nucleocapsid
isreassumed
onremoval
ofthe
RNA from
DMSO;
this reassociation would
ofcourse
be concentration
dependent.
In
conclusion,
wewould like
toadd
a noteof caution.This
discrepancy
between
sedimenta-tion
value
inordinary
sucroseand
DMSO
gradients
is notunique
toSendai RNA.
Arifand Faulkner(2) have recently shown that 42s
Sindbis
RNAsediments
only
slightly
fasterthan
28s rRNA
inDMSO
gradients, and thatthis
RNA, when
recovered from a DMSOgradient, coelectrophoreses
onpolyacrylamide
gels
with untreated 42s RNA. It seems clear that these animal viral RNAsare different fromrRNAand
bacteriophage
RNA in this respect.Even
though these RNAs have been purified ina
similar
manner asrRNA and
bacteriophage
RNA,
the
possibility
cannotbe excluded that
they
still contain material which
candifferen-tially
affect
their
buoyant density
inwaterand
DMSO (the
inability of Simmons and Strauss
[26]
tosediment
Sindbis viral RNA
as adiscrete band
inDMSO
gradients
is
note-worthy
inthis respect), thus
making molecular
weight estimation of these viral RNAs
inDMSO
gradients unreliable using rRNA and
bacteriophage RNA
asmarkers.
ACKNOWLEDGMENTS
We thank Peter Bromley for helpful discussion, and
Pierre-FrangoisSpahr for generous advice, encouragement, and support.
Thiswork wassupported by research grant no. 3.816.72 from the Fonds National Suisse de la Recherche Scien-tifique.
LITERATURE CITED
1. Acheson, N. H., E. Buetti, K. Scherrer, and R. Weil.
VOL.11,
619
on November 10, 2019 by guest
http://jvi.asm.org/
KOLAKOFSKYANDBRUSCHI
1971. Transcription of the polyoma virus genome:
synthesis andcleavage ofgiant latepolyomaspecific RNA. Proc. Nat. Acad.Sci. U.S.A.68:2231-2235. 2. Arif, B. M., and P. Faulkner. 1972.Genome of Sindbis
virus.J. Virol.9:102-109.
3. Barry, R. D., andA.G.Bukrinskaya.1968. The nucleic acid of Sendai virus and ribonucleic acid synthesis in cells infectedby Sendaivirus. J. Gen. Virol.2:71-79. 4. Blair, C. D., and W. S.Robinson. 1968.Replicationof Sendai virus, comparison of viral RNA and virus specific RNA synthesiswith Newcastle disease virus. Virology35:537-549.
5. Boedtker, H. 1960.Configurational propertiesoftobacco mosaicvirusribonucleic acid.J. Mol. Biol.2:171-188. 6. Choppin, P. W., and W. Stoeckenius. 1964. The
mor-phology of SV5 virus. Virology 23:195-202.
7. Compans, R. W., and P. W. Choppin. 1967. Isolation and properties of the helical nucleocapsid of the parainfluenzavirusSV5.Proc. Nat. Acad. Sci.U.S.A. 57:949-956.
8. Compans, R.W.,and P. W. Choppin, 1967. Thelength of the helicalnucleocapsidofNewcastle disease virus. Virology33:344-346.
9. Compans,R.W.,and P. W.Choppin.1968. Thenucleic acid ofparainfluenzavirusSV5.Virology35:289-296. 10. Duesberg, P. H. 1968. Physical properties of Rous
sarcoma virus RNA. Proc. Nat. Acad. Sci. X.S.A. 60:1511-1518.
11. Duesberg, P. H., and W. S. Robinson.1965.Isolation of the nucleic acid of Newcastle disease virus (NDV). Proc. Nat. Acad. Sci. U.S.A. 54: 794-800.
12. Erikson, R. L. 1969. Studies onthe RNA from avian myeloblastosisvirus.Virology 37:124-131.
13. Gesteland,R.F.,and H.Boedtker.1964.Some physical properties ofbacteriophage R17 and its ribonucleic acid.J. Mol. Biol.8:496-507.
14. Hosaka, Y. 1968. Isolation and structure of the
nu-cleocapsidof HVJ.Virology35:445-457.
15. Hosaka, Y.,H.Kitano,and S.Ikeguchi.1966. Studieson
the pleomorphism of HVJ virions. Virology 29:205-221.
16. Kingsbury,D. W. 1966. Newcastle disease virus RNA.I.
Isolation and preliminary characterization ofRNA fromvirusparticles.J. Mol. Biol. 18:195-203. 17. Kingsbury, D. W., andR. W. Darlington.1968.Isolation
and propertiesofNewcastle diseasevirus nucleocap-sid. J. Virol.2:248-255.
18. Kingsbury, D. W., A. Portner, and R. W. Darlington. 1970. Properties of incomplete Sendai virions and subgenomic viral RNAs. Virology42:857-871. 19. Knight, C. A., and B. R. Woody. 1958. Phosphorous
content of tobacco mosaic virus. Arch. Biochem. Biophys.78:460-467.
20. Kolakofsky, D. 1972. Transfer ribonucleic acid
nu-cleotidyltransferase andtransfer ribonucleic acid in Sendai virions. J. Virol. 10:555-559.
21. McConkey, E. H., and J. W. Hopkins.1969. Molecular weightsofsomeHelaribosomalRNAs. J. Mol. Biol. 39:545-550.
22. Mirault, M.-E., and K. Scherrer. 1971. Isolation of preribosomes fromHela cellsand their characteriza-tion by electrophoresis onuniform and exponential-gradient polyacrylamide gels. Eur. J. Biochem. 23:372-386.
23. Montagnier, L.,A.Golde,and P.Vigier.1969. Apossible subunit structure of Rous sarcoma virus RNA. J. Gen.Virol. 4:449-452.
24. Portner, A., and D. W. Kingsbury. 1970. Complemen-tary RNAs in paramyxovirions and paramyxovirus-infected cells. Nature(London)228:1196-1197. 25. Robinson, W. S. 1970. Self-annealing of subgroup 2
myxovirusRNAs.Nature(London) 225:944-945. 26. Simmons,D.T.,and J.H.Strauss. 1972.Replicationof
Sindbisvirus. I. Relative size andgeneticcontent of
26Sand 49SRNA. J. Mol. Biol.71:599-613. 27. Spirin, A. S. 1963. Some problems concerning the
macromolecularstructureof ribonucleic acids.Progr. Nucleic Acid Res. 1:301-345.
28. Strauss, J. H. Jr., R. B. Kelly, and R. L. Sinsheimer. 1968. Denaturation of RNA withdimethyl sulfoxide. Biopolymers6:793-807.
29. Weinberg,R.A.,and S. Penman. 1970.Processingof 45 S nucleolar RNA. J. Mol. Biol.47:169-178.