JOURNAL OFVIROLOGY, May 1976, p. 672-684 Copyright©1976 AmericanSocietyforMicrobiology
Vol.18, No. 2 Printed inU.SA.
Multiple Structures of Adeno-Associated Virus DNA:
Analysis
of
Terminally
Labeled
Molecules with Endonuclease
RHaeIII
DAVID T. DENHARDT,* SHLOMO EISENBERG, KATALINA BARTOK, AND BARRIE J. CARTER
Department ofBiochemistry, McGill University, Montreal,Quebec, Canada H3G 1Y6,* and Laboratoryof
Experimental Pathology, National Institute of Arthritis, Metabolism andDigestive Diseases,National InstitutesofHealth, Bethesda, Maryland20014
Received forpublication25November1975
Thedouble-stranded form ofadeno-associatedvirus(AAV)DNAhasabout20
sitessensitivetoendonuclease R *HaeIII from Haemophilus aegyptius; the
frag-ments produced fallinto about13 size classes, 8 of which contain single
frag-ments.The location of theHaeIII-producedAAVfragmentsrelativetothethree
EcoRi
fragments was determined. Using revised figures for the molecular weights of the HaeIII cleavage products of+X174 replicative form DNA, wecalculatedthat AAV DNA contains about 4,000 nucleotides. After HaeIII
diges-tionofduplex DNA terminally labeled with32Pusingpolynucleotidekinase, the
majority offragments containing a 5' 32P label were about 40 nucleotides in
length,and fragments of similarsize weregenerated from each end, suggesting that theHae siteclosesttotheendiswithinthe terminal repetition. Two
more-slowly-migrating cleavage products also bore 5' 32p end label. These three
terminally labeled specieswerealsogenerated from single-stranded AAV DNA by digestion with HaeIII, and evidence thatonemayhave anonlinear
("rabbit-ear")structureispresented.Thepredominant5'terminal basewasidentifiedas
thymine for both the plus and minus strands of AAV. Single-stranded AAV molecules could not be efficiently covalently circularized by incubation with
polynucleotideligaseorligaseplus T4 DNA polymerase.
Adeno-associatedvirus(AAV) hasagenome
composed of approximately 4,200 nucleotides (4,000, accordingtoourdata), sufficienttocode for about 150,000 daltons of protein if all the nucleotides serve acoding function (see
refer-ence 2 for a review). However, because only
about 75% of thegenomeistranscribed (8) into
stable RNA, the aggregate amount ofprotein that canbecoded foris approximately 10°
dal-tons. The fact that the largest protein in the
virus has an apparent molecular weight of about 87,000to92,000(22,31)is consistent with the possibility that the AAV genome contains
only one gene and codes for only oneprotein
that is processed to yield the virion polypep-tides.
Forreplicationand for transcription, AAV is
normallydependentonadenovirus functions of
unknown nature (9); presumably, host cell
functionsarealsorequired. Becauseofthis
de-pendence on adenovirus and host functions, AAVshouldprovide auseful tool for investiga-tionof thereplicationofmammalian DNA. It is
likely thatthere are similarities between the
mechanisms of replication of the AAV genome
and the eucaryote chromosome. In particular,
AAV may yield clues as to how the ends of linear moleculesarereplicated; since DNA
po-lymerases are incapable of replicating the
ex-treme 3' ends ofDNA templates, some
provi-sion must be made so that information is not
lostatthese ends (11, 12).
Thegenomeof theAAV virioncomprisesone
single-stranded linear molecule of either plus
or minus polarity, and when the DNA is
ex-tracted from the virions the complementary strands will reassociatetoformaduplex, if
con-ditions permit (26, 30). Koczot etal. (23) and
Berns and Kelly (3) established that the
sin-gle-stranded viral molecules could form circles that could be seen in the electron microscope;
these studies suggested the presence of an
in-verted terminal repetitionofat least1.5%of the
genome. Studies of the properties of
double-stranded AAV DNA led Gerry et al. (16) to
conclude that AAV DNA contained a limited
number ofpermutations (perhaps only two) and aterminal repetition (notinverted)
represent-ing about 1% of thegenome.
It isevidentthat the structure of AAV DNA
is complex, perhaps so that the linear
single-stranded viral molecule can be replicated. We
672
on November 10, 2019 by guest
http://jvi.asm.org/
ADENO-ASSOCIATED VIRUS DNA 673
have investigated aspects of the structure of
AAV DNA using acombinationof end-labeling
with polynucleotide kinase anddigestion with
the restriction enzyme fromHaemophilus
ae-gyptius, endonuclease R-HaeIII (27, 29), that cleaves the sequence 5' GGCC , at its center of
symmetry. Wehave used the terminology
sug-gested by Smith and Nathans (34) except that
wehave shortened it slightly for convenience. Endonuclease R-HaeIII will be referred to as
HaeHI; the fragments produced by HaeIII are designated A through M in this article,ordered
according to their mobility on our acrylamide-agarosegels. Further work is required before it can be concluded that this order also corre-sponds to their molecular weights (36).
Evi-dence is presented that AAV molecules have complementary sequences neareach terminus,
but that the terminal regions themselves, though self-complementary, are not always complementary to each other. This appearsto
lead to the formation of a terminal forked struc-turein aportion of thepopulation.
MATERIALS AND METHODS
Viral DNAs. AAV 2 DNA was prepared from AAV 2virionsproduced and purified as previously described (10). 3H-labeled and 33P-labeled kX174
replicative form (RF) DNA was purified (32) and wasgenerously provided by C. Hours.
Enzymes. The restrictionendonuclease R EcoRl wasobtained from MilesLaboratories. The restric-tion endonuclease R-HaeIII was purified as de-scribed (27), except thatthe nucleic acidswere
re-moved by precipitation with streptomycin sulfate rather than chromatography on Bio-Gel A 0.5 M. The spectrum ofOX fragments generated by this nuclease preparation and the salt concentration at which the enzyme eluted from phosphocellulose in-dicated that the activity was primarily that desig-nated HaeIII (29). Bacterial alkaline phosphatase and T4polynucleotide kinase wereprepared as de-scribed (14a). The Neurospora crassa single-strand-specific exo- and endonucleases were prepared and used as described (1).
Centrifugation. AAV DNAwaspurifiedfrom
en-zyme reactions, when necessary, by velocity sedi-mentation in neutral 5 to 20% sucrose gradients
containing1MNaCl,1mMEDTA,and50mM Tris-hydrochloride (pH 7.5). Samples (100 ,ul)were
lay-ered on 5-mlgradientsandcentrifugedfor4hat50 krpm in the SW50.1 rotor of the Beckman L265B
ultracentrifuge. Fractionswerecollectedthrougha
hole poked in the bottom of the tube, and those fractions containing the DNAwere pooledand di-luted with an equal volume of50 mM Tris-hydro-chloride (pH 8.0), and the DNA was precipitated
with0.1volume of3Msodiumacetate(pH5.5) and2
volumesofisopropanol.Precipitationwasallowedto occurfor12 h or more at -20C, and the DNAwas
collected by centrifugation at 10 krpmin the HB4 rotor of the Sorval RC2 for60 min. The DNA was
dissolved in 100 ,ul of 10 mM Tris-hydrochloride (pH 8).
Labeling of the 5' terminus. The DNA was incu-bated with1U ofbacterial alkalinephosphatase per mlat 65C for15minin30mMTris-hydrochloride (pH 8). EGTA [ethylene-bis-(Q3-aminoethyl ether)N,
N'-tetraacetic acid] was added to 6.5 mM, and the incubationat 65 Cwascontinuedfor another15min (21). Thesolution was then made 1.5 mM in potas-sium phosphate (pH 6.8), 7 mM MgCl2, 14 mM in /-mercaptoethanol, and 60 mM in Tris-hydrochloride (pH 7.4). ATP labeled in the gamma position (17) was added in a 100- to 1,000-fold excess over the molar amount of 5' ends present (0.1 to 5pmol). The specificactivity ranged between 2 x 104 and 2 x 105 counts/min per pmol in the different experiments; the final concentration of ATP was maintained be-tween 5and 10 uM. The amount ofpolynucleotide kinase required to quantitatively label the 5' ends of
acontrol polynucleotide preparation in 60min under theseconditions was determined, and this amount was added at 0 min and again at 30 min.After 1 h at 36C, EDTA was added to terminate the reaction, andthe DNA waspurified by velocitysedimentation
inneutral sucrosegradients. This step resulted in the separation of the linearduplexmolecules, used inmost of this work, from the faster-sedimenting
circular andoligomeric forms (7).
Cleavage with restriction enzymes. Endonucle-ase REcoRl wasused as described (6). Duplex AAV DNA (0.01 to1pmol) wasdigested overnightat 37C in 10mMeach NaCl, MgCl2, mercaptoethanol, and Tris-hydrochloride (pH 8) with sufficientHaeIH to givecompletedigestionin 12h. EDTAwasaddedto 20mM,bromophenol blue was added to 0.004%, and solid sucrose was addedtoabout0.5M.
Electrophoresis. The composite acrylamide-aga-rose gels contained the designated percentage of acrylamide, N, N' methylene-bis-acrylamide at
0.05the concentration ofacrylamide, 0.5%agarose
(Seakem,product of MarineColloids,distributed by
Bauschand Lomb) and E buffer (14). Polymerization wasinitiated with 0.02% ammoniumpersulfate and 0.04% N,N,N',N'-tetramethylethylenediamine. Thegels were prerun for 30 min inE buffer, and then the DNA samples were injected onto the upper surface ofthegels. The gelswere 17 cmlong in glass tubes with an internaldiameter of6 mm.The DNA was subjected to electrophoresis for 12 h at room temperature with a current of 3 mAinducedby a 60-Vpotentialdifference across the electrodes. Under
theseconditions, thebromophenolbluedyemarker
migrated about 11 to12cmintothe17-cmgel. After
completionof theelectrophoresis,thegelswere
frac-tionated into 1- or 2-mm fractions using a Gilson Aliquogel gel fractionator. The fractions were
di-gested with alkaline hydrogenperoxideovernightat 55C andneutralized, and the radioactivitywas de-termined with 10 ml of a Triton X-100-toluene-PPO (2,5-diphenyloxazole) cocktail inan Intertechnique
scintillationspectrometer withappropriatesettings
and corrections forbackgroundand cross-talk where necessary. Allgelsare plotted with the cathodeon
the leftsothat themigration of the DNAfragments
isfrom lefttoright.
Determination of the 5'-mononucleotides. The VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
674 DENHARDT ET AL.
DNA (about 0.1
/ig)
was digested with 100 ,tg of DNase I per ml (Worthington Biochem. Corp., EC 3.1.4.5, code D) for 45 min at 37 C in 5 mM MgSO4, 50 mM Tris-hydrochloride (pH 8). Glycine buffer (pH 9.1) was added to a final concentration of 60 mM, and phosphodiesterase I (Worthington Bio-chem. Corp., EC 3.1.4.1, code VPH, treated [35]toinactivate the 5' phosphatase) was added to 50
jig/
ml. Thedigestion was continued for another 60 min and then frozen. A 10- to15-Aul
sample ofthedi-gestedDNA wasapplied to a washed (distilled
wa-ter, 1 M LiCl, absolute methanol) polyethylene-imine-cellulose thin-layer plate (Macherey-Nagel
Co., Cel 300 PEI) together with20 ugeach of the
fourdeoxynucleotide monophosphates dissolved in
water. The plate wasdeveloped in the first dimen-sion with 1 M LiCl,washed with methanol,and then in the second dimension with 1 M aceticacid-3 M LiCl (9:1) (28). The location of the mononucleotide markers was determined by inspection under UV illumination. The entire plate was scanned for 32P radioactivity by autoradiography, and the amount ofradioactivity in the spots was quantitatedin 10-ml of X-100-toluene-PPO scintillation fluid after
A~~~~~
12 *A'
10
o~~~~~~~~~~~~~~~C
0 D
|g
i
F celution of the mononucleotides with 1 ml of 0.5 M ammoniumbicarbonate.
RESULTS AND DISCUSSION Characterization of the fragments of the AAV duplex produced byHaeIII. 32P-labeled double-stranded AAV DNA was digested,
to-gether with
[3HWOX
RF, with the restriction endonuclease R-HaeIH from H. aegyptius asdescribedinMaterials and Methods. The
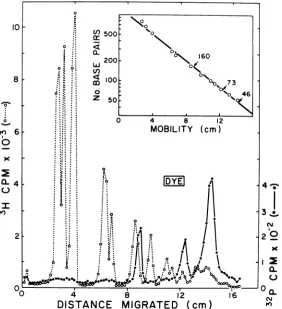
prod-ucts of the reaction wereseparated by electro-phoresis incomposite4%acrylamide-0.5% aga-rose gels, and the distribution of radioactivity throughout the gelwasdetermined by
scintilla-tion countingof thedissolved gel slices. Figure
1shows the distribution of the 32P-labeled AAV
DNA fragments (solid circles) and the
3H-la-beled
OX
RFfragments(opencircles)inthegel.The 4X RF [3H]DNAfragments were used as
molecular-weight standardssince their
molecu-lar weightshave been determined (24); 11
OX
0 2 4 6 8
lo
12
14 16DISTANCE
MIGRATED
(cm)
FIG. 1. Acrylamide-agarose gelelectrophoresis pattern of theHaeIIIcleavageproducts ofthe AAV duplex. AAV duplex [32P]DNA and 4X174 RF [3H]DNA were digested together with HaeIII and subjected to electrophoresis in a 4%acrylamide-0.5% agarose gel as described in Materials and Methods. Migration is from left to right;1-mm fractions were collected. Symbols: (0) 3H-labeled
OX1
74 RF cleavageproducts, (0)32P-labeled AAVduplex cleavage products. Inset: Calibration curve of the size of the 3H-labeled
OX
RFcleavage fragments (-) against the square root of the mobility. The size of the(X genome was taken as 5,500 base pairs (33).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.77.459.302.587.2]ADENO-ASSOCIATED VIRUS DNA 675
cleavage fragments were produced, two of
which had very similar mobilities and were
found inpeakno. 6at5.4cm.
The fragments produced from the AAV
du-plex fell into the 13 major mobility classes,
identifiedbythe letters Athrough M in order of
increasingmobilityasshown in Fig. 1.As
illus-trated in the inset to Fig. 1, an approximate
linearrelation exists between thelogarithmof
the size (e.g., the number of base pairs) of the
OX
markersand thesquarerootofthedistanceeach fragment has migrated into the gel. This
allows anapparentmolecular weightto be
as-signedto each of the fragments A through M,
andthesearegiven in Table1;thesemolecular
weightswerecalculatedassumingthat the
cor-rect size of the
OX174
genome is 4,800nucleo-tides (la).
As a first step towards locating these
frag-mentsonthe AAVgenome, theHaeIII
restric-tion enzyme pattern of the three AAV
frag-mentsproduced by digestion withendonuclease
R-EcoR1 was determined. This nuclease
cleaves the AAVduplexinto three fragments:
EcoR1 A,EcoRl1B, and EcoRlC, with
molecu-larweights of 1.6x 106, 1.1x 106, and 1.3x 105,
respectively (6).EcoRl A is theright-hand
ter-minus,EcoRliB is theleft-hand terminus, and
EcoRliC originates from themiddle of the
ge-nome (7); theycanbeseparated by
sedimenta-tion velocitycentrifugation on neutralsucrose
gradients. The HaeIII digestionpattern of the
intact duplex and the two largest fragments
producedbyEcoRi areillustrated in Fig. 2.
OX
RF DNA was added before the digestion with
R-HaeIIItoprovidemolecular-weightmarkers,
and thedigestsweresubjectedto
electrophore-sis on composite 3% acrylamide-0.5% agarose
gels. The three panels show, from top to
bot-tom,thedistribution ofradioactivityinthegels
of the fragmentsresulting from intact duplex,
the EcoRlDA segment, and the EcoRl B
seg-ment.The resolution in these gels isnotasgood
asinFig. 1because the acrylamide
concentra-tion was lower, and 2-mm rather than 1-mm
fractionswerecollected;nevertheless,the
simi-larity of the two patterns obtained from the
intact duplex (32P labeled) in Fig. 1 and the
intact duplex [radioactively labeled with
[3H]thymidine and density-labeled with
5-bromouraciltoabout 90%substitution]inFig.2
isapparent. Thetwopeaks labeled TA andTBin
the lower two panels contain the fragments
having as one terminus the end produced by
EcoRl. (Additionaldata in support ofthis
as-signmentare presentedbelow inFig. 5and6.)
The molecular weights of the various
frag-ments were determined from a calibration curve established using the known molecular
TABLE 1. Characteristicsofthecleavage productsof duplex AA V DNA produced by endonuclease
R HaeIII
HaeIII No. ofbase No. offrag- EcoRl
assign-sizeclass pairs" ments" ment
A 700 (640) 1
B 584 (550) 1 A
C 454(445) 2 A, B
D
253
(280) 1 AF 210(214) 1 A
G 148(140) 2 A, A
H 113 (105) 2 A, B
I 92(83) 1 B
J 83(76) 1 B
K 71 (63) 1 '/2(?) B ('/2A?)
L 59 (51) 3 A, B, B
M 51(41) 4 A, A,B, B
TA 257 1
TB 183 1
The first number was determined from the square root relationship plotted in Fig. 1 (normal
duplex, 32P labeled); the numbers in parentheses
were derived using the linear relations shown in
Fig. 3 [bromouracil-substituted duplex, [3H]thy-mine-labeled]. The number of base pairs is calcu-lated assuming
4X174
RFhas 4,800 base pairs (la). bDependsontheassumption thatmigrationrateisproportionaltothemolecularweight only.
weights of theHaeIIIfragmentsof AXRF
pres-ent inthegel.Inthiscase, asillustratedinFig.
3, agood linear relationwasobtainedbetween thelogarithm of the number of base pairsinthe fragment and themobility; whenplottedversus
the square rootof themobility, the fitwasnot
as good. We do not have any explanation for
why aplotagainstthesquare root in one case
(Fig. 1, 4% acrylamide) but not in the other
(Fig. 2, 3% acrylamide) allowsabetterfit,but Williamson (37) reported a similar
phenome-non. It may be a function of the acrylamide
concentration. The molecular weights esti-mated from the plots aretabulated inTable 1. The number offragments derived from one
genomeineachpeakcanbecalculatedfroman
analysis of the total radioactivity present in
each band; ifonly onefragment is present in
eachband, then theamountofradioactivityin
thatbandshouldbe proportionalto the size of
thefragment. The assumption implicit inthis
analysisisthat themobility of thefragmentis
proportionalto the sizeofthefragment andis
not affected by the base composition or
se-quence; this may be a poor assumption,
espe-ciallyforthe smallerfragments. Ananalysisof
the twogelsoftheintact AAVduplexshownin
Fig. 1and2ispresentedinFig. 4. Forthedata
in Fig. 1, a better linear relation is obtained
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
676 DENHARDT ET AL. J. VIROL.
IntactA B C (D,E)F G H IUJ K L M
AAVsst99t999ttt
:3 4 5 6 7 8 9 10
§X
markers0:
o . ... .
n,
EcoRlA B C TALT
G H t10
F
s
3 4 56t
7 8 9 10 IX markers5
LL * o
0
44
7 8 10IXmarkers
0
z
W5
DISTANCEMIRTD(c)
0
4
8
~~~2
1
6
FIG. 2. Acrylamide-agarosegelelectropherogramoftheHaeIIIcleavageproducts ofthe AAVduplexand theEcoR1 Aand EcoRl-B fragments. TheAAV DNApreparations were labeled with[3H]thymidine and substituted with 5-bromodeoxyuridine. 32P-labeled OX RF was digestedand subjected to electrophoresis
simultaneouslywith theAAVDNAina3%acrylamide-0.5%agarosegel.Thegelwasfractionatedinto2-mm
portions. A toMand3 to10indicate the distinctivecleavageproductsoftheAAVduplexandOX174RF
fragments, respectively. TA and TBaretheHaeIIIcleavageproductshavingone
EcoRi-produced
terminus.Symbols:(O) 3H,(0)32P.
11 1. 11
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.503.116.403.64.564.2]ADENO-ASSOCIATED VIRUS DNA 677
when thelogarithm of thetotalradioactivityin
theband is plotted versus the squarerootof the distance migrated, whereas for the data of Fig.
2 a better fitisobtained when the mobility is used directly.Fromtheseplots,aswellas simi-lar plotsof thedatainpanelsBand C of Fig.2
(datanotshown), itappearsthatfragments A,
B,D, E,F, I, and J are unique; onlyone frag-mentineach mobility class originates from the
AAVduplex. There are two copies each of frag-ments C and H, one each derived from the
EcoRlFA and EcoRl1B segments. Two
frag500
-00QF
.-e)
< 500
Lii
on
m
cr 100
m500
z
NO A
\XB
c
Intact
0cC
D, E
00F
"o G
B
Eco
RI.A
keKL
TA
,E
e
oF o G\ H
'o
cEco
RI B
><M
bTo
°<< K
, .~
100l
0 4 8 12
MOBILITY
(cm)
FIG. 3. Sizeofthe HaeIIIcleavageproducts.From thedatapresentedinFig. 2,calibrationcurveswere
determined byplottingthe numberofbasepairs in the Xl74fragments (assumingatotalof5,500base
pairs) againstthe distanceeachfragmentmigrated.
Fromtheposition ofeach AAVcleavageproduct,the apparentnumberofbasepairsin thatfragmentwas
estimated.
(MOBILITY,
mm)/2
(o .o)a
z
m
z
02
a.
4 8 12 16
(MOBILITY,
cm)
(°- )FIG. 4. Numberof fragmentsfromonegenomein
eachmobility classofAAVHaeIII cleavageproducts.
The upper curve(0) isaplot of theradioactivity in
eachpeak against the squarerootofthemobility;the dataarefromFig.1. The lowercurve(L)isaplot of
the radioactivity in eachpeak against the mobility; the dataarefrom Fig.2.PeaksC, G, and H(andD +
E inthe lowercurvewheretheywere notresolved)are
replotted after division by 2; Lisreplottedafter
divi-sionby 3; and Misreplotted afterdivision by4.
mentsof the mobility class G are presentinthe
duplex, and both are found in the EcoRl-A segment. The two smallest fragments, L and M, are difficult to quantitate accurately
be-causeof the broadness of the bands and evident
heterogeneity. The best fit to all the gels is obtained, however, when it is assumed that thereare threefragments inL and fourinM, distributed as indicated in Table 1. Figure 1
also indicates that there may be onefragment running fasterthanM.Theonly fragment that is consistently ambiguous is K; possibly it is
migrating anomalously because ofan unusual
base composition orpossibly inthepopulation
ofAAVduplexesanonintegralnumber of
frag-mentsof thatsize areproducedbecause of
het-erogeneity (note that the terminally labeled component beta [see below] migrates in this position) in the population. We cannot distin-guish these alternatives atpresent.
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.254.450.69.334.2] [image:6.503.53.244.206.570.2]678 DENHARDT ET AL.
Identification of the terminalfragments
us-ing polynucleotide kinase. The HaeIII
frag-mentsofAAV DNA possessingthe termini of
theintactmoleculewereidentifiedby labeling
those termini with 32P prior to the digestion
withHaeIII. The DNA wasfirst dephosphoryl-ated with alkaline phosphatase and then
re-phosphorylated using polynucleotide kinase
and [Ly-32P]ATP. The terminally labeled
mole-culesweredigested withHaellI,and the
result-ingfragmentswereseparated by electrophore-sis in the composite acrylamide-agarose gels.
The distribution ofradioactivityinthese gelsis
shown in Fig. 5. Most of the 32P-labeled
frag-ments derived from the intact molecules
mi-gratedonthe leadingedge of the peak
contain-ingthe fragments in the sizeclassM.
Invaria-bly asecond peak containing 10 to 25% of the
32Pwasobservedto migrate justahead of
frag-ment G. Often there wasalsoathirdpeak,very
closeto K, thatcontained5 to 10% of the total
a. 0161
Io
x
CL,
32P. In most gels, at least 70% ofthe total32P
labelrecoveredwasfoundinthetwopeaksnear
G andM. Theapparentmolecularweights ofa
duplex that would migrate at these positions
weredetermined using the AAV fragments as
standards;asshownintheinset(Fig. 5),aplot oflog molecularweightversusmobilitygave a
reasonablystraight line. Agel patternsimilar
(3Hdistributionapparently identical; 32P
pres-ent inall three peaks in theabove-mentioned
proportions) to that shown in Fig. 5 was
ob-tained from adigest of the circular and
oligo-meric DNA sedimenting ahead of the duplex
DNA(equivalenttofractions3 to 8ofFig. 5a in
reference 7). This suggests that the minor32P peaks are not derived from unusual, rapidly sedimentingmolecules and that the oligomeric molecules haveasimilarsetofterminal
config-urations.
Thefact that between 60 and80% of the:32P from the intact duplex migrated in an
appar-4 8 12
DISTANCE
MIGRATED
(cm)
4 .;a
3
1
N
2 -x
I 2
0a.
N
0aL,
cV
FIG. 5. Acrylamide-agarose gel electropherogram of the cleavage products produced by digestion with HaeIII of terminally labeled AAV duplex DNA. The3H-labeled AAV duplex was labeled with 32P at the
5'-terminiusingpolynucleotide kinase, digested withHaeIII,andsubjected to electrophoresis on 4%
acrylamide-0.5%agarose gels asdescribed in Materials and Methods. Symbols: (0) 3H, (0) 32P. Inset: Plot of the size of each AAVfragment versus its mobility. The arrows indicate the position and apparent size of the32P-labeled
fragments.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.122.404.294.603.2]ADENO-ASSOCIATED VIRUS DNA 679
ently homogeneous peak suggested that the
two HaeHI terminal fragments were of the samesize, and thus within the terminal
repeti-tion. This was confirmed by examining the digestion patterns of terminally labeled
EcoRlA andEcoRlFBfragments.The data are
presented in Fig. 6; both EcoRl fragments
yielded 32P-labeled Hae fragments migrating
with theMmobility class,as well as thesecond
component migrating ahead of G. An
asymme-try in the labeling ofthe two ends is
appar-ent.TheterminiproducedbyEcoRl, labeled TA
and TB in the two panels, were labeled about
75% more efficiently than the "natural" end.
This could be forconformational reasons, such
as the presence of the protruding 5' end
pro-duced by EcoRl (19) or because the terminal A
is labeled more efficiently than the terminal
bromouracil(seebelowand reference25). Com-parison of the distribution of32P in these two
panels also reveals that each fragmentwas
con-taminated with about 10% of the other
frag-ment.
Location of fragmentA inthegenome.The
°B'
36
OC
EcoRI A
30
TA
24
° 18
RE
1I2 "FDY
1GM
....
...
L12V ISF
2size of the EcoRlC fragmentwas obtained by subjecting it to electrophoresis together with
OX
RFmarkers. A value of 183 to 192 base pairs(assuming 4,800 nucleotides in
OX
DNA) wasobtaineddependingonwhether thedatainFig.
7 wereplottedusingthe mobility orthe square
rootof the mobility (the datafitequallywell). AftertheEcoRl* C fragment was exposed tothe HaellI nuclease, no change in mobility was
detected, suggestingthat therewere noHaeIII sites withinthatfragment; however, asitevery
close to an end would nothave been detected. When the number ofbasepairs inEcoRlC is addedtothe number of basepairs in HaeIII-TA and HaeIII-TB (Table 1), an aggregate size of
627 basepairs isobtained. This isclose to the
sizeof the fragmentHaeHI -A thatwas not
pres-ent in either EcoRlA orEcoRlB. From this
resultandthe data given inTable 1wededuce
thearrangementoffragmentsindicatedinFig.
8a.
Digestion ofsingle-strandedAAV DNA with
HaeIII. The ability of single-stranded AAV DNA to form circular molecules (3) canbe
ex-16
ca
12
x
0-8 0
a-N
fn
12 16 4 8
DISTANCE MIGRATED (cm)
FIG. 6. Acrylamide-agarose gel electropherogram of the cleavage products produced by digestion with HaeIIIof terminally labeledEcoRi AandEcoRi*Bfragments. These fragmentswerepurified fromanEcoRi digestof[3H]thymidine-labeled, bromouracil-substitutedAAVduplex. TheEcoRi cleavagefragmentswere
labeled with 32Pusingpolynucleotide kinase, digestedwithHaeIII,andsubjectedtoelectrophoresis on4%
acrylamide-0.5% agarose gelsasdescribedinMaterialsand Methods.Symbols:(0) 3H, (0)32P. Thepeaks arelabeledasdescribedinFig.2.
VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.53.445.324.605.2].103 plaind
by
theformation ofstructureslike thosel0 z drawn in Fig. 8b. If such structures areformed
and if
HaelIl-sensitive
sitesare presentin theSWS. EcoRIC duplex portions, then terminally labeled
se-quencesshould be cleaved fromsingle-stranded
8~~~
.z,,,DNAby
the restriction enzyme and should.
(BLTY"246 have the same mobility as fragments derived1.(MOBILITY)'2 from the
duplex. Any
single-stranded
DNA6.1 fragments derived by cleavage of the
single-5..6t9markersed DNArand byHaelIIwould be expectedto
x migratedifferentlyinthesegels. To testthese
ideas, theexperimentshown inFig. 9was
per-Ilb
4.
v formed.a. In this experiment, 32P-end-labeled
single-N.i
@315'
stranded AAV DNA (the minus strand) was2
l
ill°~~~~~~~
digested
with HaeIIIandsubjected
toelectro-phoresisongels usingthesameconditions
em-. \ ! |0 t 5 f ployed for the duplex DNA. As can be seen
M
!ikt\
li ..;M6
_a(Fig. 9),
essentiallyall of the 32Pwasfoundin4 the same three peaks as were obtained when
DISTANCE MIGRATED (cm) end-labeled duplex DNA was digested. If the
FIG. 7. Size of the EcoRl C fragment of AAV DNA. The3H-labeled EcoRl -C cleavage productwas
subjectedto electrophoresis in 4% acrylamide-0.5%
15-agarosegelsasdescribedin Materials andMethods.
The HaeIII cleavage products of 32P-labeledOXRF
were includedas markers and the datawereplotted
assuming a genome size of 5,500 nucleotides for 0
OX174. 4
FIG. 8. (a) Partialordering of the cleavage prod-uctsofAA VduplexDNA produced byHaeIII. The conventional representation of the AAVgenome is shownatthe top, and underneath the threeEcoRi
cleavage productsaredrawnapproximatelytoscale.
Thenumberof basepairs(bp) iscalculatedfromthe
sumofthe HaeIIIfragmentsasgiven inTable1.The
location and relative sizes of A and the terminal
fragmentsTA, TB,and M*aregiven. M*appearsto
migrate slightly ahead ofM (Fig. 5). (b) Suggested structuresfor circularsingle-stranded and linear
du-plexAAVDNA.Forreasonsoutlined in thetext,the
single-stranded molecules are believed to form the
sameterminalduplexstructuresasthe linear duplex
bybase-pairing of thetermini.Proposed locations of HaeIII cleavage sites are indicated byarrows. The
variousstructuresarenotdrawntoscale althougha
is themost-slowly-migratingcomponentandyisthe
fastest.
0
0o
0
N
.
X-Io
0 4 8 12 16
[image:9.503.64.254.46.278.2]DISTANCE MIGRATED (cm)
FIG. 9. Acrylamide-agarose gel electropherogram ofthe HaeIIIcleavage productsofsingle-stranded 3H-labeled AAV DNA (the minus strand) terminally
labeled with 32P using polynucleotide kinase. The
DNA was digestedandsubjected toelectrophoresis
on4%acrylamide-0.5%agarosegels usingthesame
conditionsasused inFig.5and 6. The insetshows the electropherogram ofthe DNA before digestion
withHaeIII.Symbols: (0) 3H, (0)32P.
(0)
5'(T) + strand (light)
- strand (heavy) 5'(T) EcoRI B,-1450bp EcoRI-C,-220bp EcoRIA,-2270bp
ML ,K, A
M J, I, H, D, C,) (B, C, E, F, G, G, H, L, M,) M*
Te TA
(b)
Q='-' <9
680 DENHARDT ET AL. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.503.270.458.295.566.2] [image:9.503.63.257.373.478.2]ADENO-ASSOCIATED VIRUS DNA 681 DNA was not digested with the restriction
en-donuclease, no radioactivity entered the gel
(Fig. 9, inset), thus providing strong evidence that the three peaks seen afterHae digestion were products of Hae action. We infer from these results that the same three types of
frag-ments are generated by HaeIII digestion of the
single-stranded DNAas aregenerated from the duplex. Several discrete peaks of[3H]DNAare
also evident, suggesting that single-stranded
AAVDNA, like fl DNA (19) and
OX
DNA (5),can be cleaved at specific locations by HaeHI. Since underourconditions the single-stranded DNA fragments migrated differently from the duplex fragments, we cannot with certainty de-terminewhich peaks correspond. A gel pattern similartothatseen in Fig. 9 wasalso obtained when terminally labeled plus-strand DNA was digested.
We propose that the three terminally labeled structures(alpha, beta, and gamma,inorder of
increasing mobility) seen in the gels shown in
Fig. 5, 6, and 9 have the configurations alpha, beta, and gamma drawn in Fig. 8b. Hae sites located as indicated by the arrows would
gener-ate fragments of the various sizes found. We
think the real situation is still more complex, butintheabsence of more informationfurther speculation is not useful. To determine if there was any single-stranded DNA in the terminal regions, the DNA was exposed to the single-strand-specific endonuclease from N. crassa,
either before orafterdigestion with HaeIII. In
both experiments (datanotshown), theamount
of32P inall threepositions wasreducedtoabout
thesame extent.ThelossOf32P ismostly,ifnot
entirely, the result of the fact that the nuclease will attack duplex DNA slowly from the ends, presumably asthe result of"fraying." We
con-clude that atleast by this criteriontherewere
no single-stranded regionsinthe terminal
frag-ments.
There areanumber of experiments thatcan
bedonetocheckpredictions basedonthe hypo-thetical structures illustratedinFig. 8b.One of these is that themobility of alphashouldvary
with the pore sizeofagel indifferentfashion relative to beta, gamma, and normal linear duplex fragments. Because duplex DNA
frag-ments migratethrough the gel in an"end-on"
fashion (13) the apparent molecular weight of
alpha, relative to linear duplex markers,
should increase as the acrylamide
concentra-tion isincreased,since it willbe morestrongly
retardedbyvirtueof itsnonlinearstructure.To
test thisprediction,a seriesofgels ofdifferent
concentrations wererun,andtheapparent
mo-lecular weightsof the threeterminally labeled fragments were estimated relative to the
known AAV fragments. The data are
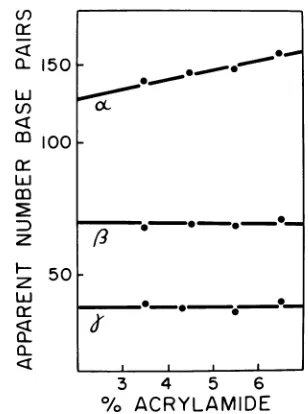
summa-rized in Fig. 10; in the 3.5% gel, alpha ran ahead of fragment
G,
whereas in the 6.5% gel it ran behind G. The other twofragments did not appear to change their mobility signifi-cantly. We therefore argue that a portion of bothsingle-stranded and double-stranded AAV molecules have the "rabbit-ear" structure showninFig. 8b.Wededuce from these results that individual single-stranded AAV DNA molecules exist with two different terminal sequences, each of which is self-complementary and capable of forming "fold-back" structures, but which are
not complementary to each other. In other words, a portion of the population does not have
acomplete inverted terminal repetition. If the terminal sequences were perfectly
complemen-tary to each other they would be expected to
form the more stable single duplex (like
gamma) rather than the double "fold-back"
structure (alpha). It appears that about
one-quarterof the moleculesinthe population con-tain non-complementary terminal sequences, although this could be an underestimate if the kinase labeling was not 100% effective. These results are not incompatible with the
conclu-(I)
at
z
LbJ
(f)
cr-0li 0m
150 60 0.0-0" *~000
CC~0~
1001-. * . *_
50-ci-O
okI I , I
3 4 5 6
%
ACRYLAMIDE
FIG. 10. Apparent size of the terminally labeled fragmentsas afunction ofacrylamideconcentration.
3H-labeledAAVduplexDNAwas terminallylabeled
with32P,digested withHaeIII,andsubjectedto elec-trophoresisongels ofdifferentacrylamide
concentra-tions, butallwith0.5%agarose. Calibrationcurves were establishedfrom the positions of the 3Hpeaks (assuming an AAV genome size of 4,000 nucleo-tides), and the apparentsizeof the 32P-labeled
frag-ments wasthenestimated. a, 8, and yarebelievedto
have thestructuresillustrated inFig. 8b. VOL.18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.503.273.427.349.556.2]682 DENHARDT ET AL.
sions(2,4, 16)that there are two or more types
of AAV molecules that differbyhaving a
lim-itedterminal permutation. However, itshould benoted that for technical reasonsithasnot so
farbeen possibletocloneAAV.
Identification of the5'terminal nucleotide.
Inthe initial attempt toidentify the5' endsof
AAV DNA, the plus and minus strands,
sepa-rated by centrifugation in CsCl equilibrium density gradients after substitution of the thy-mine with5-bromouracil, were individually
in-vestigated. The DNA was labeled with
[y-32P]ATP usingpolynucleotide kinase and then digestedwithDNaseIandvenom phosphodies-terase. Theresulting 5' mononucleotides were
resolved by two-dimensional thin-layer
chro-matography on polyethyleneimine-cellulose plates, and the32Pwaslocatedontheplates by
autoradiography. Both strands yieldedsimilar
autoradiograms. The spots from the minus-strandchromatogramwereexcised,andthe
ra-dioactivity was determined. As can be seen
from the data inTable 2, mostof the 32P was
recoveredinaregionof thechromatogram that didnotcoincide with any of the four markers.
As a control, Escherichia coli DNA
substi-tuted with5-bromouracil and uniformly labeled with 32P was similarly digested and quanti-tated. Figure lla shows an autoradiogram of
the 32P-labeled5-bromouracil-substituted DNA;
5' bromodeoxyuridylic acid migrated to the same location as the 5' terminal nucleotide of
the5-bromouracil-substituted AAV DNA. That the terminal nucleotide ofunsubstituted AAV
a )
A
®/
DNA was thymine was confirmed by labeling
the 5' end of AAV (the duplex this time) not
substituted with bromouracil; 50% of the
re-covered radioactivity was found in thymidylic
acid. Theautoradiogram is shown inFig.
lib,
and it can be seen that the onlyradioactivitydetected was contained in the four common
mononucleotides. The spots were excised, the nucleotides were eluted, and theradioactivity
presentwasmeasured.The resultsaregivenin
[image:11.503.269.459.208.286.2]Table2.
TABLE 2. Determinationofthe5'endsofAAVDNA
%32precovered
Nucleotide
Expt1 Expt2b
5'dAMP 5.9 13.6
5'dTMP 4.4 50
5'dGMP 22 18.5
5'dCMP 8.8 16.9
5'dBrUMP 59
a The minus strand of 5-bromouracil-substituted AAV DNA was used. In this experiment, theends
were quantitatively labeled as calculated fromthe
3Hand32Pspecificactivities. Autoradiogramsof the
plus and minus strands were similar, with mostof
theradioactivityin5'-dBrUMPand alesseramount
in 5'dGMP.
bThe AAV duplex not substituted with
5-bro-mouracil. In this experiment, 50% of the input3H (theDNA was labeled with[3H]thymidine)and 46% of the input 32P (representing 5' terminal
phos-phates) was recovered in the fourmononucleotides. The autoradiogram is shown in Fig. lib.
Co
CL
GC
0S..
G
FIG. 11. Autoradiograms of two-dimensional chromatograms of enzymatic hydrolysates of AAV DNA. Digestions with DNase I and phosphodiesterase I and two-dimensional chromatography on thin-layer polyethyleneimine-cellulose plateswereperformed as describedinMaterials and Methods. The reactions were completesince theonly detectableradioactivity was in mononucleotides; less than 5% ofthe input radioactivity remained atthe origin, and 50 to 75% of input radioactivity was accounted for as mononucleotides. (a)
32P-labeled,5-bromouracil-substitutedE.coliDNA.(b)AAV duplex DNA not substituted with5-bromouracil but
labeledinternally with[3H]thymidine and labeled at the 5' terminus with32P.(c)EcoRIArestriction enzyme fragment terminally labeled with 32p, substituted (about90%0o) with5-bromouracil, and labeled internally
with[3H~thymidine.
J. VIROL.
I-1 i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.503.66.460.419.561.2]ADENO-ASSOCIATED VIRUS DNA 683
TABLE 3. Formation ofprotected 5' ends by incubation with DNA polymerase and DNA ligase"
OXts7DNAb Random linearDNAN AAVDNAd
Prepn
lig lig+pol lig lig+pol lig lig +pol
Control 2.5 4.9 2 1.6 3.35 6.8
Incubated 7 27.5 3.3 8.7 3.35 11.3
Nophosphatase 205 206 91 89 43.5 59
%Protection 2.2 11 1.4 8.0 0 7.6
aThe DNA, internally labeled with
[3H]thymidine
and the 5' end labeledwith32P,was
incubated withpolynucleotide ligase (lig) or polynucleotide ligase plus T4 DNA polymerase (lig + pol) and thentreated withphosphatase (20). The numbers shown in the first three lines are the ratio of 32P counts per minute to 3H counts per minute after acid precipitation. The "control" was not incubated with ligase or ligase plus polymerase. The "No phosphatase" preparation was not reacted with phosphatase. "% Protection" is [(Incubated-Control)/NoPhosphatase] x 100.
b TheOXlinear DNA was purified from phage grown under conditions of ligase deficiency in E. coli ts7 (20).The specific activity was 8.1 x 104counts/min per,ug, and the P/H ratios given in the table have been multiplied by 103.
cThe random linearOXDNAmolecules were prepared by nicking circular single-strandedOXDNA with
DNase I (15). Lessthan one nick per molecule wasintroduced, and the linear molecules were purified by sedimentation on alkaline sucrose gradients. The specific activity of the DNA was 2.5x 103counts/min per
,gg, and the P/H ratios given in the table have beenmultiplied by 10.
d The AAV DNA in this experiment was the 5-bromouracil-substituted Hstrand. Its specific activity was
4.4 x 104counts/min per ,g,and the P/H ratiosinthetable have been multiplied by 102.
In another experiment, the separated
EcoRlA and EcoRlB fragments were
termi-nally labeled with 32p and examined. In both
cases, two spots wereobserved on the
autoradi-ograms: one in the position of
5'-deoxyadeno-sine monophosphate, as expected, since EcoRl
leavesa 5'A(18),and the secondintheposition of 5-bromodeoxyuridylic acid. An
autoradi-ogram of the EcoRlAchromatogram is shown
inFig. lic. Thisexperimentconfirms thatthe
5' ends ofboth the plus and minus strands of
AAV DNA are thymine. Consistent with the
observation thatTA is labeled more efficiently
than the natural end (Fig. 6a) is the
observa-tionthat dAMP is somewhatmoreheavily
la-beled than deoxybromouridine monophosphate. Circularization of AAV DNA. Iwaya et al.
(20) described an experiment that suggested
that linearsingle-stranded kX DNA molecules could beconvertedtocircular molecules under appropriate conditions. The experiment
in-volvedisolating linearDNAfromvirionsmade under conditions ofpolynucleotide ligase defi-ciency,labeling the5'ends with32Pusing
poly-nucleotidekinase,and thendemonstratingthat
someof the 32pcould be made resistantto
bacte-rial alkaline phosphatase by incubation with
T4DNA polymerase and T4 DNAligase. This
could have occurred if a structurelike beta in
Fig. 8b wereformed. Tosee ifAAVcould form
suchastructure, thesameexperimentwas
per-formed with single-stranded AAV DNA. The
details ofthe experimentaregiven inTable 3. Theresultwasthat AAV DNAcouldbe
"circu-larized" toabout thesame extent (5to 10%)as
the linear
OX
DNA molecule. The criteria forcircularization included resistance to alkaline phosphatase and velocity sedimentation in al-kalinesucrosegradients to the positionof
circu-lar DNA; this is not rigorous, however, since
anydimers thatmightbeformedbythe
polym-erase-ligase reaction would also exhibit these
properties. As acontrol, the same experiment
wasperformed with linear
OX
DNAmolecules having randomly located ends generated by nicking circular single-stranded DNA with DNaseI;unexpectedly,protectionof the5'endstoabout thesamelevelwasachieved. Thusit is notpossibletoconclude that the smallamount
of"circularization"observed for AAV DNA and the linear
OX
DNA from the ligase-defectivehostisevidence foranyparticularstructure at
theends. This result alsosuggeststhata major-ity of themolecules cannothave their termini
inthe (3 conformation.
ACKNOWLEDGMENTS
Wethank H. E. David Lane for the32P-labeled, bromo-uracil-substitutedE. coli DNA,Christian Hours for the32p_ labeled4X174RFDNA, Carol Kerr for the[y-32P]ATP,and Linda Pallett for typing the manuscript.
Thisresearch was supported by the National Cancer Institute ofCanada, the Medical Research Council of Can-ada, andtheUnitedStates Public Health Service.
LITERATURE CITED
1. Bartok, K., B. Harbers, and D. T. Denhardt. 1975. Isolation and characterization of self complementary sequences from 4X174 viral DNA. J. Mol. Biol. 99:93-105.
la. Berkowitz, S. A., and L. A. Day. 1975. Molecular weight of single-stranded bacteriophage fd DNA. High speed equilibrium sedimentation andlight scat-teringmeasurements. Biochemistry13:4825-4831. 2. Berns, K. I. 1974. Molecularbiology ofthe
adeno-associ-VOL. 18, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
684 DENHARDT ET AL.
ated viruses. Curr.Top.Microbiol. Immunol. 65:1-20. 3. Berns, K. I., and T. J.Kelly, Jr. 1974. Visualization of the inverted terminal repetition inadeno-associated virus DNA. J.Mol. Biol. 82:267-271.
4. Berns, K. I., J. Kort, K. H.Fife, E. W. Grogan, and I. Spear.1975.Studyof the finestructureof adeno-asso-ciatedvirusDNA withbacterial restriction endonu-cleases.J.Virol. 16:712-719.
5. Blakesley, R. W., and R. D. Wells. 1975. Single-stranded DNA from 4X174 and M13 is cleavedby certain restriction endonucleases. Nature (London) 257:421-422.
6. Carter, B. J., and G. Khoury. 1975.Specific cleavages of adenovirus-associatedvirusDNAbyarestriction en-donuclease REcoRl-characterization of cleavage products.Virology 63:523-538.
7. Carter, B. J., G. Khoury,and D. T. Denhardt. 1975. Physical map and strand polarity of specific frag-ments ofadeno-associated virus DNA produced by endonucleaseREcoRl. J. Virol. 16:559-568. 8. Carter,B.J.,G. Khoury, andJ. A. Rose. 1972.
Adeno-associated virusmultiplication. IX. Extentof tran-scription of the viral genome in vivo. J. Virol. 10:1118-1125.
9. Carter,B.J.,F.J.Koczot,J.Garrison,J. A.Rose,and R. Dolin. 1973. Separatefunction providedby adeno-virusforadenovirus-associatedvirusmultiplication. Nature (London) New Biol. 244:71-73.
10. Carter, B. J., and J. A. Rose. 1974. Transcription in vivo of a defective parvovirus: sedimentation and electrophoretic analysis of RNA synthesized by ad-eno-associated virus and itshelper adenovirus. Virol-ogy 61:182-199.
11. Cavalier-Smith, T. 1974. Palindromic basesequences andreplication of eucaryote chromosomeends. Na-ture(London) 250:467-470.
12. Denhardt, D. T. 1972. A theory of DNA replication. J. Theor.Biol. 34:487-508.
13. Dingman, C. W., M. P.Fisher,and T.Kakefuda.1972. Role ofmolecular conformation in determiningthe electrophoreticproperties ofpolynucleotides in aga-rose-acrylamide gels. Biochemistry 11:1242-1250. 14. Edgell,M.H., C. A. HutchisonIII, and M. Sclair. 1972.
Specific endonuclease Rfragments ofbacteriophage kX174 DNA. J. Virol. 9:575-582.
14a. Eisenberg,S.,B. Harbers,C.Hours,and D. T. Den. hardt. 1975.The mechanism ofreplicationofkX174 DNA. XII. Non-random location of gaps in nascent
OX174RF IIDNA. J.Mol.Biol. 99:107-123. 15. Fiers, W., and R. L. Sinsheimer. 1962. The structure of
the DNA ofbacteriophage4X174.III. Ultracentrifu-galevidence for a ringstructure. J.Mol. Biol. 5:424-434.
16. Gerry, H. W., T. J.Kelly and K. I. Berns. 1973. The arrangementof nucleotide sequences in adeno-associ-ated virus DNA. J. Mol.Biol. 79:207-226.
17. Glynn, I. M., and J. B. Chappell. 1964. A simple method for thepreparation of32P-labeledadenosine triphosphate of high specific activity. Biochem. J. 90:147-149.
18. Hedgpeth, J., H. M.Goodman, and H. W. Boyer. 1972. DNA nucleotide sequence restricted by the R1 endo-nuclease. Proc. Natl. Acad.Sci.U.S.A. 69:3448-3452. 19. Horiuchi, K., and N. D. Zinder. 1975. Site specific cleavage ofsingle-stranded DNA by aHemophilus restriction endonuclease. Proc. Natl. Acad. Sci. U.S.A. 72:2555-2558.
20. Iwaya, M., S. Eisenberg, K. Bartok, and D. T.
Den-J. VIROL. hardt. 1973. Mechanism of replication of single-strandedOX174DNA. VII.Circularization of the pro-genyviral strand. J. Virol. 12:808-818.
21. Johnson, P. H., A. S. Lee,and R. L. Sinsheimer. 1973. Production of specific fragments of 4X174 replicative form DNAby arestriction enzyme from Haemophilus parainfluenzae, endonuclease HP. J. Virol. 11:596-599.
22. Johnson, F.B., H. L.Ozer, and M. D. Hoggan. 1971. Structural proteins of adeno-associated virus type 3. J.Virol. 8:860-863.
23. Koczot, F. J., B. J. Carter,C. F. Garon, and J. A. Rose. 1973. Self-complementarity of terminal sequences withinplus or minusstrands of adeno-associated vi-rus DNA. Proc.Natl. Acad. Sci. U.S.A. 70:215-219. 24. Lee, A. S., and R. L. Sinsheimer. 1974. Acleavage map
ofbacteriophage 4X174 genome. Proc. Natl. Acad. Sci. U.S.A. 71:2882-2886.
25. Lillehaug, J. R., and K. Kleppe. 1975. Kinetics and specificity of T4 polynucleotide kinase. Biochemistry 14:1221-1225.
26. Mayor, H. D., K. Torikai, J. L.Melnick, and M. Man-del. 1969. Plus and minussingle-strandedDNA sepa-rately encapsidated inadeno-associatedsatellite viri-ons.Science166:1280-1282.
27. Middleton, J. H., M. H. Edgell, and C. A. Hutchison III. 1972.Specific fragments ofOX174DNAproduced byarestriction enzymefromHaemophilus aegyptius, endonuclease Z. J. Virol. 10:42-50.
28. Randerath, K., and E. Randerath. 1967. Thin-layer separation methods for nucleic acid derivatives, p. 323-347. In L.Grossmanand K. Moldave (ed.), Meth-ods in enzymology, vol. 12A. Academic PressInc., New York.
29. Roberts, R. J., J. B.Breitmeyer,N. F.Tabachnik, and P. A. Myers. 1975. A second specific endonuclease fromHemophilus aegyptius. J. Mol. Biol. 91:121-123. 30. Rose,J.A.,K. I.Berns, M. D.Hoggan,and F. Koczot. 1969.Evidence for asingle-stranded adeno-associated virus genome:formation of a DNAdensityhybrid on release of viral DNA. Proc. Natl. Acad. Sci. U.S.A. 64:863-869.
31. Rose, J. A., J. V. Maizel, Jr., K. Inman, and A. J. Shatkin. 1971. Structural proteinsof adenovirus-as-sociated viruses. J. Virol. 8:766-770.
32. Schekman,R.W.,M.Iwaya,K.Bromstrup,andD. T. Denhardt. 1972. Mechanism ofreplication of4X174 DNA. III. An enzymicstudy of the structure of the replicative form II DNA. J. Mol. Biol. 57:177-199. 33. Sinsheimer, R. L. 1968.Bacteriophage 4X174and
re-lated viruses. Prog. Nucleic Acid Res. Mol. Biol. 8:115-170.
34. Smith,H.O., and D. Nathans. 1973.Asuggested no-menclature for bacterial host modification and re-striction systemsand their enzymes. J. Mol. Biol. 81:419-423.
35. Sulkowski,E., andM.Laskowski.1971.Inactivationof 5'nucleotidaseincommercial preparations of venom exonuclease (phosphodiesterase). Biochim. Biophys. Acta 240:443-447.
36. Thomas, M., and R. W. Davis. 1975. Studies on the cleavageofbacteriophage lambda DNA with EcoRl restrictionendonuclease.J. Mol. Biol. 91:315-329. 37. Williamson,R. 1970. Properties ofrapidly labeled DNA
fragments isolated from the cytoplasm of primary cultures ofembryonic mouse liver cells. J. Mol. Biol. 51:157-168.
on November 10, 2019 by guest
http://jvi.asm.org/



![FIG....HaeIIIareacrylamide-0.5%digestlabeled of labeled of 6. products of with[3H]thymidine-labeled, Acrylamide-agarose withterminally as 32P described using agarose labeled...](https://thumb-us.123doks.com/thumbv2/123dok_us/1559614.108577/8.503.53.445.324.605/haeiiiareacrylamide-digestlabeled-labeled-products-thymidine-acrylamide-withterminally-described.webp)