0022-538X/93/031169-06$02.00/0
Copyright © 1993, AmericanSocietyfor
Microbiology
Integration Is
Essential for Efficient Gene Expression of
Human
Immunodeficiency Virus Type
1
HIROYUKI SAKAI,' MEIKOKAWAMURA,' JUN-ICHI SAKURAGI,12SAYURISAKURAGI,' RIRISHIBATA,1AKINORI ISHIMOTO,' NOBUMI ONO,2 SHIGEHARU UEDA,2
ANDAKIOADACHI1*
Institute forVirusResearch, Kyoto University, Kyoto 606,1andResearch Institute forMicrobial Diseases, Osaka University, Osaka 565,2 Japan
Received 7August 1992/Accepted 25 November 1992
A mutant of human immunodeficiency virustype 1 which carries a frameshift insertion in the integrase/ endonucleaseregion ofpolgenewasconstructed invitro. Upontransfection intocells, althoughthismutant exhibitedanormalphenotypewith respecttoexpressionofgag,pol,andenvgenesandtogenerationofprogeny
virions,noreplication-competent virus inCD4-positivecellsemerged.Anassayforthesingle-step replication ofa defective viralgenomedependent on trans complementation byrev proteinwasestablished and used to monitorthe early phase of viralinfection process. Viral cloneswith a mutation in the vif, vpr, or vpu gene
displayedno abnormality intheearly phase. In contrast, theintegrase mutantdid notdirecta markergene
expression afterinfection.Togetherwithanobservation that themutant lackedtheabilitytointegrate,these results indicated that theintegration wasrequiredforefficient viralgeneexpressionandproductiveinfection of humanimmunodeficiency virus type 1.
Thereplication cycleofretrovirusesconsists oftwomajor phases (4, 29). The first half of thecycleis directed toward theintegration ofacompleteDNAcopyof viralgenesinto hostcell DNA.Shortlyafterentryintosusceptible cells, the viral single-strandedRNAgenomeisconverted intoalinear
double-stranded DNA. The viral DNA intermediate then migratestothe cellnucleus and iscovalently integrated into
a host chromosome. The second phase begins with the expression of the genes encoded in the provirus and ends with theproductionofinfectiousprogenyviralparticles.The two events in the early phase of the replication cycle,
reverse transcription and integration, distinguish
retrovi-ruses fromother classes of virus. Thesetwo processesare
mediated by the two enzymes (reverse transcriptase and integrase) encodedbyavirussingleopen readingframe,pol
(5, 7).
IntegrationofaDNAcopyof theviralgenomeinto host
cellular DNA isan essentialstepinthe lifecycleofmost,if not all, retroviruses (4, 12, 29). Integration appears,
more-over,tobeimportantfor thetranscriptionof viral DNA into new copies of the viral genome and mRNAs that encode viral proteins (4, 12, 29). Integration-defective mutants of retroviruses are able to produce a DNA copy of the viral
genome,but thereplicationof themutantsinthetargetcells
is almostnegligible (6,14, 19, 20, 24).Therearesomereports
onthefunctional role of theintegraseof human
immunode-ficiencyvirus type 1 (HIV-1) inthe replication cycle. Viral clonescarryingamutation in theintegrase regionofpolgene wereunable to integrate (25)andwerereplicationdefective
(2, 3, 12, 25). Surprisingly, in one article (25), the uninte-grated HIV-1 DNA was reported to serve as an efficient template for HIV-1 antigen production. Because the
inte-grase of HIV-1 is very likely to function in the early
replication phase asdo those of the otherretroviruses, this result might represent a unique property of the HIV-1 integrase.
* Correspondingauthor.
In thiscommunication, an integration-deficient mutantof HIV-1 was constructed and analyzed in detail in several transfection and infection experiments. We demonstrate here thatthe HIV-1 integrase functions in the early replica-tion phase and that integration is important for viral gene expression.
MATERIALSANDMETHODS
Cell culture and DNAtransfection. Ahumancolon carci-noma cell line, SW480 (1), and a monkey kidney cell line, COS-1 (9), were maintained in Dulbecco's modified Eagle
medium supplemented with 10% heat-inactivated fetal calf
serum.CD4+humanT-cellleukemic celllines, A3.01 (8)and H9 (21), were maintained in RPMI 1640 medium
supple-mented withheat-inactivated fetal calfserum.For transfec-tion,uncleavedplasmidDNAwasintroduced intoadherent (SW480 andCOS-1) andnonadherent (A3.01 andH9) cells bythecalcium-phosphate coprecipitation (11, 30) and mod-ifiedDEAE-dextran (27)methods, respectively.
Infection. The infectivityof progenyvirions produced in the transfected SW480 cells was assayed in A3.01 cells. Culturesupernatantswerefiltered(poresize,0.45 ,um),and
appropriatevolumes were added to 106 cells aspreviously described(8).
RT assays. Virion-associated reverse transcriptase (RT) activity was measured as described previously (31). For quantitation, spots on DE81 paper were cut out, and RT activitywas determinedbyscintillation counting.
CAT assays. The chloramphenicol acetyltransferase (CAT)assayhas beenpreviouslydescribed(10).CAT levels were
assaypd
in equivalent amounts of cell lysates from transfected SW480cells and infected A3.01 cells.DNA constructs. An infectious proviral clone of HIV-1, pNL-432, and its mutants, designated pNL-Afl
(pol/inte-grasemutant), pNL-Nd(vif mutant), pNL-Af2 (vpr mutant),
andpNL-Ss (vpu mutant), have been previously described (1, 2, 17). pNL-nCATandthemutant clones ofpNL-nCAT
were constructed from pNL-432 (1) and itsmutant clones,
1169
on November 9, 2019 by guest
http://jvi.asm.org/
which have been previously described (2, 17). To make a series of pNL-nCAT constructs, the CATcoding sequence of pSV2CAT (784-bp HindIII-Sau3AI fragment) (23) was inserted into the XhoI (nucleotide [nt] 8887) and
KpnI
(nt 9005) sites in the nef coding region of parental clones, and therev coding sequencewas inactivated byinsertion of an 8-bp XbaI linker (Toyobo Inc., Osaka, Japan) into the BamHI site (nt 8465) (see Fig. 2). pNL-nCAT-Nh was generated from pNL-nCAT by digestion with the NheI restriction enzyme, by blunt ending with T4 DNA poly-merase,andbyresealingwith T4 DNAligase. Construction of pNL-gCAT and prevl has been describedpreviously(22, 23).EM.Transfected SW480 cells wereprocessed for electron microscopy (EM) by the standard method (2). Briefly, the cells were prefixed with Karnovsky's fixative (1.0% paraformaldehyde-3.0% glutaraldehydein 0.2 Mcacodylate buffer [pH 7.4]) at 4°C for 1 h, postfixed with 1% osmium tetroxide at 4°C for 1 h, and then dehydrated in a graded ethanol series and embedded in epoxy resin. Ultrathin sectionswereprepared,stainedwithuranylacetateandlead citrate solution, and observed under a Hitachi HU12A electronmicroscope.
Extractionof extrachromosomalandintegrated virus DNA. Extrachromosomal virus DNA in the infected cells was extracted at 10 h postinfection bythe method of Hirt (15). High-molecular-weightcellulargenomicDNAwasextracted from cells 2 days after infection by a standard proteinase K-phenol spooling method. RNAwas removed fromDNA solution by digestionwith RNase H and RNase T1 (Sigma Chemical Co., St.Louis, Mo.).
PCR analysis of viralDNA.DNAsampleswerepretreated withDpnI restriction endonuclease (New England Biolabs, Inc., Mass. USA) to remove plasmid DNAs potentially contaminated. Polymerase chain reaction (PCR) amplifica-tionwasperformedwithtwoprimers,V3DF(nt6990to7011 ofpNL-432) and SK69 (nt 7927 to 2907 ofpNL-432) (18). Samples(2,ugofDNA)weresubjectedto24cyclesof PCR ina50-,u reaction mixture. Compositionofreaction mixture was as the supplier suggested for Tth DNA polymerase (ToyoboInc.). Eachcycleconsisted of 1 min of denaturation (93°C), 2 min of annealing (57°C), and 3 min ofextension (72°C). Amplifiedproductswere runthrough a3%NuSieve GTG agarose gel (FMC BioProducts, Rockland, Md.) and analyzed by Southernblottinghybridization.The
32P-labeled
SalI (nt 5785)-XhoI (nt 8887) fragment produced by the random primerDNAlabeling kit (Takara Shuzo Co., Ltd., Kyoto,Japan)wasused asaprobe.Nucleotide sequence accession number. Nucleotide se-quence data of pNL-432 are from GenBank data base accession number M19921.
RESULTS
TransfectionanalysisofHIV-1integrase mutant.Aproviral DNAclone of HIV-1, designated pNL-Afl, which carriesa frameshift insertion within the integrase region of pol gene (2)was transfected into CD4-negative SW480 cells (1) and examinedforitsexpression.Wepreviously reported that the mutant expressed viral structural proteins and produced progeny virions normally in transfected SW480 cells, as judged byWestern immunoblotting analysis and RTassay, respectively(2). As shownin Fig. 1A, this clone could also direct the synthesis of progeny virions morphologically similartowild-type (wt) particlesupon transfection, andno
significant
difference between the amounts of progeniesAfA1 (IN)
4432
(WT
432(WT)
B
input 1 3 5 7 9 11 14 16mock
Af 1
(IN)
* [image:2.612.309.545.80.382.2]432(WT)
,*
0
FIG. 1. Characterization of an integrase mutant of HIV-1. SW480 cells were transfected with pNL-432 (HIV-1 wt) (1) or pNL-Afl (HIV-1 integrasemutant) (2), and48 hlater,sampleswere prepared forEM(A) and for infection (B). (A)EMpicture of virions produced by transfected SW480 cells. Afl(IN),virus from pNL-Afl;432(WT),virus frompNL-432.Bar = 100nm.(B) Infection of CD4-positivecells withvirus.Forinfection,A3.01 cells(106)were infected with 105 RT units of cell-free virus obtained from trans-fected SW480cells (input). RTproductionin the culture fluidsof infected cellswas monitored atintervals. Virus designationis the
sameaspresentedinpanelA.Values indicatedaysafter infection.
produced by wt and mutant clones was observed. The NL-Afl virus obtained from transfection, however, was unable to establish a productive infection in
CD4-positive
A3.01 cells. RT activity was not detected in the cells inoculatedwith the NL-Afl onanydayspostinfection (Fig. 1B). These resultswere consistent with those presented in previousreports,includingours(2, 3, 12,25), andsuggested that the viralintegraseprotein playsanimportant functional role in theearly phase of viral replication cycle, atleast in the SW480-A3.01 system. To examine theearlyreplication
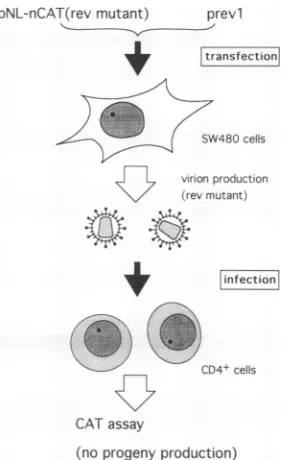
phasefurther, anewassay systemtoquantitateviral infec-tivity (from virus entry to transcription) similar to those previously described (13, 28) was established. This system utilizedreplication-defectiverev-minusproviralclones con-taining the CAT gene in the nef region (Fig. 2) and wasdependent on trans complementation for
production
of in-fectiousvectorvirus(Fig. 3).Whentherev wassupplied in trans, infectiousviruses,which carrythe CAT gene andare capable of undergoing one round of replication, would be generated (Fig. 3). Infection of cells with the virus and the CAT activity obtained would provide a measure of the efficiency withwhich the early infection phase of the viruson November 9, 2019 by guest
http://jvi.asm.org/
Stop codon
Irevf1F-n
Fv7lf
-t-at
-l nef|
pol
11
jIdl
Rjejnv
Aflii NdeIAflii SSpI Nhei
Frameshift
_ - rev - ]
vif
|1
-tat - |nefj \ pol Ivprl
ElI
env3D
O 50 100
% CONVERSION
I
I~~~~~~~~~~~~~~~~~~~~
rev
I+
pNL-432 (WildType)
0 0.5 1
[image:3.612.69.308.77.435.2]RELATIVE RT PRODUCTION
FIG. 2. Replication-defective proviral clones carrying a CAT
geneused forasingle-roundinfectionassay.(A)Genomestructures
of pNL-nCAT and pNL-gCAT. pNL-nCAT constructed in this
studycontainaCATgenein thenefgeneregionandanXbaI linker
insertion in the 2ndcodingexonof therevgene.Asaresultof the linkerinsertion, therevproteinwould stopatthe60th aminoacid,
and theenvprotein wouldhavea4-amino-acid insertionatthe site.
The mutant clones of pNL-nCAT, designated pNL-nCAT-Afl, pNL-nCAT-Nd, pNL-nCAT-Af2, pNL-nCAT-Ss, and pNL-nCAT-Nh, have frameshift mutations at the restriction enzyme sites
indicated. Generation and characterization of pNL-gCAT were
described previously (22). (B) rev dependency of marker gene
expression.pNL-nCATandpNL-gCAT (5 ,ug)weretransfected into
SW480 cells with(+)orwithout (-)revexpression plasmidprevl (10p.g).Twodays after transfection, CAT activityin the celllysates (upper graph)andRTproductioninto culture supernatantsrelative
tothatby pNL-432 (5jig)(lower graph)weredetermined. RTcounts produced bypNL-432were2,885cpm.
proceeds. As shown in Fig. 2B, while the pNL-nCAT construct (wt) forthis assay expressed CATactivity inde-pendently of the presence or absence of the rev protein withintransfected SW480 cells, itsprogenyproduction into aculture supernatantwas revdependent. Aseries of pNL-nCAT mutant constructs (Fig. 2A), which carry the same alteration in eitherthepol (integrase)orthe othergenes(vif,
vpr, vpu, andenv [gpl2O])aspreviously reported (2, 17) in
addition to the rev gene mutation, were transfected with
prevl (HIV-1 rev expression plasmid [22, 23]) into SW480 cells todetermine the effect of the mutationon the
expres-pNL-nCAT(rev mutant) prev 1
transfecton
SW480cells
virion production
trevmutant)
infection
CD4'cells
CATassay
(noprogenyproduction)
FIG. 3. A single-step infection assay. Virions recovered from SW480 cells cotransfected with replication-defective pNL-nCAT constructs(Fig. 2)andrevexpressionvectorprevl(22, 23)areable to undergo one round of replication but not multiple cycles of infection in CD4-positivecells. Earlyphaseof viral replication can be monitoredby the highlysensitive CAT assay.
sion of the marker CAT gene and the progenyproduction. Asis clear inFig. 4,noneof the mutations affected CAT and virusproduction, which is in agreement with our previous data(2, 17).To determine whether these results representa cell-specific phenomenon,several celllines, including CD4-positive A3.01 and H9, were transfected with wt or the integrase-minus mutantclone andmonitored for RT produc-tion. As shown inFig. 5,nosignificant difference, again, in the levelof progenyproductionin anyof thecell lines tested was observed. None of the progeny viruses produced in transfectedCOS-1, A3.01, and H9 cells were infectious (data notshown).
Ofparticular note is that the integrase mutant exhibited the wtphenotype in all transfection experiments described above. Because transfection ofaviral DNA clone into cells would bypass the early phase of viral replication (adsorp-tion, penetra(adsorp-tion, uncoating, and reverse transcription), these resultsarecertainly consistent with the requirement of the integrase proteinin theearlyinfection stage.
Analysisofearlyinfection phase ofthe mutant clones.The effect of mutations in the viral genes on the early stage of virusreplicationwasstudiedby the system described inFig. 3. Cell-free virussamples, collected onday 2 posttransfec-tion into SW480 cells, were inoculated into CD4-positive A3.01cells, and2 days later, celllysateswere prepared for CAT assay.Becausethegpl20envplaysacriticalrolefor the first step in the infection cycle, the env mutant carrying a frameshift insertion in the upstream region ofCD4-binding domain (Fig. 2A) was used as a negative control for this experiment. As shown in Fig. 6, approximately 100-fold increases in A3.01 cellCATactivitywereobserved between the positive (wt) and negative (env) control samples. In agreementwith thefindingthat the vpuproteinisrequired in the late infectionphase
(maturation/release)
(16), themuta-A pNL-nCAT
gag
pNL-gCAT
gag
B
revpNL-nCAT
pNL-gCAT
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.379.520.79.309.2]A
pNL-nCAT (wild type)
pNL-nCAT-Nh (env)
pNL-nCAT-Af1 (in t)
pNL-nCAT-Nd (vif)
pNL-nCAT-Af2 (vpr)
pNL-nCAT-Ss (vpu)
-~~~~---
~~~~~~~~~~~~~~~~~~~~~~~~~I
O 0.5 1.0
RELATIVE CAT ACTIVITY
I~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
I
~~~~~~~~~~~~~~~~~~~~~I
mock
pNLnCAT
pNLnCAT-Afl
mock
pNLnCAT
CD4-negative SW480
CD4-negativeCOS-1
pNLnCAT-Afl
B
pNL-nCAT (wild type)
pNL-nCAT-Nh (env)
pNL-nCAT-Af1 (in t)
pNL-nCAT-Nd (vif)
pNL-nCAT-Af2 (vpr)
pNL-nCAT-Ss (vpu)
0 0.5 1.0
mock
pNLnCAT
I1
CD4-positive A3.01
pNLnCAT-Afl
mock
pNLnCAT
CD4-positive H9
pNLnCAT-Afl
[image:4.612.55.299.71.354.2]RELATIVE RT PRODUCTION
FIG. 4. Expression of marker gene by transfectionofaseries of pNL-nCAT constructs. Theprovirus-CATclones(Fig.2)and prevl (revexpressionvector) were cotransfected into SW480cells,and 48 hlater, CAT activityin the cell lysates (A)and RTactivityin the culture supernatants (B) were determined. Relative activity is shown (that of wt pNL-nCAT = 1). CAT and RT activities
ex-pressed by the cellscotransfected with pNL-nCAT and prevl(black bars)were 24.8% (conversion ratio) and 3,124 cpm, respectively. Mutant designations and mutated genes (inparentheses) are indi-cated.
0 0.5 1.0
RELATIVE RT PRODUCTION
FIG. 5. Progenyproductionin variouscell lines transfected with theintegrasemutant.Four cell linesweretransfected withapair of pNL-nCAT(wt) plusprevl(rev supplier)orpNL-nCAT-Afl (inte-grase mutant) plusprevl, and 48 h later,RTactivityin the culture supernatantswasdetermined. Relative RT titer is shown. RTcounts
produced by various cell lines cotransfected withpNL-nCAT and prevl were 3,285 cpm (SW480), 1,898 cpm (COS-1), 1,825 cpm (A3.01), and 965 cpm(H9).
tion invpuhadnoeffectontheproductionofCAT. A similar
level of CAT activity was also observed in A3.01 cells infected with the viforvprmutantviruses. In contrast, no
significant increase in A3.01 CAT activity (relative to the activity of the negative control) was detected when the
integrasemutantwasusedas aninoculum. Theexperiments wererepeated three timeswithessentiallythesameresults.
Furthermore, the results were reproduced in two other T-cell lines(H9 andM8166)andamonocyticcellline, HL60
(data not shown).
To determine whether viral DNA is synthesized and integrated into achromosomal DNAof A3.01 cells infected with variousvirusclones,PCRanalysiswasperformed(Fig.
7). Hirt's DNA (15) and high-molecular-weight genomic DNAwereextracted from theinfected cellsatthe
appropri-atetime. Theenvsequence inthesampleswasamplified by thePCR procedure andprobed by a
3"P-labeled
viral DNAfragment. While a937-bptargetof the HIV-1envsequence could be amplified in both Hirt and genomic fractions of A3.01 cells infected with wt virus, the env sequence was
detected inneitherfraction of thecells infectedwith theenv mutantvirus,asexpected (Fig. 7B). Consistent with thedata inFig. 6,mutantviruses of thevif,vpr,andvpugenes were able to synthesize viral DNA and integrate into host cell DNA, Although HIV-1 DNA was produced normally in
A3.01 cells infected with the integrase mutant, no proviral
DNAintegrated into cellular DNA was found. This result demonstrated theintegration-defective propertyof our inte-grasemutant.
DISCUSSION
The results presented in this study demonstrate that unintegrated viralDNAis anextremely inefficienttemplate for HIV-1expressioninthe natural infection system and that integration mediated by viral integrase protein is essential for HIV-1 productive infection. An integration-defective (Fig. 7) integrase mutant constructed in vitro (Fig. 2B) displayedthe wtphenotypein alltransfection assays(Fig. 4 and5),as wasthecasefor theoriginal integrasemutant(Fig. 1)(2). Theintegrasemutantvirus obtainedbytrans comple-mentation (Fig. 3), however, was unable to express the markerCAT gene after infection ofCD4-positivetarget cells in a single-step replication system (Fig. 6). Although the conclusion described above represents a sharp contrast to that ofaprevious report(25), it is consistentwith the data obtained with the other retroviruses (6, 14, 19, 20). If integration is not required for efficient expression of viral genes,asclaimed in aprevious article (25),onemust assume that the integrase has some functional role in the late infection phase because all integrase
mnutants
described so farareunabletoestablishaproductiveinfection. Our data inon November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.332.522.75.374.2]pNL-nCAT (wild type)
pNL-nCAT-Nh (env)
pNL-nCAT-Afl (in t)
pNL-nCAT-Nd (vif)
pNL-nCAT-Af2 (vpr)
pNL-nCAT-Ss (vpu)
0 0.5 1.0
RELATIVE CAT ACTIN
FIG. 6. Analysis ofearly phase of viral replicati4 round infectionassay. Virussamples obtainedfrom(
ofpNL-nCAT constructs and prevl (Fig. 3) were CD4-positive A3.01 cells, and 48 h later, CAT acti lysates was monitored. Relative activity is shown detected in thecellsinfected with the wt NLnCAT v (conversion ratio). Mutant designations and mut; parentheses)are indicated.
Fig. 1, 5, and 6 do not support this hypothesis ularbasis forrequirementofintegration toeffici viralgenes remains tobe investigated.
Anotherimportant finding in this report istU
vif,
vpr, and vpu proteins appear not to have role in theearlyinfection phase (Fig. 6). As fo vprproteins, this result was well expected. The mutantwasreportedtobe defective in thever3
viral replication cycle (16). The growth kineti mutantviruses in CD4-positive target cells were delayed relative to those of wt virus (17), anr difficult todetect the mutational effect. Incon twoproteins,thevif
proteinwaspostulated to t theearly phaseofHIV-1infection in CD4-positA
Ftatv
tatJi
revE
SailI V3D-F
4-SK69
B
-- Z
HIRT GENON
FIG. 7. PCRanalysisofgenomic and HirtDNAs
A3.01 cells infected with various viruses. (A) L
primers V3D-F and SK69 within an env gene. L
restrictionsites (SallandXhoI) used to cut out a
32P-labeledprobe (usedforpanel B) is alsoindicatec
blot analysis ofthe amplified products by PCR. A2 infectedwithvirus asdescribed in the legendtoFi1
was extracted from infected cells for PCR ampli
appropriate time (see Materialsand Methods). The
were run through a 3% agarose gel and analyze( hybridization. Theresultsobtainedwith DNAsamp
chromosomalandgenomicfractionsareshown inpa
GENOMIC, respectively. Arrowheads indicate thes
amplified productspredictedfromthe location ofth
VITY
on by a single-cotransfections
The
vif
mutant virus obtained from transfected SW480 cells was normal with respect to infectivity as judged by a single-step infection assay (Fig. 6). This result showed that thevif
protein isnot required in the early infection stage. A more detailed studyis necessary toprecisely determine the functional role of the HIV-1vif
protein.ACKNOWLEDGMENTS
M.K. is an awardee of a Research Resident Fellowship from the Japanese Foundation for AIDS Prevention. This work was sup-ported in part by a grant-in-aid for AIDS research from the Ministry ofEducation, Science andCulture ofJapan.
REFERENCES
inoculated into 1. Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. Willey, A. Lvity in the cell Rabson, and M. A. Martin. 1986. Production of acquired immu-CAT activity nodeficiency syndrome-associated retrovirus in human and non-,iruswas28.2% human cells transfected with an infectious molecular clone. J. ated genes (in Virol. 59:284-291.
2. Adachi, A., N. Ono, H. Sakai, K. Ogawa, R. Shibata, T. Kiyomasu, M. Masuike, and S. Ueda. 1991. Generation and characterization of the human immunodeficiency virus type 1
s. The molec- mutants. Arch. Virol. 117:45-58.
iently
exmress
3. Clavel, F., M. D. Hoggan, R. L. Willey, K. Strebel, M. A.ently express Martin, and R. Repaske. 1989. Genetic recombination of human immunodeficiency virus. J. Virol. 63:1455-1459.
iatthe HIV-1 4. Coffin, J. M. 1990. Retroviridae and their replication, p. 1437-an important 1500. In B. N. Fields and D. M. Knipe (ed.), Virology, 2nd ed. ir the vpu and Raven Press, New York.
eproviral vpu 5. Craigie, R.,T.Fujiwara,and F. Bushman. 1990. The INprotein ylate stageof of Moloney murine leukemia virus processes the viral DNA ics of the vpr ends and accomplishes their integration in vitro. Cell
62:829-conlylightly 837.
only slightly 6. Donehower,
L.
H.,
and H. E.
Varmus.
1984. A
mutantmurine
it
wou be leukemia virus with a single missense codon inpolis defective Itrast to these in a function affecting integration. Proc. Natl. Acad. Sci. USA berequired in 81:6461-6465.Livecells (26). 7. Farnet, C. M., and W. A. Haseltine. 1990. Integration of human immunodeficiency virus type 1 DNA in vitro. Proc. Natl. Acad. Sci. USA87:4164-4168.
8. Folks, T., S. Benn, A. Rabson, T. Theodore, M. D. Hoggan, M. ij~I ne
IMartin,
M.Lightfoote,
and K. Sell. 1985.Characterization ofacontinuous T-cell line susceptible to the cytopathic effects of the acquired immune deficiency syndrome (AIDS)-associated retro-XhOI virus. Proc. Natl. Acad. Sci. USA 82:4539-4543.
9. Gluzman, Y. 1981. SV40-transformed simian cells support the replication of earlySV40 mutants. Cell 23:175-182.
10. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomeswhich express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051. Z< n 11. Graham, F. L., and A. J. Van der Eb. 1973. A new technique for
the assay of infectivity of human adenovirus5 DNA. Virology 52:456-467.
12. Haseltine,W.A. 1989. Development of antiviral drugs for the treatmentof AIDS: strategies and prospects. J. Acquired Im-mune Defic. Syndr. 2:311-334.
41C
13. Helseth, E., M. Kowalski, D.Gabuzda, U.Olshevsky,
W. Hasel-prepared from tine, and J. Sodroski. 1990. Rapid complementation assays ocation of the measuring replicative potential of human immunodeficiency ,ocation of the virus type 1 envelope glycoprotein mutants. J. Virol. 64:2416-fragment for a 2420.i.(B)Southern 14. Hippenmeyer, P. J., and D. P. Grandgenett. 1984. Requirement 3.01 cells were of the avian retrovirus pp32 DNA binding protein domain for g. 1, and DNA replication. Virology137:358-370.
fication at the 15. Hirt, B. 1967. Selective extraction of polyoma DNA from PCR products infected mouse cell cultures. J. Mol. Biol. 26:365-369. d by Southern 16. Klimkait, T., K. Strebel, M. D. Hoggan, M. A. Martin, and J. M. lesfrom extra- Orenstein. 1990. The human immunodeficiency virus type 1-spe-inelsHIRT and cific protein vpu is required for efficient virus maturation and size (937 bp) of release. J. Virol. 64:621-629.
eprimers. 17. Ogawa, K., R. Shibata, T. Kiyomasu,I.Higuchi, Y. Kishida, A.
M.
I
L-il.
z z x
5; S. .f. .f
z--.- -< .5z :;. ul
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.62.303.75.202.2] [image:5.612.62.298.459.598.2]Ishimoto, and A. Adachi. 1989. Mutational analysis of the human immunodeficiency virus vpr open reading frame. J. Virol. 63: 4110-4114.
18. Ou, C.-Y., S. Kwok, S. W. Mitchell, D. H. Mack, J. J. Sninsky, J. W. Krebs, P. Feorino, D.Warfield, and G. Schochetman. 1988. DNAamplificationfor direct detection of HIV-1 in DNA ofperipheralbloodmononuclearcells.Science239:295-297. 19. Panganiban, A. T., and H. M. Temin. 1983. The terminal
nucleotides ofretrovirusDNA arerequired for integration but notvirusproduction. Nature(London) 306:155-160.
20. Panganiban, A. T., and H. M. Temin.1984. The retrovirus pol geneencodesaproductrequired forDNAintegration: identifi-cation of a retrovirus int locus. Proc. Natl. Acad. Sci. USA 81:7885-7889.
21. Popovic, M., M. G. Sarngadharan, E. Read, and R. C. Gallo. 1984. Detection, isolation and continuous production of cyto-pathic retroviruses from patients with AIDS and pre-AIDS. Science 224:497-500.
22. Sakai, H., R. Shibata, J. Sakuragi, T. Kiyomasu, M. Kawamura, M.Hayami, A. Ishimoto, and A. Adachi.1991. Compatibility of rev gene activity in the four groups of primate lentiviruses. Virology 184:513-520.
23. Sakai, H., H. Siomi, H. Shida, R. Shibata, T. Kiyomasu, and A. Adachi. 1990. Functional comparison of transactivation by human retrovirus rev and rex genes. J. Virol.64:5833-5839. 24. Schwartzberg, P., J.Colicelli,andS. P. Goff.1984.Construction
and analysisofdeletionmutations in thepolgeneof Moloney murine leukemiavirus: a new viral function required for pro-ductive infection. Cell 37:1043-1052.
25. Stevenson, M., S. Haggerty, C. A. Lamonica, C. M. Meier,S.-K. Welch, and A.J. Wasiak. 1990.Integrationisnotnecessaryfor expression of human immunodeficiency virus type 1 protein products. J.Virol. 64:2421-2425.
26. Strebel, K., D. Daugherty, K. Clouse, D. Cohen, T. Folks, and M. A. Martin. 1987. The HIV "A" (sor) gene product is essentialfor virusinfectivity.Nature(London) 328:728-730. 27. Takai, H., and H. Ohmori. 1990. DNA transfection ofmouse
lymphoid cells bythe combinationof DEAE-dextran-mediated DNAuptakeandosmotic shockprocedure. Biochim. Biophys. Acta1048:105-109.
28. Terwilliger, E. F., B. Godin, J. G. Sodroski, and W. A. Hasel-tine. 1989. Construction and use of a replication-competent human immunodeficiency virus (HIV-1) that expresses the chloramphenicol acetyltransferase enzyme. Proc. Natl. Acad. Sci. USA 86:3857-3861.
29. Varmus, H. E., and P.0.Brown.1989.Retroviruses,p.53-108. In M.H. Howe and D. E. Berg(ed.),Mobile DNA. American SocietyforMicrobiology, Washington, D.C.
30. Wigler,M., A.Pellecer, S.Silverstein, R. Axel, G. Urlaub, and L.Chasin. 1979. DNA-mediated transferof the adenine phos-phoribosyltransferase locus into mammalian cells. Proc. Natl. Acad.Sci. USA76:1372-1376.
31. Willey, R. L., D. H. Smith, L. A. Lasky, T. S. Theodore, P. L. Earl, B. Moss, D. J.Capon, and M. A. Martin. 1988. Invitro mutagenesisidentifiesaregion within the envelopegeneofthe humanimmunodeficiency virus that is criticalforinfectivity.J. Virol. 62:139-147.