JOURNAL OF VIROLOGY, Apr. 1977, p.74-85 Copyright©1977 AmericanSociety for Microbiology
Vol.22,No.1 Printed in U.S.A.
Analysis of Intracellular Feline Leukemia Virus Proteins
II.
Generation
of Feline
Leukemia Virus
Structural Proteins from
Precursor Polypeptides'
GREGORY F. OKASINSKI AND LELAND F. VELICER*
Department ofMicrobiology and Public Health, Michigan State University, East Lansing, Michigan 48824 Received for publication 17 August 1976
The synthesis and processing of feline leukemia virus (FeLV) polypeptides
werestudied inachronically infected feline thymustumorcellline, F-422, which produces the Rickardstrain ofFeLV. Immune precipitation with antiserum to FeLVp30 andsubsequent sodium dodecylsulfate-polyacrylamidegel
electropho-resis (SDS-PAGE) were used to isolate intracellular FeLV p30 and possible
precursor polypeptides. SDS-PAGE of immune precipitates from cells
pulse-labeled for2.5min with[355]methioninerevealedthe presenceof a 60,000-dalton
precursorpolypeptide(Pp6O)aswell as a30,000-daltonpolypeptide. Whencells
were grown inthepresenceof theproline analogueL-azetidine-2-carboxylicacid,
a70,000-daltonprecursorpolypeptide (Pp7O)wasfoundinadditiontoPp6Oafter
a 2.5-minpulse. The cleavage of Pp6O could be partially inhibited by the general
protease inhibitor phenyl methyl sulfonyl fluoride (PMSF). This partial
inhibi-tionwasfoundto occuronly if PMSFwas presentduring pulse-labeling.
Intra-cellular Pp7O and Pp6O and FeLV virion p70, p30, p15, p1l, and p10 were
subjectedtotrypticpeptideanalysis. The resultsof this trypticpeptideanalysis demonstrated that intracellular Pp7O and virionp70 were identical and that both contained the tryptic peptides of FeLV p30, p15, p1l, and plO. Pp6O contained thetrypticpeptides of FeLVp30, p15, andplO,butlacked the tryptic
peptidesofp1l.The results of pactamycingeneordering experimentsindicated thatthe small structural proteins of FeLVareorderedp11-pl5-plO-p30. The data indicate that the small structuralproteins of FeLV are synthesizedas partofa
70,000-daltonprecursor. Acleavage schemefor thegenerationofFeLVp70, p30,
p15, p1l, andplO fromprecursorpolypeptides isproposed.
The generation of stable virion proteins from large precursor polypeptides is a well-docu-mented phenomenon observedinmanyanimal virus-infected cells. Post-translational cleavage
of precursor polypeptides was initially
de-scribed forpoliovirus-infected cells (8, 23). This phenomenon has been showntobewidespread, occurring in cells infected by coxsackievirus
(11), Sindbisvirus (21, 22),Semlikiforestvirus
(4, 12, 15),mengovirus (20),and
encephalomyo-carditis virus (3, 14). The synthesis of large
precursor polypeptides has beenproposedas a
mechanism by which polycistronic mRNA is
translatedineukaryotic cells (2).
During the past 3 years several reports of
precursorpolypeptidesinoncornavirus-infected
cells have appeared. Avian myeloblastosis
vi-rus-infected primary chicken fibroblasts
con-tain a76,000-daltonprecursorpolypeptide(27).
Trypticpeptide analysis of thisprecursor poly-'Articleno. 7743from theMichiganAgricultural Exper-imentStation.
peptidereveals thepresence of four virion
pro-teinswithin the76,000-daltonmolecule(27, 28).
Immuneprecipitation of Rauscher leukemia vi-rus (RLV) proteins from infected cellswasused
to demonstrate 82,000- and 65,000-dalton
pre-sumptive precursor polypeptides in JLS-V9 cells (26) and 200,000-, 90,000-, 80,000-, and
65,000-daltonpolypeptides inJLS-V16cells (1, 16).Jamjoometal. (9)demonstratedthat RLV
contains anon-glycosylated 70,000-dalton
poly-peptide thatisidentical to anintracellular
pol-ypeptide,immuneprecipitable byantiserumto
RLV. The virion and intracellular
70,000-dal-ton polypeptide contain RLV p30 tryptic
pep-tides, and the authors suggest a possible
pre-cursor-productrelationship.
Recently, we identified a 60,000-dalton
pre-cursor polypeptide (Pp6O) of feline leukemia
virus (FeLV) p30 (18). We employed both
im-muneprecipitation ofFeLVpolypeptidesfrom
pulse-labeled cells and trypticpeptide analysis
ofPp6OandFeLVp30todemonstratea
precur-74
on November 10, 2019 by guest
http://jvi.asm.org/
sor-productrelationship. In thisreportwe
pre-sent evidence that: (i) there exists a
70,000-dalton precursor polypeptide (Pp7O) of FeLV
p30, p15, p1l, andplO; (ii)thecleavageofPp7O
results in the generation of Pp60 and p1l; (iii)
the cleavageofPp6O can bepartiallyinhibited
by the general protease inhibitor phenyl
methyl sulfonyl fluoride (PMSF); (iv)
intracel-lular Pp7O and FeLV virion p70contain
identi-cal tryptic peptide profiles; and (v) the gene
order of the low-molecular-weight FeLV
struc-turalproteinsis pll-p15-plO-p30.
Our evidenceindicates that the small
struc-tural proteins of FeLV differ from those of
avianvirusesinbotharrangement withinand
cleavagefromprecursorpolypeptides. FeLV
ap-pears tobesimilartoRLV inthat bothcontain
uncleaved precursors of structural protein (9;
present publication). The inhibitionof precur-sor cleavage by a general protease inhibitor reported here seemstobe uniqueforFeLV.
(Most ofthis work was submitted by G. F.
Okasinski inpartial fulfillment of the
require-ments for the degree ofDoctor of Philosophy.
This work was presented in part at the ICN-UCLA Winter Conferences on Molecular and
Cellular Biology, 14-19 March 1976, Squaw Valley, Calif., and at the Cold SpringHarbor
meeting on RNA Tumor Viruses, 26-30 May
1976, ColdSpring Harbor, N.Y.)
MATERIALS AND METHODS
Source of cells and virus. Thechronicallyinfected feline thymus tumor cell suspension (F-422) was
used throughout these experiments. This cell line
produces the Rickard strain of FeLV andwas
propa-gatedaspreviouslydescribed (7).
Radioactive labeling of cells. F-422 cells were pulse-labeled with [35S]methionine (1 uCi/106cells)
or3H-aminoacid mixture(2jiCi/106 cells).Thepulse wasterminated and cells werechased in the pres-enceof10timesthe normal concentration of
methio-nineaspreviously described (18).
Purification of FeLV. 14C0 or3H-amino acid-la-beled FeLV and unlaacid-la-beled FeLVwerepurifiedfrom F-422 culture fluids by discontinuous sucrose gra-dientcentrifugationaspreviouslydescribed (18).
Preparation of subcellular fractions. Particulate
fractions (PF) and cytoplasmic extracts (CE) were
preparedfromNonidet P-40(NP-40)-disruptedcells
aspreviouslydescribed(18). Briefly, cellswere
dis-rupted withlysisbuffer (0.5% NP-40-0.15 M NaCl-0.01 MTris [pH7.4]) andcentrifugedat2,400rpm for5 min in anInternational PR-6centrifuge. The supernatantwasremoved andcentrifugedat100,000
x g for 1 h. The100,000 xgsupernatant (cytoplas-micextract)wasremoved,andthepelletwas solubi-lized withlysisbuffercontaining0.2%deoxycholate (PF). Both the CE andPF weresonicallytreated for 2 to 3 minin a 150-W Bransonultrasonic cleaner (Branson Instruments Co., Stamford, Conn.) and
centrifugedagainat100,000 xg for1h.
Immuneprecipitation.Rabbit antiserumtoFeLV
p30 was prepared and clarified as previously de-scribed (7, 18). Rabbit antiserumto bovine serum albumin (BSA) was obtained from E. Sanders
(Michigan State University). Clarified antisera wereaddedtosubcellularfractionsordisrupted vi-rusandincubated for30minat37°C andthen
over-nightat4°C.Immuneprecipitateswerecollectedby
sedimentation through 1mlof 10% sucrose (wt/wt) inlysisbufferat2,000rpmfor 20 min inan Interna-tional PR-6centrifuge. Thiswasrepeatedone addi-tionaltime,and thefinalprecipitatewassolubilized for sodiumdodecyl sulfate-polyacrylamidegel
elec-trophoresis (SDS-PAGE) as previously described (18).
Doubledetergent disruptionofFeLV. Labeledor unlabeledpurified FeLVwasdisrupted by suspen-sion of viral pelletsinlysisbuffercontaining0.5%
deoxycholatefollowedby5min ofsonictreatmentin a 150-WBranson ultrasonic cleaner. Thedisrupted
viruswascentrifugedat100,000xgfor1hin anSW 50.1rotor(Beckman),andthe supernatantwasused as a source oflabeled or unlabeled soluble FeLV
polypeptides.
SDS-PAGE. Electrophoresis in the presence of SDS was.doneaspreviouslydescribed (18), usinga 9%polyacrylamidegeldescribedbyFairbanksetal. (5). Thegelswerefractionated and assayed for
ra-dioactivity, using 3a70b scintillation cocktail (RPI
Corp., Elk Grove Village, Ill.) as previously de-scribed (7).
Tryptic peptide analysis. Tryptic peptidesof
im-mune precipitated polypeptides and labeled FeLV
polypeptides, eluted fromSDS-polyacrylamidegels,
werepreparedaspreviouslydescribed (18). A1-mg
amount ofBSA wasadded to FeLVproteins sepa-ratedby gelfiltration in the presence of 6 M guani-dinehydrochloride (GuHCl),and theproteinswere
precipitatedbydilution of GuHClto0.6Mfollowed
by the addition oftrichloroacetic acid to a final concentration of 25%. Tryptic peptides were then
prepared as previously described (18).
Cation-ex-change chromatographyoftryptic peptides wasdone
byusingahigh-pressurecolumn of type P
chromo-beads (Technicon) as previously described (18).
Tryptic peptides were lyophilized and stored at
-76°C untilchromatography.
Pulse-labeling in the presence of pactamycin.
Pactamycinwasobtained from George B. Whitfield
of theUpjohnCo. Thedrugwasstored as a 5 x 10-5 Msolution in1 mMacetic acid at-20°C.
A total of 250 x 106 cells wereincubated for 30s
withpactamycinat afinal concentration of 5 x 10-7
M andpulse-labeled with 3H-amino acid mixture(2
,4Ci/100
cells)or"4C-aminoacidmixture (0.21iCi/106cells)inafinal volume of 5 ml.
RESULTS
EffectofPMSFonthecleavage of Pp6O.To determine the effect of PMSF oncell viability
andprotein synthesis duringtheshortperiod of exposuretobe employed in these
experiments,
cellswereincubated withPMSFfor 2 min and
thenpulse-labeledfor 2.5 min in the presenceof
on November 10, 2019 by guest
http://jvi.asm.org/
76 OKASINSKI AND VELICER
PMSF. Both hot trichloroacetic acid-precipita-bleradioactivity and trypan blue dye exclusion weremonitored. The results of this experiment (Fig. 1)indicated that PMSF reduced trichloro-aceticacid-precipitableradioactivity by 40% (at
50 jug/ml)during the short period of exposure used in these experiments and reduced cell via-bility by 10to 15%.
Pulse-chase-labeling experiments weredone inthe presenceof PMSFtodeterminewhether PMSF, a general protease inhibitor, could in-hibit the cleavage of Pp6O. In experiment 1
(Fig. 2)cells werepulse-labeledfor2.5 minand
then chased for 30 min in the presence or
ab-senceof PMSF. SDS-PAGE of immune
precipi-tatesfrom subcellular fractions ofpulse-labeled
cells demonstrated the presence ofPp6O in the
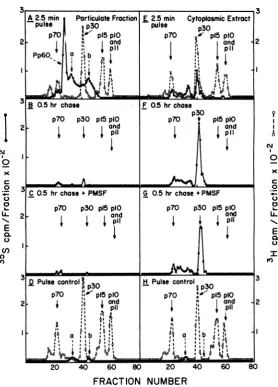
PFandp30inthe CE (Fig. 2A andE)as previ-ouslyreported(18). When cellswerechased for 0.5 hinthe presenceorabsence ofPMSF, Pp6O
wasabsentfrom both subcellularfractionsand p30wasfoundmainlyinthe CE(Fig. 2B, F, C,
andG), indicatingno inhibition ofPp6O cleav-age.
Theeffect ofPMSFonPp6O cleavagewasalso
examined with the protease inhibitor present duringpulse-labeling. Inthis experiment(Fig.
3) cells were incubated with PMSF for 2 min
and thenpulse-labeledfor 2.5min inthe
pres-ence of PMSF. SDS-PAGE of immune
precipi-tates from subcellular fractions of cells
pulse-100
III
4) 0
al
100 E
.0
50
75 o
.6
0..
50 <
25
250 0
0 10 20 30 40 50
1sg PMSF
FIG. 1. Effect ofPMSFoncellviability and
pro-teinsynthesis. Six aliquots ofcells (50 x106cellsper
aliquot)wereincubatedwith0, 5,10,15,20,or50pg
ofPMSFpermlfor2minandthenpulse-labeled for
2.5min with 10 uCiof[35S]methionineperaliquot in
thepresenceofPMSF. The pulse-labelingwas
termi-nated, and the cells were chased for 3.0 h in the
presenceofPMSFasdescribed inthetext.Cell via-bility was determined by trypan blue dye exclusion immediately afterthepulse-label (4.5-minexposure
toPMSF) andat1.5and3.0h. Theeffect of PMSF
onprotein synthesiswasdeterminedbyassayinghot
trichloroacetic acid-precipitable radioactivity from NP-40-disruptedcells.
labeledinthe presence of PMSFdemonstrated the presenceofPp6Oinboththe PF and CE and
little, if any, p30 (Fig. 3A andD). When cells
pulse-labeled in the presence of PMSF were
chased in the presenceor absence of PMSF for
0.5 h, Pp6O was present in both subcellular fractionsaswas asmallamountofp30, indicat-ingpartial inhibitionofPp6Ocleavage.
Nonspecific immune precipitationwas
moni-tored by incubating labeled subcellular frac-tions with5,ug of BSA and 200 ,ul of anti-BSA. Figures 2D and H depict typical results
ob-tained from subcellular fractions of pulse-la-beled cells. We routinely detect only two poly-peptides (labeledaandb) in all control experi-ments. Identical results were obtained from subcellular fractions of cells incubated with PMSF (data not shown).
Effect of L-azetidine-2-carboxylic acid on
the synthesis ofFeLV precursor polypeptides. Wehave previously demonstrateda precursor-productrelationship between Pp6O and p30 (18). Several amino acidanalogues have been tested for their effectonthe appearance and process-ing of FeLV precursor proteins. Of the
ana-logues tested (fluorophenylalanine,
canavan-ine, andL-azetidine-2-carboxylic acid), only
L-azetidine-2-carboxylic acid has yielded any de-tectable effect. SDS-PAGE ofimmune
precipi-tates from subcellular fractions of cells pulse-labeledinthepresence of L-azetidine-2-carbox-ylic acid demonstrated the presence of large amounts of70,000-daltonpolypeptide (Pp7O)as
wellasreducedlevels ofPp6O and p30 (Fig. 4).
A comparison of these results (Fig. 4) with those seeninFig. 2AandEindicated that the 70,000-daltonpolypeptideseen inrelatively low amounts in the absence ofanalogues was now
the principal immuneprecipitable polypeptide whenpulse-labeling was donein the presence of L-azetidine-2-carboxylic acid. This suggests that theprolineanalogue may be inserted into the initial translation product and subse-quently inhibit the cleavage step, resultingin
theformation of Pp6O. Control experiments em-ploying immune precipitation of BSA with
anti-BSA yielded results similartothose shown
inFig. 2Dand H (data not shown).
Immune precipitation of p30 and p70 from disruptedFeLV.The presence ofan intracellu-lar 70,000-dalton polypeptide that is both im-muneprecipitable by anti-p30 and electropho-retically identical to virion p70 led us to exam-ine virion p7O for p30 antigenic determinants.
In aprevious report(18)wedemonstrated that the anti-p30 serumcouldonlyimmune
precipi-tateviralp30 from 0.5% NP-40-disrupted virus.
Subsequent experiments have shown that 0.5%
J. VIROL.
I
on November 10, 2019 by guest
http://jvi.asm.org/
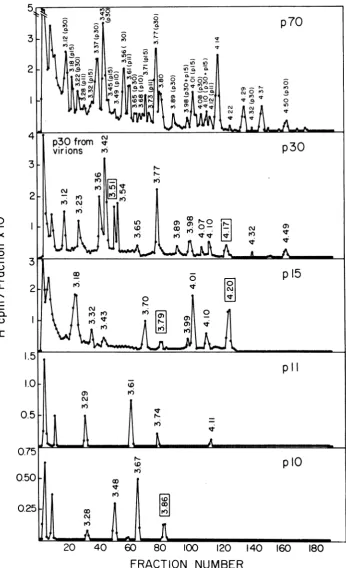
[image:3.508.88.232.393.528.2]20 40 60 80
I I
CY
x
c
0
0 L.
U-N.
E
0.
m
to
20 40 60 80
FRACTION NUMBER
FIG. 2. SDS-PAGE of immune precipitates fromPFand CEof cells incubated with andwithoutPMSF afterpulse-labeling.Atotalof 200 x106cells werepulse-labeledwith 200 pACiof[35S]methioninein4 mlof growth medium. Thepulse-labeledcellsweredivided intofour aliquots,twoof whichwerechasedfor 0.5 h
either withorwithout PMSF(50 pg/ml)and theremaining aliquots whichwereusedasasourceof
pulse-labeled polypeptides. PF and CE were prepared from both pulse-labeled andpulse-chase-labeled cells.
Polypeptides were immuneprecipitated from each fraction with anti-p30 and co-electrophoresed with
3H-aminoacid-labeled FeLVasdescribed in the text. Solidarrows indicate thepositions ofnon-glycosylated
FeLVpolypeptides. (A) and (E) PF and CE, respectively, from pulse-labeled cells; (B) and (F)aPFandCE,
respectively, from cells chased in the absence of PMSF; (C) and (G) aPFandCE, respectively, from cells
chasedinthepresenceof50pgPMSFperml; (D) and (H) SDS-PAGE of immuneprecipitates fromaPF and
CE,respectively, of pulse-labeled cells incubated with5 pgof BSA and200 piof anti-BSA.
NP-40alone doesnotsolubilize viralp70 toany
considerabledegree (datanot shown).
"4C-amino acid-labeled FeLV was disrupted
bytreatmentwith 0.5% NP-40 and sodium
de-oxycholate and then immuneprecipitated with
anti-p30. The detergent disruption procedure
employed in these experiments solubilizes the
major FeLV polypeptides (Fig. 5A), although
only 60 to 70% of the FeLV p70 is solubilized
(data notshown). SDS-PAGE of the resulting
immune precipitate demonstrated two labeled
FeLV polypeptides. Both FeLV p30 and p70
wereimmuneprecipitated (Fig. 5B). FeLVp70
wasonlypartially immune precipitated (30% of
thetotalsolubilizedp70 countsper minute) in
thisexperiment. The dataindicated that FeLV
p70 contained some p30 antigenic
determi-nants. Control experiments using5 ,4gof BSA
and 200 1.l of anti-BSA showed virtually no
precipitation of labeled viral proteins,
indicat-I
CY
x
c
0
0
h-Li
N'.
E
0.
I0
A2.5 min Particulate Fraction E 2.5min CytoplasmicExtract
puSe0 P30 pulse p30
B 0.5 hr chose MS F 0.5 hrchose;PS
p70
p3
p15
plO p70 f p15 plO2 ; and * janid
l~~~~~~~~1
11
3 0.5 hr chase Fontrol}0.5 hrchose +3
p70 p30 p15 plO p70 * p15plO 2- p70 p30 ~p15andl 7 3 1 andp~l
JQ Pulse
control)
H Pulse control,mp30 p30
p70O p15plO p70 p15pIO
2 l and land 2
S I3
4m;
046a,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.508.117.396.63.451.2]78 OKASINSKI AND VELICER
N
0
x c
3' 0
0
LI-0. U) to
to
[image:5.508.123.403.70.380.2]FRACTION NUMBER
FIG. 3. SDS-PAGE of immune precipitates from PF and CE of cellspulse-labeledwith[35S]methioninein the presenceand absence of PMSF. A total of 300 X 106 cells weresuspended in 6 ml of growth medium deficientinmethionine but containing 50 mgofPMSF per ml for2min. The cellswerethenpulse-labeledfor 2.5 minand divided into three aliquots, and two aliquots werechasedinthe presenceorabsenceofPMSF. PF and CEwerepreparedand then immuneprecipitatedwith anti-p30 andelectrophoresedasdescribed in the text.(A) and(D)aPFandCE,respectively, from cellspulse-labeledinthe presenceofPMSF; (B) and (G)a PFand CE, respectively, from cells pulse-labeled and chased in the presenceofPMSF (50pglml);(C) and(F)
aPF andCE, respectively,fromcellspulsedinthe presenceofPMSF and chased in the absenceofPMSF.
20 40 60 80 20 40 60
FRACTION NUMBER
FIG. 4. SDS-PAGE ofimmuneprecipitates the CE and PF of cells pulse-labeled [35S]methionine inthepresenceofL-azetidine-2 boxylicacid.Priortopulse-labeling,100 X 106 were incubatedfor45 min ingrowth medium
cient in methionine and proline but containing azetidine-2-carboxylic acid (021 mglml). The
were pulse-labeled for 2.5 min with 100 [35S]methionine in thepresenceofL-azetidine-2
boxylicacid(021 mg/ml),and thenaCEand
_.
ing little or no nonspecific trapping (data not3 shown).
°2
Tryptic peptide analysis of intracellular2 C
Pp7O,
Pp6O,
and FeLV virionp70,
p30,
p15,
[image:5.508.64.259.475.568.2]p1l, and p10. Intracelular 3H-amino
acid-la-| beledPp7OandPp6Otrypticpeptideswere sub-jected to high-pressure cation-exchange
chro-e
matography
on acolumn oftype
Pchromobeads(Fig. 6). The trypticpeptidefractionsare
iden-from tified
by
the elutionpH
of thepeak
fractionwith
(Fig.
6).Pp7O
contained 29 detectablepeptide
S-car- fractions, whereas Pp6O was resolved into 25
cells
defi-ng
L-cells Ci of
?-car-aPF
were prepared. The fractions were incubated with
300piofanti-p30and5 pgofdisruptedFeLV. The
immune precipitates were collected and co-electro-phoresed with 3H-amino acid-labeled FeLV as de-scribed in the text. Solid arrows indicate the
posi-tionsof labeled FeLVpolypeptides.
A +PMSF Particulate Fraction D+PMSF Cytoplasmic Extract
2.5min pulse plO 2.5 min ~ 1
an u Pp60 |a
p70 - 30
pi5
ir pulse Pp60 and2- :p70 p30 p11pil
Pp60
Lasa
a ~~~~~~~~~~~~b
B +PMSF E+PMSF
Pp60
0.5 hr chase
casd
hrei
and2 p70 p30 p15 pil p70 p30 p15p1l
Pp60
C -PMSF,. F -PMVSF Pp6O
0.5hr chase plO 0.5 hr
and chase andl
2- p70 p30 pI5 piI p70 p30 pI5 p~l
Pp60
b
~~~k~~ b
20 40 60 80 20 40 60 80
1.5
.2 1.0
IL E 05
on
W6
A Particulate Fraction 8Cytoplasmic Extract
p70 P30 p15 PlO p70 p30 pi5pIO
i iand j and
Pp70 1 P070 i
P060 PL60
abb
ir
J. VIROL.
_-
on November 10, 2019 by guest
http://jvi.asm.org/
0
0
u_ 8
U BImmunePrecipitatedp30 p70
E p7O p30p15 plO
i' 6- p
1,J
4-2
0 20 40 60 80
FRACTION NUMBER
FIG. 5. SDS-PAGE of immune precipitatedp30 andp70 fromdetergent-disrupted FeLV. "4C-labeled FeLV wasprepared from 25 x 106cells incubated
with25 uCiof "4C-labeled amino acid mixture for24 h in12.5 mlofgrowth medium. The viruswas
puri-fied as described in the text. The viral pellet was
suspendedin 0.6 mlof lysis buffer containing0.5% NP-40 and 0.5% deoxycholate by 3 min of sonic oscillation in a150-W Branson Ultrasonic Cleaner
(Branson Instruments Co., Stamford, Conn.). The disrupted virus was then centrifuged for 1 h at 100,000 x g. (A) FeLV polypeptides (20,000 cpm) wereprecipitated with 9 volumes ofacetoneand 50
pgofBSA ascarrier and thenelectrophoresed; (B) FeLVpolypeptides (20,000cpm) wereimmune
pre-cipitatedwith 100
41
of anti-p30 and electrophoresedasdescribed in thetext.
peptide fractions. Twenty-three of the 25
pep-tidefractionspresentinPp6Owerealso foundin
Pp7O. Two Pp6Otrypticpeptide fractions, with elution pH values of 3.85 and 3.94, were not
detectableinthetryptic peptide profileofPp7O. Pp7Ocontained,inadditiontothe23Pp6O
tryp-tic peptide fractions, 6 peptide fractions, with
elutionpHvalues of3.28, 3.62, 3.73, 3.81,4.12,
and 4.14. Because Pp7O and FeLV vision p70
were both immune precipitable with anti-p30
(Fig. 4 and 5)and because Pp70 contained the
tryptic peptidefractions ofPp6O (Fig. 5), tryptic peptide analysis of the structural proteins of FeLVwasdonetodetermine: (i)whether
intra-cellular Pp7Oand FeLV virion p70 were
simi-lar, and(ii)whether the additional tryptic pep-tide fractions seen in Pp7O, but not in Pp6O,
could be assigned to any of the FeLV structural proteins.
Tryptic peptide analysis of FeLV virion p70 yielded29 peptide fractions (Fig. 7) thatwere
indistinguishable from the 29 peptides of intra-cellular Pp70. The results demonstrated that intracellular Pp7O andvirion p70 (Fig. 6 and 7) were identical by tryptic peptide analysis. A total of 13 of 15 p30 tryptic peptide fractions were present in virion p70 and intracellular Pp60and Pp7O (Fig. 6 and 7). Virion p15
con-tained 9 tryptic peptide fractions, of which 7
were present in virion p70 and intracellular Pp60 and Pp70 (Fig. 6 and 7).The assignmentof the p15 tryptic peptide fractions with an elution pH of 3.79 remains uncertain due to the low level ofradioactivity found in the p15 tryptic
pepide fractions and thehigh level of radioac-tivity present in the p70 tryptic peptide frac-tions, withelution pH values of3.77and 3.80.
The pH values are accurate to ±0.01 pH unit
and thus could allow assignment of this p15
peptidetoeither the p70peptide fractions 3.77 or3.80.Wechose to consider thispeptideas not
.s
A Pp70
EY0-0020'wt4q 60 80 0 20 14060- ISO
FRACT ION NUMBER
FIG. 6. Tryptic peptideanalysisofPp70and Pp60 eluted from SDS-polyacrylamide gels. Pp70 was prepared by SDS-PAGE of immune precipitates from the PF and CE of 500 x 106cells pulse-labeled for 2.5 min with 500 ,uCiof3H-amino acid mixture in the
presence of L-azetidine-2-carboxylic acid. Pp6O was
prepared in a similar manner from the PF of 500 x 106 cells pulse-labeled(without L-azetidine-2-carbox-ylic acid) with 500 ,uSCi of 3H-amino acid mixture. Both polypeptides were eluted from gels, and tryptic peptides were prepared asdescribed in the text. Tryp-tic peptides were eluted from a column of type P chromobeads, with a linear gradient of pyridine ace-tate (pH3.1to 4.9) at a flow rate of 30 mlh. Various
peptide peaks are identified by the pH of elution determined from reading pH values with a PHM 26 expanded-scale pH meter(Radiometer, Copenhagen, Denmark). Percent recovery ranged from 70 to 80% of the radioactivityelutedfrompolyacrylamide gels.o,
ElutionpH valuesofPp6O trypticpeptidesabsentin
Pp70.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.508.83.222.58.322.2] [image:6.508.262.454.328.468.2]Cl
cJ
go
0
4-O
0
U-E
Q. 0
F')
FRACTION
NUMBER
FIG. 7. Tryptic peptide analysis ofFeLVvirionp70,p30,pl5, pll, andp1O.FeLVp7O,p30,and pl5were prepared by SDS-PAGE of3H-aminoacid-labeled FeLV. FeLVp1l andplOwerepurified by gel filtration of labeled FeLVin GuHCl. Tryptic peptides of thefive virion proteins were preparedasdescribedinthetext. Tryptic peptides were elated and pH values weredetermined as describedin the legend to Fig.6.Percent recovery ranged from 60 to 80% of theradioactivity elated from polyacrylamide gelsorgelfiltration columns. Symbols: A, elation pH values of tryptic peptides absentinvirionp70andPp7O ofFig. 6; ( ),p70tryptic peptides presentin bothp70andvirionproteinsp30, p15,p1l,andplO.
80
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.508.92.439.37.606.2]being present within p70 because of the rela-tively low level of radioactivity. The tryptic
peptidefractions of p1l had elution pH values of
3.29, 3.61, 3.74, and4.11 (Fig. 7). Three of the
p1l tryptic peptide fractions were present in
both virion p70 and intracellular Pp7O, but
wereabsentinthePp6Otryptic peptide profile
(Fig. 6 and 7). Although the p1l peptide
frac-tion eluting at pH 4.11 could not be assigned
unequivocallydue to lowerradioactivity and a
variation of ±0.01 pH units, the presence of a
pH4.12peptidefraction in Pp7O andp70that is
absent in Pp6O is consistent with the other three p1l fractions. These data show that both
virionp70 and intracellular Pp7O contain p1l,
whereasPp6O lacks p1l. Tryptic peptide
analy-sis of virion plO revealed fourtryptic peptide fractions, withelution pH values of 3.28, 3.48,
3.67, and 3.86. Two of the plOtryptic peptide
fractions (pHvalues 3.48and3.67) were present inp7O, Pp7O, andPp60,indicating the presence
of plO in all three molecules. The plO tryptic
peptide fraction with an elution pH value of
3.86 was not present in p70 or Pp7O, but was
presentinPp60, whereas theplOpeptide
desig-nated 3.28isalso seen in p70 and Pp7O, but is
absent in Pp6O. We judge the presence of the plO peptides3.86 and3.28 and thePp6O peptide 3.85 (Fig. 6) tobe expectedanomalies of tryptic
digestionof precursor andproduct proteins.
The tryptic peptide analysis of intracellular
and virion polypeptides was done in parallel
rather than by a double-label procedure,
be-cause of the relatively small amounts of plO
and p1l available from FeLV and the expense
of generating largequantities ofprecursor poly-peptidesof FeLV. Tohelpinsure uniformityin thisparallel analysis, allof thetryptic
peptides
of the proteins examined (Fig. 6 and 7) were
prepared in parallel and stored as
lyophilized
tryptic peptides until chromatography (no
longer than 3 weeks). Tryptic peptides were
eluted with buffers prepared in large
quanti-ties,and the column oftype Pchromobeadswas
washed with sodium hydroxide,
pyridine,
andstarting bufferbefore each run. FeLVplOand
p1l werepurifiedbygelfiltrationinGuHCl(7).
Todetermine the effect ofGuHCl-treated
pro-teinsonthetrypticpeptideprofiles,tryptic
pep-tides were preparedand analyzed from FeLV
p30andp15 preparedfrombothSDS-PAGE and
gelfiltrationsinGuHCl.Polypeptidesprepared
by bothproceduresyieldedqualitatively
identi-cal results when their tryptic peptides were
analyzed by high-pressure cation-exchange
chromatography (data notshown). The
repro-ducibility of the pH valuesassignedtothe tryp-ticpeptides wasexamined from severaltryptic
peptide profiles of FeLV p30 andwasfound to
vary by ±0.01 pH unit (datanotshown).
Pactamycin gene ordering. The order of FeLVp30, p15, p1l, andplO within Pp7O was
determinedbypreferential labeling of the
car-boxy terminal end of newly synthesized poly-peptides. Ourexperimental approachwas simi-lartothatemployed by other workerstoorder the proteins of poliovirus(24, 25), encephalomy-ocarditis virus (3), and recently avian myelo-blastosisvirus (AMV)(28). Inourexperiments pactamycinwasusedtoselectivelyinhibit initi-ation of polypeptide synthesis, while permit-tingchainelongation. Cellswereincubated for
30 swith5 x 1O-7 Mpactamycin, a
concentra-tionrequiring 10 to 15 min tocompletely inhibit
incorporation oflabeled aminoacids, and then
pulsed-labeled in the presence of pactamycin with3H- or
'4C-amino
acid mixtures for5min.Aparallel culturewaspulse-labeledinthe
ab-sence of pactamycin with the opposite (i.e.,
either 3H- or 14C-amino acid mixture) labeled
aminoacidmixture. Afterpulse-labeling, both cultureswerechased for20hand released virus
waspurified from thegrowth medium.Labeled
FeLVproteins wereseparated by gel filtration
inthe presence of6 MGuHClto allow
resolu-tion of FeLV p1l and plO, which are not
re-solvedby SDS-PAGE (7).The relativelabeling ratiosweredetermined from thepercentage of label in each protein synthesized in the pres-enceof pactamycinoverthepercentageoflabel
in each protein synthesized in the absence of
pactamycin.
This experimental approach, based on an
analysisofpolypeptidesinextracellularvirions aftera20-h chase, wasnecessitated by the lack ofhigh-titer monospecificantiseratoFeLVplO
andp1l, which is neededtoeffectively
quanti-tateintracellularpolypeptides. Control
experi-ments with immune precipitation by antisera
to whole virusdemonstratedthat the intracel-lular p30, p15, and p11-plO peaks detected on
SDS-PAGE shortly after a pulse were not de-tectable 20h later(G. Okasinski, unpublished
data). These results were consistent with our
publisheddata (18)concerningincorporationof
p30 into extracellular virus in the absence of pactamycin. Immune precipitation, with
anti-serumtop30andp15,ofsubcellular fractions of
cells pulse-labeled inthe presenceof
pactamy-cinandchased for1hresultedinnodetectable
Pp60 (data not shown). The results of these
controlexperiments indicated thatpactamycin didnotdetectably effect eitherthe cleavage of
Pp6O or the release ofviral polypeptides from
these cells.
Table 1 depicts the results ofa pactamycin
on November 10, 2019 by guest
http://jvi.asm.org/
82 OKASINSKI AND VELICER
TABLE 1. Effect ofpactamycin on therelative labeling of FeLV virionproteinsa Viral protein Relative labeling ratiob
p11 0.44
p15 0.65
plO 1.06
p30 1.21
a Cells werepulse-labeled inthe presence or
ab-sence ofpactamycin and then chased for 20 h. Puri-fied virus wasdisrupted, and proteins were chro-matographed by gel filtration in the presence of 6 M GuHCl.
bRatios are thepercentage oflabel in each pro-tein synthesized in the presence of pactamycin over the percentage of label in each protein synthesized
intheabsence ofpactamycin.
experiment. An examination of this data
indi-catesthat pactamycintreatmentledto a
prefer-entialloss of labelinp1l and p15 relativetoplO and p30. The relative labeling ratios obtained
inthis experimentindicatethefollowing order for FeLV proteins: N-pll-pl5-plO-p30-C.
DISCUSSION
In this communication we have presented evidence that the majornon-glycosylated
struc-tural proteins of FeLVaresynthesized as part
of a large precursor polypeptide. The largest precursordetected in these experimentswas a
70,000-dalton polypeptide, present both
intra-cellularly (Fig. 4) andas acomponentof FeLV (Fig.5,6,and 7).Trypticpeptide analysis dem-onstrated thatPp7O and virion p70were identi-caland that bothcontainedthe trypticpeptides
of FeLV p30, p15, p1l, and p10 (Fig. 6 and 7).
Thepresenceof p1l tryptic peptides in
intracel-lularPp7O and the absence of these tryptic
pep-tides in Pp6O indicate that the conversion of
Pp7O
-*Pp6O
involves the release ofp1l from Pp7O. Wehavealsopresented evidencesuggest-ing
that the conversion ofPp7O
-0Pp6O
canbe inhibited by L-azetidine-2-carboxylic acid (Fig.4), whereas the conversion of
Pp6O
-0 FeLV structuralproteinswaspartiallyinhibitedbyageneralproteaseinhibitor(Fig. 2and 3).
Pulse-labeling experiments in the presence of
pacta-mycin indicated that the order of small FeLV
structural proteinsispll-pl5-plO-p30(Table1).
The presence of a virion-associated protein
(p70) with atryptic peptide profile identicalto
intracellular Pp7O indicates the incorporation
of anuncleavedprecursor into mature virions.
A similar finding has recently been
demon-strated for Rauscher leukemiavirus(RLV) and RLV-infected cells (9) as well asevidence fora
possibleuncleaved precursoras acomponentof
the feline leukemia virus pseudotype of
Molo-ney sarcomavirus (19). Subviral particles
pre-paredfrom FeLV containp70within the core of
FeLV rather than as an envelope component (Behnke and Velicer, manuscript in prepara-tion).
Although immune precipitates (using anti-p30) frompulse-labeled cells routinely contain
low levels of a labeled 70,000-dalton moiety (Fig. 2and3) (18),thispolypeptide becomes the principal immune precipitable polypeptide
when cells aregrowninthe presence of
L-azeti-dine-2-carboxylic acid. Weattribute the routine presence of Pp7O at lowlevels inimmune pre-cipitates during normal pulse-labeling
condi-tions(Fig. 2A andB)and withlong-term
label-ing conditions (18) to reflect an intracellular
level of uncleaved Pp7O that eventually
be-comes incorporated into FeLV as virion p70.
However, a portion ofthis Pp7O may also be destined for furthercleavage. Theproline
ana-logue L-azetidine-2-carboxylic acid could affect cleavage of Pp7O in two ways. The analogue could beincorporated directlyintothe cleavage enzymeand alter theactivity of the enzyme,or the analogue could be incorporated into the
substrate (Pp7O). In our experiment the cells
were preincubated with L-azetidine-2-carbox-ylic acid for45min prior topulse-labeling (Fig.
4). Similarresults areobtained when thecells
arepreincubatedfor 0, 5, and 10 min with the
analogue (datanotshown). Ifthe proline
ana-logue were altering the cleavage enzyme
di-rectly, the enzyme would have to have an
ex-tremely short half-life. Themostlikely
alterna-tive is that L-azetidine-2-carboxylic acid is
in-corporateddirectly into Pp7O and thus prevents
the conversion of
Pp7O
-0Pp6O.
The effect of the general protease inhibitor
uponthe cleavageofPp6O does notprovide an
unequivocal interpretation. We cannot
attri-butethe ratherdramatic changesinthe levels
and distribution ofPp6O (Fig. 3) to a general
effect of PMSF on cell viability because the
decrease in cell viability is rather small.
Al-though PMSF does reduce the level of protein
synthesis, thisreduction appearstobe
quanti-tative rather than selective for viral protein
synthesis. Furthermore, Fig. 3 shows that a
changeinthe totallengthofexposure from 4.5 min (Fig. 3Aand D plus C and F)to34.5 min
(Fig. 3B and E) did notcause any significant
changesintheelectrophoreticprofiles. Figures
2and3show thatapartialinhibitionofPp6O by
PMSF occurs only if PMSF is present during
and shortly after the synthesis ofPp6O. It is
difficult to attribute the inhibition of Pp6O cleavage to asimpleinactivation ofaprotease
duetotheshorttimeframe ofactivityseenin J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
these experiments. Figure 2 shows that Pp60 becomes committedtocleavagewithinthe first few minutes after synthesis, whereas Fig. 3 indicates that PMSF inhibition requiresonlya
shortexposure toPMSFduring pulse-labeling.
BecausePMSFinhibition requires thepresence
of theprotease inhibitor onlyduring
pulse-la-beling, we interpret its activity to an aberra-tionofthe normal juxtaposition ofasubstrate Pp6O and enzyme, possiblyinduced by the in-teraction of PMSF with either Pp6O or a
pro-tease. In support ofour interpretation is the
change in distribution ofPp6O within the PF and CE. Under normal conditions Pp60is lim-ited to the PF, whereas, in the presence of PMSF, Pp6O is foundin both the PF and CE immediately after pulse-labeling. Lindell has recently reported (13) that the addition of PMSFtoisolated nuclei inhibits the release of RNA polymerase, which also indicates that PMSF may have additional activities aside from theproteaseinactivationactivity. Further experimentationinvolvingisolation of intracel-lular membranes and precursor polypeptides
will be required to more clearly elucidate the activity of PMSF on Pp6O cleavage. Previous work with both AMV-infected cells(27,28)and RLV-infected cells (26) has not provided evi-dence forany inhibition ofprecursor cleavage bygeneral proteaseinhibitors. However, RLV
precursor cleavage may be inhibited by
tolyl-sulfonylphenylalanylchloromethylketoneand
carbobenzyloxy-phenylalanyl chloromethyl
ke-tone(R. Arlinghaus, personal communication). Our results with FeLV-infected cells may
re-flect differences in experimental design (i.e.,
concentrations ofprotease
inhibitor)
or indif-fering susceptibilities of avian, murine, and feline oncornavirusprecursor
polypeptides.
Trypticpeptide analysis of intracellular
pre-cursorpolypeptides and virion structural
pro-teins(Fig.6and7) demonstratesthat themajor
structural
non-glycosylated
polypeptides
of FeLVaresynthesizedaspartofa70,000-daltonprecursor. The presence ofp30, p15, p1l, and
plO tryptic peptides within the Pp7O molecule and the absence of the
p1l
trypticpeptides
inPp6O indicate that the conversion of Pp70 --Pp6Oresultsinthe formation ofp1l.These data
wouldsuggest that
Pp7O
containsthep1lmoiety
at either its NH2-terminal or COOH-terminal end andthat removalofp1l results in the
for-mation ofPp6O.
The results ofpactamycin gene ordering
ex-periments(Table 1) confirm thesuggested
posi-tionofp1l withinPp70. These results combined with the trypticpeptide analysisand immune
precipitation experiments lead us to propose
(Pp 70)
p11 p15 plO, p30
,/ ,' " (Pp60)
..1'p1 + p15 p O. p30
(Pp25?) '
p15 plO + p30
(p70) i X
pil p15 plO, p30 ,' p15 + p1O
FIG. 8. Proposed scheme forthegeneration of the non-glycosylated structural proteins of FeLV. The order of FeLV proteins within Pp7O was that ob-tained from pactamycin gene ordering experiments. The existence of Pp25 is uncertain.
the cleavage scheme shown in Fig. 8 for the
generation ofFeLV non-glycosylated structural
proteins. Thefirst step in this scheme involves
thegeneration ofp1l and Pp6Ofrom Pp7O.
Be-cause p1l is the NH2-terminal moiety, this
cleavage may occur on a nascent polypeptide
andthus could explainthedetection of only low
levels of Pp7O undernormallabeling conditions
(Fig. 2A and B). The absence of any anti-p30
immune precipitable intracellular polypeptides
otherthan Pp6O and p30 (Fig. 2) argues that
p30is generatedbydirectcleavage ofPp6O.The
order of FeLVstructural proteins also supports
this interpretationin thatp30is
carboxy-termi-nal onthe Pp6O moiety. The generation ofp30
from Pp6O should result in the formation of a
precursor polypeptide of about 25,000 daltons
(Pp25) containingp15andplO. Immune
precipi-tation of subcellular fractions from pulse-la-beled cells usinganti-pi5under thesame condi-tions used for anti-p30 also yields Pp7O and Pp6O, but not a 25,000-dalton precursor (G.
Okasinski,unpublisheddata). This observation leads us to believe that p15 and plO may be rapidly generated directly from a would-be
Pp25.The finalfeatureof this proposed
genera-tion of structural proteinsisthe incorporation ofPp7O into assembled virusasp70.
A comparison of the arrangement and
pro-posed processing of FeLV structural proteins with that recently reported for AMV reveals
some distinct differences. The order of AMV structural proteins withina 76,000-dalton
pre-cursor(Pr76) isN-p17-p28-p14-C,withthe
posi-tion of plO being uncertain (28). The major group-specific antigen (p28) is located inter-nally within AMV Pr76, whereas the
corre-sponding FeLV protein is COOH-terminal on FeLVPp7O.Vogtet al. (28) have proposed that
the first cleavage ofPr76resultsinprecursors
of66,000 and 12,000 daltons (Pr66 and Prl2),
the latter arising fromcleavageatthe COOH-terminal end and ultimately giving rise to AMVp14. ThusAMVprecursorprocessingmay
differ from that proposed forFeLV in thatthe
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.508.289.432.68.141.2]84 OKASINSKI AND VELICER
first cleavage is atthe COOH-terminalend in
the former andattheNH2-terminalend in the
latter. Avian Pr76, thus, requires complete or
almost complete synthesis prior to the first cleavage, whereas feline Pp7O could be cleaved
as a nascent polypeptide. These differences
could explain the relative ease of avian Pr76
isolation and the absence of abundant feline Pp7Ounder normallabeling conditions. Pr66is
believed to be further processed to a 60,000-dalton precursor (Pr60). At this point thetwo
proposed schemes become very similar. It is believed that AMV p28, plO, and p17are gener-ateddirectly fromAMV Pr6O.This process may be analogous to the generation of FeLV p30,
p15,andplOfrom FeLV Pp6O. Although Vogtet
al. (28) havedetecteda32,000-daltonprecursor (Pr32), it is notbelievedtobeanessential step
inthe generationof AMV proteins. Recent pub-lished work with RLV precursor proteins has also failed to detect precursors of RLV p30
smaller than 65,000 daltons (1,16, 26),although
this point isstill under investigation and there is some unpublished evidence for smaller
pre-cursors(Arlinghaus, personalcommunication).
Thus FeLVappears todiffer from AMVinboth
the arrangement of structural proteins in
pre-cursorpolypeptides and inthe initial cleavage from precursors, but may be similar in the processing of60,000-dalton precursor polypep-tides. The incorporation ofaprecursor
polypep-tide into assembled virions alsoappears tobea
unique phenomenon of FeLV and RLV (9), while absent from AMV.
The identification of precursor polypeptides
inFeLV-infected cells is consistentwithsimilar
reports ofprecursor polypeptides in
AMV-in-fected cells (18, 27) andinRLV-infected cells (9, 16, 26). Precursorpolypeptides similartothose identifiedinAMV-and RLV-infected cells have also been synthesized in cell-free translation systems (6, 10, 17, 29). Inaddition tothe
pres-ence of precursor polypeptides in cells
produc-tively infected with oncornaviruses, these
pre-cursorshaveinevery instancebeenindicatedto
have a membrane association (9, 16, 18, 26-28). Although the synthesis of oncornavirus pro-teins in productively infected cells results in
the formation ofhigh-molecular-weight,
mem-brane-associated precursor polypeptides, the size of detectable precursors and their
subse-quent processing appears to vary. Whenamino
acid analogues have been utilized to inhibit
precursor processing, the results have varied
fromno effect in AMV-infected cells (27)with
fluorophenylalanine andcanavanine to
inhibi-tionofformation ofa65,000-daltonprecursorin
RLV-infected cellswith canavanine andno
ef-fect withfluorophenylalanine and
L-azetidine-2-carboxylicacid (26).Inourhands L-azetidine-2-carboxylic acid resultsinanaccumulationof Pp7O (Fig. 5), whereas canavanine and fluoro-phenylalanine yield no detectable alterations
inelectrophoretic profiles of immune precipita-ble precursor polypeptides (G. Okasinski,
un-published data). Our finding of partial inhibi-tion of Pp6O cleavage by a general protease inhibitor (Fig. 3and 4) is, to ourknowledge, the first suchpublishedreport andserves topoint
out what may be fundamental differences in
various oncornavirus precursor polypeptides
and/orinhost cell influencesonprecursor proc-essing.
The synthesis and processing of RLV
precur-sorpolypeptides as discerned by two different groupsofworkers usingdifferent host cellsmay bestexemplify the effects of host cells on
pre-cursorprocessing (16, 26). One groupof
work-ers, using JLS-V9 cells infected with RLV,
could identifypresumptive precursor polypep-tides with molecular weights of 82,000 and
65,000. These precursor polypeptides are
be-lieved to be precursors of RLV p30, p15, and
pl2b (26). Results ofimmune precipitation of
intracellular protein with anti-RLV indicated that very little, ifany, RLV p30 was present intracellularly and suggested that production
ofp30from precursor polypeptides is
immedi-ately followed by assembly of RLV p30 into extracellular virus. When JLS-V16 cells were examined for the presence of RLV precursor
polypeptides,four large polypeptides were iden-tified, with molecular weights of 200,000, 90,000, 80,000, and 65,000, respectively (1, 16). JLS-V16 cells, however, do contain readily identifiable intracellular RLVp30, suggesting relatively larger pools of intracellular RLVp30.
These differences inthe number of identifiable precursor polypeptides, their size, and their processing rates (as judged by intracellular
RLVp30levels) all argue that due caution must be exercised in developing a general mode of oncornavirusproteinsynthesis and processing.
ACKNOWLEDGMENTS
We thankAlice Swanson for technical assistance. This research was supported by Public Health Service grant CA-12101 from the National Cancer Institute and grant DRG-1230 from the Damon Runyon Memorial Fund forCancerResearch, Inc., and a grant from the Elsa U. Pardee Foundation. L.V. is a recipient of research career development award CA-70808 from the National Cancer Institute.
LITERATURE CITED
1. Arcement, L. J., W. L. Karshin, R. B. Naso, G. Jamjoom, andR. B.Arlinghaus. 1976.Biosynthesis ofRauscher leukemia viralproteins: presence ofp30 and envelope p15 sequences in precursor polypep-tides.Virology69:763-774.
2. Baltimore, D. 1971. Expression of animal virus
ge-nomes.Bacteriol. Rev. 35:235-241.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
3. Butterworth, B. E., L. Hall, C. M. Stoltzfus, and R.R. Rueckert. 1971. Virus-specific proteins synthesized in encephalomyocarditis infected HeLa cells. Proc. Natl. Acad. Sci. U.S.A. 68:3083-3087.
4. Clegg, J. C. S., and S. J. T. Kennedy. 1975. Initiation of synthesis of the structuralprotein of Semliki forest virus.J. Mol. Biol. 97:401-411.
5. Fairbanks, G., T. L. Steck, and D. F. H. Wallach.1971. Electrophoretic analysis of the major polypeptides of the human erythrocyte membrane. Biochemistry 10:2606-2617.
6. Gielkens, A. L. J., D. Van Zaane, H. P. J. Bloemers, and H. Bloemendal. 1976.Synthesis of Rauscher
mu-rine leukemia virus-specific polypeptides in vitro. Proc. Natl. Acad. Sci. U.S.A. 73:356-360.
7. Graves, D. C., and L. F. Velicer. 1974. Propertiesof felineleukemia virus. I.Chromatographic separation andanalysis of the polypeptides. J. Virol. 14:349-365. 8. Jacobson, M. F.,and D. Baltimore. 1968. Polypeptide cleavageinthe formation ofpoliovirus protein.Proc. Natl. Acad. Sci. U.S.A. 61:77-84.
9. JamJoom,G., W. L.Karshin, R.B.Naso,L.J. Arce-ment, and R. B. Arlinghaus. 1975. Proteins of Rauscher murine leukemia virus: resolution of a
70,000 dalton, non-glycosylated polypeptide contain-ing p30peptidesequences.Virology 68:135-145.
10. Kerr, I. M., U. Olshevsky, H. F. Lodish, and D.
Balti-more. 1976. Translation of murine leukemia virus RNA incell-freesystemsfromanimal cells. J.Virol. 18:627-635.
11. Kiehn, E. D., and J. J. Holland. 1970. Synthesisand cleavage of enteroviruspolypeptidesin mammalian cells. J.Virol. 5:358-367.
12. Lachmi, B.,N.Glanville,S.Keranen,and L.
Kaarai-nen. 1975.Tryptic peptide analysisof nonstructural andstructuralprecursorproteinsfrom Semlikiforest virus mutant-infectedcells. J. Virol. 16:1615-1629. 13. Lindell,T.J. 1975. Inhibition ofeukaryotic
DNA-de-pendent RNA polymeraserelease from isolated nuclei
andnucleolibyphenylmethylsulfonylfluoride. Arch.
Biochem.Biophys. 171:268-275.
14. McLean, C., and R. R. Rueckert. 1973. Picornaviral
geneorder: comparison ofarhinovirus with a
car-diovirus. J. Virol. 11:341-344.
15. Morser, M. J., and D. C. Burke. 1974. Cleavage of virus-specific polypeptidesincells infectedwith
Sem-liki forest virus.J. Gen. Virol. 22:395-409. 16. Naso, R. B., L. J.Arcement, and R. B.Arlinghaus.
1975. Biosynthesis of Rauscher leukemia viral
pro-teins. Cell 4:31-36.
17. Naso, R. B., L. J. Arcement, T. Gordon Wood,T. E. Saunders,and R. B. Arlinghaus.1975.Thecell-free translation of Rauscher leukemia virus RNA into highmolecular weight polypeptides. Biochim. Bio-phys. Acta383:195-206.
18. Okasinski, G.F., and L. F. Velicer. 1976. Analysis of intracellularfeline leukemia virus proteins. I. Identi-fication ofa60,000-dalton precursorof feline leuke-mia virusp30.J. Virol. 20:96-106.
19. Oskarsson, M. K., W. G. Robey, C. L. Harris, P. J. Fischinger, D. K. Haapala, and G.F.Vande Woude. 1975.A p60 polypeptideinthe feline leukemiavirus pseudotype of Moloney sarcomavirus with murine
leukemia virus p30 antigenic determinants. Proc. Natl. Acad. Sci. U.S.A.72:2380-2384.
20. Paucha, E., J. Seehafer, andJ.S. Colter.1974. Synthe-sis ofviral-specific polypeptides inMengo virus-in-fected Lcells: evidence for asymmetric translation of theviralgenome.Virology 61:315-326.
21. Scheele, C. M., and E. R. Pfefferkorn. 1970. Virus-specific proteins synthesized in cells infected with RNA+ temperature-sensitive mutantsof Sindbis
vi-rus.J. Virol.5:329-337.
22. Schlesinger, S., and M. J. Schlesinger.1972.Formation ofSindbis virus proteins: identification ofa precursor
foroneof the envelope proteins. J. Virol.10:925-932. 23. Summers, D. F., and J. V. Maizel, Jr.1968. Evidence forlargeprecursorproteins inpoliovirus synthesis. Proc. Natl. Acad. Sci. U.S.A. 59:966-971.
24. Summers, D.F.,and J. V. Maizel, Jr.1971. Determina-tion of thegene sequenceof poliovirus with
pactamy-cin. Proc.Natl. Acad. Sci. U.S.A. 68:2852-2856. 25. Taber, R., D. Rekosh, and D. Baltimore. 1971. Effect of
pactamycin on synthesis of poliovirus proteins: a
method for genetic mapping. J. Virol.8:395-401. 26. Van Zaane, D., A. L. J. Gielkens, M. J. A.
Dekker-Michielsen, and H. P. J. Bloomers.1975. Virus-spe-cific precursor polypeptides in cells infected with
Rauscherleukemia virus.Virology 67:544-552. 27. Vogt, V. M.,and R. Eisenman.1973.Identification ofa
large polypeptide precursor ofavian oncornavirus
proteins. Proc. Natl. Acad. Sci. U.S.A. 70:1734-1738. 28. Vogt, V. M., R. Eisenman, and H. Diggelmann. 1975. Generation ofavian myeloblastosis virus structural proteins by proteolytic cleavage ofa precursor
poly-peptide. J. Mol. Biol. 96:471-493.
29. Von Der Helm,K., and P. H. Duesberg. 1975. Transla-tion of Roussarcomavirus RNAinacell-freesystem from ascites Krebs II cells. Proc. Natl. Acad. Sci.
U.S.A.72:614-618.
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG. 3.2.5PFandthedeficientatext. PF SDS-PAGE ofimmune precipitates from PF and CE ofcells pulse-labeled with [35S]methionine in presence and absence of PMSF](https://thumb-us.123doks.com/thumbv2/123dok_us/1546462.107179/5.508.64.259.475.568/pfandthedeficientatext-ofimmune-precipitates-ofcells-labeled-methionine-presence-absence.webp)