0022-538X/85/120655-05$02.00/0
Copyright © 1985, American Societyfor Microbiology
Sequence Alterations in Temperature-Sensitive M-Protein
Mutants
(Complementation Group III) of Vesicular Stomatitis Virus
Y. GOPALAKRISHNA ANDJ. LENARD*
DepartmentofPhysiology and Biophysics, University of Medicine and Dentistry of New Jersey-Rutgers Medical School,
Piscataway, NewJersey 08854
Received 6 May 1985/Accepted 29 July 1985
Sequencesweredeterminedof thecoding regionsofthe M-proteingenesof theGlasgowandOrsaystrains
of vesicular stomatitis virus(Indiana serotype) and oftwogroupIII (M-protein) mutantsderived from each
wildtype.Synthetic primerswereannealedwith viral genomicRNAand extendedwithreversetranscriptase.
The resulting high-molecular-weight cDNA was sequenced directly. Both Glasgow and Orsay wild types
differedip 13 bases froma cloneof the SanJuan strain sequencedby J. K. Rose and C. J.Gallione(J. Virol.
39:519-528, 1981). Six of these base changes caused amino acid changesineach wild type,whereassevenwere
degenerate. The Orsay aild Glasgowsequencesresembled each othermorecloselythan either resembled that
of Rose andGallione, differingineightnucleotides and four amino acids. Each of the
four
mutants,however,differedfromitspargntwild type inonlyone ortwopointmutations. Everymutation causedachangeeither
fromortoachargedaminoacid;thechangefor tsG3lwasLys (position 215)toGlu,thechangeforts023was
Gly (position 21)'toGlu,thechangefor ts089wasAla(position 133)toAsp,thechangesfor tsG33wereLys (position 204)toThr and Glu(positioh 214) toLys. The chargedifferencespredicted fromtheseaminoacid
changeswas confirmedby nonequilibrium pH gradient electrophoresisfortsG31, tsG33, ts023, and thetwo
wildtypes. Thesemutations affect residuesspanning nearly85% ofthe linearsequence, althoughthemutants
possessnearly identical phenotypic properties.
The Mprotein of vesicular stomatitis virus (VSV) is oneof the threemajor proteinsfound inpurified virions, accounting for nearly one-third of the total protein in purified viral preparations (1). Mprotein appears to berequired for viral
budding
(21, 22, 25),implying that it must interact with both viral nucleocapsids and membranes. Interaction with the viral nucleocapsid is indicated by the M protein's ability toinhibit
viral transcription, both in detergent disrupted viral preparations(3, 6, 26)and ininfectedcells (5, 11).M-protein binding to nucleocapsid depends largely on electrostatic interactions, since inhibition can be reversed by theaddition of salt orpolyanions such aspolyglutamicacid(2, 3, 6, 26). The interaction ofMprotein withlipidmembranes of intact virions has been demonstrated by cross-linking withradio-labeled,
lipids (15) and by labeling with membrane-specific probes (10, 27). Thewild-type Mproteindoesnotappeartopenetratedeeplyinto
the
membrane, however, sinceit doesnot react with a probe localized close to the center ofthe viral bilayer(24).
The temperature-sensitive mutants of VSV provide an
important tool forthestudy of individual viralproteins.VSV has five transcribed genes, and its mutants fall into five complementation groups (16); group III corresponds to
mutations in the M protein (7, 8, 16). Earlier studies from this laboratory have characterized some properties of the four group IIImutantssequencedin the presentstudy (9, 10, 17, 26). Allfourmutants behaved in apractically identical fashion. Aweakening of electrostatic interactions between
mutant M protein and nucleocapsid (as compared with the wild type) was indicated by (i) a decrease in the M protein's ability toinhibit polymerase activity in detergent-disrupted virions (9, 26), (iii a decrease in spatial proximity between M protein and the nucleocapsid N protein in intact mutant virionsasindicated by cross-linking experiments (9, 10), and
*Correspondingauthor.
(iii)anincrease inRNAsynthesisincells infectedwithgroup III mutants at permissive temperature, as shown by others (5, 11). Most surprisingly, this decrease in M protein-nucleocapsid interaction was associated with a coordinate increase in the association ofmutantM protein with
mem-branes. Thiswasdemonstratedby (i)anincreaseinlabeling ofmutant as compared with
wild-type
Mprotein
in intact virions byamembrane soluble photoactivated probe(9, 10) and (ii) a decrease in G-protein mobility on the surface ofmutant as compared with wild-type-infected cells at the permissive temperature, measured by fluorescence photo-bleachingrecovery (9, 17).
The presentstudywasundertakentodetermine theamino acid changes that underlie the altered properties of the
mutant M proteins. We show that each mutant phenotype
canbeaccountedforbyasingle amino acid substitutionthat decreases the M protein's positive charge or increases its negative charge.Since the mutatedresiduesspannearly 85% ofthelinearsequence, it appears thatmost ofthe molecule is involved in its function.
MATERIALS ANDMETHODS
Cellsandviruses.TheOrsayandGlasgowvariantsofVSV Indiana and the group III mutants were obtained from A. Huang andA.Flamand.Virusesweregrownin BHK-21cells and isolated and purified as described previously (13). All stockswereplaque purified,andtheir temperaturesensitivity wasdetermined by plaque assay on Vero cells at 31 and 39°C.
Isolationofviral
genomic
RNA.Purifiedviruswaspelleted andsuspended (2to 3 mg/ml) by brief sonication in 10 mMTris-1 mM EDTA (pH 7.0). The virus was
deproteinized
withproteinase K (SigmaChemical Co.; 0.04 to 0.5 mglml) in1% sodium
dodecyl
sulfate and 0.25 M NaClat55°Cfor 10 min. The mixture was extracted twice with phenol-chloroform and once withchloroform,
and the aqueous phasewas precipitated twice with ethanol.655
on November 10, 2019 by guest
http://jvi.asm.org/
45 55 65 75 85 95
T T C A T C A T G A G T T C C T T A A A G A A G A T T C T C G G T C T G A A G G G G A A A G G T A A G A A A T C T A A G
MET SER SER LEU LYS LYS ILE LEU GLY LEU LYS GLY LYS GLY LYS LYS SER LYS
105 115 125 135 145 155
A A A T T A G G G A T C G C A C C A C C C C C T T A T G A A G A G G A C A C T A A C A T G G A G T A T G C T C C G A G C LYS LEU GLY ILE ALA PRO PRO PRO TYR GLU GLU ASP THR ASN MET GLU TYR ALA PRO SER
165
G C T C C A A T T G A C A A A T C C ALA PRO ILE ASP LYS SER
175
T A T TYR
225 235
T T A A G A T A T G A G A A A T T C T T C LEU ARG TYR GLU LYS PHE PHE
185 195 205 215
T T T G G A G T T G A C G A G A T G G A C A C T C A T G A T C C G A A T C A A PHE GLY VAL ASP GLU MET ASP THR HIS ASP PRO ASN GLN
245 255 265 275
T T T A C A G T G A A A A T G A C G G T T A G A T C T A A T C G T C C G T T C PHE THR VAL LYS MET THR VAL ARG SER ASN ARG PRO PHE
285 295 305 315 32-5 335
A G A A C A T A C T C A G A T G T G G C A G C C G C T G T A T C C C A T T G G G A T C A C A T G T A C A T C G G A A T G
ARG THR TYR SER ASP VAL ALA ALA ALA VAL SER HIS TRP ASP HIS MET TYR ILE GLY MET
345 G C A G G G A A A C G T
ALA GLY LYS ARG
355 C C C T T C T A C
PRO PHE TYR
365 375 385 395
A A G A T C T T G G C T T T T T T G G G T T C T T C T A A T C T A A A G G C C LYS ILE LEU ALA PHE LEU GLY SER SER ASN LEU LYS ALA 405
A C T C C A G C G G T A T T G THR PRO ALA VAL LEU
465
G C T T A T T T G C C A C A T ALA TYR LEU PRO IJIS
525
A G A A G A C C A T T C A A T ARG ARG PRO PHE ASN
415 G C A G A T
ALA ASP
425 435 445 455
C A A G G T C A A C C A G A G T A T C A C G C T C A C T G T G A A G G C A G G
GLN GLY GLN PRO GLU TYR HIS ALA HIS CYS GLU GLY ARG
475 485 495 505 515
A G A A T G G G G A A G A C C C C T C C C A T G C T C A A T G T AC C A G A G C A C T T C
ARG MET GLY LYS THR PRO PRO MET LEU ASN VAL PRO GLU HIS PHE
535 545 555 565 575
A T A G G T C T T T A C A A G G G A A C G A T T G A G C T C A C A A T G A C C A T C T A C ILE GLY LEU TYR LYS GLY THR ILE GLU LEU THR MET THR ILE TYR
585 595 605 6 15 625 635
G A T G A T G A G T C A C T G G A A G C C G C T C C T A T G A T C T G G G A T C A T T T T A A T T C T T C C A A A T T T ASP ASP GLU SER LEU GLU ALA ALA PRO MET ILE TRP ASP HIS PHE ASN SER SER LYS PHE
645 655 665 675 685
T C T G A T T T C A G A G A G A A G G C C T T A A T G T T T G G C C T G A T T G T C G A G G A A A A G SER ASP PHE ARG GLU LYS ALA LEU MET PHE GLY LEU ILE VAL GLU GLU LYS
705 715 725
G C T T G G G T C C T G G A T T C T G T C C G C C A C T T C A A A T G A
ALA TRP VAL LEU ASP SER VAL ARG HIS PHE LYS *00
695 G C A T C T G G A ALA SER GLY 735
FIG. 1. NucleotidesequenceofcDNAinthetranslatedregion of theGlasgow M-proteingene andpredictedamino acidsequence.The foursynthetic primers used in thisstudyareunderlined. Numberingis from3' endofthe M geneas reportedbyRose andGallione(19).
5'-End labeling ofprimers. Four synthetic primers were
synthesized by D. H. L. Bishop, University of Alabama, Birmingham. Thesequenceswerebased onthe sequence of M-protein mRNA published byRose and Gallione(19) and chosen tohybridizewithregions about180residues apart in the viralgenome(seeFig. 1). Theprimersweretreated with calf intestinal phosphatase (45°C for 45 min). After heat inactivation of thisenzyme,theprimerswerelabeledattheir 5' ends with T4 polynucleotide
kinase
(P-LBiochemicals)
and
[-y-32P]ATP
(ICN
Pharmaceuticals Inc.).Primer extension. Labeled primer was annealed to viral genomicRNAtemplate by incubation at room temperature for10minin the presence ofmethylmercurichydroxideand extended by using reverse transcriptase (Seikagaku Amer-ica, Inc.). The reaction mixture (50 ,ul) contained 50 ,ug of viralgenomicRNA, 50 x 106cpmof end-labeled primer, 10
mM methylmercuric hydroxide, 10 mM dithiothreitol, 50 mMTris hydrochloride(pH8.3), 6mMmagnesiumchloride, 110 mM potassium chloride, the four deoxynucleoside tri-phosphatesat 1 mMeach,and 40 Uofreversetranscriptase. The reaction. was incubated at 37°C for 2 to 3 h and terminated by theaddition of5 ,ulof 500mM EDTA.
Sequenceanalysis. The template in the extended reaction
product was hydrolyzed by the addition of 0.3 M sodium
hydroxide (30 minat50°C).Afterextraction withphenoland ether, the dried reaction productwastaken up in 20to30,ul of 10 mM Tris-1 mM EDTA(pH 7.5) and passedthrough a column(30 by 0.9cm) ofSephadex G-100to removeshorter cDNA strands arisingfrom specific stops, whichwere
pre-sentin everyreactionproduct.Thevoid volumewaspooled andprecipitated withalcohol in the presence of 20to 30,ug of tRNA as carrier. The dried material was dissolved in waterand sequenced bythemethod of Maxam andGilbert (12).
Electrophoresis. Nonequilibrium pH gradient electropho-resis (14) was carried out by a modification described by Sanders et al. (20) for resolution of very basic proteins. Virions were solubilized in buffer containing 0.3 M NaCl with protamine as described in footnote 4 of reference 20. Only the first (pH gradient) dimension was run, permitting side-by-side comparison of M protein from different mu-tants.
RESULTS
The nucleotide sequence of the M-protein gene of the Glasgow wild type of VSV, with the deduced amino acid
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.64.561.67.469.2]TABLE 1. Differences between M-genesequencesof Glasgow andOrsay strains of VSV Indiana andpublished sequence Base Triplet
sequence'
Amino acid change number" (19) Glasgow Orsay (position)136 AGC AAC AAC Ser- Asn(32)
200 ACC ACT ACT _C
201 TAT CAT CAT Tyr His(54)
210 AAT CAT Asn His(57)
359 AAA AAG AAG
-438 ACT GCT GCT Thr - Ala(133)
446 TGC TGT TGT
-470 CAT CAC
-473 AGG AGA AGA
-552 ATT GTT Ile-- Val(171)
575 TAC TAT
-596 GCA GCC
620 TTC TTT
681 AAA GAA Lys-* Glu(214)
698 GCG GCT GCT
-714 ATC GTC GTC Ile-- Val(225)
717 AGC CGC Ser-*Arg(226)
aNumberingas inFig.1.
IAlteredbase is in boldface type.
c-,Identicaltopublishedsequence.
sequence, isshown in Fig. 1. It is importantto notethat the sequencing strategy used yields a consensus sequence
di-rectly; clones, each one necessarily derived from a single
copy of thegene, were not used. This hasproved fortunate inthelight of therecentdiscovery by Schubertetal. (23) of extensiveheterogeneity in cDNA clones prepared from the VSVLgene.The sequenceshown inFig. 1 differs from that
reportedfor this region of the Mgeneof the San Juan strain
by Rose and Gallione (19) in 13 locations, involving six amino acidsubstitutions(Table 1). The Orsay wildtypealso differs from that of Rose and Gallione by 13 nucleotides, resulting in six amino acid changes (Table 1). The Glasgow andOrsay wildtypes differfrom each other somewhatless, by eight nucleotides and four amino acid changes (Table 1). Thedifferences betweenoursequencesand that of Rose and Gallione (19)may arise(i) from differences in the strains of
VSV used or (ii) from the fact that Rose and Gallione
reported the sequenceofasingle cDNA clone derived from
M protein mRNA, whereas we obtained consensus
se-quencesby using thegenomeas template.
The first primer was chosen to hybridize with a region
encompassing the first 13 residues of the coding region to
minimize the possibility of hybridization with undesired regions of the viral genome, e.g., sequences coding for the
ribosome-binding regions. We have therefore been unableto
verify directly the first five amino acids (Met-Ser-Ser-Leu-Lys) in the sequence deduced by Rose and Gallione (19).
However, thesequence mayreasonably be assumedtobegin with Met in the mutants as well as in the wild type. In addition, the fact that extended productswereobtainedfrom
all templates with this primer attests to the fact that its 3'-terminal nucleotide (A) is complementary to the viral
genomeinall themutants;since this is theonly nucleotide of the Lys triplet present in the primer and the other two nucleotides were sequenced in this study, Lys must be
presentasamino acid 5 in all themutants. Only three amino acidresidues, in positions 2 through 4, thus remain undeter-mined. However, in an early experiment carried out in collaboration with J. Perrault and M. McClure, Washington UniversityMedical School, St. Louis, Mo., thesequenceof
[image:3.612.316.560.89.162.2]thefirst 67nucleotides of the ts023Mgene,whichincluded
TABLE 2. Base and amino acidchangesin M-protein mutants of
vSv
Mutant Wildtype Baseno. Base Aminoacid changea change"(position)
tsG31 Glasgow 684 A G Lys Glu(215)
tsG33 Glasgow 652 A C Lys Thr(204)
681 G A Glu Lys(214)
tsO23 Orsay 103 G - A Gly- Glu(21)
tsO89 Orsay 439 C A Ala - Asp (133)
aRelativetotheappropriate wild type.
coding sequences for the first nine amino acids, was deter-minedby extension of a 33-nucleotideprimer(NS gene bases 757 through 790) originallyused by Rose insequencing the NS-M intergene region (18). The tsO23 sequence differed fromthat of Rose and Gallione in asingle nucleotide (G at
position 13 to A) in the noncoding region of the gene. Thus, nodifferences were found between the amino acid sequence of the N-terminal region of Mprotein deducedbyRoseand Gallione and that deduced by us for ts023.
The sequence differences between thegroup III mutants and theirrespective wild types are shown in Table 2. tsG31 differs from the Glasgow wild type inonly a single nucleo-tide, which changes lysine at position 215 to glutamic acid. Similarly, ts023 and tsO89 each differ from the parent Orsay wild type in a single position, 21 and 133, respectively; in each anacidic amino acid substitutes for a neutral one. tsG33 has two changes, lysine at position 204 to threonine and glutamic acid at position 214 to lysine. However, in the wild-typesequence deducedbyRoseandGallioneandin the Orsay wild type, a lysine residue ispresent atposition 214. It may be, therefore, thatsubstitution atthis position does not give rise to the temperature-sensitive phenotype; the only relevant change would then be thelysine-to-threonine changeat position 204.
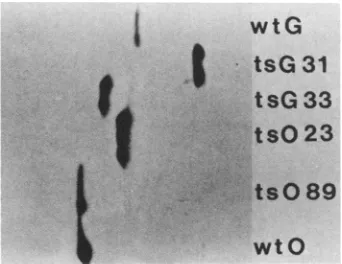
Anonequilibrium pH gradientelectrophoresis gel showing the relative migrationof the two wild-typeandfour mutant M proteins used in this study is shown in Fig. 2. All six proteins could becompared in asingle slab gel, since M is theonlybasicproteinin theVSV virion. The identity of the
wtG
tsG 31
~~~~t
S23
0
.1
t.t089
[image:3.612.61.299.92.297.2]wtO
FIG. 2. One-dimensional nonequilibrium pH gradient electro-phoresis gel comparing M proteins from wild-type and mutant strains ofVSV, stained with Coomassie blue. The pH increases towardthe bottom ofthegel.
on November 10, 2019 by guest
http://jvi.asm.org/
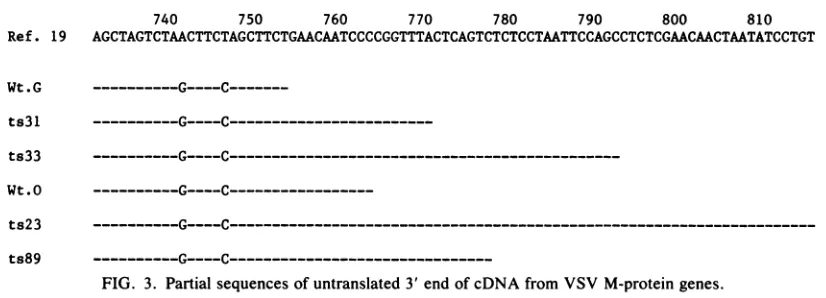
[image:3.612.354.525.547.679.2]740 750 760 770 780 790 800 810 Ref. 19 AGCTAGTCTAACTTCTAGCTTCTGAACAATCCCCGGTTTACTCAGTCTCTCCTAATTCCAGCCTCTCGAACAACTAATATCCTGT Wt.G
---G----C---ts3l
---G----C---.-ts33 ---G----C---.-Wt.0
---G----C---ts23 ---G----C---ts89
FIG. 3. Partial sequences of untranslated3' endof cDNA from VSV M-protein genes.
single prominent band in each lane as M protein was confirmed by two-dimensional electrophoresis (data not shown). The gel in Fig. 2 confirms several predicted rela-tionships betweenMproteins: (i)the Orsaywildtypeis more basic than the Glasgow wild type, (ii) tsG33 is more basic than the Glasgow wild-type, (iii) tsG31 is less basic than Glasgow wild type, and (iv) ts023 is less basic than the Orsay wildtype.The Mprotein ofts089,on theother hand, is identicaltothat of itsparentOrsay wildtype,whereas the deducedsequencepredictsthatitshouldinsteadbeidentical to ts023; we have no explanation for this discrepancy. The finding ofa single M-protein band ineachlane is consistent with an earlier observation (4), suggesting that phosphory-lationisrestricted to a small fraction (5 to 10%) of viral M protein; the single M-protein bands in Fig. 2 therefore presumably correspond tothe unphosphorylated form.
Partial sequences of the 3' noncoding regions of the M-gene cDNA which were also obtained in this study are shown in Fig. 3. Two differences from the published se-quence(19) werefound in all sixgenes: A(position741) to G,and T (position746) to C.
DISCUSSION
In comparing the sequences for wild-type M protein of threedifferent strains-thetwoin thispaperandthe oneof Rose and Gallione
(19)-it
is noteworthy that of the eight amino acid changes observed, only two involved highly basicoracidicresidues,andneither of these wassharedby bothGlasgow and Orsay wildtypes. Inposition 214, the Lys ofRoseandGallione is GluintheGlasgow wildtype but not in the Orsay wild type. Further, thisresidue is also Lys in tsG33 (which has anadditional charge change; Table 2). It appears that achangefrompositivetonegative chargeinthisposition
does not affect the wild phenotype. The other change was at position 226: Ser of Rose and Gallione becomes Arg in the Glasgow strain, but not in the Orsay strain. Itmay be that this position, onlyfour residues from the C terminus, is notessential forproteinfunction.By contrast, all ofthe nucleotidechanges in the mutants causedamino acid changes, and these allinvolved basic or
acidic residues. Allfourmutantshad onepositionatwhicha
charge change of-1 or -2occurred (Table2). The one +2 change found, atposition 214in tsG33, is compatible with the wild phenotype, as discussed above. It is noteworthy that, in all the sequences examined, residues 214 and 215 were both charged; a charge of either +2 or 0 in these
positions
isapparentlycompatiblewith thewildphenotype, butachargeof -2 is not, as shownby tsG31 (Table 2).Allfourmutantshavepreviously beenfoundtohavevery
similar phenotypes, characterized by temperature sensitiv-ity, decreased binding tonucleocapsids,and increased bind-ingtomembranes (7, 10, 16, 17, 24, 26). Thefactthat these two coordinate affinity changes arise from a single amino acid substitution suggests that the decreased binding to nucleocapsids may represent the primary effect ofthe mu-tation, since mutations more commonly decrease specific interactions than increasethem. The mutations are so widely distributedalong the polypeptide chain, however, that they are difficult to interpret in terms ofa specific binding site. Since all of the changes involvechargedresidues, and since Mproteinbindsnucleocapsids by ionic interactions (26),it is possible that all the mutations perturb the nucleocapsid binding site directly.Inthis case,thesitemusteitherbevery large, or, more likely, arise from a specific folding ofthe proteinthatbringsthesewidelyseparatedresidues together. Alternatively, the mutations may cause conformational changes that destabilize the protein conformation or de-creasetheaccessibility ofthebinding site.Ineithercase, the unusually high rateof spontaneous reversion found forthe group III temperature-sensitive mutants (16) suggests that the specificity requirements forwild-type function may not be very stringent, since manydifferentpoint mutationscan evidentlyactequivalently to restorethe wild (temperature-stable) phenotype. Experiments are in progress to charac-terize temperature-stable revertants of the mutants de-scribedinthispaper.
ACKNOWLEDGMENTS
The experttechnical assistance of Roger Vanderoef isgratefully acknowledged. WeareindebtedtoM. M.Sanders for instruction in theisoelectric geltechnique and toY.Ninomiya for instruction in
sequencing methodology. J.L. is indebted to J. Perrault and M. McClure forproviding the resources of theirlaboratory andtheir personal expertise during his sabbatical leave at Washington University Medical School.
This workwassupported by Public Health Service grant AI-13003 from the National Institutes ofHealth.
LITERATURE CITED
1. Bishop, D. H. L., and M. S. Smith. 1978. Rhabdoviruses, p. 167-280. In D. P. Nayak (ed.), Molecular biology of animal viruses. MarcelDekker, Inc., NewYork.
2. Carroll, A. R., and R. R. Wagner. 1978. Reversal by certain polyanions of an endogenous inhibition of the vesicular stomatitis virus-associated transciptase. J. Biol. Chem. 253: 3361-3363.
3. Carroll,A.R.,and R. R.Wagner. 1979.Roleof themembrane (M)protein in endogenous inhibition of in vitrotranscription by
vesicular stomatitisvirus. J. Virol.29:134-142.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.101.514.67.216.2]4. Clinton, G. M., B.W.Burge,andA.S. Huang. 1978. Effects of phosphorylation and pH on the association of NS protein with vesicularstomatitis virus cores. J. Virol. 27:340-346.
5. Clinton, G. M., S. P. Little,F. S. Hagen, andA. S.Huang.1978. The matrix (M) protein of vesicular stomatitis virus regulates transcription. Cell 15:1455-1462.
6. Combard, A., and C. Printz-Ane. 1979. Inhibitionof vesicular stomatitis virus transcriptase complex by the virion envelope M protein. Biochem. Biophys. Res. Commun. 88:117-123. 7. Knipe,D., H. F. Lodish, and D. Baltimore. 1977.Analysis of the
defectsof temperature-sensitive mutants of vesicular stomatitis virus: intracellular degradation of specific viral proteins. J. Virol.21:1140-1148.
8. Lafay, F. 1974. Envelopeproteinsof vesicular stomatitis virus: effect of temperature-sensitive mutations in complementation groupsIII and V. J. Virol. 14:1220-1228.
9. Lenard, J., D. A.Mancarella, T. Wilson, J.A. Reidler,P. M. Keller, and E.L. Elson. 1982. The M protein of vesicular stomatitis virus: variability oflipid-protein interaction compat-ible with function. Biophys. J. 37:26-28.
10. Mancarella, D. A., andJ. Lenard. 1981. Interactions of wild-type and mutant M protein of vesicular stomatitis virus with viral nucleocapsid and envelope in intactvirions. Evidence from
[I25]iodonaphthyl azide labelling and specific cross-linking. Biochemistry 20:6872-6877.
11. Martinet, C., A. Combard, C. Printz-Ane, . nd P.Printz. 1979. Envelopeproteins and replication of vesicular stomatitis virus: in vivo effects of RNA+ temperature-sensitive mutations on viral RNA synthesis. J. Virol. 29:123-133.
12. Maxam, A. M., and W.Gilbert. 1980. Sequencing end labelled DNAwith base-specificchemical cleavages. Methods Enzymol. 65:499-560.
13. Miller, D. K., and J. Lenard. 1980. Inhibition of vesicular stomatitis virus infectionby spikeglycoprotein. Evidence foran intracellular,G-protein-requiring step. J. Cell Biol. 84:430-437. 14. O'Farrell, P. G., H. M. Goodman, andP.H. O'Farrell. 1977. High resolution two-dimensional electrophoresis of basic as well as acidicproteins. Cell 12:1133-1142.
15. Pepinsky, R. B., andV.M. Vogt. 1979.Identification of retro-virusmatrix proteins bylipid-protein cross-linking. J. Mol. Biol. 131:819-837.
16. Pringle, C. R. 1975. Conditional lethal mutants of vesicular stomatitis virus.Curr. Top. Microbiol. Immunol. 69:85-116.
17. Reidler, J. A.,P. M.Keller, E. L. Elson, and J. Lenard. 1981. A fluorescence photobleaching study of vesicular stomatitis virus infected BHK cells. Modulation ofG protein mobility by M protein. Biochemistry 20:1345-1349.
18. Rose, J. K. 1980. Complete intergenic and flanking gene se-quences from the genome of vesicular stomatitis virus. Cell 19:415-421.
19. Rose, J. K.,andC. J.Gallione. 1981. Nucleotide sequences of the mRNA'sencoding the vesicular stomatitis virus G andM proteins determined fromcDNAclonescontaining the complete coding region. J. Virol. 39:519-528.
20. Sanders, M. M., V. E. Groppi,Jr., and E. T. Browning. 1980. Resolution of basic cellularproteins including histone variants by twodimensionalgelelectrophoresis: evaluation of lysineto arginine ratios and phosphorylation. Anal. Biochem. 103: 157-165.
21. Schnitzer, T. J., C.Dickson, and R. A. Weiss. 1979. Morpho-logical and biochemical characterization of viral particles pro-duced by the tsO 45 mutant of vesicular stomatitis virus at restrictive temperature. J. Virol. 29:185-195.
22. Schnitzer,T.J.,and H.F. Lodish.1979.Noninfectious vesicular stomatitis virusparticles deficient in the viral nucleocapsid. J. Virol. 29:443-447.
23. Schubert, M., G. G. Harmison, and E. Meier. 1984. Primary structureof the vesicular stomatitis viruspolymerase (L) gene: evidence for a high frequency of mutations. J. Virol. 51: 505-514.
24. Stoffel, W., C. Schreiber, andH. Scheefers. 1978. Lipids with photosensitive groups as chemical probes for the structural analysis of biological membranes: onthelocalization oftheG and Mprotein of vesicular stomatitis virus. Hoppe-Seyler'sZ. Physiol. Chem.359:923-931.
25. Weiss,R.A.,and P. L. P. Bennett.1980.Assembly of membrane glycoproteins studied by phenotypic mixingbetween mutantsof vesicular stomatitis virus and retroviruses. Virology 100: 252-274.
26. Wilson, T.,andJ. Lenard. 1981. Interaction of wild type and mutantM protein of vesicular stomatitis virus with nucleocap-sids in vitro. Biochemistry 20:1349-1354.
27. Zakowski, J. J., and R. R. Wagner. 1980. Localization of membrane-associated proteins in vesicular stomatitis virus by use of hydrophobic membrane probes and cross-linking re-agents. J. Virol.36:93-102.