0022-538X/91/094629-07$02.00/0
CopyrightC) 1991,American SocietyforMicrobiology
The
Minute Virus
of
Mice
Capsid
Specifically
Recognizes
the 3'
Hairpin
Structure of
the
Viral
Replicative-Form
DNA:
Mapping
of the Binding Site by
Hydroxyl
Radical Footprinting
KURTWILLWANDAND BERNHARD HIRT*
Swiss InstituteforExperimental Cancer Research, CH-1066 Epalinges, Switzerland Received 29January1991/Accepted24May 1991
Theterminal hairpin structures of the DNA of minute virus of mice(MVM) are essential for viral replication. Here weshow that the hairpin 3' terminus ofMVM replicative-form DNA bindsspecifically toemptyMVM capsids. Binding of the same terminalDNA sequence in itslineardouble-stranded(extended)conformationwas notobserved. After heat denaturation and quick cooling of 3'-terminal extended-form fragments,notonly the virionstrand but also thecomplementary strand was found to bind to the capsid, presumably becauseeach strandre-formedasimilar hairpinstructure.Nobinding affinityfor thecapsidwasfoundtobeassociatedwith hairpin or extended 5' termini or with any other region of the viral DNA. Hydroxyl radical footprinting analyses revealed three protected nucleotide stretches forminga bindingsite at thebranch point of thetwo 3'-terminal hairpin arms looping out from the DNA stem (T structure). Single basechangeswithin this site did notaffect the binding.Inband shiftexperiments, specific bindingto theT structure wasdemonstrated forVP1 butnotforVP2.
The minute virus ofmice (MVM) belongs to the genus Parvovirus of the family Parvoviridae (20). These are non-envelopedicosahedral viruses which replicate autonomously in mammalian cells. The MVM genome is linear single-stranded DNA, 5,149 bases in length (2). At both termini, short self-complementary sequences are folded into stable hairpin structures (3, 18). The 3'-terminal hairpin serves as template andprimer forcomplementary-strand synthesis in vivo to produce a double-stranded replicative-form (RF) DNA (4).The ends of the RF DNA molecules occur either in thehairpin conformationor in the so-called extended form, in which the terminal DNA stretch is base paired to a complementarystrand forming a regular, linear terminus (3). Replication models which account for these conformational differences have been developed (6, 25).
The RFDNAsof the autonomous parvoviruses Aleutian mink disease virus (ADV) and bovine parvovirus have been shown to interact with viral capsid proteins via their 3' termini. The binding was observed only for 3' ends in the hairpin conformation but not for extended forms (15, 29). The ADV DNAhad been shown to interact with only one of thetwo ADVcapsid proteins, VP1 (28, 29). Here we show that the 3' terminus of the MVM RF DNA in its hairpin conformationalso binds to empty viral capsids and purified VP1. Using the hydroxyl radical footprinting technique, we havemapped the binding site to the central branch point of the T-shaped hairpin. We have tested other sequences forming similar T-shaped structures. These included the left-hand end of the MVM complementary strand, a 3'-terminal canine parvovirus (CPV) hairpin DNA fragment, and anoligodeoxynucleotide whichdiffered fromthe MVM genome inpositions 69 and 70. All theseDNAs werefound tobind to the MVMcapsidaswell. In contrast, we could not detectbindingof thecapsidtotheconformationally different 5'-terminal MVM DNA hairpin. Linear double-stranded DNAfragments, whetheroriginatingfromthe 3' end,the5'
*Correspondingauthor.
end, orotherregions of the MVM RF DNA, also were not foundtobind.
MATERIALSANDMETHODS
Materials. Restriction enzymes and DNA-modifying en-zymes werepurchased from Boehringer.Radiolabeled prod-ucts were from Du Pont. Products for radical footprinting werepurchased from Sigma.
Cellsand viruses. The cultivation ofmouse A9 fibroblast cells and thepropagation of MVMp, the prototype strain of MVM,wereperformedasdescribed elsewhere (1). CPVwas grown in feline NLFK cells.
Purification of MVMp particles. MVMp was purified on CsCl gradients according to the method described in refer-ence 28. Fractions were collected from the bottom of the tube, assayed for their density, dialyzed against 10 mM HEPES
(N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid) (pH8.0)-50mMNaClandanalyzed by sodiumdodecyl sulfate(SDS)-polyacrylamide
gel electrophoresis (PAGE) and Coomassie blue staining. The fraction containing the maximalamountofviralproteins, usually correspondingtoa densityof 1.35g/cm3,
wasconsideredtocontainpure empty viralparticles and was used in all experiments. The protein concentration was determined by fluorescencespectropho-tometry.
Purification ofviral proteins. Viral proteins were
purified
in SDS-PAGE gels by using 25 mM Tris-190 mM
glycine-0.1%SDSasrunning buffer. Theywerelocalized
by
staining
aparalleltrack withCoomassie blue andwereelectroeluted, usingthesamebuffer,in thepresence of 300
p.g
ofRNaseas carrier. After 3 h ofelectroelution at 25 mA, 1-mlsamples
were dialyzed against 1 mM HEPES (pH 8.0)-5 mM NaCl and lyophilized to a final volume of 100 ,ul. Proteins were
precipitatedwith 5volumes of acetone,dried,anddissolved in binding buffer(10mM HEPES [pH
8.0],
50 mMNaCl).
IsolationofMVM and CPV RFDNAs. At 24 h
postinfec-tion, whole-cell DNA was isolated by the method of Hirt 4629
on November 10, 2019 by guest
http://jvi.asm.org/
Ti
jsty
I
5'
CD
Le) ) GOD
CD 10 014
v-Ina0O
CM4
_ ~ CM CM
IUL t (7
f)'.. 0)
0I)CI) NtI aC I
le t-oo
Iqr
0 0 ~ 0 0.*1 .. 0 I
3'
VIUVI VI5'
S v~~~~- Ix olM-m_ v-o I4 b s|, IV ...9@
CD
Cl*.
I,'-".~~~~~~~~~~3'
5'.
1-1000 2000 3000 4000
Ava
II
Xba
I
r0
_-i-It-MboII
][1 5
,
Mbo
I
HI
5000 nucleotides
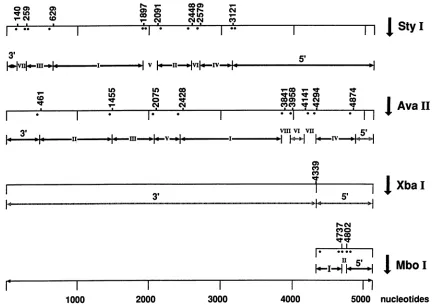
FIG. 1. Restriction sitesontheMVMpgenome.Thepositionsofthe 3' end labelsareillustratedby points.Thefragmentswhich havenot
beenlabeledareindicatedby hatchedarrows.
(11). Viral monomer RF DNAs were purified on 5to 30%
sucrosegradients (22).
DNAdigestion andendlabeling of fragments. DNA(1 ,ug)
was digested in the appropriate restriction buffer, phenol
extracted, ethanol precipitated, and dried. 3'-End labeling
was performed with Escherichia coli DNA polymerase I
(largeKlenowfragment) and a-32P-labeleddeoxynucleoside triphosphates. Specific fragmentswerepurifiedin
polyacryl-amidegels andrecovered by electroelution.
Gel retardation assay. Band shift experiments were
per-formedundertheconditions described previously (29). Sam-pleswereanalyzed byelectrophoresison5%polyacrylamide
gels ranginginlength from 10to35 cm.
Hydroxyl radical footprinting. The protocol of Zorbas et al. (30)was followedexceptfor differences in the composi-tion ofbinding buffer, the finalreaction volume, and the time ofreaction. The DNA-proteincomplexwas formed in 85 ,ul
of10 mM HEPES (pH 8.0)-50 mM NaCl as described for
band shift experiments. After 1 h of incubation at room
temperature,the solutionwasmade 1 mM in sodium
ascor-bate,20 p.M inEDTA, 10 p.M in(NH4)2Fe(SO4), and 0.3% in H202. Thefinalreaction volume was 100 ,u1. After addition
oftheH202 solution, the reactionwascarriedoutfor 2min andstopped with 10 ,ul of100 mM thiourea-20mM EDTA. When the DNAprotein complex wasto be separated from unbound DNA, 10% glycerol and 0.1% bromophenol blue
were added and the sampleswere immediatelyloaded onto
5% polyacrylamide gels. After electrophoresis, the DNA complex and free DNA were recovered by electroelution.
Both samples were phenol extracted, ethanol precipitated,
dried, and analyzed in 6% polyacrylamide-8 M urea
se-quencing gels.
Sequencingof DNA.Sequencingreactionswerecarriedout by themethod of Maxam and Gilbert(14).
Synthesisofoligodeoxynucleotides. Thesequencebetween nucleotides 39 and 79 of the MVM genome, including the part of the 3'-terminal hairpin which forms the T-shaped structure, was synthesized as a single 41-mer. After
self-annealing, the DNA was 3' end labeled by using Klenow fragment and [32P]dCTP.
RESULTS
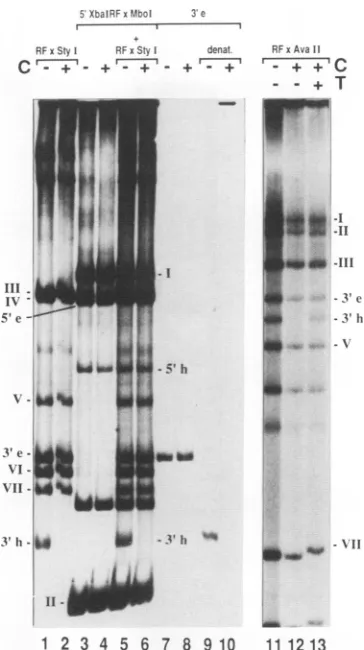
3'-Terminal hairpin of MVM RF DNA binds to MYM capsids. We carriedoutband shift experiments with MVM RF DNA enzymatically digested with StyI. The fragments
were3' end labeled by using radioactive dATP,asshown in
Fig. 1.Afterelectrophoresisina5%polyacrylamide gel,the
pattern seen in Fig. 2, lane 1, was obtained. When the digestion products were electrophoresed after incubation with MVM capsids, the band representing the 3'-terminal hairpin fragment disappeared (lane 2, band h), suggesting binding of this fragment to the capsid. In contrast, the mobilityof therespective extended-form fragment (140 bp, bande)remainedunchanged, indicatingthat thepresenceof theterminalsequence inahairpin conformationis essential for binding. The specificity of this binding is demonstrated by theunchangedmobilities of the otherdigestion products comingfrom differentregionsof the genome. However, in theexperiment described above, we could not analyze the
00) 0)
IVO
I . .
Nr 00) 0) 0) q U
0o 0. .
3'
lm-
1-c"J
*-+I.*-Iv
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.89.521.73.379.2]3' e
denat. RFxAva II
ri,7^_r
+
~~~
+ T-+
-+r
-I
0-IlwI~~~~~~-
L-l
~ ~
~~~~~~~
Ii.S'h~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
->3'h ' w
I'I
1 2 3 4 5 6 7 8 9 10 111213
FIG. 2. Gel mobility shift analysis of MVM RF DNA fragments. Restriction fragments were generated by digestion withStyI,XbaI, and MboI orAvaII. The 3'-end-labeled digestion products were separated in a 5% polyacrylamide gel (lanes 1 to6 and 11 to13). The gel-purified 3'-terminal extended-form fragment of the StyI digest was run in lanes 7 and 8, and its denatured form was run in lanes 9 and 10. For band shift analyses, samples wereincubated with0.05 ,ugof empty MVM capsids (C) (lanes 2, 4, 6, 8, 10, 12, and 13). The sample in lane 13 was additionally incubated with a100-fold molar excess of T oligodeoxynucleotide (T). h, hairpinform; e, extended form.
5'-terminal MVMStyI fragments because they were too long (2,000 bp). To produce short 5'-terminal fragments which could be tested in a band shift assay, the right-hand XbaI digestion product of MVM RF DNA was cleaved withMboI (Fig. 1). Figure 2, lane 3, shows the electrophoresis patterns of the resulting 3'-end-labeled fragments (one additional band with a length of about 110 bp is of unclear origin). The addition of MVM capsids had no influence on the position of the bands, including those representing hairpin or extended 5' termini (lane 4). To identify the latter, we have redigested the fragments with
AvaIl,
which cuts at nucleotide 4874 and therefore cleaves the 5'-terminal fragments labeled at nucle-otide 4802 and leads to the disappearance of the correspond-ing bands, which was in fact observed (data not shown). To confirm the result described above, both sets of MVM DNA fragments, i.e., those produced by StyI digestion and those resulting from cleavage with MboI, were loaded in the same lane (lane 5). After the addition of MVM capsids(lane 6), all bands remained unchanged except for that representing the 3'-terminal hairpin. AnAvaII
digest of MVM RF DNA wasloaded without priortreatment (lane 11) or afterincubation with empty capsids (lane 12). The specific disappearance of theband of the 3' hairpin fragment (about 400 bp, lane 12) was prevented when a 100-fold molar excess of a synthetic
oligodeoxynucleotide
that spanned the region betweennu-cleotides 39 and 79 on the MVM genome was added (lane 13). This oligodeoxynucleotide (T oligodeoxynucleotide) is expected to form the T-shaped structure characterized by
two
hairpin
armsloopingoutfromthestemofthe molecule(compare withFig. 7). FragmentIVoftheAvaIl digest isnot visible in Fig. 2(lanes 11, 12, and 13), although it should be labeled(Fig. 1). Onthe other hand, there are twoadditional fragments of unclear origin (Fig. 2, between fragment V and VII). If we suppose that the restriction site at nucleotide 4874 is not GGACC, as published (2), but GGTCC, the extended- andhairpin-form5'-terminalfragments could have been labeled with dATP and thus they, instead offragment IV, would have given the observed result.
Inlane7, thegel-purified extended 3'-terminal fragment is visible. As expected, there was no shift of this fragment produced by added MVM capsids (lane 8). However, on adding capsids to heat-denatured and quick-cooled samples, all thematerialwas movedtothe top of the gel (lanes 9 and 10). Since the fragment possesses a 3' recessed right-hand endwhich has been filled in by Klenow polymerase, the label is within the complementary strand. The unpaired comple-mentary strand can, like the virion strand, form a T-shaped hairpin structure (compare Fig. 7). This structure, which permits binding to the capsid, has never been observed in vivo. The same result as that seen in Fig. 2, lanes 9 and 10, was obtained with DNA labeled in thevirion strand with T4 polynucleotide kinase (datanot shown).
Hydroxyl radical footprinting. The 3'-terminal hairpin frag-ment ofthe Styldigest (compare Fig. 1 and 2) was used for radical footprinting experiments (Fig. 3). The cleavage pat-tern ofthe free DNA is shown in lane 1 and those of the DNA-protein complex are in lanes 2 and 3. Threeregions of DNA, clearly seenin lane 3, were foundto be protected by bound capsid. This experiment was repeated, and the posi-tions of protection (lane 9) were mapped relative to the migrationof sequencingreaction products (lanes 5 to 8). The material analyzedinlanes 3 and 9 had been recovered from a polyacrylamide gel in which theDNA-protein complex was separated from unbound DNA after the radical reaction. In lane 10, the unbound DNA samplerecovered fromthe same gel was loaded. This sample is specificallyenriched in DNA molecules which are nicked in the binding regions (see Discussion). Lane 11 shows free DNA thatwas cleaved in the absence of any protein.
The capsid binds at the branch point of the terminal T structure. The protected area includes 5 nucleotides in the centerof the T (noted as C in Fig. 3) as well as 6 nucleotides in both the 3'- and5'-oriented hairpin arms (depicted in Fig. 4). Additional nucleotides are semiprotected: 3'-CACT-5' between nucleotides 38 and 41, C in position 48, G in position 62 andTinposition 74. In the 5'-oriented armof the T, wefound a sequence slightly different from that published by Astell et al. (3): starting with nucleotide 68, thesequence observed was not 3'-CTG-5' but 3'-CGT-5'.
Binding of VP1 to the Tstructure. The empty MVMcapsid is formedby twodifferent proteins designated VP1 and VP2. We wanted to know whetherbothoronlyoneof them would bind to the viral DNA. The two capsid proteins were separated by SDS-PAGE and gel purified as described above. A gel retardation experiment was carried out using the MVM StyI digest. The result is given in Fig. 5. Lane 1
5XbalRFxf0bol
I I
RFxStyl RFxSty
C - '+- +' ,§
ll
IV'
5' e
t i L
UU0
i~~
N'.,
W;!d
it
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.87.269.72.397.2]A
Sty1-3-h O H
+ +
]5'
]C
797
IM. 0
1:.
] 3
B
StyI-3-h
C Py G Pu- -'
++ C
c
__*
-_ _ i~~
MP
sk
_r I
Z CL a- m C: > > >
3'h
1 2 3 4 5 6 7 8 9 10 11
FIG. 3. Hydroxyl radicalfootprintinganalysis oftheinteraction between the MVMcapsid and the MVMRF3'hairpin terminus. (A) Lane 1,hydroxyl radical-inducedcleavage of the 3'-terminal MVM RFStyIhairpinfragment;lanes2and3,cleavage patternsofDNA capsid complexes formed by1 ,ug(lane 2)or2,g(lane 3) of empty MVM capsids. Priorto theiranalysis, thecomplex andfree DNA had beenpurified in polyacrylamide gels. (B) Mappingofprotection sites. Lane 4, pBR322Hinflfragments (154 and75bp); lanes5 to8, Maxam andGilbertsequencing reactions of theStyIfragment; lane 9, protection by bound MVM capsids against hydroxyl radical-mediated cleavage; lane 10, DNA from the same reaction that migrated as free DNA during gel purification; lane 11, direct cleavage of free DNA in the absence of capsid.
50 AR G I
'C
G ,G
CG 40 30 20
GIC'C 3I G
GA3
I I *3TCPWACGTCACTTACGT
ACATG..
.CCAR
60 CG 1IIIIIIIIIIIIIII 11111
AGTGTGCAGTGARTGCAA
GTGTAC...
GGTTCI A
A 80 140
AG 90
FIG. 4. Schematic representation of the interaction between the MVMcapsid and the 3'-terminalhairpinMVM RFStyIfragment. Large black boxes represent areas of complete protection, cross-hatched boxes indicatesemiprotected regions, and asterisks indicate positions ofthe 3' end labels.
1 2 3 4 5
FIG. 5. Mobility shiftanalysisof MVMRFStyIfragments.The StyIdigestwas runin lane1. Lane 2shows thepattern of thesame digest in the presence of 15jigof RNase. MVMcapsid proteinsVP1 orVP2 wereadded in lanes3 and4,respectively.BothVP1andVP2 wereaddedin lane5.e, extendedform; h,hairpin.
shows the electrophoresis pattern of the digest in the ab-senceofprotein. Lane 2 isacontrol which confirms thatthe mobilities of the fragments remain unchanged in the pres-enceof 15 ,ug of RNase. RNasewasalso addedtotheviral protein samplesas acarrier. The80-bp3' hairpin fragment partially disappearedin the presence of VP1 (lane 3), sug-gesting thatit bound to theprotein. In contrast, no loss of material was visible for the extended terminal fragment, indicating the involvement of the 3' hairpin structure in bindingtoVP1. VP2 hadnosimilareffect when addedtothe DNA(lane 4),althoughit hadbeenappliedinapproximately a fivefold excess overVP1, corresponding to the stoichio-metric ratio of the viral proteins within the capsid (23). Interestingly, when VP1 and VP2 in the amountsapplied in lane 3 or4 were mixed together, incubated for 30 min in binding buffer,and thenadded to the DNA,even moreof the 3' hairpin fragment disappeared(lane 5).
Theexperimentwasrepeatedwith thesyntheticT oligode-oxynucleotide (usedascompetitorinFig. 2, lane 13). Itwas shiftedbyVP1,confirmingthat theviralprotein binds within the hairpinT structure. Noshift of the T oligodeoxynucle-otidewasproducedbyVP2(datanotshown).
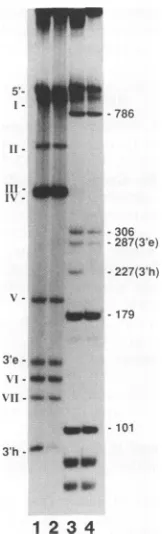
Sequence requirements for binding. Asshown above(Fig. 2),after heatdenaturation andquickcooling, bothstrandsof
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.413.476.66.428.2] [image:4.612.94.275.73.398.2] [image:4.612.68.298.568.678.2]5.-!F.7 86
i'
'179
3'e-l
N1-46_
VI a
-o
-1013h-F w__
GNP
1 234
FIG. 6. Gel mobility shift analysis ofrestriction fragments of MVMandCPVRF DNAs.Restrictionfragmentswereproduced by digestion ofMVM RF DNAwith StyI or of CPV RF DNA with AccI.The3'-end-labeled digestion products were separated in a5% polyacrylamide gel (lanes 1 and 3). For mobility shift analysis, sampleswereincubatedwith0.05 ,ugofempty MVMcapsids(lanes 2and4). e,extendedform; h,hairpin.
MVM RF DNA were found to bind to the viral capsid, indicating that theDNAsecondary structurerather thanits nucleotidesequenceis important forbinding. Furthermore, bindingwasdemonstrated foranoligodeoxynucleotide com-prisingasequence published elsewhere (3) which was sub-sequently found to be slightly different from the MVMp sequence we observed. To see whetherfurther changes in thenucleotide sequence would have any influence on bind-ing,weusedanAccIdigest of CPVRF DNAinaband shift experimentwith empty MVM capsids. As shownin Fig. 6, theMVMcapsid bindstothe3'-terminal hairpin fragment of AccI-digested CPVRF DNA(lane 4, band h) with the same specificityasit bindstoitshomologous 3'-terminalhairpinin StyI-digestedMVM RF DNA(lane 2).Lanes 1and 3contain therespective free-DNA samples.InFig.7, the sequencesof thefourDNAswhichhavebeenfoundto bindtothe MVM capsid are depicted. Each sequence can form a T-shaped secondary structure which is essential forthe formation of the DNA-protein complex. Differences from the MVM se-quence within the T
oligodeoxynucleotide
and the CPV DNAarenoted.DISCUSSION
Wehaveanalyzedthebinding ofempty MVMcapsids to the 3'terminus of MVM RF DNA. Thecapsid recognizesthe specific structure of the 3'-terminal hairpin and does not interactwith the 3'endinitsextended form. Thebindingis duetoone ofthe twocapsidproteins, VP1. Like the entire capsid,gel-purified VP1 wasfoundtodiscriminate between
A,CG
Gc' TCATCAC...3'
A
AGT GTG...
"cc AG
3 TERMINALTSTRUMRE OFMVMCp)DNA
A,Gc
CG/oGCG
Gc' TCATCAC.. .3'
C \ AGT GTG... A G
STRUCTMRE OFT-OLIGODEOXYNUCLEOTIDE
T C0
Co TCA CAC...3'
III III
GC\ AGTAGTG...
GC\suGC
cc
\'C\\GC
T C
T-STRUCTURE OF THECOMPLEMENTARYSTANDOFTET-TERMINUSOFMV (p)DNA
R G
G TCATCAC...3'
-~ III III
AGT GTG... A G
3'- TERMINALTSTRUMREOFCPUDNA
FIG. 7. Schematicrepresentation ofthe secondary structuresof fourDNAsbindingto the MVMcapsid.The left-handpartsofthe virionandcomplementarystrands of MVM RF DNA are shown at thetop. Thepositions ofsingle base changes between the sequence of the MVM virion strand and two other DNA species, the T oligodeoxynucleotide and the 3'-terminal sequence of the CPV virion strand,areshown.
the extended andhairpin forms ofthe 3' terminus, binding onlytothe latter. The aminoacidsequencesof VP1 and VP2 areidentical exceptforaregionattheamino terminus which isuniquefor VP1 (24). This suggests that theamino-terminal region is involved in binding to the hairpin. In band shift experiments (Fig. 5, lane 3), the VP1-DNAcomplexes did not enter the gel. Complexes with the T oligodeoxynucle-otide were visible as clear bandsjust at the topofthe gel (data not shown). This suggests that VP1 formed large aggregates. The existence ofparvovirus capsid protein ag-gregateshasbeen observedinvivo(17). Although nospecific DNA-bindingactivity wasfound forVP2alone, its addition toVP1markedlyenhanced thebindingtothe 3' hairpin (Fig. 5, lane 5). The incubation of the viral proteins before the addition ofDNAmayhave resulted in thereconstitution of a capsidlike structure with higher affinity for the hairpin region.
Thespecific binding ofthe MVMcapsidtothe MVM DNA 3' terminus isanalogous to previous findings obtained with ADV and bovine parvovirus (15, 29), suggesting that this interaction plays a general role during the recognition of parvovirus DNA by the parvovirus capsid. In many other systems, the specificrecognitionofaviralgenomeby viral capsid proteins serves to ensure the
encapsidation
of the viral genome and preventspackaging ofextraneous nucleic acid (13,21,27;forareview,seereference5).Thesynthesis
ofsingle-stranded
parvovirus
progeny DNAanditspackag-ing are simultaneous processes whichrequire the presence
of
preformed
emptyviralcapsids
(9, 16, 19). Thebinding
ofthe parvovirus RF DNA 3' end to the
parvovirus
capsid
mightthus serve for both the synthesis of progeny strands and their concomitant
encapsidation.
A model for these processes hasbeenproposed elsewhere(29).It hasbeenassumed thata
T-shaped
structure, character-izedbytwohairpinarmsloopingoutfrom the DNA stem, is formed by intramolecular basepairing
within the MVM DNA 3' terminus (3).Figure
4 showsa schematic represen-tation of this structure,including
thecapsid-binding site,
asmapped by radical
footprinting.
Thecapsid
binds toon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.142.223.74.341.2] [image:5.612.318.554.74.252.2]quences which are held close together within the branch
point region
ofthe Tandprotectseachof the threejunctions.This
finding
is in strong support of the T-type structurepredicted
by the sequence analysis. An extended,non-branched arrangement of the same binding sequences
changes
their positions in space and may thus preventinteraction with thecapsid.The branchpoint regionof the T structure is not in aB-type helix. The DNA will be bent at
the
junctions
like DNA within cruciform structures (7).These bends may represent signals which contribute to the
recognition
ofthecapsid
asit has been observed for othersystems
(reviewed
in reference 26).Hydroxyl
radicals attack the DNA within the minorgroove (10). Hence, the efficient protection of the DNA within the capsid-binding region suggests that the capsid binds totheminor groove. In contrast, we couldnotdetect
protection
of thebinding
region against dimethylsulfate-induced DNA cleavage: one single G at position 47 was foundtobe
protected (data
notshown).
SinceG residuesaremethylated
atthe N-7position
in themajor
groove(14), theeffect of
minor-groove-binding proteins
on Gmethylation is limited.Samples
treated withhydroxyl
radicals generallycon-tained both the
DNA-capsid
complex and unbound DNA. Afterthehydroxyl
radical reaction, thecomplexwas sepa-rated from free DNAby
PAGEbefore it wasanalyzed inasequencing gel.
When the free DNA was also recoveredfrom the polyacrylamide gel and analyzed in parallel, a
peculiar
patternwasobserved(Fig. 3,lane10).Itlookedlikea
negative footprint,
showingsignals
only in those regionswhich were
protected
in the complex (Fig. 3, lane 9). Toexplain
thispattern,
we assumethat there is anequilibrium
between theunbound-DNA
pool
and DNA inthecomplex.
Ifafree-DNA molecule which is nickedin the
binding region
has alower
affinity
forthecapsid,
thismoleculewill remainlonger
in the free-DNApool
than will molecules with anintact
binding region.
Its contribution to the free-DNAdigestion
patternwillresultinahigher signal intensity
withinregions
correspondingto the capsid-binding site. Thus, theinformation
implied
in the patterninlane 10isequivalentto data obtainedfromaninterferenceexperiment:
the strongest bandscorrespond
to those sequences which areindispens-ablefor
binding.
The MVM
capsid
was found to bind to four differentT-shaped species containing
different nucleotide sequences(Fig. 7), indicating
thatbindingcantolerate certain changesin the
primary
sequence.Thequestionsarise:is anyT-form-ing
sequence acceptable, or are there more constraints tobinding?
There are reasons to favor the first possibility. Parvovirusesareverywell conserved with regard totheir3'hairpin
structures but much less so with regard to theprimary
sequencewithinthesestructures (8). Furthermore,the
replication
of adeno-associated virus DNA clearlyde-pends
onthe presenceof the Tstructure withinthe invertedterminal repetition but not on the sequence within the
repetition
(12). Binding experiments with modifiedse-quences will
provide
information on the importance of theprimary
structure.ACKNOWLEDGMENTS
We thankP. Beard and J. Benhattarforsuggestions and discus-sions. We are grateful to H. Zorbas, Institute for Biochemistry,
UniversityofMunich, Munich, Germany, for invaluablehelp with the radical footprintingtechnique. G. Siegl kindly provided CPV. We thank B. Bentele for technical assistance and A. Emery-Goodman foroligodeoxynucleotide synthesis.
This workwassupported inpartbyagrantfromtheCommission oftheEuropeanCommunitytoK.Willwandandbyagrantfrom the Fonds National Suisse delaRechercheScientifiquetoP.Beard and B. Hirt.
REFERENCES
1. Antonietti,J.-P.,R. Sahli,P.Beard,and B. Hirt.1988. Charac-terization ofthecelltype-specific determinant in thegenomeof minute virus of mice.J. Virol. 62:552-557.
2. Astell, C. R., E. M. Gardiner, and P. Tattersall. 1986. DNA sequence of the lymphotropic variant of minute virus ofmice, MVM(i), andcomparison withtheDNA sequenceof the fibro-tropicprototype strain. J. Virol. 57:656-669.
3. Astell, C. R., M. Smith, M. B. Chow, and D. C. Ward. 1979. Structure of the 3'-hairpin termini of four rodent parvovirus genomes: nucleotide sequence homology at origins of DNA replication. Cell 17:691.
4. Bourguignon, G. J., P. J. Tattersall, and D. C. Ward. 1976. DNA of minute virus of mice: self-priming, nonpermuted, single-stranded genome with a 5'-terminal hairpin duplex. J. Virol. 20:290-306.
5. Casjens,S. 1985.Nucleicacidpackaging byviruses,p.75-147. In S. Casjens (ed.), Virus structureand assembly. Jones and BartlettPublishers, Inc.,Boston.
6. Cotmore, S. F., and P. Tattersall. 1987. The autonomously replicating parvoviruses of vertebrates. Adv. VirusRes. 33:91-173.
7. Gough, G. W., and D. M. Lilley.1985.DNAbendinginducedby cruciform formation. Nature (London)313:154-156.
8. Hauswirt, W. W. 1984. Autonomous parvovirusDNAstructure
and replication,p. 129-152. In K. I. Berns (ed.),The parvovi-ruses. Plenum Press, NewYork.
9. Hermonat, P. L., M. A. Labow, R. Wright, K. I.Berns, and N. Muzyczka. 1984. Genetics ofadeno-associated virus: isolation andpreliminary characterization ofadeno-associated virustype 2 mutants.J. Virol.51:329-339.
10. Hertzberg,R.P., and P. B. Dervan. 1984.Cleavage of DNA with methidiumpropyl-EDTA-iron(II): reactionconditions and prod-uct analyses. Biochemistry 23:3939-3945.
11. Hirt, B. 1967. Selective extraction of polyoma DNA from infected mouse cell cultures. J. Mol. Biol. 26:365-369. 12. Lefebvre, R. B., S. Riva, andK.I. Berns. 1984. Conformation
takes place over sequence in adeno-associated virus DNA replication. Mol. Cell. Biol. 4:1416-1419.
13. Lever, A., H. Gottlinger, W. Haseltine, and J. Sodroski. 1989. Identification ofa sequencerequiredfor efficient packagingof human immunodeficiency virus type 1 RNA virions. J. Virol. 63:4085-4087.
14. Maxam, A. M., and W. Gilbert. 1980. Sequencingend-labeled DNA with base-specific chemical cleavages. MethodsEnzymol. 65:499-560.
15. Metcalf, J. B., R. C. Bates, and M. Lederman. 1990.Interaction of virally coded protein and a cell cycle-regulated cellular protein with the bovine parvovirus left terminus ori. J. Virol. 64:5485-5490.
16. Muller, D.-E., and G. Siegl. 1983. Maturation of parvovirus Lulll in a subcellular system. I. Optimal conditions for in vitro synthesis and encapsidation of viral DNA. J. Gen. Virol. 64:1043-1054.
17. Paradiso, P. 1983. Analysis of the protein-protein interactionsin the parvovirus H-1 capsid. J. Virol. 46:94-102.
18. Rhode, S. L., and B. Klaassen. 1982. DNA sequence of the 5'-terminus containing the replication origin of parvovirus rep-licative form DNA. J. Virol 22:778-793.
19. Senapathy, P., J. D. Tratschin, and B. J. Carter. 1984. Replica-tion ofadeno-associated virus DNA: complementation of natu-rally occurring rep- mutants by wild-type genome or an ori-mutant andcorrection of terminal palindrome deletions. J. Mol. Biol. 179:1-20.
20. Siegl,G., R. C. Bates, K. I. Berns, B. J. Carter, D. C. Kelly, E. Kurstak, and P. Tattersall. 1985. Characteristics andtaxonomy ofparvoviridae. Intervirology 23:61-73.
21. Stow, N. D., E. C. McMonagle, and A. J. Davison. 1983.
on November 10, 2019 by guest
http://jvi.asm.org/
Fragments from bothtermini of the herpes simplex virustype 1
genome contain signals required for theencapsidation of viral DNA. Nucleic Acids Res. 11:8205-8220.
22. Strauss, S.E., E. D.Sebring,and J. A. Rose.1976.Concatemers of alternating plus and minus strands are intermediates in
adeno-associated virus DNA synthesis. Proc. Natl. Acad. Sci. USA 73:742-746.
23. Tattersall, P., P. J. Cawte, A. J. Shatkin, and D. C. Ward. 1976. Three structural polypeptides coded- for by minute virus of mice,aparvovirus. J. Virol.20:273-289.
24. Tattersall, P., A. J.Shatkin,andD. C. Ward. 1977. Sequence homology between thestructural polypeptides of minute virus of mice.J. Mol. Biol. 111:375-394.
25. Tattersall, P., and D. C. Ward. 1976. Rolling hairpin model for replication ofparvovirus and linearchromosomalDNA.Nature (London) 263:106-109.
26. Travers, A. A. 1989. DNA confirmation and protein binding.
Annu. Rev. Biochem. 58:427-452.
27. Vlazny, D. A., A. Kwong, and N. Frenkel. 1981. Site-specific cleavage/packagingof herpes simplex virus DNA and the selec-tive maturation of nucleocapsids containing full-length viral DNA. Proc. Natl. Acad. Sci. USA79:1423-1427.
28. Wiliwand, K., and O.-R. Kaaden. 1988. Capsid protein VP1 (p85) of Aleutian disease virus isamajor DNA-binding protein. Virology166:52-57.
29. Willwand, K., and O.-R. Kaaden. 1990. Proteins of viral and
cellularorigin bind tothe Aleutian disease virus(ADV) DNA 3'-terminal hairpin: presentation ofascheme for encapsidation of ADV DNA. J. Virol.'64:1598-1605.
30. Zorbas, H., L. Rogge, M. Meisterernst, and E.-L. Winnacker.
1989. Hydroxyl radical footprints reveal novel structural
fea-turesaround theNF1 binding site in adenovirus DNA. Nucleic Acids Res. 17:7735-7748.