ORF57 Protein Is Regulated by Protein Phosphorylation and
Homodimerization
Vladimir Majerciak,aNatalia Pripuzova,a*Calvin Chan,aNicholas Temkin,aSuzanne I. Specht,bZhi-Ming Zhenga Tumor Virus RNA Biology Section, Gene Regulation and Chromosome Biology Laboratory,a
and Laboratory of Cell and Developmental Signaling,b
National Cancer Institute, NIH, Frederick, Maryland, USA
ABSTRACT
Kaposi’s sarcoma-associated herpesvirus (KSHV) ORF57 plays an essential role in KSHV lytic infection by promoting viral gene expression at the posttranscriptional level. Using bioinformatic and biochemical approaches, we determined that ORF57 con-tains two structurally and functionally distinct domains: a disordered nonstructural N-terminal domain (amino acids [aa] 1 to
152) and a structured␣-helix-rich C-terminal domain (aa 153 to 455). The N-terminal domain mediates ORF57 interaction with
several RNA-protein complexes essential for ORF57 to function. The N-terminal phosphorylation by cellular casein kinase II (CKII) at S21, T32, and S43, and other cellular kinases at S95 and S97 residues in proximity of the caspase-7 cleavage site, 30-DETD-33, inhibits caspase-7 digestion of ORF57. The structured C-terminal domain mediates homodimerization of ORF57, and
the critical region for this function was mapped carefully to␣-helices 7 to 9. Introduction of point mutations into␣-helix 7 at
ORF57 aa 280 to 299, a region highly conserved among ORF57 homologues from other herpesviruses, inhibited ORF57 ho-modimerization and led to proteasome-mediated degradation of ORF57 protein. Thus, hoho-modimerization of ORF57 via its C terminus prevents ORF57 from degrading and allows two structure-free N termini of the dimerized ORF57 to work coordinately for host factor interactions, leading to productive KSHV lytic infection and pathogenesis.
IMPORTANCE
KSHV is a human oncogenic virus linked to the development of several malignancies. KSHV-mediated oncogenesis requires both latent and lytic infection. The KSHV ORF57 protein is essential for KSHV lytic replication, as it regulates the expression of viral lytic genes at the posttranscriptional level. This report provides evidence that the structural conformation of the ORF57 protein plays a critical role in regulation of ORF57 stability. Phosphorylation by CKII on the identified serine/threonine residues at the N-terminal unstructured domain of ORF57 prevents its digestion by caspase-7. The C-terminal domain of ORF57, which is
rich in␣-helices, contributes to homodimerization of ORF57 to prevent proteasome-mediated protein degradation. Elucidation
of the ORF57 structure not only enables us to better understand ORF57 stability and functions but also provides an important tool for us to modulate ORF57’s activity with the aim to inhibit KSHV lytic replication.
K
aposi’s sarcoma-associated herpesvirus (KSHV) ORF57 (alsoknown as Mta) is expressed early in the KSHV lytic cycle and is required for the efficient expression of a subset of viral genes, including KSHV PAN, ORF59, K8, viral interleukin-6 (vIL-6),
ORF47, and others (1–7). A KSHV genome lacking ORF57
ex-pression is associated with a defective lytic cycle incapable of
pro-ducing infectious virions (8,9).
KSHV ORF57 functions as a posttranscriptional regulator of viral gene expression by affecting RNA stability (PAN, ORF59, and ORF47), splicing (ORF50 and K8), polyadenylation (ORF59),
and translation (vIL-6) (1,2,4–7,10) but appears not to promote
RNA export (11,12). Whether ORF57 directly promotes KSHV
genome instability in infected cells (13) remains to be confirmed.
Although all ORF57 functions involve ORF57 association with an RNA target, this association also requires cellular proteins to
func-tion as ORF57 cofactors (14,15), and each of the ORF57-specific
functions depends on a specific cofactor(s). This has been dem-onstrated by the observation that ORF57 stabilizes PAN RNA via
interaction with PABPC1 (16), that ORF57 mediates K8 splicing
by interaction with SRSF3 (7), that ORF57 enhances ORF59
ex-pression by the supex-pression of SPEN-induced nuclear
hyperpoly-adenylation (4), and that ORF57 promotes vIL-6 translation by
preventing Ago2, a major component of RISC complexes, from
interacting with a microRNA binding site in vIL-6 RNA (6).
ORF57 interacts with Aly/REF (12,14,17,18), a ubiquitously
expressed nuclear protein which functions as a molecular chaper-one to regulate dimerization, DNA binding, and transcriptional
activity of basic region-leucine zipper (bZIP) proteins (19,20). It
Received28 December 2014Accepted30 December 2014
Accepted manuscript posted online7 January 2015
CitationMajerciak V, Pripuzova N, Chan C, Temkin N, Specht SI, Zheng Z-M. 2015. Stability of structured Kaposi’s sarcoma-associated herpesvirus ORF57 protein is regulated by protein phosphorylation and homodimerization. J Virol 89:3256 – 3274.doi:10.1128/JVI.03721-14.
Editor:R. M. Sandri-Goldin
Address correspondence to Zhi-Ming Zheng, [email protected].
* Present address: Natalia Pripuzova, Tumor Vaccines and Biotechnology Branch, Division of Cellular and Gene Therapies, CBER, FDA, Silver Spring, Maryland, USA.
Supplemental material for this article may be found athttp://dx.doi.org/10.1128 /JVI.03721-14.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.03721-14
on November 7, 2019 by guest
http://jvi.asm.org/
was initially viewed as an RNA export cofactor (21,22), but this interaction is not necessary for RNA export of viral intronless RNAs. Several lines of evidence support the latter conclusion. First, depletion of Aly/REF from HEK293 nuclear extract does not affect the ORF57 interaction with KSHV intronless ORF59 RNA, and small interfering RNA knockdown of Aly/REF from HeLa or HEK293 cells does not affect ORF57-mediated enhancement of
ORF59 expression (14). Second, an ORF57 mutant with a
defi-ciency in Aly/REF binding retains its ability to accumulate KSHV
target mRNAs (12). Third, the Aly/REF-ORF57 interaction has
been demonstrated to be nonessential for KSHV lytic replication but contributes to target RNA stability independently of effects in
RNA export (23,24). Fourth, a recent well-designed study
con-cluded that ORF57 does not provide the specific RNA export func-tion and is not a bona fide export factor for KSHV intronless RNAs
(11). Finally, knockout of Aly/REF expression does not affect
mRNA export inDrosophila melanogastercells andCaenorhabditis
elegans(25,26) and in herpes simplex virus 1 (HSV-1) infection
(27).
Posttranscriptional regulators with similar activities to KSHV ORF57 are also encoded by other members of the herpesvirus
family. These include well-characterized HSV-1 ICP27 (28),
hu-man cytomegalovirus virus (HCMV) UL69 (29), Epstein-Barr
vi-rus (EBV) EB2 (or EB-SM) (30), and herpesvirus saimiri (HVS)
ORF57 (31). While all homologues in the family share many
com-mon activities, they diverge with regard to specific functions and target specificities. Therefore, they do not complement each oth-er’s function to rescue virus infection by a homologue-deficient
genome (9,32). Although the functions of KSHV ORF57 and its
homologues became more understood over recent decades, the regulation of their unique activities, in particular the contribution of their protein structures to their unique activities, remains to be elucidated. To date, there is no crystal structure for any member in this family. Because ORF57 has no homology to any known cellu-lar proteins and bears only low amino acid sequence homology to its close homologues, it remains a great challenge to solve the ORF57 three-dimensional (3D) structure and correlate its struc-ture with its function.
In this study, we conducted a comprehensive analysis of the ORF57 structure, both computationally and experimentally, and determined that the ORF57 protein contains two structurally and functionally distinct domains: an intrinsically disordered
N-ter-minal domain and a structured C-terN-ter-minal domain. We found that the distinct structures in the identified domains dictate ORF57 binding activities toward its self-association and interac-tions with cellular cofactors.
MATERIALS AND METHODS
In silicoprotein sequence analysis.Protein secondary structures were predicted with PSIPRED version 3.0 (a PSI-BLAST-based secondary structure prediction tool publicly available athttp://bioinf.cs.ucl.ac.uk /psipred/) (33). Links to all programs used for prediction of the intrinsi-cally disordered region can be found athttp://www.disprot.org/predictors .php. The kinase-specific eukaryotic protein phosphorylation sites were predicted by using the NetPhosK 1.0 server (http://www.cbs.dtu.dk /services/NetPhosK/) (34). The sequence homology analysis was per-formed by using ClustalW2, which is located athttp://www.ebi.ac.uk /Tools/msa/clustalw2/.
Cells.Human HEK293, HeLa, and monkey COS-1 cells were culti-vated in Dulbecco’s modified Eagle medium supplemented with 10% fetal bovine serum (FBS; HyClone). KSHV-positive BCBL-1 (35) and JSC-1 (36) B cells were maintained in RPMI 1640 medium with 10% FBS. The KSHV lytic cycle was induced by treatment with 3 mM so-dium butyrate (JSC-1 cells) or 0.6 mM soso-dium valproate (BCBL-1 cells) for 24 h. All transfections were carried out with Lipofectamine 2000 (Invitrogen) or LipoD293 (SignaGen Laboratories) as recom-mended by the manufacturers.
Plasmids and mutagenesis. The eukaryotic expression vectors pFLAG-CMV-5.1 and p3⫻FLAG-CMV-14 (Sigma) and pEGFP-N1 (Clontech) were used. These previously described plasmids were used to express various forms of ORF57 protein and EBV EB2, as follows: pVM7 (ORF57-FLAG), pVM68 (ORF57-3⫻FLAG), pVM24 (ORF57-FLAG amino acids [aa] 1 to 251;⌬C wild type [wt]), pVM45-51 (ORF57-FLAG aa 1 to 251;⌬C with single, double, or triple nuclear localization signal [NLS] mutations), pVM35 (ORF57-GFP [green fluorescent protein]; mtNLS-1, -2, and -3 [mutated NLSs]), pVM20 (ORF57-GFP aa 167 to 455;⌬N), pcDNA-ORF57, and pGS113 (EB2-myc) (14,37). The mu-tagenesis of conserved residues in the ORF57 helix 7 region was per-formed by overlapping PCR using the primers listed inTable 1. The am-plicon with the designed mutation(s) in ORF57 was recloned into p3⫻FLAG-CMV-14 to generate the following plasmids: pVM83 (ORF57-3⫻FLAG, E287A, E288A), pVM84 (ORF57-3⫻FLAG, E287P, E288P), pVM85 (ORF57-3⫻FLAG, W292A), pVM86 (ORF57-3⫻FLAG, W292P), and pVM99 (ORF57-3⫻FLAG, K345A).
[image:2.585.42.555.77.216.2]Antibodies and recombinant ORF57 protein.Rabbit polyclonal an-tibody targeting the ORF57 N terminus (aa 119-PEKRPRRRPRDRLQ-132) was described before (8). To generate the antibody against the
TABLE 1Oligonucleotides used in ORF57 mutagenesis
Oligonucleotide Sequencea Mutation(s) Plasmid
oVM193 5=-CTTTCGTGGCGGCACAAATGACGTGGGCC-3= E287A, E288A pVM83
oVM194 5=-CATTTGTGCCGCCACGAAAGCCCCAAGCG-3= E287A, E288A pVM83
oVM195 5=-CTTTCGTGCCGCCACAAATGACGTGGGCC-3= E287P, E288P pVM84
oVM196 5=-CATTTGTGGCGGCACGAAAGCCCCAAGCG-3= E287P, E288P pVM84
oVM197 5=-CAAATGACGGCGGCCCAGACGGTTGTGC-3= W292A pVM85
oVM198 5=-CTGGGCCGCCGTCATTTGTTCCTCCACGA-3= W292A pVM85
oVM199 5=-CAAATGACGCCGGCCCAGACGGTTGTGC-3= W292P pVM86
oVM200 5=-CTGGGCCGGCGTCATTTGTTCCTCCACGA-3= W292P pVM86
oVM273 5=-AGCACTGATCGCACAGGTGGCATATTTGGTA-3= K345A pVM99
oVM274 5=-ATGCCACCTGTGCGATCAGTGCTAGCTCGCT-3= K345A pVM99
oVM68b 5=-TACTCAGAATTCACC/ATGGTACAAGCAATGATAGACATGG-3=
oVM69b 5=-ATCGTGGATCC/AGAAAGTGGATAAAAGAATAAACCCTTG-3=
aUnderlined portions of sequences indicate point mutations. b
oVM68 is a forward primer with an EcoRI site, and oVM69 is a backward primer with a BamHI site (a slash divides the restriction cutting site from the viral primer sequence).
on November 7, 2019 by guest
http://jvi.asm.org/
ORF57 C terminus, rabbits were immunized with a synthetic peptide corresponding to aa 394 to 412 (394-ARGQELFRTLLEYYRPGDV-412) of the ORF57 protein sequence. Other antibodies used in the study were anti-hnRNPU (Ab10297; Abcam), anti-RNase helicase A (Ab26271; Ab-cam), anti-nucleolin C23 (Ab13541; AbAb-cam), anti-FLAG M2 (F1804; Sigma), anti-c-Myc (M4439; Sigma), anti-neomycin phosphotransferase II (06-747; Millipore), Lys48-specific anti-ubiquitin rabbit monoclonal antibody (clone Apu2; 05-1307; Millipore), and anti--tubulin (T5201; Sigma). Recombinant full-length ORF57 protein containing a C-terminal FLAG tag was expressed in Sf21 insect cells by using a baculovirus vector and purified by immunoaffinity column chromatography with an anti-FLAG antibody (38).
Limited proteolysis and Edman protein sequencing.Lyophilized clostripain and proteinase K were obtained from Sigma and reconstituted as recommended by the manufacturer. The ORF57 limited proteolytic digestion was carried out in a 10-l reaction mixture containing 4g of purified ORF57-FLAG protein or whole-cell extract resuspended in im-munoprecipitation (IP) buffer (50 mM HEPES [pH 7.4], 150 mM NaCl, 1 mM EDTA, 2.5 mM EGTA, 10% glycerol, 0.1% NP-40) supplemented with increasing amounts of clostripain or proteinase K. The digestion reaction mixture was incubated for 3 min at room temperature and stopped by addition of 1l of 1 M protease inhibitor phenylmethylsulfo-nyl fluoride and 10l of 2⫻SDS protein sample buffer containing 10% (vol/vol) 2-mercaptoethanol (2-ME). After digestion, the samples were separated via SDS-PAGE and stained with GelCode Blue stain reagent (here referred to as Gel Blue; Thermo Scientific) or analyzed by Western blotting. To determine the N-terminal amino acid sequence of the ORF57 protease-resistant (PR) region, the digested ORF57 products with 0.1 U of clostripain were subjected to Edman degradation chemistry using an ABI Procise 494 sequencer (Alphalyse Inc.).
Immunoprecipitation and mass spectrometry.The whole-cell ex-tract used for IP was prepared by resuspending cell pellets in ice-cold RSB-150 buffer (10 mM Tris-HCl [pH 7.5], 150 mM NaCl, 2.5 mM MgCl2, 0.1% NP-40). After brief sonication, the extract was cleared by centrifugation for 10 min at 10,000⫻gat 4°C, diluted in IP buffer, and incubated with antibody-coated agarose beads overnight at 4°C. After extensive washing, the immunoprecipitated complexes were eluted with IP buffer supplemented with 150 ng/l of 3⫻FLAG peptide (Sigma) for 1 h at 4°C or with 2⫻SDS protein sample buffer supplemented with 5% (vol/vol) 2-ME. In some cases, the prepared cell extract was pretreated before IP with RNase A (Ambion) or DNase I (New England Biolabs) for 30 min at room temperature. Immunoprecipitated proteins were sepa-rated on SDS-PAGE and detected by Gel Blue staining or by Western blotting. The identity of immunoprecipitated proteins was determined by tandem mass spectrometry (MS/MS) after in-gel trypsin digestion (Prot-Tech, Norristown, PA).
In vitrophosphorylation and dephosphorylation.ORF57 proteinin vitrophosphorylation was carried out in a 10-l reaction mixture con-taining 400 ng of recombinant ORF57-FLAG protein, 1⫻casein kinase II (CKII) buffer (20 mM Tris-HCl [pH 7.5], 50 mM KCl, 10 mM MgCl2), 1 mM cold ATP or 25Ci [␥-32P]ATP, and 125 U of recombinant CKII
(New England BioLabs). The reaction mixture was incubated for 1 h at 30°C and stopped by addition of 10l of 2⫻SDS protein sample buffer with 10% (vol/vol) 2-ME. The32P-labeled ORF57 was separated by
SDS-PAGE, transferred to a polyvinylidene difluoride (PVDF) membrane, and detected by using a PhosphorImager (Molecular Dynamic). The band corresponding to phosphorylated ORF57 was then cut from the mem-brane for further analysis.In vitrodephosphorylation was performed in a 50-l reaction mixture containing 10l of cell extract or 1g of recom-binant ORF57-FLAG protein diluted in 1⫻protein phosphatase (PPase) buffer (50 mM HEPES [pH 7.5], 10 mM NaCl, 2 mM dithiothreitol [DTT], 0.01% Brij 35) supplemented with 1 mM MnCl2and 1,000 U of
Lambda bacteriophage-PPase (New England Biolabs). The reaction mixture was incubated at 30°C for 1 h. The-PPase activity was inhibited
by addition of 10 mM sodium orthovanadate (Na3VO4) or 50 mM EDTA
to the reaction mixture.
Mapping of ORF57 phosphorylation sites.The phosphorylated resi-dues in the ORF57 protein were mapped by using two complementary methods. First, recombinant ORF57-FLAG protein was subjected to tryp-tic digestion, and the resulting peptides were evaluated by matrix-assisted laser desorption ionization–time of flight analysis. A capillary liquid chro-matography-electrospray ionization-MS (LC-ESI-MS) setup with an ion trap mass analyzer (ProtTech) was used to determine the exact position of phosphorylated residues. Second, the recombinant ORF57 protein was firstin vitrophosphorylated with recombinant CKII in the presence of [␥-32P]ATP as described above. Radiolabeled ORF57 was separated by
SDS-PAGE, transferred to a PVDF membrane, and detected with a PhosphorImager. The membrane containing the phosphorylated ORF57 protein was then subjected to proteolytic digestion with trypsin or endo-proteinase GluC. The phosphopeptides carrying32P labeling were isolated by reverse-phase high-performance liquid chromatography (HPLC) and subjected to phosphoamino amino acid analysis and phosphopeptide mapping by using Edman degradation as described previously (39,40).
In vitrocaspase cleavage assay.ORF57in vitrocaspase cleavage was performed as described previously (38). Briefly, a 20-l reaction mixture containing 200 ng of recombinant ORF57-FLAG protein diluted in 1⫻ cleavage buffer {100 mM NaCl, 50 mM HEPES [pH 7.4], 10 mM DTT, 1 mM EDTA, 10% glycerol, 0.1% 3-[(3-cholamidopropyl)-dimethylam-monio]-1-propanesulfonate} and 2.5 U of active recombinant caspases (EMD Millipore) was incubated for 4 h at 37°C. The reaction was stopped by addition of an equal amount of 2⫻SDS–10% 2-ME sample buffer, and the cleavage products were analyzed by Western blotting after separation by SDS-PAGE.
Chemical protein cross-linking.All cross-linking experiments were performed with freshly made solution of disuccinimidyl suberate (DSS; Thermo Scientific) protein cross-linker dissolved in dimethyl sulfoxide (DMSO) as a 50 mM stock solution. Forin vitrocross-linking, recombi-nant ORF57-FLAG protein was diluted in IP buffer. Alternatively, the cells expressing ORF57 protein forin vivocross-linking were resuspended in phosphate-buffered saline (PBS). Both samples were then incubated with increasing amounts of DSS for 30 min at room temperature, along with DMSO (vehicle) used as a negative control. The cross-linking reaction was stopped by addition of 1l of 200 mM Tris-HCl (pH 7.4) and incubated for 15 min at room temperature to quench the cross-linker. The obtained protein complexes were separated by SDS-PAGE and analyzed by Gel Blue staining or by Western blotting.
Inhibition of ORF57 dimerization by synthetic peptides.A dimeriza-tion interface of ORF57 homodimers was mapped by using a library of short synthetic peptides from the ORF57 C terminus (Table 2), which were synthesized by Peptide 2.0 Inc. (Chantilly, VA). The prepared pep-tides were dissolved in water to prepare a 5 mM stock solution. To allow the ORF57-peptide interaction at a ratio of 1:3, 200 ng (⬃5 nM) of re-combinant ORF57 protein was first preincubated with⬃15 nM (3l of 5 mM stock) of individual peptides in a 10-l reaction mixture on ice for 30 min. Subsequently, for peptide dose-response evaluation, the same amount of recombinant ORF57 was incubated with increasing amounts of individual peptides. The formation of ORF57 homodimers was then tested by addition of DSS cross-linker at a final concentration of 500M. The cross-linking reaction was carried out as described above, and the cross-linked ORF57 complexes were analyzed by Western blotting.
In vivonuclear translocation assay.Thein vivonuclear translocation assay was performed as previously described (41). Briefly, 1⫻105COS-1
cells grown of glass coverslips in a 6-well plate were cotransfected with 200 ng of GFP-tagged “reporter” vector together with 400 ng of FLAG- or c-Myc-tagged “tester” plasmid. Twenty-four hours after transfection, the cells were fixed with cold ethanol at⫺20°C for 20 min, followed by im-munofluorescent staining with anti-FLAG M2 or anti-c-Myc antibody (Sigma) in combination with a secondary anti-mouse Alexa Fluor 546-conjugated antibody. Cell nuclei were counterstained with Hoechst 33258
on November 7, 2019 by guest
http://jvi.asm.org/
dye (Sigma) for 30 min at room temperature. The protein localization was determined by using a Zeiss LSM510 META laser-scanning confocal mi-croscope (Zeiss), and the efficiency of translocation was determined as the percentage of double-positive cells with nuclear translocation ORF57-GFP protein.
RT-PCR.The expression levels of ORF57 and glyceraldehyde-3-phos-phate dehydrogenase (GAPDH) RNAs were determined by two-step re-verse transcriptase PCR (RT-PCR) carried out on 1g of total RNA iso-lated with TriPure reagent (Roche) after treatment with Turbo DNase (Ambion). Reverse transcription was performed with murine leukemia virus (MULV) RT in the presence of random hexamers followed by PCR amplification using AmpliTaq (ABI) and the following gene-specific primers: oVM65 (5=-ATGCTCGCAGGGAGTCTGAG-3=) and oVM68 for ORF57 RNA and oZMZ269 (5=-GTCATCAATGGAAATCCCATCACC-3=) and oZMZ270 (5=-TGAGTCCTTCCACGATACCAAA-3=) for cellular GAPDH RNA for sample normalization.
Protein stability assay and proteasome inhibition.To determine ORF57 protein stability, HeLa cells were transfected with vectors express-ing the wild-type (pVM68) or dimerization-defective mutant (pVM85) form of ORF57 protein. Twenty-four hours after transfection, the cells were treated with 50g/ml of cycloheximide (CHX) for 0, 1, 2, or 4 h. At each time point, the cells were harvested directly in 2⫻SDS–2-ME protein sample buffer, and the level of the remaining ORF57 protein was deter-mined Western blot analysis after normalization to neomycin phospho-transferase II or cellular-tubulin. To inhibit ORF57 proteasome-medi-ated degradation, ORF57-expressing HeLa cells were treproteasome-medi-ated 24 h after transfection with 20M proteasome inhibitor MG132 (Sigma) for 4 to 6 h or with DMSO (vehicle, negative control). The cells were then either lysed directly in SDS protein sample buffer for Western blotting or in RSB-150 buffer for IP analysis.
RESULTS
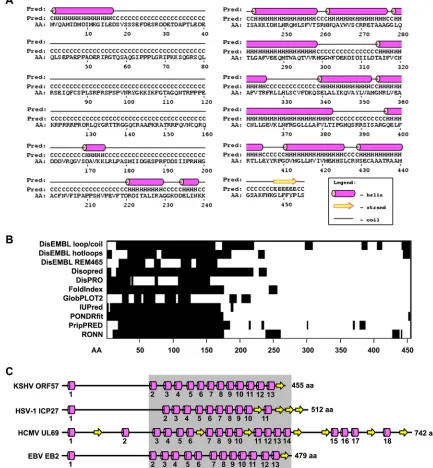
Secondary structure of KSHV ORF57 is highly conserved
among its homologues.The secondary structure of the KSHV
ORF57 protein (GenBank accession numberYP_001129410.1)
predicted by PSIPRED (version 3.0) displays 13␣-helices derived
from 170 residues, representing⬃37.3% of ORF57 amino acid
residues, and one-sheet derived from only 6 aa residues,
count-ing for⬃1.3% of ORF57 amino acid residues (Fig. 1A). The
ma-jority (279, or⬃66.3% of total ORF57 amino acid residues) of
ORF57 are random coils lacking any secondary structure. The
identified␣-helices are unevenly distributed in the ORF57
poly-peptide and are mostly clustered on the C-terminal half of ORF57,
with only one␣-helix at the very end (aa 2 to 16) of the N terminus
of ORF57. This leaves the N-terminal half of the ORF57 protein mostly unstructured. We noticed that the predicted ORF57 struc-ture is slightly different from that produced with other prediction
algorithms (data not shown) and from the published one (15)
because an updated version of PSIPRED (version 3) was used in this study.
A protein lacking the secondary structure generally indi-cates the presence of “intrinsically disordered” regions (IDR) with a characteristic sequence composition that contains high numbers of polar residues to cause low hydrophobicity and a high net charge. These features prevent the IDR from folding spontaneously into stable tertiary structures, thereby leading to “natively unfolded” domains with an extended conformation
and high flexibility (for a review, see reference42). Analyses of
ORF57 by 11 available prediction algorithms (Fig. 1B) all
con-sistently gave the same long IDR in the ORF57 N terminus
covering approximately 200 aa residues. Also shown inFig. 1B,
a few shorter IDRs were predicted in the other parts of the ORF57 protein, but their predictions varied from one algo-rithm to another.
Despite some functional conservation, ORF57 homologues share very low amino acid sequence homology. To evaluate the possible structural similarities among ORF57 homologues, the secondary structures of HSV-1 ICP27 protein (GenBank
accession numberBAE44982.1) from the alphaherpesviruses,
HCMV UL69 (GenBank accession numberACZ79980.1) from
from the betaherpesviruses, and EBV EB2 (GenBank accession
numberYP_401659.1) from gammaherpesviruses were predicted
by PSIPRED (see Fig. S1A to C in the supplemental material). The predicted secondary structures from these three ORF57
homo-logues (Fig. 1C) showed high similarity to the ORF57 structure
and were characterized by a low density of structural motifs
en-riched in␣-helices versus-sheets at 11:4 in ICP27, 18:7 in UL69,
and 13:1 in EB2 and an unevenly clustered distribution of the structural motifs in their C termini. As expected, the secondary structure of EBV EB2 closely resembles that of KSHV ORF57. ICP27 deviates slightly from ORF57, but UL69, the largest
homo-logue, exhibits an additional 4␣-helices and 3-sheets from its
unusually long C terminus. Together, we conclude that the KSHV ORF57 protein and its homologues all contain two structurally distinguishable domains, an “intrinsically disordered”
N-termi-nal domain and a structured␣-helix-rich C-terminal domain.
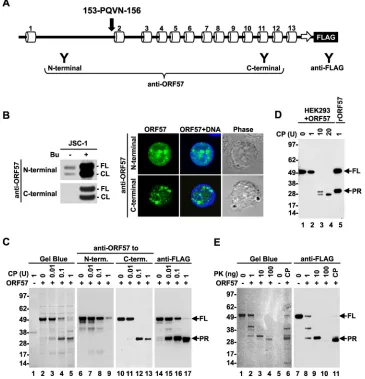
Limited proteolysis of the ORF57 protein.Limited
proteoly-sis has been widely used to determine the conformation of both
isolated proteins and protein complexes (43). The principle of this
[image:4.585.41.551.77.184.2]approach is to observe protease digestion of only flexible, unstruc-tured protein regions, not the rigid globular domains, at low con-centrations of proteases. This allows the identification of protein domains based on their distinct structures. The limited proteolysis
TABLE 2ORF57-derived small synthetic peptides tested for inhibition of ORF57 homodimerization
Peptide name Sequence Length (aa) Positiona Charge pI
vm1 IDGESPRFDDSIIPRHHGA 19 183–201 0 (neutral) 5.41
vm2 FTDRDITALIRAGGKDDE 18 218–235 ⫺2 (acidic) 4.19
vm3 NRLVEACNLLGEVKLNFR 18 355–372 1 (basic) 8.55
vm4 TSRHNQAYWVSCRRETAAAGG 21 258–278 3 (basic) 9.50
vm5 QTLGAFVEEQMTWAQTVVRH 20 280–299 0 (neutral) 5.48
vm6 TRFRLLHLSCVFDKQSE 17 324–340 2 (basic) 8.56
vm7 RRSISARGQELFRTLLEYYR 20 389–408 3 (basic) 11.09
vm8 EHHSLCRNSECAAATRAA 18 422–439 2 (basic) 7.42
vm9 TRAAMGSAKFNKGLFFYPLS 20 436–455 3 (basic) 10.90
aThe peptide position represents its amino acid residues in the KSHV ORF57 protein.
on November 7, 2019 by guest
http://jvi.asm.org/
of ORF57 was first carried out using purified recombinant ORF57-FLAG protein and clostripain, an arginine-specific pro-tease (endoproteinase Arg-C). ORF57 contains 45 arginine resi-dues, the most frequent residue, accounting for almost 10% of all amino acid residues in ORF57. These arginine residues are evenly distributed along the entire ORF57 polypeptide, with the longest
interval between two arginines being⬃28 residues. After
diges-tion with increasing amounts of clostripain, the cleavage products
of ORF57 were resolved by SDS-PAGE (Fig. 2C) and stained with
Gel Blue for total protein determination. This assay revealed a
progressive digestion of the full-length (FL) ORF57 (⬃50 kDa) by
clostripain along with the appearance of a new protein band of
approximately 30 kDa (Fig. 2C, lanes 3 to 5), which is smaller than
clostripain, of size⬃38 kDa (Fig. 2C, lane 1). The data indicated
that the 30-kDa band was from ORF57 and might represent the protease-resistant region (PR) of the ORF57 protein.
Subsequently, we analyzed the PR band derived from ORF57 by Western blotting using a set of anti-ORF57 antibodies
specifi-FIG 1Predicted secondary structure of KSHV ORF57 protein and its conservation among ORF57 homologues. (A) Diagram of the predicted secondary structure of the ORF57 protein based on the PSIPRED algorithm. C, coil; H,␣-helix; E,-sheet/strand. (B) Positions and sizes of the intrinsically disordered regions (black boxes) in the ORF57 protein predicted by multiple algorithms (shown on the left). (C) Conservation of the secondary structure among ORF57 homologues HSV-1 ICP27, HCMV UL69, and EBV EB2, based on PSIPRED prediction (see also Fig. S1 in the supplemental material). Numbers under each ␣-helix represent the relative positions from the N to C terminus. Numbers at the end of each protein represent the protein length in amino acid residues. The gray box marks the region with increased sequence conservation. The drawings are not to scale.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.71.504.73.541.2]cally designed to target either the N- or C-terminal regions of
ORF57 (Fig. 2A). Despite both antibodies recognizing full-length
and caspase-7-cleaved ORF57 in butyrate-activated JSC-1 cells,
based on a Western blotting assay (Fig. 2B, left panel) and
immu-nofluorescent staining (Fig. 2B, right panel), only the anti-ORF57
C-terminal antibody, not the anti-ORF57 N-terminal antibody,
reacted with the PR band (Fig. 2C, compare lanes 11 to 13 versus 7
to 9). We noticed that the full-length ORF57 level gradually
de-creased, along with the increasing doses of clostripain (Fig. 2C,
lanes 6 to 13), and was accompanied by an increasing level of the
PR product (Fig. 2C, lanes 10 to 13). These data demonstrate that
the PR product from clostripain digestion represents the C
termi-nus of the ORF57 protein. This conclusion was further verified with an anti-FLAG antibody against the FLAG tag fused to the
ORF57 C terminus (Fig. 2C, lanes 14 to 17). To map the N
termi-nus of the PR region, the cleaved PR products were sequenced by Edman protein sequencing; we identified PQVN as the last four N-terminal amino acid residues of the PR product, which mapped to aa 153 to 156 of ORF57, with two arginines at aa 151 to 152
immediately upstream, as the clostripain digestion site (Fig. 1A
and2A).
To confirm that ORF57 forms a similar structure under native conditions in cells, ORF57-FLAG was expressed in HEK293 cells and the whole-cell extract was used in a clostripain digestion at
FIG 2Mapping of the ORF57 protein structure based on limited proteolysis. (A) Diagram of the ORF57 secondary structure based on PSIPRED prediction (see Fig. 1A), with a FLAG tag on the ORF57 C terminus and positions of the epitopes (Y) for separate anti-ORF57 and anti-FLAG antibody recognition. Positions and sequences of the residues as detected by Edman protein sequencing of the ORF57 PR region (PR) are shown above␣-helix 2. (B) Western blot analysis of ORF57 in uninduced or 3 mM butyrate-induced JSC-1 cells with an anti-ORF57 N- or C-terminal antibody (left panel). FL, full-length; CL, cleaved. Indirect immunofluorescence staining of the butyrate-activated JSC-1 cells (right panel) was performed using an anti-N- or C-terminal ORF57 antibody in combination with an Alexa Fluor 488-conjugated secondary antibody (green). The cell nuclei were counterstained with Hoechst 33258 (blue). (C) Limited proteolysis of recombinant ORF57-FLAG protein with increasing amounts of clostripain (CP). ORF57 without digestion was used as a negative control. The cleavage products were separated by SDS-PAGE and visualized with Gel Blue staining (left panel) or by Western blotting using anti-ORF57 to the N terminus or C terminus and anti-FLAG antibodies (right panels). (D) Clostripain digestion of ectopic ORF57-FLAG protein in cell extract from HEK293 cells prepared 24 h after transfection with a pVM7 vector. The cleavage products were analyzed by Western blotting using an anti-FLAG antibody. rORF57, recombinant ORF57. (E) Cleavage of recombinant ORF57-FLAG protein with increasing amounts of proteinase K (PK). ORF57 cleaved with 1 U of clostripain served as a control. The cleaved products were analyzed with Gel Blue staining and by Western blotting using an anti-FLAG antibody as described for panel C. FL, full length. Numbers on the left are molecular markers (in kDa).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.110.475.70.449.2]higher doses. Western blot analysis of the cleavage products with an anti-FLAG antibody revealed PR products with similar sizes as
those derived from the recombinant ORF57 (Fig. 2D, compare
lane 3 to lane 5), indicating that the ORF57 protein expressed in the cells in the presence of other cellular proteins displays a similar structure as recombinant ORF57 protein. However, we saw an additional band derived from further digestion of the PR products
with increasing doses of clostripain (Fig. 2D, lanes 3 and 4).
Fi-nally, we confirmed the clostripain-specific digestion of ORF57 and PR production by the limited proteolysis of recombinant ORF57 protein by using a more promiscuous proteinase K, a broad-spectrum serine protease expected to cleave ORF57 at 201
sites, as predicted with the ExPASy PeptideCutter software (http:
//web.expasy.org/peptide_cutter). Similar to clostripain, the in-creased amounts of proteinase K led to progressive digestion of full-length ORF57 and increased production of the PR product
similar in size to the one from clostripain digestion (Fig. 2E,
com-pare lanes 4 and 6). Moreover, these PR products from proteinase K digestion were also recognizable with an anti-FLAG antibody (Fig. 2E, lanes 7 to 11). Collectively, our data from limited prote-olysis using two proteases in combination with Edman protein sequencing indicate that the ORF57 conformation consists of an unstructured, protease-sensitive N terminus from aa 1 to 152 and a structured protease-resistant C terminus from aa 153 to 455.
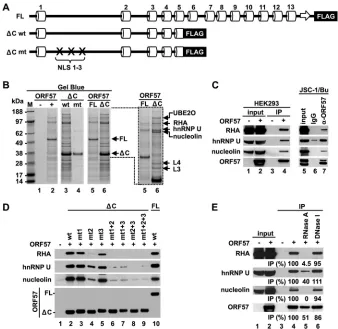
The N terminus of ORF57 is essential for interaction with
RNA binding proteins.The flexibility and accessibility of the IDR
would be characteristic for high binding potentials (44). To test
the identified IDR in the ORF57 N terminus for its interaction with cellular proteins, the full-length (FL) ORF57 protein was
compared with a C-terminal-truncated ORF57 (⌬C ORF57)
con-taining only the N-terminal half (aa 1 to 251) (Fig. 3A) for the
binding activities in the presence of HEK293 total cell extract by co-IP. We found that the protein binding profiles between FL
ORF57 and ⌬C ORF57 were similar (Fig. 3B), but a group of
proteins subsequently identified by MS analysis, including UBE2O (ubiquitin-conjugating enzyme E2O), RHA (RNA heli-case A), hnRNP U (heterogeneous nuclear ribonucleoprotein U), nucleolin, and ribosomal proteins L4 and L3, appeared to
prefer-entially bind to⌬C ORF57 (Fig. 3B, lanes 5 and 6). The binding of
such proteins may vanish by the introduction of point mutations
into both NLS 1 and 2 of the⌬C ORF57 (Fig. 3B, compare lanes 4
and 3). We previously showed that the simultaneous mutations in any two of three ORF57’s NLSs would be detrimental for ORF57 function, even though the mutant ORF57 (mtORF57) remains a
nuclear protein (14). Together, these data indicate that the
N-ter-minal IDR of ORF57 is a major domain that interacts with multi-ple cellular proteins, which might be affected minimally by the C-terminal structural domain of ORF57.
RHA, hnRNP U, and nucleolin were subsequently chosen for further verification. We confirmed their interactions with both ectopic ORF57 in HEK293 cells and native ORF57 in
KSHV-in-fected JSC-1 cells by co-IP (Fig. 3C, lanes 4 and 7). Further
anal-yses of⌬C ORF57 with point mutations in single, double, or triple
NLS motifs for the binding of RHA, hnRNP U, and nucleolin
demonstrated that, despite⌬C ORF57 exhibiting a comparable
binding activity with FL ORF57 for these three proteins, the NLS2
in⌬C ORF57 appears to play a major role for binding of RHA and
nucleolin, and introduction of point mutations into the NLS2
remarkably reduced the binding activities of⌬C ORF57 (Fig. 3D,
lane 4). Introduction of point mutations into the NLS1 or NLS3 of
⌬C ORF57 did not affect its interaction with any of the three
proteins (Fig. 3D, compare lane 2 versus lanes 3 and 5). In
con-trast,⌬C ORF57 with combined mutations in both NLSs or all
three NLSs lost the binding activity for all three proteins (Fig. 3D,
lanes 6 to 9), which correlates with the reported activity of ORF57 in the enhancement of intronless KSHV ORF59 expression and of
KSHV K8 splicing (5,14).
Given that ORF57 in KSHV-infected cells is often associated with specific ribonucleoprotein complexes, including RNAs and
proteins (15), the role of nucleic acids in the ORF57 interaction
with RHA, hnRNP U, or nucleolin was tested in pulldown assays using HEK293 cell extract with or without RNase A or DNase I treatment. As expected, DNase I digestion of the cell extract pro-duced no effect on the ORF57 association with RHA, hnRNP, or
nucleolin (Fig. 3E, compare lane 6 to lane 4), but RNase A
diges-tion of the cell extract almost completely disassociated ORF57
from RHA and nucleolin (Fig. 3E, compare lane 4 to lane 5),
indicating that their interaction with ORF57 is RNA mediated. In
contrast, such RNase A digestion of the cell extract led to⬃60%
reduction of hnRNP U from association with ORF57 (Fig. 3E, lane
5), suggesting the presence of a partial protein-protein interaction between ORF57 and hnRNP U. Together, our data indicate that the disordered region in the ORF57 N terminus plays an essential role in ORF57 interactions with a wide range of cellular factors, either directly or indirectly.
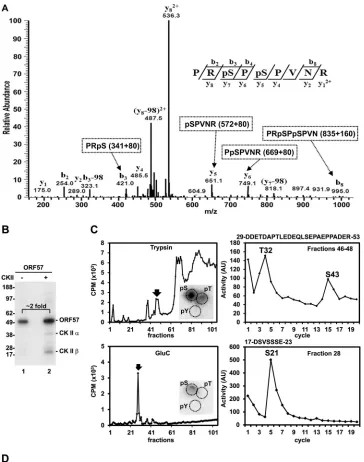
The N-terminal region of ORF57 is posttranslationally
mod-ified by phosphorylation.Posttranslational protein modification
represents an important regulatory mechanism for many cellular processes. It has been demonstrated by proteome analysis that the IDR regions of a given protein are rich in posttranslational
mod-ifications, such as phosphorylation (45,46). The KSHV ORF57
protein is phosphorylated by cellular CKII, but its
phosphoryla-tion sites remain to be identified (47). To experimentally map
ORF57 phosphorylation sites, we took two complementary meth-ods. First, a mass spectrometry analysis of phosphopeptides gen-erated by trypsin digestion of recombinant ORF57 protein led to the identification of the phosphopeptide 93-PRpSPpSPVNR-101, which shows the phosphorylation of ORF57 at S95 and S97
resi-dues (Fig. 4A). Web-based prediction of kinase phosphorylation
sites in ORF57 by using the NetPhosK 1.0 server predicted glyco-gen synthase kinase 3 (GSK3) as a potential kinase responsible for the phosphorylation of both the S95 and S97 residues, p38MAPK kinase for S95 and ribosomal S6 kinase (RSK) for S97. Unexpect-edly, neither of the two phosphorylation sites identified in ORF57 were predicted to be the sites phosphorylated by previously
re-ported CKII (47).
Therefore, in our next analysis, the recombinant ORF57
pro-tein was first incubatedin vitro with recombinant CKII in the
presence of radiolabeled [␥-32P]ATP. This led toin vitro
phos-phorylation of ORF57 and autophosphos-phorylation of CKII␣and
subunits (Fig. 4B, lane 2). We also unexpectedly observed ORF57
phosphorylation in the absence of CKII, with⬃2-fold-lower
in-tensity than that in the presence of CKII (Fig. 4B, compare lanes 1
and 2). Thisin vitroautophosphorylation of the ORF57 protein
appeared to occur in the absence of any other phosphorylated
protein (Fig. 4B, lane 1). To further map the positions of
phos-phorylation sites, the CKII-phosphorylated ORF57 protein was digested with trypsin or endoproteinase GluC, and the fraction-ated phosphopeptides with high activity were subjected to Edman peptide sequencing and phosphoamino acid analysis. This led to
on November 7, 2019 by guest
http://jvi.asm.org/
identification of three additional ORF57 phosphorylation sites at the S21, T32, and S43 residues. Phosphoamino acid analysis of the fractionated phosphopeptides confirmed the specific phosphory-lation of serine (Ser or S) and threonine (Thr or T), but not ty-rosine (Tyr or Y), which is in agreement with CKII being a serine/
threonine-selective protein kinase (Fig. 4C, insets). Interestingly,
the same analyses of autophosphorylated ORF57 protein in the absence of CKII also identified the phosphorylation of ORF57 at the S21, T32, and S43 residues (data not shown). The similarity of these mapped phosphorylation sites to the ones in CKII-phos-phorylated ORF57 suggests ORF57 phosphorylation by a residual amount of insect CKII copurified with the recombinant ORF57 protein from Sf21 cells, as functional conservation of CKII has
been reported among eukaryotes (48). Nevertheless, this study led
us to experimentally identify five phosphorylation sites from the
ORF57 N terminus (Fig. 4D) which are all localized in the IDR
region, a preferential region for posttranslational modification of the ORF57 protein.
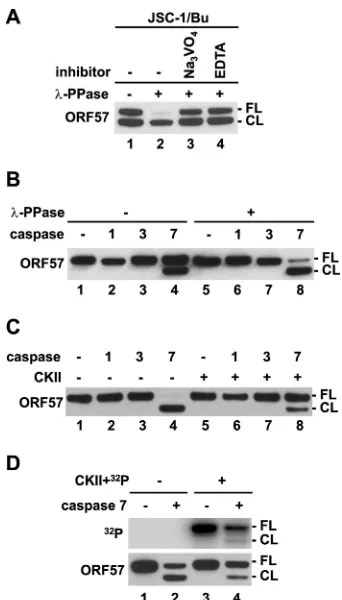
Phosphorylation and caspase-7 cleavage of KSHV ORF57. Protein phosphorylation is a reversible posttranslational modifi-cation which affects protein localization, folding, binding activi-ties, stability, etc. It has been shown that phosphorylation
modu-lates substrate recognition by cellular caspases (49). The KSHV
ORF57 protein contains a caspase-7 cleavage site at aspartic resi-due D33, and its cleavage by caspase-7 produces a truncated,
func-tionally attenuated form of ORF57 (38). Given that all identified
CKII phosphorylation sites in ORF57 reside in close proximity to the caspase-7 cleavage site, with phosphorylation site T32 being positioned directly within the caspase-7 recognition site 30-DETD-33, we examined how phosphorylation and
dephosphory-FIG 3The intrinsically disordered N terminus of the ORF57 protein mediates binding of ORF57 to cellular cofactors. (A) Diagrams of the secondary structure of full-length ORF57-FLAG (FL) and C-terminally truncated ORF57-FLAG (⌬C, aa 1 to 251) with wild-type (wt) or mutated (mt) NLS. (B) Binding activities of the ORF57 proteins described for panel A. Total cell extract from HEK293 cells transfected with individual ORF57-expressing vectors or an empty vector was used for IP. ORF57 complexes immunoprecipitated with an anti-FLAG antibody were resolved by SDS-PAGE and visualized by Gel Blue staining. M, molecular markers (in kDa). Enlarged portion from lanes 5 and 6 shows the most abundant proteins copurified with the⌬C ORF57 protein, identified by mass spectrom-etry. UBE2O, ubiquitin-conjugating enzyme E2O; RHA, RNA helicase A; hnRNPU, heterogeneous nuclear ribonucleoprotein U; L3 & L4, ribosomal proteins L3 and L4. (C) Interactions of ectopic ORF57 from HEK293 cells or endogenous ORF57 from JSC-1 cells with RHA, hnRNP U, and nucleolin, determined by IP with an anti-FLAG (left) or anti-ORF57 (right) antibody. The cells without ORF57 expression or with nonspecific immunoglobulins (IgG) for IP were used as negative controls. The immunoprecipitated proteins were analyzed by Western blotting using appropriate antibodies. Lytic KSHV infection in JSC-1 cells was activated by sodium butyrate (Bu). (D) Role of three nuclear localization signals (labeled as 1, 2, and 3) in the ORF57 interaction with RHA, hnRNP U, and nucleolin. Western blot analysis was conducted for RHA, hnRNP U, and nucleolin pulled down by an anti-FLAG antibody from HEK293 cells expressing three different forms of ORF57-FLAG fusion proteins (FL-wt,⌬C-wt, or⌬C-mt). The cells transfected with an empty vector were used as a negative control for IP specificity. (E) Role of RNA in ORF57 interaction with cellular cofactors. ORF57-associated proteins were immunoprecipitated from total cell extract of HEK293 cells expressing ORF57-FLAG by using an anti-FLAG antibody without or with prior treatment with RNase A or DNase I. Relative amount (as a percentage) of the immunoprecipitated proteins from nuclease-treated cell extract was determined by Western blotting compared to that from nuclease-untreated extract.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.122.460.65.394.2]lation of ORF57 regulate ORF57 cleavage by caspase-7. The whole-cell extract prepared from KSHV-infected JSC-1 cells ex-pressing ORF57 after lytic reactivation by sodium butyrate was
treated with nonspecific-PPase. Because KSHV reactivation in
JSC-1 cells activates cellular caspases to cleave ORF57 protein
(38), JSC-1 extract without -PPase treatment usually exhibits
two forms of ORF57, an FL form and a faster-migrating cleaved
form (CL), at a ratio of approximately 1:1 (Fig. 5A, lane 1). We
FIG 4Phosphorylation sites identified in KSHV ORF57. (A) Identification of ORF57 phosphorylation sites by mass spectrometry. MS/MS analysis of abundance and mass ofbandyions obtained by fragmentation of ORF57 phosphopeptide aa 93 to 101 identified phosphorylated serine residues at S95 and S97. The protein sequences corresponding to individual ions and calculation of their masses are shown in the dashed boxes, with a mass of the phosphogroup equal to 80 Da. (B) SDS-PAGE autoradiograph ofin vitro-phosphorylated recombinant ORF57-FLAG protein by recombinant CKII in the presence of radiolabeled [␥-32P]ATP.
Numbers on the left represent the positions of protein markers (in kDa). (C) Mapping of CKII phosphorylation sites in ORF57.In vitro-phosphorylated recombinant ORF57 protein was digested with trypsin (top) or endoproteinase GluC (lower), and the resulting phosphopeptides were purified by reverse HPLC based on their activity (left). The phosphopeptides fractionated with high activity (marked by arrows in the left panels) were separately subjected to Edman protein sequencing (right panels) or phosphoamino acid analysis (insets). The activity (in arbitrary units [AU]) of each Edman sequencing cycle is shown on the right, together with the corresponding peptide sequence, which is shown above each panel. Insets in the left panels show phosphoamino acid analysis of the CKII-phosphorylated, trypsin-digested, or GluC-digested recombinant ORF57 fragments in the combined fractions from 46 to 48 or in the fraction 28 by 2D chromatography. pS, phosphoserine; pT, phosphothreonine; pY, phosphotyrosine. (D) A partial protein sequence of ORF57, with all five phosphorylated residues identified by two methods.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.111.475.65.527.2]found that-PPase treatment of the same extract strongly
en-hanced ORF57 cleavage (Fig. 5A, lane 2), and two-PPase
inhib-itors, sodium orthovanadate and EDTA, blocked the
-PPase-induced ORF57 cleavage (Fig. 5A, compare lane 2 to lanes 3 and
4), indicating a direct effect of dephosphorylation on ORF57 cleavage. To further confirm this observation, recombinant
ORF57 protein was treated with or without-PPase followed byin
vitrocleavage assays with recombinant caspase-1, -3, or -7 (Fig.
5B). As expected, ORF57 in the absence of-PPase pretreatment
was cleaved by caspase-7 by⬃50% (Fig. 5B, compare lane 1 to lane
4), whereas dephosphorylation of the ORF57 by-PPase
signifi-cantly increased ORF57 cleavage (to⬃90%) by caspase-7 (Fig. 5B,
compare lane 5 to lane 8). Under both conditions, ORF57 was not
cleavable by caspase-1 or -3 (Fig. 5B, lanes 1 to 3 and 5 to 7). These
data suggest that protein dephosphorylation promotes ORF57 cleavage specifically by caspase-7.
To further confirm the inhibitory effect of phosphorylation on ORF57 cleavage by capsase-7, recombinant ORF57 was first
phos-phorylatedin vitroby CKII followed by caspase cleavage with
re-combinant caspase-1, -3, or -7 (Fig. 5CandD). Again, ORF57
without CKII treatment was cleaved by caspase-7, but not by
caspase-1 or -3 (Fig. 5C, lanes 1 to 4, andD, lane 2). As expected,
in vitrophosphorylation of ORF57 by CKII inhibited the cleavage
of ORF57 by caspase-7 (Fig. 5C, lane 4 versus lane 8, and D, lane 2
versus lane 4). Moreover, we found that, in the presence of [␥
-32
P]ATP, CKII predominantly mediated the incorporation of
␥-32P into the FL ORF57 to protect it from caspase-7 cleavage, and
not much so into the cleaved ORF57 (Fig. 5D, compare top panel
versus lower panel in lane 4), suggesting that other phosphoryla-tion sites remaining in the cleaved ORF57 are less sensitive to CKII kinase. In summary, this study demonstrated that phosphoryla-tion in the ORF57 N-terminal IDR inhibits ORF57 cleavage by cellular caspase-7 and plays an important role in regulation of ORF57 protein stability.
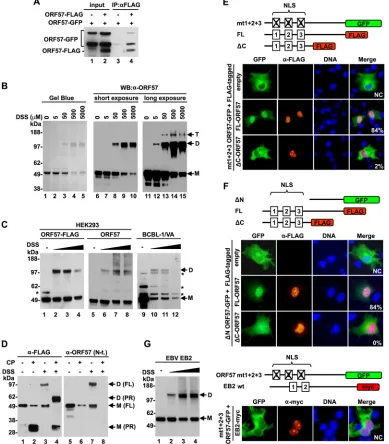
ORF57 homodimerization via its structural C terminus.We
next wished to determine the role of the structural C terminus in ORF57 self-interaction and function, because KSHV ORF57 and its homologues have been shown to form multimeric complexes
by self-interaction via their C terminus (41, 50–52). However,
these studies did not provide any hard evidence to show ho-modimerization of ORF57. To do this, we first repeated the asso-ciation of two forms of ORF57 proteins tagged either with GFP or with FLAG by co-IP after their coexpression in HEK293 cells. Western blotting with an anti-ORF57 N terminus antibody that recognized both forms of ORF57 revealed efficient pulldown of ORF57-GFP by an anti-FLAG antibody from the cells fected with ORF57-FLAG, but not from control cells
cotrans-fected with an empty vector (Fig. 6A, compare lane 3 to lane 4),
indicating the association of ORF57-GFP with ORF57-FLAG in the cells. To exclude the possibility that the observed association between two tagged ORF57 proteins was not mediated by other
cellular proteins, we used recombinant ORF57 protein in anin
vitrohomodimerization assay. The recombinant ORF57 protein diluted in IP buffer was incubated at room temperature, and the expected ORF57 homodimers were stabilized by the addition of DSS, a noncleavable bivalent chemical cross-linker that is com-monly used to detect direct protein-protein interactions. The re-sulting complexes were resolved by SDS-PAGE. Gel Blue protein
staining showed ORF57 monomers (M) of⬃50 kDa without DSS
(Fig. 6B, lane 1). With increasing doses of DSS, the intensity of ORF57 monomers reduced gradually along with the appearance
of the predicted ORF57 homodimers, of⬃100 kDa (Fig. 6B, lanes
2 to 5). Both ORF57 monomers and dimers were specifically
de-tectable with an anti-ORF57 N-terminal antibody (Fig. 6B, lanes 6
to 15). A longer exposure time for the Western blot assay revealed
a small amount of ORF57 homotrimers of⬃150 kDa (Fig. 6B,
lanes 13 to 15). Since DSS is membrane permeable, the formation of an ORF57 dimer was also examined in HEK293 cells expressing ectopic ORF57 with or without a C-terminal FLAG tag or in BCBL-1 cells with KSHV lytic infection and native ORF57 expres-sion. As expected, Western blotting with an anti-ORF57 antibody showed that the samples without DSS contained only monomeric
ORF57 (Fig. 6C, lanes 1, 5, and 9), whereas DSS treatment led to
the formation of ORF57 homodimers in both types of cells
FIG 5Phosphorylation inhibits ORF57 cleavage by caspase-7. (A and B) De-phosphorylation of ORF57 promotes caspase cleavage of ORF57. (A) Cell ex-tracts from JSC-1 cells induced with sodium butyrate (Bu) were treated with -PPase in the absence or presence of -PPase inhibitors orthovanadate (Na3VO4) or EDTA. Cell extract without any treatment served as a negative
control. The cleaved ORF57 was detected by Western blotting using an anti-ORF57 N-terminal antibody. FL, full length anti-ORF57; CL, cleaved anti-ORF57. (B) Recombinant ORF57-FLAG protein with or without-PPase pretreatment was incubated with recombinant caspase-1, -3, or -7, and the cleavage prod-ucts were detected by Western blotting as described for panel A. ORF57 pro-tein without caspase digestion served as a negative control. (C and D) Phos-phorylation of ORF57 inhibits caspase cleavage. Recombinant ORF57-FLAG protein without or with priorin vitrophosphorylation by recombinant CKII was cleaved with recombinant caspase-1, -3, or -7 and analyzed by Western blotting as described for panel A. Autoradiograph (upper panel) of caspase-7-digested recombinant ORF57 without or with phosphorylation by CKII plus [␥-32P]ATP (D, upper panel). The same membrane was then blotted with an
anti-ORF57 antibody (D, lower panel). ORF57 without caspase digestion served as a control.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:10.585.77.248.64.364.2]FIG 6Homodimerization of ORF57 via the structured C-terminal domain. (A) Self-interaction of ORF57in vivo. ORF57-GFP and ORF57-FLAG fusion proteins were coexpressed in HEK293 cells. ORF57-FLAG was immunoprecipitated with an anti-FLAG antibody, and proteins in the pulldown were analyzed by Western blotting with an anti-ORF57 N-terminal antibody. Cells cotransfected with ORF57-GFP and empty FLAG vectors (-) served as negative controls. (B) ORF57 homodimerizationin vitro. Recombinant ORF57-FLAG protein was incubated with increasing amounts of the bivalent chemical cross-linker DSS. The cross-linked complexes were resolved by SDS-PAGE and analyzed with Gel Blue staining or by Western blotting using an anti-ORF57 N-terminal antibody. M, monomer; D, dimer; T, trimer. (C) ORF57 homodimerizationin vivo. HEK293 cells expressing ORF57 with or without a C-terminal FLAG tag or KSHV-infected, valproic acid (VA)-activated BCBL-1 cells were incubated with increasing concentrations (50, 500, and 5,000M) of DSS, and the cross-linked proteins were analyzed by Western blotting with an anti-ORF57 N-terminal antibody. *, uncharacterized protein band. (D to F) Role of the structured C-terminal domain in ORF57 homodimerization. (D) Proteolytic digestion of recombinant ORF57-FLAG protein with clostripain (CP) with or without cross-linker DSS. The cleavage products were detected by Western blotting with an anti-FLAG or anti-ORF57 N-terminal (N-t.) antibody. ORF57 without CP digestion or DSS cross-linking served as negative controls. M, monomer; D, dimer; FL, full length. (E and F) Results of ORF57 translocation assays, with the upper panels showing the diagrams of ORF57 proteins fused with GFP or FLAG tags (color boxes). Numbered boxes in ORF57 are three NLSs, with a cross indicating its mutation. ORF57 localization in COS-1 cells cotransfected with a GFP-tagged ORF57 mutant (mt1⫹2⫹3 ORF57-GFP) or⌬N ORF57 (⌬N ORF57-GFP) as a GFP reporter together with a FLAG-tagged full-length or⌬C ORF57 vector. Cells cotransfected with the GFP reporters and an empty FLAG vector served as negative controls. Twenty-four hours after transfection, the cells were fixed and immunostained with an anti-FLAG antibody and then an Alexa Fluor 546-labeled secondary antibody (red). Localization of the ORF57-GFP fusion was detected with direct fluorescence (green) of the cells. The cell nuclei were counterstained with Hoechst 33258 dye (blue). Fifty cells positive for the coexpression of both ORF57-GFP and ORF57-FLAG proteins on each slide were examined for subcellular localization of the ORF57-GFP fusion. The percentage of cells positive for nuclear localization of ORF57-GFP due to dimerization of ORF57-GFP with ORF57-FLAG is shown in the right corner. NC, not counted. (G) EBV EB2 can be a homodimer, but it does not heterdimerize with KSHV ORF57. (Left) Formation of EB2 homodimer in HEK293 cells expressing c-myc-tagged EB2 protein. The cells at 24 h after transfection were harvested in PBS and incubated with increasing concentrations (50, 500, and 5,000M) of DSS forin vivocross-linking. EB2 complexes were detected by Western blotting using anti-c-myc monoclonal antibody. (Right) Cytoplasmic ORF57-GFP translocation assays with myc-tagged EBV EB2. COS-1 cells were cotransfected with the cytoplasmic ORF57-GFP (mt1⫹2⫹3 GFP) reporter along with c-myc-tagged EBV EB2 protein (EB2-myc). Twenty-four hours after transfection, nuclear localization of ORF57-GFP or EB2-myc was examined by immunofluorescent staining using an anti-FLAG or anti-c-Myc antibody in combination with an Alexa Fluor 546-labeled secondary antibody (red). Cell nuclei were counterstained with Hoechst dye (blue).
on November 7, 2019 by guest
[image:11.585.99.488.38.482.2](Fig. 6C, lanes 2 to 4, 6 to 8, and 10 and 11). We were unable to detect ORF57 homotrimers in the cells, presumably because of its
much lower abundance. We noticed that DSS at 5 mM forin vivo
cross-linking sometimes induced large protein aggregates that were hardly separable by SDS-PAGE.
The contribution of the structural C terminus to ORF57
dimerization was further investigatedin vitroby DSS cross-linking
followed by clostripain digestion. We assumed that, if ORF57 ho-modimerization were mediated by its C terminus, proteolytic re-moval of the N terminus of ORF57 after DSS cross-linking would result in a smaller homodimer derived from the C terminus. In-deed, Western blot analysis of the cross-linked ORF57 after clos-tripain digestion with an anti-FLAG antibody to the ORF57
C-ter-minal FLAG tag identified a dimer⬃60 kDa derived from two
cross-linked C termini (PR), each⬃30 kDa (Fig. 6D, compare
lane 4 to lane 2). This dimer derived from the C termini after clostripain digestion was not detectable by Western blotting with
an anti-ORF57 N terminus antibody (Fig. 6D, lane 8), further
confirming its origin.
Using anin vivonuclear translocation assay as our third
ap-proach to verify ORF57 dimerization in cells, we conducted the assay with a cytoplasmic “reporter” protein lacking a functional
NLS due to a mutation (mt1⫹2⫹3 ORF57-GFP) or deletion of
166 aa from the ORF57 N terminus (⌬N ORF57-GFP)
coex-pressed with a “tester” protein that contained a functional NLS
(ORF57-FLAG or⌬C ORF57-FLAG) in COS-1 cells (Fig. 6EandF
protein diagrams) (14). Analysis of 50 cells positive for the
coex-pression of both ORF57-GFP and ORF57-FLAG proteins showed
that ORF57-GFP reporters were efficiently (⬃84%) translocated
from the cytoplasm to the nucleus of COS-1 cells in the coexpres-sion of the full-length ORF57-FLAG, but not much so in the
co-expression of the⌬C ORF57-FLAG (Fig. 6EandF). The nuclear
translocation was ORF57 specific, because EBV EB2, a close ORF57 homologue from human gammaherpesvirus, did not show such a function, although EBV EB2 homodimerizes in
trans-fected cells (Fig. 6G). Together, these data provide convincing
evidence that the ORF57 C terminus is essential for formation of ORF57 homodimers but incapable of forming a heterodimer with EBV EB2.
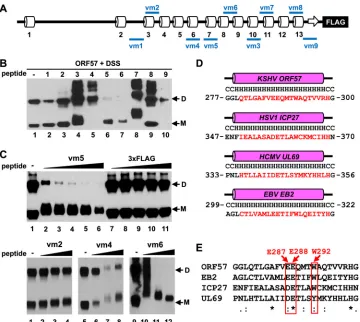
ORF57 homodimerization via the C-terminal␣-helices 7 to
9.Considering that small synthetic peptides derived from
interac-tion interface can inhibit protein-protein interacinterac-tions (53,54) and
in the interest of identifying the ORF57 dimerization interface, we synthesized a set of 9 peptides from various regions of the ORF57
C terminus targeting both structural (␣-helices) and unstructural
(loops) regions (Fig. 7A; see also Fig. S2 in the supplemental
ma-terial) using the following criteria: maximum length of 21 aa res-idues with a minimal 30% hydrophobic resres-idues to enable high
solubility in water (seeTable 2for details). The activity of
individ-ual peptides was tested in thein vitrodimerization assay described
above, using recombinant ORF57 protein. As shown inFig. 7B,
peptides vm1 and vm2 exhibited no effect on ORF57 ho-modimerization (lanes 2 and 3), neither for peptides vm3, vm7, nor vm8, but the latter three peptides induced the formation of larger ORF57 complexes, presumably due to their excessive
cross-linking to ORF57 (Fig. 7B, lanes 4, 8, and 9). Peptides vm4 and
vm9 also displayed no effect on ORF57 homodimerization but appeared to induce the instability of monomeric ORF57 (lanes 5 and 10). Interestingly, peptide vm5, derived from ORF57
C-ter-minal aa 280 to 299 (␣-helix 7), and peptide vm6, from aa 324 to
340 (an unstructural region between␣-helices 8 and 9), were
in-hibitory to ORF57 homodimerization (lanes 6 and 7). By
compar-ison with 3⫻FLAG peptide, serving as a negative control (Fig. 7C,
lanes 7 to 11, upper panel), the peptide vm5 inhibited ORF57
homodimerization in a dose-dependent manner (Fig. 7C, lanes 2
to 6, upper panel) with a decreased mobility of ORF57 monomers by several kilodaltons, indicating cross-linking of the peptide vm5 (2.33 kDa) to ORF57. As expected, similar results were observed with the peptide vm6, but not with the peptide vm2 or vm4, which
does not affect ORF57 homodimerization (Fig. 7C, three lower
panels). The vm5-corresponding regions (␣-helix 7) in ORF57
homologues, including HSV-1 ICP27, HCMV UL69, and EBV EB2, are a highly conserved region in structure and amino acid
sequence composition (Fig. 7DandE). All homologues contain a
20-aa-long␣-helix (helix 6 in ICP27 or EB2 and ␣-helix 9 in
UL69) (Fig. 7D) and several fully conserved residues (Fig. 7E). The
vm6 corresponding region among the homologues also has simi-lar features (see Fig. S2 in the supplemental material). Taken to-gether, our data indicate that ORF57 homodimerization requires the vm5 and vm6 corresponding regions.
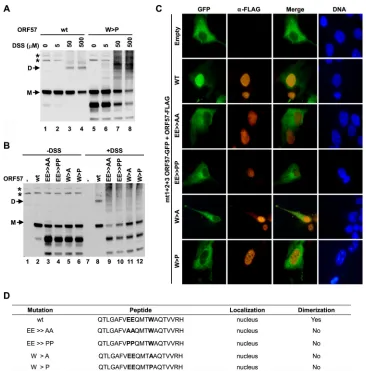
Disruption of ORF57 homodimerization via mutation of the
peptide vm5 region.To further investigate the role of the peptide
vm5 corresponding region in ORF57 dimerization, the conserved
residues E287, E288, and W292 within␣-helix 7 were mutated in
the context of full-length ORF57. The selected amino acids were mutated to either alanine, to eliminate the amino acid side chain,
or to helix-breaking proline (EE⬎⬎AA, EE⬎⬎PP, W⬎A, W⬎
P). Resulting mutants were then tested for their ability to ho-modimerize after expression in HEK293 cells by DSS-mediated
cross-linking. As shown inFig. 8A, wt ORF57 homodimers were
detectable in the samples with DSS (lanes 1 to 4), but the W⬎P
mutant failed to efficiently homodimerize at all three tested doses
of DSS (lanes 5 to 8). We noticed that, although the W⬎P mutant
was unstable, lack of dimerization of the mutant was not a result of its instability and appeared even when the expression level of the mutant was justified to that of wt ORF57 (compare lanes 6 to 8 with lanes 2 to 4 for the monomeric ORF57 level). Similarly, none of the other mutants was able to form homodimers when tested in
the same assay in the presence of DSS (Fig. 8B), with a justified
protein level close to that of wt ORF57. It is worth noting that the mutant proteins lacking homodimerization might appear as a high-molecular-weight smear in HEK293 cells in the presence of
high-dose DSS (Fig. 8AandB), indicating nonspecific
cross-link-ing to other unknown cellular proteins. The formation of ORF57 homodimer was further verified by nuclear translocation assays
using the cytoplasmic mt1⫹2⫹3 ORF57-GFP construct as a
re-porter. Compared with wt ORF57-FLAG, the mutants with all types of mutations in the peptide vm5 corresponding region
dis-played no ability to translocate the cytoplasmic mt1⫹2⫹3
ORF57-GFP into the nucleus, indicating dysfunctional
dimeriza-tion of the mutants with the reporter (Fig. 8C). Their properties
are summarized inFig. 8D. In conclusion, the peptide vm5
corre-sponding region (aa 280 to 299) at␣-helix 7 of the ORF57 protein
is critical for ORF57 homodimerization mediated by the amino acid residues highly conserved among ORF57 homologues.
Inhibition of ORF57 homodimerization induces
degrada-tion of ORF57.We noticed that introduction of point mutations
into the peptide vm5 corresponding region led to a significant
reduction of ORF57 protein expression in HeLa cells (Fig. 9A,
compare lanes 2 to 5 to lane 1), but a dimerable K345A mutant
on November 7, 2019 by guest
http://jvi.asm.org/
which contained a point mutation outside the peptide vm6
corre-sponding region did not show such a reduction (Fig. 9A, compare
lane 8 to lane 6). Compared to wt ORF57 RNA (Fig. 9A, middle
panel) and its protein level (Fig. 9A, top panel), all ORF57 mutants
showed a relative RNA level of⬃65% and a protein level of⬃5%
of that for wt ORF57 (Fig. 9A, bar graphs). Obviously, the
signif-icantly reduced level of a mutant protein was not correlated to a minimally reduced RNA level and thus could not be caused at the transcriptional or posttranscriptional level.
To examine whether an ORF57 mutant had a short half-life, we
compared wt ORF57 with the W⬎A mutant for protein stability
in HeLa cells treated with CHX in a time course manner. CHX is an inhibitor of protein biosynthesis in eukaryotes as it blocks
pro-tein translational elongation (55). Western blot analysis revealed
that a large amount (81%) of wt ORF57 protein remained
detect-able even at 4 h after blocking translation by CHX (Fig. 9B, lanes 1
to 4). In contrast, the W⬎A mutant decreased over time, with
only 16% of the protein left at 4 h after CHX blocking (Fig. 9B,
lanes 5 to 8). From our calculation using a line plot analysis, we
determined that the half-life (t1/2) of wt ORF57 is more than 4 h
(the last time point for the sample collection in the study), and the
mutant half-life is only about⬃1 h (Fig. 9B, line graph).
Interest-ingly, we found that, although the W⬎A mutant was less stable,
it resisted caspase cleavage (Fig. 9B, lanes 5 and 6), suggesting a
correlation between ORF57 homodimerization and its caspase di-gestion.
As the majority of intracellular proteins in eukaryotic cells are
degraded by cellular proteasomes (56), the effects of the
protea-some inhibitor MG132 on the expression of wt ORF57 and its
dimerization-defective mutants were tested in HeLa cells (Fig.
9C). Western blotting of protein samples showed an⬃50%
in-crease in the protein levels of EE⬎⬎AA, EE⬎⬎PP, and W⬎A
mutants and an⬃150% increase in the protein level of the W⬎P
mutant in the presence of MG132 over that in the absence of
MG132 (Fig. 9C, lanes 3 to 10 of top panel and the line graph in the
lower panel), whereas wt ORF57 protein remained constant,
ex-hibiting no visible changes under either condition (Fig. 9C, lanes 1
and 2, top panel), suggesting that the dimerization-defective
mu-FIG 7Inhibition of ORF57 homodimerization using small peptides. (A) Diagram of the predicted secondary structure of ORF57 protein along with positions of synthetic peptides vm1 to vm9 (blue lines). SeeTable 2for peptide details. (B) Examination of individual peptides that affect ORF57 homodimerization. Recombinant ORF57-FLAG protein was first incubated with each peptide in a protein:peptide molar ratio of 1:3 (5 nM ORF57:15 nM peptide) for 30 min, followed by cross-linking with DSS (500M) and analysis by Western blotting with an anti-ORF57 N-terminal antibody. M, monomer; D, dimer. (C) Dose-dependent effect of peptide vm5 on ORF57 homodimerization. Western blotting of ORF57-FLAG protein after incubation with increasing concentrations (5, 10, 15, 20, and 25 nM) of peptide vm5 or with control 3⫻FLAG peptide was conducted with an anti-ORF57 N-terminal antibody. ORF57 without cross-linking with any peptide or in the presence of increasing concentrations (5, 10, and 15 nM) of peptide vm2, vm4, or vm6 was used as a control. (D and E) Conservation of the secondary structure (D) and amino acid sequence (E) of the peptide vm5-corresponding region in ORF57 homologues. PSIPRED prediction (D) and ClustalW2 alignment (E) were used to analyze KSHV ORF57, HSV-1 ICP27, HCMV UL69, and EBV EB2 proteins. Consensus symbols (asterisk, semicolon, and period) were used according to ClustalW2 methods. In the red boxes are conserved amino acids E287, E288, and W292, mutated for further analysis.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:13.585.113.473.64.386.2]tants are prone to degradation via a cellular proteasome pathway. Given that proteasomic degradation is generally initiated by polyubiquitination of a target protein, the polyubiquitinated pro-tein often exhibits an increase in propro-tein molecular weight. How-ever, the polyubiquitinated protein may not be detectable under physiological condition due to its rapid degradation. To examine this possible polyubiquitination in the dimerization-defective
mutants, the W⬎P mutant was compared with wt ORF57 protein
in HeLa cells treated with MG132 for 6 h. As expected, the mutant protein was heavily degraded with multiple degraded protein frag-ments easily detectable by Western blotting with an anti-ORF57 N
terminus antibody (Fig. 9D, compare lanes 2 and 6 versus lanes 1
and 5 for input protein). Western blotting of wt ORF57 and the
W⬎P mutant in the IP pulldown indicated that only the W⬎P
mutant presented the higher-molecular-mass protein forms
(polyubiquinated protein forms) recognizable by an anti-ORF57
N terminus antibody (Fig. 9D, compare lanes 8 versus 7) and by a
K48-specific anti-polyubiquitin rabbit monoclonal antibody (Fig.
9D, lanes 9 and 10). The data demonstrate polyubiquitination of
the dimerization-defective W⬎P mutant.
DISCUSSION
KSHV ORF57 belongs to a family of sequentially conserved poorly but functionally well-conserved homologues found in all
herpes-viruses (15). To date, for none of the family members has a 3D
structure been solved by nuclear magnetic resonance (NMR) or X-ray crystallography, due to their poor expression in bacteria and precipitation during purification (unpublished data). In this study, we present the first comprehensive structural analysis of KSHV ORF57 protein using multiple computational prediction
FIG 8Introduction of point mutations into the peptide vm5-corresponding region in ORF57 prevents ORF57 homodimerization. (A) Homodimerization capability of wt ORF57 and its W⬎P mutant in HEK293 cells at increasing concentrations of DSS. HEK293 cells at 5⫻105cells in a 6-well plate were transfected
with 0.2g (for wt) or 2g (for mutant) ORF57-FLAG vectors. The cells at 24 h after transfection were treated with 20M MG132 for an additional 6 h and harvested in PBS forin vivocross-linking with DSS, and then cell lysates were detected by Western blotting with an anti-ORF57 N-terminal polyclonal antibody. *, a nonspecific protein band. (B) ORF57 protein carrying point mutations of E287A and E288A (EE⬎AA), E287P and E288P (EE⬎PP), W292A (W⬎A), and W292P (W⬎P) are all incapable of homodimerizing in HEK293 cells at 24 h after transfection in the presence of DSS (500M). Further deatils are provided in panel A. (C)In vivotranslocation assays in COS-1 cells cotransfected with mt1⫹2⫹3 ORF57-GFP together with a wild-type (wt) or mutant ORF57-FLAG fusion, described for panel B. Localization of ORF57-GFP (green) and ORF57-FLAG (red) proteins was examined by immunofluorescence microscopy at 24 h after transfection. The cell nuclei were counterstained with Hoechst 33258 dye (blue). (D) Summary of various mutations in the peptide vm5-corresponding region that affect ORF57 homodimerization.