JOURNAL OFVIROLOGY,
JUlY
1989, p. 3080-3086 0022-538X/89/073080-07$02.00/0Copyright©) 1989, American SocietyforMicrobiology
Expression
and
Purification
of
Biologically Active
v-sis/Platelet-Derived Growth Factor
B Protein
by
Using
a
Baculovirus
Vector
System
NEILLGIESE,' MARY MAY-SIROFF,' WILLIAMJ. LAROCHELLE,' KATHLEEN VAN WYKE COELINGH,2
AND STUART A.
AARONSON'*
Laboratory of Cellularand MolecularBiology, Building 37, Room IE24, NationalCancerInstitute,' andLaboratory of
Infectious
Diseases,
NationalInstitutte
ofAllergy
andInfectiouis
Diseases,2
Bethesda,Maryland
20814Received 6February 1989/Accepted 16 March 1989
Malignant transformation induced by simian sarcomavirus is mediated by its v-sis protein, the monkey homolog of the platelet-derived growth factor (PDGF) B chain. By use of an appropriately engineered
baculovirusexpressionvector,the v-sisproteinwasexpressedinthe insect cell lineSpodopterafrugiperda (Sf9)
atalevel 50-to 100-foldhigherthan thatobserved withoverexpressioninmammalian-celltransfectants. The sisprotein producedbySf9 cells underwentprocessingsimilartothat observedinmammaliancells,including efficientdisulfide-linked dimer formation.Moreover,the recombinant sisproteinwascapableofbindingPDGF receptorsandinducingDNAsynthesis asefficientlyasPDGF-Bsynthesized bymammaliancells. Asignificant
fraction of sis proteinwasreleasedfrom Sf9cells,which madepossibleaone-stepimmunoaffinity purification
tonearhomogeneitywitha40% recoveryofbiologicalactivity.These results demonstrate thataproteinwhose
normal processing requires both intrachain and interchain disulfide-bridge formation can be efficiently
expressedinabiologically active form ininsectcellsby usingabaculovirusvector system. The v-sis oncogene of simian sarcoma virus encodes a
homolog of the human platelet-derived growth factor (PDGF)B chain (5, 34). PDGF is an important mitogen for
cells of mesenchymal origin. Recent evidence has linked PDGF to the etiology of certain human tumors, and it has also beenimplicated in the genesis of otherdiseases, suchas
arteriosclerosis (26). Thus,adetailedknowledge of its
struc-ture might aid in elucidating the interactions required for binding and activation of its cognate receptor. Molecular genetic approaches have been utilizedtoelucidatethe struc-ture of the v-sisIPDGF-B product. It is known that the minimal v-sis transforming domain spans 84 amino acids
identical insequencetohumanPDGF-B (11, 28). This region containseight cysteine residues, fourofwhich are involved
inintrachain disulfide linkages essential for biological activ-ity, whereas the othersappeartobe required for the forma-tion ofa disulfide-linked dimeric molecule (8, 27).
Thev-sis/PDGF-Bgene product expressedinmammalian cells remains tightly cell associated, despite its processing through the secretory pathway of the cell (25). Moreover, the level ofsis protein expression obtained, although suffi-cientforcertainanalyticalstudies, hasnotbeenadequate for detailedstructuralanalysis. Efforts toexpress the proteinin
procaryotic systems have led to high yields of inactive molecules (3). This is presumably because the reducing environment of bacteria prevents the properdisulfide-bond
formation required for the synthesis of biologically active dimers.
The baculovirus system has been successfully exploited forhigh-level expressionofavariety of mammalian proteins,
several ofwhich donotassumeanactive conformationwhen
expressed in bacteria(4, 6, 9). Usingthis system, we show
that v-sis/PDGF-B protein expression occurs at 50- to 100-fold-higher levels than achieved with strongeucaryotic pro-moters in mammalianfibroblasts. Wefurthershow that the
* Correspondingauthor.
processing and functional properties of the molecule in invertebrate and mammalian cells are similar. Finally, the
utility of the baculovirus system for purification of the processed PDGF-B product by immunoaffinity chromatog-raphy is demonstrated.
MATERIALS AND METHODS
Cell lines and virus. NIH 3T3 cells expressing the v-sis
gene under the control of the mouse metallothionein pro-moter(10), designated MMT-v-sis, weremaintainedat 37°C in Dulbecco minimum essential medium(DMEM)containing
10% calf serum. The fall armyworm ovary cell line, Spodoptera frugiperda (Sf9), wasobtained from the
Ameri-can Type Culture Collection, Rockville, Md., and main-tained at 27°C in complete Grace medium(GIBCO Labora-tories).
Wild-type baculovirus, Autographa calijornica nuclear polyhedrosisvirusE-2,waskindlyprovidedbyM. Summers and colleagues (TexasA & MUniversity)andwasamplified
by infection of Sf9 cells. Extracellular virions and viral genomic DNAwereprepared asdescribed previously (32).
Antibodies. Peptide antisera against the amino terminus (anti-sis N) or carboxy terminus (anti-sis C) of the v-sis
protein have been describedpreviously (8, 13, 24). Anti-sis V4serum wasraisedagainstasynthetic peptide, designated
V4, corresponding to amino acids 139 to 154 of the sis protein. V4 was conjugated to thyroglobulin and used to
immunize rabbits as described previously (24). Native
hu-manPDGF, isolated fromplatelets,wasusedto generatethe polyclonal antiserum, anti-PDGF, as described previously
(8, 13). Anti-PDGF immunoglobulin G (IgG) was prepared
byDEAE Affi-Gel Blue chromatography (Bio-Rad Labora-tories).
Construction of baculovirus expression vector pAc373sis and generation ofrecombinant baculovirus. Enzymes were
purchasedfromBoehringerMannheimBiochemicals,andall DNAmanipulationswereperformedbystandardprocedures 3080
Vol.63,No.7
on November 10, 2019 by guest
http://jvi.asm.org/
Polyhedrin Gene
pAc373
pAc373sis
Met Pro Asp Tyr Ser Bam HI
...TAATAAAAAAACCTATAAAT ATG CCG GAT TAT TCA...GGATCC
39 49 59 229
Bam HI ...TAATAAAAAAACCCGAGATCCGCGGATCC
39 49 62
Bam HI Met
...TAATAAAAAAACCCGAGATCCGCGGATCCGTCGACGGTATCGCTGCCTGGGTCC ATG
39 49 62 93
FIG. 1. Nucleotide sequences of the wild-type polyhedrin gene and transfer vectors surrounding the site of recombination and the initiation codons. The nucleotide sequences of the polyhedrin gene and of pAc373 are as described by Summers and Smith (32). The numbering system used is based upon the first transcribed nucleotide being designated 1.
(20). The pSSVsv2 plasmid (8) was digested with Sall and XbaI to generate a 1.2-kilobase-pair fragment containing the complete v-sis coding region as well as 22 base pairs of 5' untranslated sequence and 348 base pairs of 3' untranslated sequence. This fragment was ligated to BamHI linkers and subcloned into the BamnHI site of the transfer vector pAc373 (32). The sequences surrounding the vector insert junction, including the initiation codon, are shown in Fig. 1. This plasmid, designated pAc373sis, was grown in Escherichia coli DH5ox cells and purified by CsCl density gradient centrifugation (20).
Sf9 cells were cotransfected with 1 .g of baculovirus genomic DNA and 2 jig of pAc373sis plasmid DNA by the calcium phosphate transfection method (32). Three days after transfection, extracellular virions were collected and allowed to form plaques on Sf9 cellsaspreviously described (32). Polyhedrin-negative plaques were picked and used to
infect Sf9 cells in 96-well plates (32). Cells were lysed and screened by dot blot hybridization with a v-sis probe. Recombinant virus was further purified by performing two more rounds of plaque purification. Stocks of recombinant virus were grown on Sf9 cells to a titer of107to
10'
PFU/ml foruse in all subsequent experiments.Metabolic labeling, cell fractionation, and immunoprecipi-tation. Sf9 cells were seeded at a density of 107 cells per 100-mmpetri dish in complete Grace medium and infectedat a multiplicity of approximately10with recombinant baculo-virus. At 48 h postinfection, cells were labeled for4 h with
[35SJcysteine
(220 ,uCi/mI) in cysteine-free Grace medium. MMT-v-sis cells were seeded 16 h prior to labeling at 107cells per 100-mm dish in DMEM with 10% calfserum in 50
FiM
ZnCl,. Thecells werelabeled for4 h with [35S]cysteine(125 iLCi/ml) and
[35S]methionine
(125 jiCi/ml) incysteine-and methionine-free DMEM containing 25 jm
ZnCl,.
Bothisotopes were present at 1,200 Ci/mmol (DuPont Co.). Cul-ture supernatants collected after metabolic labeling were used to identify secreted forms of recombinant sis protein.
Membranefractions were prepared by hypotonically lysing
MMT-v-sisandSf9 cellsin 1 mMphosphate buffer (pH 7.4). Nuclei were removed bycentrifugation at 1,000 x g for 10 min. Supernatants were centrifuged at 100,000 x g for 90 min to precipitate membranes. The membrane pellet was suspended in 300
[LI
of10 mMphosphate buffer (pH 7.4) and boiled for 5 min to release sis protein. Membranes were repelleted at 100,000 x gfor 45 min, and the supernatant, designated P100,was used forimmunoprecipitations.Quan-titiative recovery ofsis proteinin the P100 fractionhas been previously demonstrated (25). Aliquots of P100 (50
[lI)
orculture supernatant (1 ml) were incubated with 3 pA of antiserafor 1 h at4°C. Immunoprecipitates were recovered with the aid ofStaphylococcus aurelus protein A bound to Sepharose beads (Pharmacia, Inc.). After solubilization by boiling in 4% sodium dodecyl sulfate (SDS) with or without 1.4 M 3-mercaptoethanol, proteins were analyzed by
SDS-polyacrylamide gelelectrophoresis (PAGE) (14%
polyacryl-amide).
Assay for mitogenic activity. Mitogenic assays were per-formed asdescribed previously (16). Briefly, NIH 3T3 cells were seeded at 2 x 104 cells per well into 96-well plates in DMEM-10% calf serum. Cells were allowed to become quiescent over 4 to 6 days at 37°C. Mitogens were added directly, and incubation at 37°C was continued foran addi-tional 16 h, atwhich time 2 jiCi of [3H]thymidine (50 to90
Ci/mmol; DuPont) was added, and the cells were kept at
37°C for an additional 5 h. Cell monolayers were washed with 5% trichloroacetic acid, solubilized in 0.25 N NaOH, and countedbyliquid scintillation.
Conditioned medium was collected from 100-mm petri
dishesover a 16-hperiodby incubatingthecells in 10ml of serum-free DMEM orGrace mediumfor MMT-v-sis orSf9 cells,respectively. Sf9 cellswereinfected withrecombinant baculovirus 48h priortocollection of conditioned medium. P100 wastested directly.
PDGF receptor competition assay. Binding experiments were performed on NIH 3T3 cells by a modification of the procedure described previously (22). Briefly, cells were seeded in DMEM-10% calfserum at8 x 105cells per wellin
12-well plates. At 48h, the cells were washed with binding
buffer (DMEM, 25 mM
N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic acid[HEPES], 1mgof bovineserumalbumin per ml [pH 7.4]) and incubated for 2 h at4°C with binding
buffer containing1 ngof
[125IJPDGF-B
andvarious concen-trations of unlabeled ligands. Cultures were washed exten-sively with binding buffer and solubilized with 20 mM HEPES (pH 7.0)-1% Triton X-100-10% glycerol.Radioac-tivity present in the lysates was quantitated by
using
agamma counter. PDGF-B (Amgen) was iodinated to 20,000
cpm/[jg
byusingthe method of Bolton and Hunter(1).Immunoaffinity purification of recombinant sis protein.
Recombinant sis protein was purified on
immunoaffinity
resins by couplinganti-PDGFIgGtoAffi-Gel 10asspecified
bythe manufacturer(Bio-Rad). Sf9 cells(108)wereinfected with recombinant baculovirus, and medium from the in-fected cells (100 ml) was concentrated 10-foldby using
aCentriprep 10 (Amicon Corp.). Concentrated medium was incubatedat4°Cfor16 hwith 1ml of
antibody-coupled
resinon November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.73.547.78.212.2]3082 GIESE ET AL.
inphosphate-buffered saline (pH 7.4) and 1% Triton X-100. The resin was washed extensively with phosphate-buffered saline, and the specifically bound protein was elutedwith0.2 M glycine hydrochloride (pH 2.2).
RESULTS
High-level expression ofv-sis/PDGF-B proteininSf9 cells. A baculovirus transfer vector, designated pAc373sis, was con-structed from pAc373 and a 1.2-kilobase-pair restriction fragment containing the v-sis gene. The nucleotide se-quences surrounding the site of recombination and the initiation codons of the wild-type polyhedrin gene and pAc373sis are shown in Fig. 1. The primary v-sis transla-tional products specified by pAc373sis or simian sarcoma virus would be identical, since the first methionine codon downstream of the polyhedrin promoter was the v-sis initi-ation codon. To transfer the v-sis gene into the baculovirus genome, the pAc373sis plasmid was cotransfected with wild-type A. californica nuclear polyhedrosis virus DNA into Sf9 cells and recombinant virus was isolated as previ-ously described (32).
To assess v-sis protein expression, we metabolically la-beled Sf9 cells at various times after infection with recom-binantvirus. Maximal levels of protein were observed at 48 hafter infection (data not shown), and all metabolic labeling studies whose results are presented here were performed during this time interval. Since the great majority of sis
protein expressed in mammalian cells is associated with the plasma membrane (25), we used the P100 membrane fraction
ofSf9 cells foranalysis ofcell-associated sis protein. Under reducing conditions, we observed a protein of
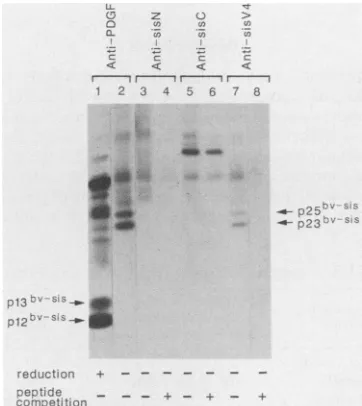
approximately 27,000daltons, p27bv-sis, which was
specif-ically detected by anti-PDGF serum as well as by antisera directed against the NH, terminus (anti-sis N), COOH terminus (anti-sis C), or an internalregion (anti-sis V4) of the v-sis precursor protein (Fig. 2). In addition to p27bv-sis, a protein of approximately 20,000 daltons, p20bv-sis, was observed as aless intense, diffuse band detected by anti-sis C and anti-PDGF but not anti-sis N. This pattern of immu-noreactivity suggested that limited amino-terminal process-ing had occurred. Similar results were observed when we
analyzed NIH 3T3 cells expressing the v-sis gene under the control of MMT-v-sis (Fig. 2). The primary translational product of v-sis, p28v-sis, is known to be glycosylated (25). The slightly altered mobility of p27bv-sis, as compared with p28v-sis, may be accounted for by differences in glycosyl-ation commonly observed in Sf9 cells when compared with mammalian cells (9, 12, 15, 30). All of these findings sug-gested that processing of the baculovirus-expressed v-sis protein was similar to that observed in mammalian cells (23). When expression levels were compared, the amount of v-sisproteindetectedimmunologically in recombinant virus-infected Sf9 cells was 50- to 100-fold larger than that present in zinc-induced MMT-v-sis cells (Fig. 2). The level of v-sis protein expressed in zinc-induced MMT-v-sis cells is the
highest we have observed in mammalian cell transfectants whenusing avariety of promoters, including simian virus40.
3-actin,
and Moloney mouse leukemia virus and simian sarcomavirus long terminal repeats (our unpublished obser-vations). Thus, the baculovirus system was considerably more efficient for expression of the v-sis product.Dimerization ofv-sis/PDGF-B protein inSf9 cells. In mam-malian cells, the primary v-sis translational product, p28v-sis,undergoes aseries of processing steps that include rapid
formation of a disulfide-linked homodimer, p56v-sis,
fol-MM?- v sls 4-!Sf9 -sisV
IT L t LL
E 0 >t 9m Z 0 > (9
.8 8 .8 8
< <: < < ~< < <
2 3 56 7* 9 10 1 12 13 14
; p270v SIS
_4--..* --2- bv sic
*...._ ,* _ _ _ _ + _ z + _
FIG. 2. Identification of the v-sis translation products in Sf9 cells. MMT-v-sis cells (lanes 1 to 7) orrecombinant virus-infected Sf9 cells (lanes 8 to 14) were metabolically labeled with [35S] methionine and/or[35S]cysteine for4h, and P100membrane frac-tions were prepared.Proteins were immunoprecipitated with anti-sis N, anti-sis C, anti-sis V4, or anti-PDGF as indicated above the lanes. Each peptide antiserum was preincubated with 15 p.g of homologous peptide asindicated below the lanes. Immunoprecipi-tates weretreatedwith 1.4 Mmercaptoethanol and analyzed on14% polyacrylamidegelscontaining SDS.
lowed by NH,-terminal and COOH-terminal cleavages to yield p35v-sis and p24v-sis, respectively (23). To evaluate disulfide-linked dimerization of v-sis protein in insect cells, we subjected immunoprecipitates to SDS-PAGE analysis undernonreducing conditions. p27bv-sis underwent efficient dimerization(Fig. 3). The major species detected by anti-sis C, anti-sis N, and anti-PDGF was approximately 54,000 daltons, p54bv-sis, consistentwith adimer of the full-length monomeric form. In addition to p54bv-sis, anti-sis C or anti-PDGF detected a dimeric form slightly larger than p35v-sis observed in MMT-v-sis cells (Fig. 3). The lack of detection of this form by anti-sis N demonstrated that it arosefrom amino-terminal processing. Anti-sis C, anti-sis N, and anti-PDGF each detected a small amount of a p27bv-sis
species, which probably represented the nascent monomer that has been detected in mammalian fibroblasts (8, 23).
Identification of v-sis/PDGF-B protein secreted from Sf9 cells. The secretion of recombinant proteins into culture fluids can provide an enriched source, greatly facilitating subsequent purification. The v-sis/PDGF-B protein is not efficiently secreted from mammalian cells (25). Nonetheless, the high level of its expression in Sf9 cells led us to investigateitsrelease into culturefluids. Thus, supernatants from metabolically labeled Sf9 cultures were subjected to immunoprecipitation and SDS-PAGE analysis. Under nonreducing conditions, anti-PDGF detected two dimeric proteins, of 23,000 (p23bv-sis) and 25,000 (p25bv-sis) daltons (Fig. 4). These proteinswere not detected by anti-sis N or anti-sis C, suggesting that proteolytic processing had oc-curred at both the amino and carboxy termini of the sis protein.
To confirm the identity ofthe secretedproteins, we used anantibody raisedagainstasynthetic peptide corresponding toamino acids 137to 152 of the sis protein, designated V4.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.325.561.78.281.2]DMMJT-v----sis
g-i-
Sf9-v -ssIz 2
2 0
1 2 3 A,
LL LL
C2 Z U
CD ,0 S CD
O .6 0
- c
<: <r < <:
5 6 7 S 9 10
muwI
,47'
4
4.
.)D6 -0. ,,*- -4!,V ,.
;..-n-,
c,35-. .4
-:eDtide
comnetitIon
+ _ + _-_ + - +
-FIG. 3. Dimerization of v-sis/PDGF-B protein expressed by recombinant virus-infected Sf9 cells. Recombinant virus-infected Sf9 cells (lanes 1 to 5) or MMT-v-sis cells (lanes 6 to 10) were metabolically labeled with[35S]methionineand/or[35S]cysteinefor 4 h, and P100 membrane fractions were prepared. Proteins were immunoprecipitated with anti-sis N (lanes 1,2,6, and 7), anti-sis C (lanes 3, 4,8,and9),oranti-PDGF (lanes 5 and 10) serum. In each case,peptideantisera werepreincubatedwith 15 p.gof homologous peptide as indicated below the lanes. Immunoprecipitates were analyzed on 14%polyacrylamide gels containing SDS.
Anti-sis V4 detected both p23bv-sis and p25bv-sis (Fig. 4). Moreover, the expected monomeric forms, pl2bv-sis and pl3bv-sis, wereobserved under reducing conditions (Fig. 4). These results demonstrated that a significant amount of v-sis/PDGF-B protein was secreted from recombinant virus-infected Sf9 cells.
Biological activity ofv-sis/PDGF-B protein produced in Sf cells. The mitogenic activity ofthe recombinant sis protein wasexamined by assaying its ability to induce[3H]thymidine incorporation in NIH 3T3 cells. Both the P100 membrane fraction and conditioned medium from recombinant virus-infected Sf9 cells induced a high level of thymidine incorpo-ration which was specifically blocked by anti-PDGF IgG (Fig. 5). Mitogenic activity was also detected in the P100
fraction from zinc-induced MMT-v-siscells, but the levelof
activity was 25- to 50-fold lower (Fig. 5A). These findings were consistent with the levels of immunoprecipitable sis protein associated with each cell type (Fig. 2). The condi-tioned medium from infected Sf9 cells was also found to contain a high level of mitogenic activity inhibitable by anti-PDGF IgG (Fig. 5B). The total amount of specific
mitogenic activity present in culture fluidsascompared with the P100fraction indicated thatapproximately 25 to50% of the sis protein produced was secreted. In contrast, MMT-v-sis cell culture fluids did not contain detectable mitogenic activity, consistent withthepreviousobservation that more than 95% of the sis protein produced in mammalian fibro-blasts remains cell associated (25).
Immunoaffinity purification of recombinant v-sis/PDGF-B protein.Todemonstrate theutility ofthisexpression system fortheisolation ofrecombinantsisprotein,wesubjectedthe secreted forms ofthe proteinto immunoaffinity purification
(see Materials and Methods). The concentration ofsis pro-teininconditioned medium from recombinant virus-infected
cells was estimated as0.26
pLg/ml
on the basisof itsmitoge-nic activity withpurified mammalian PDGF-B as astandard (Table 1). Immunoaffinity chromatography led to the recov-eryof around
40%
of thisactivity, all ofwhich was shownto be due to thesis protein onthebasisof itsneutralization by anti-PDGF serum (data not shown). Protein concentrationsbefore and afterimmunoaffinity enrichment indicated thata 150-fold purification ofsis protein had been achieved (Table 1).
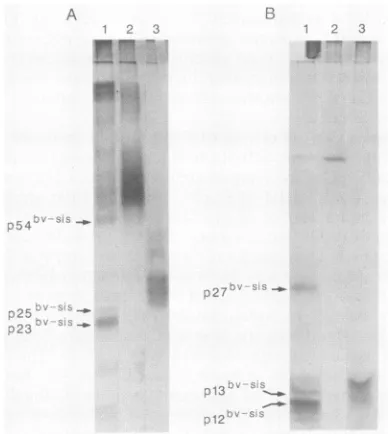
Toidentify the forms of the sis protein purified by immu-noaffinity chromatography, we analyzed the eluate fractions bySDS-PAGE and silver staining (Fig. 6). Under nonreduc-ing conditions, proteins migrating as p23bv-sis, p25bv-sis, and p54bv-sis dimers were readily observed (Fig. 6A).
Immunoblotting confirmed the identity of these forms as sis proteins (data not shown). Moreover, no proteins in this size range were detected under the same conditions from Sf9 cells infected with wild-type virus (Fig. 6). When analyzed under reducing conditions, the expected monomeric forms were observed (Fig. 6B). Analysis of purified PDGF-B produced by mammalian cells revealed a heterogeneous
population of dimer forms ranging in size from 26,000 to 32,000daltons undernonreducing conditions (Fig. 6A), and the expected monomeric forms of 12,000 to 15,000 daltons wereobserved underreducing conditions (Fig. 6B).
Amounts ofimmunoaffinity-purified recombinant sis pro-tein and PDGF-B isolated from mammalian cells were ad-justed such that eachcontained identical mitogenic activity (Fig. 6). Since similar amounts of protein were detected, these results indicatedthat the specific mitogenicactivity of the sis protein produced by Sf9 insect cells was nearly identical to that of PDGF-B protein expressed by mamma-lian cells. As a final test of the functional properties of
C5 z L >
1 2 3 4 5 6 8
ii.
- 2--p25--p2 3;"' ;"
p13z
red ctionr + -
-Peotide _ _ _
com;petition +-_ + +
FIG. 4. Immunoprecipitation of secreted forms ofv-sis protein produced by recombinant virus-infected Sf9 cells. Cells were in-fected withrecombinant virus 36 h priortometaboliclabeling with
[35S]cysteine
for 8 h.Conditioned media wereimmunoprecipitatedwithanti-PDGF(lanes1and2),anti-sis N(lanes3and4), anti-sis C (lanes5and
6).
oranti-sis V4(lanes7and8). Peptideantiserawerepreincubated withtheirhomologous peptideasindicated below the lanes. The sampleanalyzed in lane1 was reduced priorto electro-phoresisasindicated. Proteinswereanalyzed byelectrophoresis in
14% polyacrylamide gels containing SDS.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.77.282.77.280.2] [image:4.612.347.528.433.636.2]3084 GIESE ET AL.
2.0
E
et
0
0a 1.5
o
1-c 0
-am
0 1.0
0 U C
S C
E
.= 0.5
I-C.,
0.0
0 20 40 60 80 100 120 0 2 4 6
[image:5.612.143.480.74.312.2]MEMBRANE PROTEINS(10 cells) SECRETEDPROTEINS(10 cells)
FIG. 5. Mitogenic activity of v-sis/PDGF-B proteins produced bySf9orNIH 3T3 cells. Theincorporationof[VH]thymidineintoquiescent NIH3T3 cellswasdetermined after theaddition of v-sis proteins produced byrecombinant virus-infected Sf9cells(O)orMMT-v-siscells
(A). The P100 membrane fraction (A)orconditioned medium(B) preparedfrom the number of cells indicatedwas testeddirectly. Insome
cases, proteinsfromSf9 cells(A)orMMT-v-sis cells(A)were preincubatedwith anti-PDGFIgG.
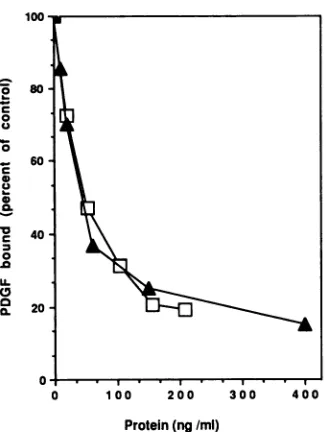
immunoaffinity-purifiedv-sisprotein,weexamined itsability
to compete with [125IlPDGF-B for PDGF receptor binding
on NIH 3T3 cells. When standardized on the basis of
mitogenic activity, sis protein produced by Sf9 cells
com-peted for PDGFreceptorbinding withanefficiency
indistin-guishable from PDGF-B isolatedfrom mammalian cells(Fig.
7).
DISCUSSION
The present study demonstrates the highly efficient expression of a biologically active growth factor in the baculovirussystem.Yieldsofthe sisproteinwereatleast 50-to 100-foldhigher than those observed in mouse fibroblasts
inwhich the transfected v-sisgene wasunder thecontrolof
a strong mammalian promoter. In addition, the level of recombinant sis protein produced by yeast expression (14)
wasatleast 10-fold lower than that observed in Sf9 cells. In
A B
1 2 3
I 4.;
,27` _
-'25
TABLE 1. Immunoaffinity purificationofv-sis/PDGF-B Protein-containing Amtof PDGF-B ig Purification Yield
substance protein"
equivalents"/mg
(fold) (%)(mg/mi) ofprotein
Conditionedmedium 0.13 2.0 1 100
Immunoaffinity 0.01 300 150 40'
column eluate
" Forproteinconcentrationsgreater than 0.05mg/ml.amountswere
deter-*minedbythe method of Lowry et al. (19). Lower protein concentrations were
determined by
A8(0
(7) or bysilver staining after SDS-PAGE. with knownquantitiesof mammalian PDGF-B as standards.
b Theamountofbiological activity equivalentto that observed for 1Figof
mammalian PDGF-B as determined by[3H]thymidineincorporationinto NIH
3T3 cells (seeMaterials and Methods).
' The averageyieldobserved in threeseparate experiments based on the
amountofbiological activityr-ecoveredin theimmunoaffinitycolumn eluate as
assayed by[3H]thymidineincorporationinto NIH 3T3 cells (see Materials and
Methods).
12
FIG. 6. SDS-PAGE of recombinant v-sis protein after
immu-noaffinity chromatography. Conditioned medium containing se-cretedproteinsfromSf9 cells infected with recombinantvirus(lanes 1) orwild-type virus (lanes 2) was concentrated 10-fold and
sub-jected toimmunoaffinity chromatography(see Materials and
Meth-ods). Eluate fractions were analyzed under nonreducing (A) or
reducing (B) conditions by electrophoresis in 14% polyacrylamide gels containingSDS. Lanes 3 contained 1 Figofmammalian PDGF-B
(Amgen), whereasanequivalent amountofrecombinant v-sis pro-teinwasanalyzedonthe basis ofbiological activity. Proteinswere
visualized bysilverstaining (Bio-Rad). E
asC
0
S._
E
U C 0
1CL
I-C.,
J. VIROL.
.3
PR
"ion November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.338.532.392.609.2]
80-soX
0 U
0040
0
U.
020-0 1oo 200 300 400
Protein(ng /ml)
FIG. 7. PDGF receptor-competing activity of recombinant
v-.si.slPDGF-B
protein. Immunoaffinity enriched v-siXs proteinpro-ducedbySf19 cells(A)wascomparedwith mammalian PDGF-B(O)
for its ability to compete with [1251]PDGF-B for NIH 3T3 cell receptorbinding.Thev-si.s proteinconcentrationwasestimatedon the basis of its mitogenic activity compared with that of PDGF-B
(Amgen). Duplicate determinations were made ateach
concentra-tion, andvalues reportedvariedby lessthan 5%s.
insect cells the sis protein underwent a
complex
series ofprocessing
steps,including
disulfide-linked dimer formation and amino- as well ascarboxy-terminal
processing. Thisprocessing
was very similartothe knownprocessing
of thesi.s
protein
inmammalian cells(23).Finally,
therecombinant sisprotein
bound PDGF receptors and induced DNA syn-thesisasefficiently
asdid PDGF-Bpurified
from mammalian cells.The sis
product
is verypoorly
released into culture fluidsby
most mammalian cell types (25). We showed that whenoverexpressed
by
Sf9 insect cells, a muchhigher
fraction(approaching
50%) was detected in culture fluids. Whether this reflects differences in levels ofproduction
or in condi-tions of incubation or some inherent differences between mammalian and insect cells is not known. In any case, the fact that asignificant
level ofsisprotein
wasreleased from Sf'9 cells made itpossible,
in a one-stepimmunoaffinity
purification
scheme,toenrich theproteinby
150-fold, witharecoveryof
40%o
ofmitogenic activity.
Previous studies have demonstrated the efficient
produc-tion ofbiologically
activepolypeptide
hormones orgrowth
factorsby using
a baculovirusexpression
system. These include beta interferon(30), interleukin-2 (29), interleukin-3(21),
erythropoietin
(35), andgastrin-releasing peptide
(17).Each of these
peptides
isamonomerinitsnative conforma-tion. Thus, v-si.s/PDGF-B is the first growth factor to beexpressed
as a dimerby using
the baculovirus system.Molecular
genetic
analysis
of the sis geneproduct
has led to evidence that fourcysteine
residues are required for intrachainlinkages
essentialfor thebiological activity
of the molecule (8, 27). In contrast, four othercysteine
residues each appear necessary for formation of a disulfide-linked dimer but are notrequired
for thebiologic
activity of the molecule (8).Understanding
of the interaction of the v-sis-encodedgrowth
factor with itsreceptor will be facilitatedby the identification ofspecificdisulfide linkages critical for sis structural and functionalproperties. The ability to over-express and purify mutantsisproteins by using the baculo-virus system should greatly facilitate structural characteri-zation. Moreover, the present study suggests that this system may aid in the expression and characterization of otherimportant dimeric polypeptide growthfactors or hor-mones such as colony-stimulating factor 1 (2), the beta transforming growth factors (31), and the inhibins (18, 33), whose structures and functions are also determined by multiple disulfide linkages.
ACKNOWLEDGMENTS
We thank Max Summers andcolleagues for plasmid pAc373 and wild-type A. (californica nuclear polyhedrosis virus. We are also grateful to KeithRobbins for helpful discussions during thecourse
of thiswork.
LITERATURE CITED
1. Bolton,A. E.,andW.M. Hunter. 1973. The labelingofproteins to high specific radioactivities by conjugation to a 1251-con-tainingacylating agent. Biochem. J. 133:529-539.
2. Das, S. K., and E. R. Stanley. 1982.Structure-function studiesof acolony-stimulating factor(CSF-1). J. Biol. Chem. 257:13679-13684.
3. Devare, S.G.,A.Shatzman, K. C.Robbins, M. Rosenberg,and
S. A. Aaronson. 1984. Expression of the PDGF-related
trans-forming protein of simian sarcoma virus in E. coli. Cell 36: 43-49.
4. Domongo, D. L., andI.S.Towbridge. 1988.Characterization of thehumantransferrin receptor produced inabaculovirus vector
system. J. Biol. Chem. 263:13386-13392.
5. Doolittle, R. F., M. W. Hunkapiller, L. E. Hood, S. G. Devare, K. C. Robbins, S. A. Aaronson, and H. N. Antoniades. 1983. Simiansarcoma virus oncgene, v-sis, is derivedfrom the gene (or genes) encoding a platelet-derived growth factor. Science 221:275-277.
6. Ellis, L., A. Levitan, M. H. Cobb,and P. Ramos. 1988. Efficient expression in insect cells of a soluble, active human insulin receptorprotein-tyrosine kinase domainbyuseofabacoluvirus vector. J. Virol. 62:634-639.
7. Fujikawa, K., and E. W. Davie. 1981. Human factor XII (Hagleman factor). Methods Enzymol. 80C:198-211.
8. Giese, N. A., K. C. Robbins,and S. A. Aaronson. 1987. The role ofindividual cysteine residues in the structure and function of the v-sis gene product. Science 236:1315-1318.
9. Greenfield, C., G. Patel, S. Clark, N. Jones, and M. D.
Water-field. 1988. Expressionof the human EGF receptor with
ligand-stimulatable kinase activity in insect cells using a baculovirus vector. EMBO J. 7:139-146.
10. Hamer, D. H., and M. Walling. 1982. Replication in viva ofa
cloned mammalian gene: cadmium induces thetranscriptionofa
mouse metallothion geneinSV40vectors.J. Mol. Appl. Genet. 1:273-288.
11. Hannink, M., M. K. Sauer,and D.J.Donoghue. 1986. Deletions intheC-terminalcoding regionof thev-sisgene: dimerization is required for transformation. Mol. Cell. Biol. 6:1304-1314. 12. Hsich, P., and P. W. Robbins. 1984. Regulation of
asparagine-linked oligosaccharide processing. Oligosaccharide processing in Aedes albopictus mosquito cells. J. Biol. Chem. 259:2375-2382.
13. Igarashi, H., C. D.Rao, M. Siroff,F. Leal, K. C.Robbins, and
S. A. Aaronson. 1987. Detection of PDGF-2 homodimers in
humantumor cells. Oncogene 1:79-85.
14. Kelly, J. D.,E. W.Raines, R.Ross,and M.J.Murray. 1985. The B chain of PDGF alone is sufficient for mitogenesis. EMBO J. 4:3399-3405.
15. Kuroda, K., C. Hauser, R. Rott, H. D. Klenk, and N. Doefler. 1986. Expression of the influenza virus haemagglutinin in insect cells by abaculovirus vector. EMBO J. 5:1359-1365.
16. Leal, F., L. T. Williams, K. C. Robbins, and S. A. Aaronson.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.94.257.73.289.2]3086 GIESE ET AL.
1985. Evidence that the v-sis gene product transforms by interaction with the receptor for platelet-derived growth factor. Science 230:327-330.
17. Lebacq-Verheyden, A.-M., P. G. Kasporzyk, M.G. Raum, K. Van Wyke Coelingh, J. Lebacq, and J. F. Battey. 1988. Post-translational processing of endogenous and of baculovirus-expressed humangastrin-releasing peptideprecursor. Mol. Cell. Biol. 8:3129-3155.
18. Ling,N.,S.Y. Ying, N. Ueno, J.Shamasaki, F. Esch, M.Hotta, and R. Guillemen. 1986. Pituitary FSH is released by a het-erodimer of the 3-subunits from the two forms of inhibition. Nature (London) 321:779-780.
19. Lowry,0.H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193:265-275.
20. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning:a laboratory manual. Cold Spring HarborLaboratory. Cold SpringHarbor, N.Y.
21. Miyajima, H., J.Schreurs,K.Otsu,A. Kondo, K. Avai, andS. Malda. 1987. Use of the silkworm Bombyx mori, an insect baculovirus vector for high-level expression and secretion of biologicallyactive mouse interleukin-3. Gene 58:273-281. 22. Nister, M., C.-H. Heldon, A. Wasteson, and B. Westermark.
1984. Aglioma-derived analog to platelet-derived growth factor: demonstration of receptor competing activity and immunologi-cal crossreactivity. Proc. NatI. Acad. Sci. USA 81:926-930. 23. Robbins, K. C., H. M. Antoniades, S. G. Devare, M. W.
Hunkapiller, and S. A. Aaronson. 1983. Structural and immuno-logical similarities between simian sarcoma virus gene prod-uct(s) and human platelet derived growth factor. Nature (London) 305:605-608.
24. Robbins,K. C.,S.G. Devare, E. P. Reddy, and S. A. Aaronson.
1982. In 'i4voidentification of the transforming gene product of simiansarcomavirus. Science 218:1131-1133.
25. Robbins,K. C., F. Leal, J. H. Pierce, and S. A. Aaronson. 1985.
The v-sis/PDGF-2 transforming gene product localizes to cell membranes but is not a secretory protein. EMBO J.
4:1783-1792.
26. Ross, R., E. W. Raines, and D. F. Bowen-Pope. 1986. The biology of platelet-derived growth factor. Cell 46:155-169. 27. Sauer, M. K., and D. J. Donoghue. 1988. Identification of
nonessential disulfide bonds and altered conformations in the v-sis protein, a homolog of the B chain of platelet-derived growthfactor. Mol. Cell. Biol. 8:1011-1018.
28. Sauer, M. K., M. Hannink, and D. J. Donoghue. 1986. Deletions in the N-terminalcoding region of the v-sis gene: determination of the minimal transforming region. J. Virol. 59:292-300. 29. Smith, G. E., G. Ju, B. L. Ericson, J. Moschera, H. Lahm, R.
Chizzonite, and M. D. Summers. 1985. Modification and secre-tion of human interleukin 2 produced in insect cells by a baculovirus expression vector. Proc. Natl. Acad. Sci. USA 82:8404-8408.
30. Smith, G. E., M. D. Summers, and M. J. Fraser. 1983. Produc-tion of human beta interferon in insect cells infected with a baculovirusexpression vector. Mol. Cell. Biol. 3:2156-2165. 31. Sporn, M. B., A. B. Roberts, L. M. Wakefield, and R. D.
Assovan. 1986. Transforminggrowthfactor-B: biological func-tion and chemical structure. Science 233:532-534.
32. Summers, M. D., and G. E. Smith. 1987. A manualof methods for baculovirus vectors and insect cell culture procedures. Bulletin no. 1555. Texas Agriculture Experimental Station, College Station,Tex.
33. Vale, W., J. Rivier, J. Vaughn, R. McClintock,A.Corrigan, W. Woo, D. J. Karr, and J. Speiss. 1986. Purification and charac-terization of an FSH releasing protein from porcine ovarian follicular fluid. Nature(London)321:776-779.
34. Waterfield, M. D., G. T. Scrace, N. Whittle, P. Stroobant, A.
Johnsson, A. Wasteson, B. Westermark, C.-H. Heldin, J. S. Huang, and T. F. Deuel. 1983. Platelet-derived growth factor is structurallyrelated to theputative transforming proteinp28s" of simian sarcoma virus. Nature(London)304:35-39.
35. Wojchowski, D. M., S. H. Orkin, and A. J. Sytkowski. 1987. Active human erythropoietin expressed in insect cells using a baculovirus vector: a role for N-linked oligosaccharide. Bio-chim. Biophys.Acta910:224-232.
J. VIROL.

![FIG. 2.cells.tionstatesSf9methionineN,lanes.homologouspolyacrylamide Identification of the v-sis translation products in Sf9 MMT-v-sis cells (lanes 1 to 7) or recombinant virus-infected cells (lanes 8 to 14) were metabolically labeled with[35S] and/or [35S](https://thumb-us.123doks.com/thumbv2/123dok_us/1325335.86349/3.612.325.561.78.281/tionstatessf-methioninen-homologouspolyacrylamide-identification-translation-recombinant-infected-metabolically.webp)