JOURNAL OF VIROLOGY, Jan.1979,p.123-133
0022-538X/79/01-0123/11$02.00/0 Vol. 29, No. 1
Envelope
Proteins and Replication of Vesicular Stomatitis
Virus: In Vivo
Effects of
RNA'
Temperature-Sensitive
Mutations on Viral RNA Synthesis
CLAIRE MARTINET,* ARLETTE COMBARD, CHRISTIANE PRINTZ-ANE, AND PIERRE PRINTZ Institut deMicrobiologie, Universite de Paris-Sud, Centre d'Orsay, 91405 Orsay Cedex, France
Received for publication 19 May 1978
Temperature-sensitive (ts) mutants of vesicular stomatitis virus belonging to
complementation groups III and V were investigated for their in vivo RNA
synthesis. Thesucrose gradient patterns of the RNA species which theyproduced
at nonpermissive temperature
(39.2OC)
were systematically compared underdifferent experimental conditions: variationofinputmultiplicity andoftimeof infection, superinfection withTparticles, and temperatureshifts.Finally,amore
precise analysis of the various RNA species synthesized was carried out. It
appeared that thecharacteristics ofRNA synthesisspecifiedat
39.20C
bytsHI ortsVmutantsdiffered from the normal RNA synthesis of vesicularstomatitisvirus
wildtype.Theircommon depression at
39.20C
invirion-like RNA (38S)produc-tion-i.e.,so-calledgenomereplication-was tentatively paralleled with the
con-comitant ts eventswhich have been previously shown to affect the two viral envelope proteins. Anoverproduction of the RNA transcriptswasdescribed for
mutantsingroupIII andposed the question ofaregulationprocess todetermine
theamountofRNAtobe transcribed.
Inthe processof
elucidating
thevirus-speci-fied events in cells infected with vesicular
sto-matitis virus (VSV), much interest has been centered on temperature-sensitive (ts)mutants
of this virus. Uptonow, information about
mu-tantsclassifiedas
RNA' (able
tomakeareadily
detectable amount of
actinomycin
D-resistant RNA) was almost limited todescription
of the proteinsynthesis they
induced at restrictivetemperature (21,
32).
Indeed,
onemayapriori
assumethattheir RNA
syntheses
would befully
comparable
tothose of the wildtypeand there-fore that these groups are rather altered in a maturationstep late in theviralcycle.
Infact,
defects inenvelope
proteins
Mand G have been demonstrated forgroupsIII andV,
respectively
(4, 9, 11,12,33).Despite the sparseness of
published
results about the RNAsynthesis
inducedby
tsII andt8Vmutants,atleastoneobservation
prompted
ustocharacterize inmore detail their intracel-lularRNAspecies:the
abnormally
highlevel of RNAsynthesis obtainedintsIII-infectedcellsat restrictive temperature, either in cumulativemeasurement (20) orin a sucrose
gradient
pat-tern (29).
Therefore,
acomparative
analysis
ofthe RNA
syntheses
inducedby
tsIII and tsVmutants was systematically
conducted,
and analterationintheirsyntheseswasdemonstrated.
MATERIALS AND METHODS
Cells and viruses. AU the analyses of RNA syntheses in vivowereperformedonHeLacell mono-layers grownasalready described (22) andonchicken embryo cultures whenalow level of residualcellular proteinsynthesiswasrequired.
VSV (Indiana serotype) strainsinvestigated were:
awild type which gives twiceasmanyplaquesat 30 as at39.20C,parentofthe mutants; tsmutantsbelonging
tocomplementationgroupm [ts023(III),
ts076(Il),
and
ts089(Ill)],
andts mutantsbelongingtocomple-mentation group V [tsO45(V), ts044(V), and
tsO110(V)]. Allmutantstocks, containing only
stan-dard Bparticles,werechecked forlow percentage of
revertants and low residualgrowthat39.2°C andby
complementationtestsasreported previously(22).
Defective interferingTparticleswerepreparedby
infecting chicken embryo monolayers with a third
high-multiplicity passage ofwild-type VSV. After a
low-speedcentrifugation toremove cells and debris,
the virions(Band Tparticles)werepelletedat33,000
rpmfor 1 hat40CinaSpinco45 rotor.Thepellets
weresuspended in asaline buffer (0.01 M Tris [pH
7.4], 1 M NaCl, 1 mM EDTA).Thesuspensionwas
layeredonto a 5 to40%sucrosegradientinthesame
buffer andcentrifuged (18,000rpmfor1.5hat4°Cin
a SpincoSW25 rotor). The band of Tparticleswas
removedbysuction, andtheyieldof Tparticlesinthe
preparationwas calculated in equivalent PFU from
optical densityat 260nm (7). The Tparticleswere
checked for homogeneity in size, corresponding to
shortTparticleswith19SRNA.
123
on November 10, 2019 by guest
http://jvi.asm.org/
124 MARTINET ET AL.
Sucrose gradient fractionation of uridine-la-beledproducts. HeLacells wereinfectedatthe mul-tiplicity specified for eachexperiment and treated with actinomycin D. After labeling with [3H]uridine (20
MCi/ml), cytoplasmic
extracts were prepared asal-readydescribed (22). Fromthisstarting materialtwo
differentanalyses could be conducted.(i)RNAspecies
weresolubilized from thecytoplasmic extractby ad-dition ofsodiumdodecyl sulfateto afinal
concentra-tion of1%;theywerefractionatedby centrifugationin a15 to 30% sucrosegradientat21,000rpm for 15 hat
17°Cin aSpinco SW27.1rotor,asdescribedelsewhere (2).(ii) Thecytoplasmicextractwasalsofractionated
on a 15 to30%sucrose gradient inhighsalinebuffer
(0.01 MTris[pH 7.4],0.5 MNaCl,0.05 MMgCl2at 27,000 rpmfor3h at4°C in the Spinco SW27.1rotor.
Determination of the radioactivity in an aliquot of each fractioncollected enabledus tolocate the
poly-somes (positioned onboth sides of the 120S nucleo-capsidpeak) and the free mRNA present as ribonu-cleoprotein structuresmigratingjust above the ribo-somal optical density peak. Fractions were pooled (polysomeson theonehandand free mRNAon the other); the RNAwasphenolextracted(22), and
poly-adenylic acid-containing mRNA's were isolated
ac-cordingtotheiraffinityforoligodeoxythymidylicacid [oligo(dT)]-celluloseasdescribed below.The mRNA's
were analyzed on a 15 to 30% sucrose gradient in
sodiumdodecyl sulfateasusual.
Analyses of viral RNA. After separationon su-crose gradients, the various VSV RNA species were
analyzedin severalways.
(i) Hybridization. Samples of[3H]RNAmeltedat
100°C for2minwerehybridized with unlabeled RNA extracted withphenol frompurifiedvirions.Annealing
was conducted in0.01 M Tris-0.5 M NaCl, pH 7.4
(final volume, 40 d), in the presence of 0.5
Pi
of diethylpyrocarbonate. Annealing was performed at70°C for atleast 24 h. Then the amount of RNase-resistant radioactivity in each annealedsample was
determined aftera30 mindigestion with pancreatic
RNase A(10lg/ml,finalconcentration).
(ii)Melting. Samplesof[3H]RNAwereheated in 90%dimethyl sulfoxide for4min at60°C toseparate anypossiblecomplexedRNA strandsfromeach other
(31). ThenRNAswererecoveredby precipitationwith 2volumes ofethanol in the presenceof0.2 MNaCl
and separation on a 15 to 30% sucrose gradient as
describedabove.
(iii)Bindingto oligo(dT)-cellulose. Samples of [3H]RNAweremixedwitholigo(dT)-cellulose (0.2g) in0.01 MTris-hydrochloride (pH7.4)-0.5 MKCl.The reaction begansimplyas a batch kept for15 min at room temperature; then thesuspension was poured
intoasmallcolumn,andthefirst effluentwaspassed twice more overthe column. The column wasthen washed with about 4 ml of 0.01 M Tris-0.5 M KCl
while a slow flow rate was maintained throughout.
Adsorbed RNAwaselutedwithabout 1 ml of 0.01 M Tris. The measurement of theacid-precipitable
radio-activityofeach fraction madepossibletheselectionof
appropriate fractions, which werepooled and
even-tuallyanalyzedona15 to30%sucrosegradient.
(iv) Methylation. Infectedcells weremaintained
attheappropriate temperature (30 or 39.2°C) for 2 h
inEagleminimal essential medium withonly 10 ttM
methionine butsupplemented with 10jig of
actino-mycin D per ml and 2% fetal calf serum. At 2 h
postinfection (p.i.), cells were labeled with 10 yM
[methyl-3H]methionine (160 .tci/ml)inthe above me-dium without coldmethionine but in thepresence of 10mMsodiumformateand 10AMguanineand ade-nine. Inparallel, cultureswere labeled with
[3H]uri-dine (20,uCi/ml) inthesame mediumadjustedto 10
,M withmethionine.At4 hp.i.,cellswerescrapedoff the sides ofthe flasks and treated withproteinase K
(100lg/ml) for 1 h at 37°C, and RNA was twice extracted with phenol-chloroform and then alcohol
precipitated.
Analysis of viralproteins. Conditions for
infec-tion, labelingofviralproteins in chickenembryo cells, andtreatment of thecytoplasmicextractbefore elec-trophoresis,as well astheelectrophoretic procedure,
wereexactly thesame aspreviously described (2). Chemicals. Additionalcompounds,notmentioned
previously (2, 3, 22),weredimethylsulfoxide, fromE. MerckAG, Darmstadt, Germany;oligo(dT)-cellulose,
from Collaborative Research Inc., Waltham, Mass.;
anddiethylpyrocarbonate, from Hopkins & Williams, Chadwell-Heath, England.Actinomycin Dwas a
gen-erous gift from MerckSharp & Dohme, West Point,
Pa. Radioisotopes were obtained from the
Commuis-sariata l'EnergieAtomique, Saclay,France.
RESULTS
Results reported below refer to mutants
tsO23(III)andts045(V).It mustbepointedout
that within each of these complementation
groupsonlyonephenotype has been constantly
observed, whatever the mutant considered:
tsO23(III), tsO76(III), or tsO89(III) and
ts045(V), ts044(V),ortsO110(V).
Characteristics of RNA synthesis
in-duced at 39.2°C by mutants ofgroups m
and V. (i) Global pattern in sucrose
gra-dient. Auniformlabelingfrom 1 to 4 h p.i. with
[3H]uridinewasappliedtoequivalentbatches of
cells infected in duplicate with tsO23(III) or
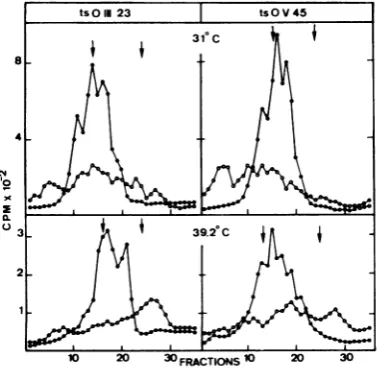
tsO45(V)andmaintained at either 30 or 39.20C.
Cytoplasmic viral RNAs were analyzed by
su-crose velocity gradients. At 39.2°C both
tsO23(III)andtsO45(V) synthesized all the
char-acteristic RNA species normally seen at the
permissive temperature, butwithdifferent
pro-portions (Fig. 1). The levels of synthesis at
39.2°Cof the 13-15 and28S RNAs were approx-imately the same as those at 30°C. Some
in-crease at high temperature could even be
de-tected inthe 13-15S peak. Conversely, the
syn-thesis of the 38S RNA seemed to be depressed
at39.20C.Thesemodificationsarecharacteristic
oftsO(III) and tsO(V) mutants; in the case of
infection withwild-type VSV, the RNA profiles
wereonly slightlymodified with the increase of
temperature (data not shown).
(ii) Effect of input multiplicity on RNA
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ENVELOPE PROTEINS AND REPLICATION OF VSV 125
multiplicity dependent,
andthey
became dis-2 tsOV 45tinctly higher
than thewild-type
ratio above the moderatemultiplicity
values that weusually
used
(50
PFU percell). Looking
to thecorre-sponding
profiles,
we observed that theover-production
here illustrated at39.2°C
wasfully
1 It V1 due to excessive synthesis of the
so-called
mRNA's
(see,
e.g.,Fig.
1),
especially
for thetslI
mutant. In all cases the
38S
peak
was much more moderate at39.2 thanat300C
andneveraccounted for the observed increase of the
39.2/30°C
ratio.However,
the amount of 38Sto
, , , RNArose asthemultiplicity
wasincreased._tsO IU 23 [1
(iii)
Effect of time on RNAsynthesis.
SinceX tsO IN 23 a long labelingperiod represents a steady-state
X. t
\condition,
shorterpulses
were thenperformed.
2 At differenturidine
times
of the infection cycle,[3H]-wasaddedtothe
medium,
and the totalresulting
labeling
in each RNAcategory
wasmeasuredastheareaof each
gradient peak.
Thisalloweda
ready
comparison
of thehourly
rates ofsynthesis
of38, 28,
and 13-15S at 30 and1
39.2°C
by
eachof thetwo mutants(Fig.
3).
Thecurves obtained at
300C
for 13-15Sillustrated
the well-knownpredominance
of the mRNA'sduring
the firststages
of thenormal
cycle
(24,
26).
The 38S RNAemerged
moreslowly
andculminated
ratherlate,
just
beforebeing
released10 20 30 40 as
part
of the virions. Because of theirsimilarity,
FRACTIONS results obtainedat39.2°Cwith eitherts023(Il)
FIG. 1. In vivo RNA synthesis directed by
ts023(III)
andts045(V).HeLacellswereinfected
for 245minatroomtemperaturewith eitherts023(III)or
ts045(V) (100PFUpercell). Thentheyreceived
com-plete Eagle essential medium with 10pgof
actino-mycinD perml,
[H]uridine
(20ACi/mi)
from1 to4hp.i. Cellswerewashed,scraped
off
theflasks, pel-
/leted, anddisruptedinreticulocytestandard
buffer.
Thensodiumdodecylsulfatewasadded
(1%[wt/volJ
z /finalconcentration)tosolubilize
cytoplasmic RNAs,
whichwerethen analyzed byvelocity centrifugation ina15to30%sucrosegradientwith sodiumdodecyl
sulfate at21,000rpmfor15h in theSpincoSW27.1
_-rotor at 17°C. Arrowson thefigure show thepeak
positionsofthe 18and28S ribosomal markers
from
HeLa cells. Symbols:0, 30°C;
0,
39.20C. d20 100 200 40000O iO
synthesis.
The influence of themultiplicity
of MULTIPLICITY OF INECTIO0infectionon anextended
labeling
asabove(1
to FIG. 2. Effect ofmultiplicity
of infection on total 4 hp.i.)
wasinvestigated
for both groups of RNA synthesis by mutantsfrom
groups IIIand V. mutants.By
summing the totalradioactivity
For eachmultiplicity of infectionused,the amountof
within each of the three mainpeaks
in the viral RNA was calculated at either 30or39.2°C
bygradient
profiles,
we could evaluate theratioof summing the total radioactivity of the three main the RNA induced at 39.20Cto that induced at peaks observedin a sucrose gradient profile as in300C and follow its variation versusthe multi-
Fig.
1. The evaluated ratio of counts per minute atplctfinfection (Fig. 2). The ratio for our
39.20C
to countsperminuteat300
Cwasthenplottedplicity
ofsinfein
independeofor
mul versus the multiplicity of infection.Symbols:
0,wild-type
strain,
beingindependent
of the mul-ts023(III);
0,ts045(V).
The broken line correspondstiplicityover a widerange, was taken as aref- to the value observed for a wild-typestrain close to erence
(equal
to=0.6).
Strikingly,
for bothgroup
the strainfrom
which themutants wereisolated andIIIandgroup Vmutantsthe ratioswere
strongly
whichwas notcompletely
heat-resistant.VOL. 29, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.74.215.67.370.2] [image:3.501.255.449.377.541.2]J. VIROL.
126 MARTINET ET AL.
10
'C~~~~~~~~ ~ ~ ~ ~ ~ ~ ~ ~ ~~~~~~~~~~~~~~~~~.
5
,,,,~~~~~~~I,i,L,
[image:4.501.59.248.57.306.2]1 2 3 4 1 2 3 4 1 2 3 4 HOURS p.i
FIG. 3. Effect oftimeonthesynthesis ofeachRNA
species. HeLa cells were infected with either
ts023(III)orts045(V) (multiplicityof infection = 100
PFUper cell).After adsorptionatroomtemperature,
the cells received complete Eagle medium supple-mented with actinomycin D. For each mutant, two seriesof monolayers were used: one was incubated
at 30°C, and the other was incubated at 39.20C. [3H]uridine(20jiCi/ml)wasaddedatdifferenttimes ofinfection, and cytoplasmic extract wasprepared
after 1 h oflabeling. RNAs were solubilized and
layeredontolinear 15to30%osucrosegradients. The
totalradioactivitywascalculatedforeachpeak
ob-served aftergradientfractionationandwasplotted
atthetimevaluecorrespondingtothemeantimeof
the uridinepulse considered. Symbols: 0, 30°C, 0,
39.20C.
or tsO45(V) may be now discussed together.
During the first hours (up to2 to3 hp.i.), the
synthesis of all viral RNAs was impaired at 39.2°Cwithoutaselective inhibitionofanypeak and thus led to a qualitatively normal RNA
profile. Later in the cycle, this normal
equilib-rium between the three main RNA peaks
be-came
progressively
alteredinthefollowingway.First,noincreasedratecould beobserved in 38S
formation: this RNAappearedmerelyata
con-stant restrictedrate. Onthecontrary, mRNA's weresynthesizedwithincreasing speedand
be-came several times more important than they
wereatpermissivetemperature.Onemaynotice
the different slopes of thecurvesof the 13-15S
RNA, which showed again that mutant
tsO23(III) synthesized larger amounts of this
RNA than didmutantts045(V).
Question of a replicative defect in tsHl
and tsV mutants. The
observations
reported
above have
demonstrated
thedeficientamountof38S RNA
produced
at 39.2°Cby
eithertsIII
or tsV mutants. The
possibility
of an actualdefective step in the
replication
process of the viralgenomewill
nowbeexamined.
(i)
Superinfection
withTparticles.
Inter-ferencewith the
production
of viralstandard
Bparticles
ofVSVbyits Tparticles
isthought
tooccur via a
competition
atthelevel oftherep-lication ofthe twotypes ofgenomes (7, 16, 18).
This is
characterized
inviralRNAsynthesis
bytheappearance inthe
gradient
profile
ofapre-dominant
19S
peak,yielding
newT-particle
ge-nomes (26). Such a deviation is shown in the
control
(wild
typeormutantsatpermissive
tem-perature)panel
ofFig.4.Inthisexperiment
thesuperinfection
with Tparticles
wasdone asin-dicated
byStampfer
etal.(26):
Tparticles
were added2.5 haftertheBparticles
at anequivalent
multiplicity
to that of the Bparticles
alreadyadsorbed.
Cells
wereexposed
to[3H]uridine
from 3 to6hp.i.Wedidnotobserve
(Fig.
4) anychange at
all
in the 39.2°C RNAprofiles
of tsO23(III) ortsO45(V),and no inhibition oftheexaggerated
13-15SRNAsynthesis
oraccumu-lation of T genomes could be detected, even when the resolution of the
gradient
wasim-proved
byextendingthe time ofcentrifugation
(bottompanels).
Therefore, atnonpermissive
temperature the
replication
ofTparticles
wasprevented
intsO23(III)-
ortsO45(V)-infected
cells.
Thisresult arguesforanalteration of the replicative process of these mutants. The twofollowing experiments
wereperformed
to findoutwhich
possible
stepcouldbeimpaired.
(ii) Melting of28S RNA material. A firstexplanation
oftherestrictedappearance ofthegenome-like
RNA among the RNAsynthesizedby tsO(III) or tsV mutants was a piling up of
most of the 38S RNA. This might occur by
prevention
of itsleaving thepool ofreplicating
andtranscribing complexes present in the 28S
regionofa
gradient
(25, 26). Therefore, after alabelingfrom 2 to 5 hp.i.,cytoplasmic extracts
from
cells
infected with either tsO23(III) ortsO45(V)werefractionatedinsucrosegradients
asusual. The fractions sedimenting inthe 28S
regionwere
pooled
andseparated
into twoali-quots,oneof which was melted in 90%dimethyl sulfoxide before being ethanol
precipitated,
aswas done directly for the other. Both
precipi-tatedRNAswereanalyzedin 15 to30%sucrose gradients (Fig. 5). As a control, the 28SRNAs
synthesized
at 30°C by each mutant showedaftermelting the sameproportionofradioactive
38S RNA as is
normally
presentinsuchmate-rial. In case of infection at 39.2°C by either
mutant, the melting procedure released
on November 10, 2019 by guest
http://jvi.asm.org/
ENVELOPE PROTEINS AND REPLICATION OF VSV
FRACTIONS
FIG. 4. SucrosegradientpatternsoftheRNA synthesizedintsmutant-infectedcellssuperinfectedwith
VSV Tparticles. HeLa cellswere infectedwith eithertsO23(III), tsO45(V), orwild-type(WT) Bparticles (multiplicity of infection=50PFUpercell)andincubatedat39.2°C. After2.5hp.i.themediumwasdiscarded
andthecells weresuperinfected with defective interfering Tparticlesatanequivalentmultiplicitytothat of
theBparticles; half ofthecultureswereparallely mock infected with saline buffer. Thirty minutes later, the
inoculumwassubstitutedwithmediumcontainingactinomycin D.At 3h afterthefirst infection,[3HJuridine wasadded, andcellswereharvestedat6hp.i.CytoplasmicextractswereprocessedasforFig. 1. In thecase
ofthethree bottompanels,the RNAswereanalyzedin thesame sucrosegradientbutat33,000rpmfor17 h
inaSpincoSW41rotor at17°C. Symbols:0,single infection; *,superinfectionwithTparticles.
8
4
03
2
[image:5.501.105.388.65.306.2]10 20 30FRACTIONS10 20 30
FIG. 5. Analysisofproducts obtained bymelting
28S RNAs. Viral RNAs induced by tsO23(III) or
tsO45(M
and labeled with[3H]uridine from2to 5 hwereanalyzedasindicated inFig.1.After localiza-tionofthe28Speak byprecipitatingaliquots along thegradientwithtrichloroaceticacid, thefractions
tially light RNAs (i.e., <38S) andnot-ascould
beexpected-an increasedamountof 38S RNA.
The significance of this result could be
rein-forced when the RNA complexes recovered in
the 28S region were shown not to be artifacts
arising from a random association during our
wholeexperimentalprocedure. Indeed, addition
of radioactive 38S RNA extracted from VSV
purified virions to cold VSV-infected cellsjust
atthe timewhentheywerescraped into
reticu-locyte standard buffer before beingsubmittedto
theusualextraction-fractionationprocedure did
notlead to the appearance ofradioactivity in
ofthisarea werepooledandseparatedinduplicate:
one wasdirectlyprecipitated byadditionof2volumes
of ethanol, whereas the otherfirstreceived
dimeth-ylsulfoxidetoafinalconcentrationof90% andwas
melted at 60°C for4 min. Both aliquots were
sus-pended in reticulocyte standard buffer-1% sodium
dodecyl sulfateandanalyzedon sucrosegradientsas usual. Cold ribosomalmarkerswereaddedtoeach
gradient. Symbols: 0,original28SRNA; 0,melted 28SRNA.
lSs
Li1
ts0 IN 23
1
tsOV4531C4c
3- i i
~
~~~39.2'C
4 4I
CA
Ie
127
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.55.244.407.593.2]128 MARTINET ET AL.
the28S region ofa 15to30%sucrose gradient.
From this experiment, the hypothesis ofsome
hidden38S RNA within the 28Scomplexes has
toberuledout.
(iii) Polarity of RNAs synthesized at
39.2°C by tsO23(III) and ts045(V). If the
replicative defect demonstrated above forboth
group III and group V mutants consisted in a
deficient 38Sgenome completion, accumulation
ofnegative strands could be expected in other
regions of thegradient. Therefore, the polarity
of the molecules present in each radioactive
peak detectedat39.2°Cwasdetermined by
an-nealing them with unlabeled virion genome,
after melting of the radioactive material. The
comparison (Table 1) of the negative-strand
yields thus obtained with the normalones (i.e.,
in the wild type at 30 or 39.2°C or in each
mutant at 30°C) didnot reveal anyspecial
ac-cumulationat39.2°C ofnegative strands in each
RNAcategoryofthe15 to30%sucrosegradient,
whatever the mutant used. In fact, more 38S
RNA could be annealed toviral genome than
normally when it had been synthesized at
39.2°C. But this was notreally significant,
be-cause-despite the special care taken to avoid
contamination with short nascent RNA
strands-the 38S RNA analyzed presented
about 11%self-annealing.
Effects of temperature shifts. The use of
temperature shifts demonstrated successfully
that the G and M proteins behave abnormally
at 39.2°C during the maturation of tsO23(III)
and ts045(V), respectively (11, 12). Therefore,
similar temperature shifts were imposed on
tsO45(V)- or tsO23(III)-infected cells, and the
viral RNA syntheses were then analyzed. For
bothmutants,wecompared (Fig. 6) the profiles
oftheRNAs labeled from 5 to 6.5h p.i. under
threedifferent conditions: (i)labelingat30°C in
cellsmaintainedatthistemperaturethroughout,
(ii) labeling at 39.2°C in cells which had been
transferred from 30to 39.2°C at 2.5h p.i., and
(iii) labelingat30°C incellswhich hadjustbeen
incubatedbackat30°C afterafirstshift from 30
to39.20C. Attention hastobepaidtothe
differ-ence of the scale used for each mutant.
Com-paredtonormal 300Cprofiles (whichwerevery
similar for both mutants), the simple shiftfrom
30 to 39.20C induced a marked change in the
RNAprofiles of bothmutants,wherethe
alter-ations already described were particularly
sig-nificant: an evident disequilibrium took place
which favored the transcribed RNAsatthe
ex-pense of 38S RNA accumulation (especially
weak). In thecaseofts023(III), the 13-15Speak
couldamount toasmuchas20 times itsnormal
yield, even by 1 hafter the temperature shift.
TABLE 1. Annealingof intracellular viral RNAs with virion RNA'
RNaseresistance(%)b
Virus Type of labeled Afterannealing
RNA
Beoane-
withgenome
Self-annealing
ing RNA
Wildtype at 30 or39.2°Cormutants at 13-15S 7+10 99±8 9±6
300C 28S 12±8 88±10 26±5
38S 2±5 26±7 30±4
ts023(III)at39.20C 13-15S 2±3 100±5 8+ 5
28S 13±5 100±9 21±7
38Sc 11±7 40±2 11±9
tsO45(V)at39.2°C 13-15S 9±2 89±8 10 ± 14
28S 15±3 78±12 22±5
38Sc 7±5 35±7 16±6
Afterextraction and isolationof3H-labeledviral RNAs asdescribedin Fig. 1,materialfrom each peak (at
least1,500cpm) wasmixedwithunlabeledRNAextractedfromVSVvirions.Annealing wasallowedat70°C for atleast24h, andtheproportionofradioactivityrenderedresistant to a30-mindigestionwithpancreaticRNase wasestimatedbyacid precipitation.Theexcess of genome RNA was checked byhybridizationofincreasing levels ofthis one withtotal labeledinfected-cellRNA. The valueobtainedfortsO23(III)-infectedcells at 39.2°C
beingthehighest, it was chosen as abasisto calculate thequantity of genome RNA to be added in each
hybridization sample(=3,ug).
bMean values of several
experiments.
cThe 38Speakwasalwayscontaminatedwith short nascent RNAs
essentially
ofpositivepolarity. In the caseofthepoorly defined38S RNApeak observedduringanalysisof the RNAsynthesizedat 39.2°C by either mutant,thiscontaminationwasespecially inconvenientforsubsequent characterization.Therefore, the fractionscorresponding to the 38S zone in afirst gradientwerepooled, meltedin 90% dimethylsulfoxide, and then
fractionatedonasecond15to30% sucrosegradient.The onlymolecules migratingagain as 38Smaterialwere
annealed.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.501.66.452.382.531.2]ENVELOPE PROTEINS AND REPLICATION OF VSV 129
,31c 39o2c
~~~~~~~~x
31oc 3i92c
oO~ ~ ~5E~S ~~~~2
01 2 3 4 5 6h a.
*dsorption
10 20 30 FRACTIONS 10 20 30
FIG. 6. RNA species synthesized bytsO23(III)- and ts045(V)-infected cells underdifferent temperature
conditions.After infection with ts023(III)orts045(V),actinomycin D-treatedcells wereincubatedat30°C.At
2.5andthenat5hp.i., the cellswereshiftedupto39.2°Cand then downto30°Casindicatedin theleftpanel.
Allsampleswerelabeled with[3H]uridine(20
ACi/ml)
from5to6.5hp.i.Cytoplasmicextractswerepreparedand analyzed as in Fig. 1. Symbols: 0, permissive temperature; 0, upward shift conditions; A, doubk
temperatureshift.
By extending the timne centrifugation of the
gra-dient, the various RNA species inthe 13-15S
peak could be resolvedas three distinct peaks
(i.e., 17, 15, and llS), which had the same in-crease (Fig. 4, bottom panels). Except for this
difference inmagnitudeofRNAproductionafter
the shift, the profiles for both mutants were
rather similar. On thecontrary,thetwo mutants completelydiffered in thecaseofadouble tem-perature shift. In ts023(III)-infected cells,
al-teredRNAsynthesis persisteddespitethereturn
to permissive temperature. Conversely, when
shifted backto
300C,
thetsO45(V)-infected
cellsledtoarapid change towardsa newunbalance
between the three peaks of the RNA profile,
yieldingatremendous amount of38S RNA at theexpenseof the 13-15S RNA. An actual burst in 38S RNA led to the normal level of 38S
accumulation within 1.5 h.
Overproductionof RNAby tsmmutants
at39.20C. The temperatureshiftup justshowed
moredramaticallythe excessivesynthesisof 28
and 13-15S RNAs by tsO23(III) at 39.2°C, as
described aboveonseveral occasions. The
ques-tion arises whether this really concerned the
transcription process,which would be exagger-ated owingto the defect ofgroup m mutants.
First, the possibilitywasruled outthat the in-crease of radioactivity in the 28 and 13-15S
species made at 39.20C was simply due to an increase of the specific radioactivity, as that
could besuspectedwithregardtotheabnornal
effect ofgroupIIImutantsonthepermeability
of chickenembryocellstouridine(5)at39.20C.
Infact, this phenomenonwasnotobserved on
HeLa cells (5; personal observation);
further-more,byextendingthespintime(asforFig. 4),
the viral RNAs in 13-15S peak could be
sepa-rated from the 188 ribosomal RNA, and the
ratio ofcountsperminutetoopticaldensitywas
found to be identical for viral RNA peaks at both30and39.2°C (datanotshown).Therefore, at39.2°C themutant tM023(I) actuallyinduced thesynthesis ofanexcessnumber of13-15S,as
of28S, molecules. They were then examinated
fortheirputativemessengercharacter.
(i) Analyses of28and 13-15S RNAs.One
ofthe characteristics ofVSV mRNA's is their complementaritytothe viriongenome.InTable 1,onecan seethatthe positive polarity of28and
13-158 RNAs induced at 39.20C by ts023(II)
wasconsistent with this criterion. Asasecond
characteristic, the presence of polyadenylated
tails was checked by chromatography on
oligo(dT)-cellulose, andtheamountof polyad-enylic acid-containing RNAs in the 28 and 13-15S material recoveredat39.20C (from 1to
3hp.i.) wascomparedwith thenormalamount
observed at 300C. Virtually no difference in
thesetwoproportionswasrevealed(Fig. 7). One
may conclude from these tworesults that the
overproduction of 28 and 13-15S RNA by ts023(III)at39.20Cwasrelated withtheprocess oftranscription.
(ii) Translation.Toseewhetheranexcessof
proteinsynthesistookplaceconcomitantly,cells
were infected with ts023(llI) under the same
conditionsasinFig. 7,and viralprotein synthesis
was followed by incorporation of [3H]leucine
(Fig. 8). Approximatelythesameamountofeach
viralpolypeptide (exceptforthe Lprotein,but
this is a general feature of all mutants) was
translatedat30 and
39.20C,
despitethe doubleamountof available mRNA observedat
restric-31°c O
avL__ I I .,.^ F
VOL. 29, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.501.50.444.64.212.2]130 MARTINET ET AL.
E0.
39.2°
C20
10
10 20 30
FRACTIONS
FIG. 7. Sucrosegradientanalysis
ofpolyadenyla-tedRNAsynthesizedbyts023(III)at30and39.2°C.
Total cytoplasmic RNAs from cells infected with
tsO23(III)at30 or39.20Cwereeachdivided intotwo
aliquots. One aliquotwas directlyanalyzed by
su-crose gradient centrifugation; the other one was
loadedontoanoligo(dT)-cellulosecolumntoadsorb the RNAcontainingpolyadenylatedtracks.After elu-tion, these RNAswereanalyzedincomparisonwith their counterparts. Symbols: 0,totalRNA; 0, poly-adenylicacid-containingRNA.
tive temperature in this
experiment.
Thisseemsto be correlated with the distribution of the mRNAmoleculeswithinthe
cytoplasm. Indeed,
twopools
of viral mRNA have been demon-strated: (i)polysomal
and(ii)
free mRNA inribonucleoprotein structures
(6).
Therefore,
weprepared
bothfractions fromtsO23(III)-infected
cells as described in Materials and Methods.
From the RNAs extractedfrom both fractions,
mRNA'swere
separated according
totheirbind-ingto
oligo(dT)-cellulose.
Results(Table 2)ob-tained at
300C
showed the normal distributionof RNA
transcripts:
about 20%weretrappedaspolysomes, whereas most ofthe molecules
re-mained free in the
cytoplasm,
inaccordance withHuang et al. (6). This proportion wasnot
pre-served in thecaseof infection withtsO23(III)at
39.2°C:no moremRNAwasfoundaspolysomal
RNA,but theamount of free mRNAwastwice
that observed at
300C.
Asmethylation
of viralmRNAseems tobeanimportantfactor for their
subsequent translation, at least in vitro (1, 28),
we checked the extent of methylation of the
mRNAsynthesizedat39.2°C bytsO23(III) (Fig.
9). Whereas in this experiment the mRNA's
10 20 30
FRACTIONS
40 50
30
20
100
x
0,
30
20
[image:8.501.67.251.50.338.2]10
FIG. 8. Synthesis of virus-specified proteins in cells infected with ts023(III). Monolayers infected with tsO23(III) werecovered with 1/10(vol/vol) di-luted minimal essential medium and incubated at
either 30 or 39.2°C. [3H]leucine (10 uCi/ml) was
added 1.5 h later. Cells were harvested at3 hp.i. Proteinswereextractedfrom the cytoplasmiccontent
of disrupted cellsbyboilingwith10%oacetic acid-0.5 Murea-1% sodiumdodecylsulfate-0.1% 2-mercapto-ethanol. Extracted3H-proteinsweresubjectedto elec-trophoresis in 7.5% neutral sodium dodecyl sulfate-acrylamide gels along with "C-proteins
ex-tracted from purified virions as markers (arrows). The insertshowsthe RNAsyntheses under thesame
conditions at30°C(-- -)or39.20C ( ).
TABLE 2. Locationof mRNA in
tsO23(III)-infected
cellsacpm
Location Temp
Oligo(dT)-(OC) Total cellulose bound
Polysomes 30 19,600 1,400
39.2 12,000 1,300
Free ribonu- 30 74,000 6,900
cleoprotein
39.2 153,000 13,500
aCytoplasmic extracts from tsO23(III)-infected
cellsat 30and39.2°Cwerefractionatedon 15to30%
sucrosegradients inhigh saline buffer(27,000rpm,3
h, 4°C intheSpinco SW27 rotor). Fractions lighter than the 80S ribosomal peak and fractions of the
polysomalarea werepooled separately,and their RNA wasextractedaccordingtothephenol method.Their content in mRNAwas estimated by the amount of RNA boundtooligo(dT)-celluloseasinFig.8.
RNA NS 39'2C
RbA~~~~~~~~NS
'-I
Lc
30'C
tI
! *as ,', , X
oXWoeo o.owO,o, o
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.501.262.458.123.305.2]ENVELOPE PROTEINS AND REPLICATION OF VSV 131
N*0 a
FRACTIONS
FIG. 9. Velocity sedimentation analysis of
meth-ylated viral RNA fromts023(III)-infectedcells. Four
batchesofHeLa ceUswereinfectedwith
ts023aIII)
(=50PFUpercell);twowereincubatedat30°C,and
the otherswereincubatedat39.20Cfor2h inEagle minimalessential medium withonly 10C2mM methi-onine, 2% fetal calfserum,andactinomycinD.Labels
([3H]uridine or[methyl-3H]methionine) were
intro-ducedat2hp.i. inthesamemediumsupplemented with 10mM sodiumformateand 10 l guanineand
adenine. At4hp.i.theRNAsweredeproteinizedwith
proteinase K and then twice extracted with
phenol-chloroform. Theywereanalyzedon15to30%
sucrose gradients as usual. Symbols: 0, 30°C; 0,
39.20C.
made at 39.20C were at least six times more numerousthan thosemade at
300C,
theyieldof themethylgroupstheybore didnotexceedthatobserved at 300C. Moreover, the methylated
mRNA ratio at 39.2 to 300C wds only 0.5,but
this reflected only a difference in methionine
metabolism between thetwotemperatures,asit
could be seen on the figure at the levelof the
tRNA peaks and as we observed for the wild-typeVSV(datanotshown).
DISCUSSION
ts mutants of groups III and V induce at restrictive temperature a large amount of viral RNA, which determines their RNA' phenotype (10). After sucrose gradient analysis, however,
an abnormal profile is observed for the RNA synthesized at39.2°C: for both groups the rela-tive proportion between the three main RNA species, i.e., 38, 28, and 13-15S RNA, ischanged whencompared with what is seen at
30°C.
The patternobtainedresembles thatdescribed under othercircumstances with mutants or even with the wild type (2, 17, 30) andshows also a dimi-nution ofthe yield of thevirion-like RNA, coun-terbalanced by enhancement of the RNAtran-scripts.
Such a result hasgenerally
been ex-plained by the existence of anequilibrium be-tweengenomereplication and transcription (17, 22), which arethought to share the same tem-plate and the same enzyme(s). In the case of tsIIIand tsVmutants, high temperature appar-ently promotesupsetting of the normal equilib-rium. The quantity of the 38S RNA, which is the final replication product, is reduced, more especially as the absence of budding out at 39.20C would have lead to its expected accu-mulation. Furtheranalyses
here reported dem-onstrate that the paucity of the 388 RNA ob-served inagradient profile
cannotbe related toa
trapping
of this RNA species among other viralproducts. Therefore, the synthesis of new genome RNA looks tobereally
affected when infection with tsIIIortsVmutantsis carried outat39.20C. The
inability
ofsuperinfecting
T par-ticles toreplicate
at 39.2°C when mixed withtsHI ortsVmutantssustains the idea that
rep-lication of these mutants isdeficient at restric-tive temperature. Kinetic curves enable us to
explain
moreprecisely
what occurs at 39.20C: there isnot an arrestof thesynthesis
ofthe38S RNAbut,
ratheranabolishment of the normal amplification of thereplication
processby
limi-tation of the number ofsynthesized
molecules. The rate ofsynthesis
seems to berestrictedtoitsinitial valueas
if,
just
after the entry of theparental
virions,
thereplication
roundbegan
nornally
at39.20C
within thecell.Aninterpre-tation of these observations is to suppose two
statesof the defective
polypeptide
ingroup IIIandgroupV mutants
depending
onthetemper-ature at whichtheyhave beensynthesized: the
alteration of the new
polypeptides synthesized
at
39.20C
would lead to a restriction of thesynthesis of new VSV genomes, whereas the
parental proteins
brought
by
virions grownat300C
would operatenormally
in this process. Thislatterassumption
is favoredby
twoobser-vations: for both tsIII and tsV mutants the
VOL. 29, 1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.501.50.236.55.404.2]132 MARTINET ET AL.
amount of38S RNA at39.2°Cisdependenton
inputmultiplicity,and in the caseofgroup V the
parental G protein can be reused in rescue of
UV-irradiated virions (4).
The implication of proteinsalteredin
comple-mentation groups III and V in the process of VSVreplication is apuzzling question. Indeed,
it hasbeen demonstratedthatproteins Gand M
are not functional at39.20C in mutants of groups
V and III, respectively (9, 11, 12). Our results
seem toindicatethatthetwoenvelope proteins
of VSV may also have a role in the
intracyto-plasmic synthesis of viral RNA. Such a
state-ment may appear surprising; however, there is
a good agreement between what is observed
about the synthesisof 38S RNA after different
temperature shifts and thereversibility (orlack
ofit) of thealterationinduced byhigh
temper-ature in polypeptide M or G for tsIII or tsV
mutants.Lafay and co-worker (11, 13) have
dem-onstrated that formutants of group III the M
protein does not recover a functional state at
permissive temperature when it has been
syn-thesizedat39.20C,whereas under the same
con-ditions theGprotein ofgroup Vmutantsrapidly
reversestowards normal behavior (12). In
par-allel, we observed that the 38S RNA did not
recovernormalsynthesis (atleast notfor1.5h)
when tsIII-infected cells were transferredfrom
39.2 to 300C; on the contrary, in ts-V-infected
cells the return to300C
immediately
(in anotherexperiment,notshownhere,we sawthat 30 min wasenough) producestherecuperationof a
nor-mal level of38S RNA synthesis. Thus,we can
draw a
parallel
between the replication of38SRNA and the behavior of M and G proteins.
Little is known about the mechanism of new
genome synthesis, but it wasshown to require
thenucleocapsid. Ourownprevious studies
deal-ing with mutants in groups I, II, and IV (2, 3,
22), tentatively correlated with the three
pro-teins ofthecapsid, haveshown theirimplication
in the replicative process. With the present
knowledge, thereis no reason toassignadirect
roletothe Mand Gproteinsin the same process,
but one may consider that these two envelope
proteins have indirect intervention. Perhaps,
owingtotheirspecificabilitytobe inserted into the cellularmembranes, they would determine some special sites in which the nucleocapsid apparatus would be devoted more to the
pro-duction of genomic RNA than to that of
mRNA's. Thelong cytoplasmic lifetime of the G
protein and its specific insertion in the
mem-brane compartment of the rough endoplasmic
reticulum(8, 9, 23) as soon as it is translated are
factsaprioriconsistent with such an hypothesis.
The second point to be discussed now also
implies that the M protein might have a role
other than the simple construction of the viral
envelope. We have alreadymentioned that re-striction of 38S RNA synthesis favors
produc-tion of the mRNA because of the upsetting of
theequilibriumbetween thereplicatingand the
transcribing nucleocapsids. This phenomenon
may well account for the increase of the 13-15S
peak observed for mutants of group V at39.20C,
butnotfor that for mutants ingroup III because
of thestriking increase of thequantityof 28 and
13-15S RNA species (up to 20times the 300C
yield) observed as soon as tsIII-infected cells are incubated at 39.2°C. However, this increase does not take place when primary transcription is
studied at39.2°C (personal observation).In this
case, onlyparental proteins synthesizedat
per-missivetemperature areimplicated, and,as
dis-cussedabove forreplication of38S RNA, they
might have preserved their expected function.
Theexcessivesynthesisof 28 and 13-15S RNAs
otherwiseobserved at 39.20Cconcerns actually
the process oftranscription, since these RNAs
appear to becomplementarytothe viral genome
and possess polyadenylic acid-terminal
se-quences.However,theycannotbe characterized
as polysomal RNAs: only a fraction of these
RNAs is recovered aspolysomes, leading
never-thelesstothe normalquantityof mRNA bound to ribosomes and to the same quantity of
trans-lated viral polypeptides as those during the
course of an infection at 300C. Whether this
impliesthat the translational machinery of
eu-caryoticcells isfullysaturatedwhen functioning
(assuggested by Laskeyetal.[14]) orthat viral or cellular regulation controls the quantity of
protein synthesiscannotbe presently resolved.
It must be noticed that such regulation could
occur via the degree of methylation of the
mRNA's,whichappeared defectivefor most of
them.Anyway, it is clear thatafterthealteration
of the M protein an exaggeration of the VSV
transcriptionrateappears. This in turn suggests
thatthepotential transcription capacitypresent
inVSV-infectedcells is notfullyexpressed
dur-ingthe courseofanormalcycleand that the M
proteinisimplicatedinsomeway in this
repres-sion. Inrelation to this statement, previous
re-sults may be considered. Inhibition of the in
vitrotranscriptionactivity by envelope proteins
has been shown in avirus related to the
rhab-dovirus class (15). Uncoatednucleocapsid cores
from VSV appear to be more active in the in
vitro polymerase reaction than are complete
VSV virions (27), and Perrault and Kingsbury
specifiedthat aninhibitor of this reaction, which
is not theG protein, is present in the envelope
of VSV (19).These data support our conclusion,
althoughitmightbeconceivable thatregulation
of the transcription activity by the M protein
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ENVELOPE PROTEINS AND REPLICATION OF VSV 133 would not benecessarily observed in thevirion
butwould rather beanintracytoplasmic process. This putative role for the M protein as well as the reason for the lack of translation of the excess of mRNA made at 39.2°C by tsIII mu-tants,iscurrently under investigation.
ACKNOWLEDGMENTS
This work was partially supported by grants from the Centre National dela RechercheScientifique (L.A.136) and theCommissariat a l'Energie Atomique.
We thank C. LaBonnardiere and M. Caboche, Station de Virologie et d'Immunologie, Institut National de la Recherche Agronomique 78650 Thiverval Grignon, France, for helping us in the preparation of themethylation assays. We also thank Annick Friedmann forher excellent technical assistance.
LITERATURE CITED
1. Both,G.W., A. K.Banerjee,and A. J. Shatkin.1975. Methylation-dependenttranslation of viral messenger RNAs in vitro. Proc. Natl. Acad. Sci. U.S.A. 72: 1189-1193.
2. Combard, A.,C.Martinet,C. PrintzAne,A. Fried-man, and P. Printz.1974.Transcriptionand replica-tion of vesicular stomatitus virus: effects of tempera-ture-sensitive mutations incomplementation groupIV. J.Virol. 13:922-930.
3. Combard, A., C. Printz-Ane, C. Martinet, and P. Printz.1977.Temperature-sensitive defect of vesicular stomatitis virus incomplementationgroupII. J. Virol. 21:913-923.
4. Deutsch,V. 1976. Parental Gprotein reincorporation by aVesicular StomatitisVirustemperature-sensitive mu-tant ofcomplementation group V at nonpermissive temperature.Virology69:607-616.
5. Genty, N. 1975. Analysis of uridine incorporation in chickenembryo cells infected byvesicular stomatitis virus and its temperature-sensitive mutants: uridine transport. J.Virol.15:8-15.
6. Huang,A.S.,D.Baltimore,and M.Stampfer.1970. Ribonucleic acidspeciesof Vesicular Stomatitis Virus. III. Multiple complementary messenger RNA mole-cules.Virology42:946-957.
7.Huang, A. S.,and E. K. Manders. 1972. Ribonucleic acidsynthesis of vesicular stomatitis virus. IV. Tran-scriptionbystandard virus in the presence of defective interfering particles.J. Virol. 9:909-916.
8. Knipe,D.M.,H. F.Lodish,and D. Baltimore. 1977. Localization oftwocellular forms of the vesicular sto-matitis viralglycoprotein.J.Virol. 21:1121-1127. 9. Knipe,D.M.,H. F.Lodish,and D. Baltimore. 1977.
Analysis of the defects oftemperature-sensitive mu-tantsofvesicular stomatitis virus: intracellular degra-dation ofspecificviralproteins.J. Virol.21:1140-1148. 10. Lafay, F. 1969.Etudedesmutantsthermosensibles du virus de la Stomatite Vesiculaire: classification de quelquesmutantsd'apris descritkres de fonctionne-ment.C.R.Acad. Sci.268:2365-2388.
11.Lafay,F. 1971. Etude des fonctions du virus de la Sto-matiteVesiculairealtereesparunemutation thermo-sensible: mise en6vidence de la proteine structurale affecteepar lamutation ts 23.J. Gen. Virol.13:449-453. 12.Lafay,F.1974.Envelope proteinsofvesicularstomatitis virus: effectoftemperature-sensitivemutations in com-plementationgroupsIIIand V. J.Virol.14:1220-1228. 13.Lafay, F.,and A.Berkaloff. 1969.Etudedesmutants
thermosensibles du VSV:mutantsde maturation. C.R. Acad.Sci.269:1031-1035.
14. Laskey,R.A.,A. D.Mills,J. B.Gurdon,andG. A. Partington.1977.Proteinsynthesisinoocytesof
Xen-opuslaevis isnotregulated by the supply of messenger RNA. Cell 11:345-351.
15.Marx, P. A., A. Portner, and D. W. Kingsbury. 1974. Sendai virion transcriptase complex: polypeptide com-position and inhibition by virion envelope proteins. J. Virol. 13:107-112.
16.Palma, E. L., S. M. Perlman, and A. S. Huang. 1974. Ribonucleic acid synthesis of Vesicular Stomatitis Vi-rus. VI. Correlation of defective particle RNA synthesis with standard RNA replication. J. Mol. Biol. 86: 127-136.
17.Perlnan, S. M., and A. S. Huang. 1973. RNA synthesis ofvesicular stomatitis virus. V. Interactions between transcription and replication.J.Virol.12:1395-1400. 18. Perrault, J., and J. J. Holland. 1972. Absence of
tran-scriptase activity oftranscription-inhibiting ability in defective particles of Vesicular Stomatitis Virus. Virol-ogy50:159-170.
19. Perrault, J., and D. T. Kingsbury. 1974. Inhibitor of Vesicular Stomatitis Virus transcriptase in purified vir-ions. Nature (London)248:45-47.
20. Pringle, C. R., and L. B.Duncan. 1971. Preliminary physiologicalcharacterization of temperature-sensitive mutants ofvesicular stomatitis virus. J. Virol. 8:56-61. 21. Printz, P., and R. R. Wagner. 1971. Temperature-sen-sitive mutants of vesicular stomatitis virus: synthesis of virus-specific proteins. J. Virol. 7:651-662.
22. Printz-Ane, C., A. Combard, and C. Martinet. 1972. Study of the transcription of the replication of vesicular stomatitis virus by using temperature-sensitive mu-tants.J. Virol. 10:889-895.
23. Rothman,J.E., and H. F. Lodish. 1977. Synchronized transmembrane insertionandglycosylation of a nascent membrane protein. Nature (London) 269:775-780. 24. Schincariol,A.L.,and A. F.Howatson. 1970.
Repli-cation of Vesicular Stomatitis Virus. I. Viral specific RNA andnucleoprotein in infected Lcells. Virology 42: 732-74"
25. Schincariol,A.L., and A. F. Howatson. 1972. Repli-cation of VesicularStomatitisVirus. II.Separationand characterization ofvirus-specific RNA species. Virology 49:766-783.
26. Stampfer, M.,D.Baltimore,and A. S.Huang.1969. Ribonucleic acid synthesis of vesicular stomatitis virus. I.Speciesofribonucleic acid found inChinesehamster ovary cells infected withplaque-forminganddefective particles. J. Virol.4:154-161.
27. Szilagyi,J.F.,and L.Uryvayev. 1973.Isolation of an infectiousribonucleoproteinfrom vesicular stomatitis virus containing an active RNAtranscriptase. J. Virol. 11:279-286.
28. Toneguzzo, F.,and H. P.Ghosh.1976.Characterization andtranslation ofmethylated andunnethylated vesic-ular stomatitis virus mRNA synthesized in vitro by ribonucleoproteinparticles from vesicular stomatitis vi-rusinfected L cells. J. Virol.17:477491.
29. Unger,J.T.,and M. E. Reichmann. 1973.RNA syn-thesis in temperature-sensitive mutants of vesicular stomatitis virus. J. Virol. 12:570-578.
30. Wertz,G.W.,and M.Levine.1973.RNAsynthesis by vesicularstomatitis virus and asmallplaque mutant: effects ofcycloheximide.J.Virol. 12:253-264. 31. Wild,T. F.1971.Replicationof vesicular stomatitis virus:
characterization of the virus-induced RNA. J. Gen. Virol. 13:295-310.
32. Wunner,W.H.,andC. R.Pringle.1972.Protein syn-thesis inBHK21cells infected with vesicular stomatitis virus. I.tsmutantsof the Indianaserotype.Virology 48:104-111.
33. Zavada, J.1972.Pseudotypeofvesicularstomatitisvirus with the coatof murineleukaemia andof avian myelo-blastosis viruses. J.Gen.Virol. 15:183-191.
VOL. 29, 1979

![FIG. 3.species.PFUperts023(III)mentedtheseries[3H]uridineatofafterlayeredtotalservedatthe 30°C, Effect of time on the synthesis ofeach RNAHeLacellswereinfectedwitheither or ts045(V) (multiplicity of infection= 100 cell)](https://thumb-us.123doks.com/thumbv2/123dok_us/1520024.104599/4.501.59.248.57.306/pfuperts-mentedtheseries-uridineatofafterlayeredtotalservedatthe-effect-synthesis-rnahelacellswereinfectedwitheither-multiplicity-infection.webp)



![FIG. 9.proteinasephenol-chloroform.ylatedsucroseonine,minimalducedtheadenine.with(=50batches([3H]uridine Velocity sedimentation analysis of meth- viral RNA from ts023(III)-infected cells](https://thumb-us.123doks.com/thumbv2/123dok_us/1520024.104599/9.501.50.236.55.404/proteinasephenol-chloroform-ylatedsucroseonine-minimalducedtheadenine-velocity-sedimentation-analysis-infected.webp)