0022-538X/79/12-0779/11$02.00/0 Vol. 32, No. 3
Membrane Proteins Specified by Herpes
Simplex
Viruses
V.
Identification of an Fc-Binding Glycoprotein
ROBERT B. BAUCKE ANDPATRICIA G. SPEAR*
Department ofMicrobiology,The University of Chicago, Chicago, Illinois 60637 Received for publication 7 June 1979
Aglycoprotein with affinity for the Fc region ofimmunoglobulinwas isolated from extracts of cultured cells infected with herpes simplex virus type 1, and
experimentsweredone to characterize its properties and to investigate whether
it could account for the Fc-binding activity previously demonstrated on the
surfaces of intact herpes simplexvirus-infected cells. The technique of affinity
chromatographywasusedtoidentifyandisolate the Fc-bindingglycoproteinand
to demonstrate the specificity of its interaction with
inmunoglobulin
G-Fc.Although threeelectrophoreticallydistinguishable Fc-binding polypeptides were
identified by affinity chromatography, these three species appear to be different
forms of the same translation product, based on comparisons of proteolytic
digestionproducts andon the kinetics ofappearance of each form after a brief
pulse with radioactive amino acids. The results suggest that one polypeptide,
designated pE, is processedtoyield
gEj,
which is in turn processed to yieldgE2.Both
gE,
and gE2 areglycosylatedmembrane proteins and both can be labeledby the lactoperoxidase-catalyzedradioiodination of intact infectedcells,indicating
thepresenceof these proteins in surfacemembranes of the cells. Increases in the amountsofgE,and gE2at thecellsurface were found toparallelthe increase in
Fc-binding activity of intact infectedcells.
Cells infected with either serotypeofherpes
simplex virus (HSVtype 1 [HSV-1]or HSV-2)
display membrane receptors thathave affinity
for the FcregionofimmunoglobulinG(IgG) (11,
19, 26, 28, 29, 31,32). These viruses induce the
expressionof thereceptorsinavarietyof differ-entcelltypesthatnormallydonotdisplaysuch
bindingcharacteristics, i.e., culturedepithelioid
orfibroblastic cells. The increase innumberof
receptorsparallels theproduction of HSV
pro-teinsand,infact, has been showntodependon
viralgeneexpression (9,29,31).Morerecently,
ithasbeenreported thattwoother human
her-pesviruses, cytomegalovirus (15, 21, 23, 30) and
varicella-zoster virus
[M.
OgataandS. Shigeta,Abstr. Annu. Meet. Am. Soc. Microbiol. 1979,
S(H)59,p.304],alsoinducereceptorscapableof
bindingtothe FcregionofIgG.
Immunoglobulins from several mammalian
species,includingman, have been showntobind
toHSV-infected cells(31, 32).Assumingthatall
theseimmunoglobulinsbind via their Fcregions
tothe same virus-induced receptor (whichhas notbeen proven in everycase),one couldinfer thatthe receptor hasaffinity forahighly
con-served portion of the antibody molecule. The fact thatidentification and characterization of the virus-induced receptor hassofardepended
on its affinity for IgG should not obscure the
possibility,however, that itmayhave evolvedto
interact with some other molecule, related in
structure toIgG.
Speculationsaboutthefunction(s)of this
her-pesvirus-inducedcell surfacereceptorhave been
basedon theassumptionthatbindingofIgGis
pertinenttoitsroleinviralreplicationor
path-ogenesis. Ithas beensuggested thatbindingof
IgGtothereceptors onthesurfaces of infected
cells could interfere with immune cytolysis (7,
16), and evidence has recentlybeen presented
thataggregated rabbitIgGcaninhibit both
an-tibody-dependent, complement-mediated and
antibody-independent,cell-mediated
cytolysis
ofHSV-infected cells in vitro (1). In addition, it
has beensuggested that the binding ofIgG to
theherpesvirus-inducedreceptors mayinfluence theexpressionofviralgeneproductsin infected
cellsandcouldperhapsleadtomalignant
trans-formationortheestablishment oflatency (8, 16,
31).Withregardtothishypothesisit isof
inter-est that incubation of HSV-infected cells with
relatively high concentrations of nonimmune
rabbit IgG can reduce the yield of infectious progeny recovered from thecells(8).Itremains to be determined, however, whether these
ob-served effects of
immunoglobulin
aredirectly
779
on November 10, 2019 by guest
http://jvi.asm.org/
relevanttothe primary function of the herpes-virus-inducedreceptor.
Regardless of the function of thisreceptor,its affinity for IgGcanbeexploited for the isolation
and characterization of themolecule(s)
respon-sible for theFc-binding activity. Sakuma etal. (23) recently presented evidence that
cytomeg-alovirus receptor-IgG complexescanbe
precipi-tatedby anti-IgG and that thecytomegalovirus Fc-binding receptor is a glycoprotein with an apparentmolecularweight of 43,000.
In this report we describe the identification
andisolation,by affinity chromatography, ofan
HSV-1-induced Fc-binding glycoprotein and also provide evidence that it could accountfor
atleastsome,andpossiblyall, of the Fc-binding
activity detectedonthe surfaces ofinfectedcells.
This Fc-binding glycoprotein accumulates in
two electrophoretically distinguishable forms, whoseapparentmolecularweightsare
approxi-mately 80,000 and 65,000, and is distinct from the HSV-1glycoproteins that have been
previ-ously characterized (6, 12, 17, 22, 24, 25, 27).
MATERIALS AND METHODS Cells and viruses. The cel lines used in these studieswere HEp-2andBHK-21,clone 13, obtained from Flow laboratories (Rockville, Md.). Both cell types were grown in the Dulbecco modification of Eagle minimal essential medium supplemented with 10% heat-inactivated fetal bovine serum. The virus
strains used were HSV-1(HFEM)syn, a syncytial
plaquevariant thatweisolated froma nonmutagen-ized stock ofHSV-1(HFEM) providedbyA.Buchan (University of Birmingham, Birmingham, England), and HSV-2(GP), asyncytial plaque variant isolated
fromHSV-2(G) by Cassaietal. (4). Although HSV-l(HFEM)syncausesextensive fusionof BHKcells,it exhibits much lessfusion-inducing activityinHEp-2 cells. HSV-2(GP),onthe otherhand,induces exten-sive fusion ofHEp-2 cells. Both virusstrains were
propagatedinHEp-2cellsand,fromtheir initial iso-lation, have been passaged onlyafew times at low
multiplicitiesof infection.
Chemicals and radioactive precursors. Re-agentsfor electrophoresis, including acrylamide and the cross-linker N,N-diaVlyltartardiamide, were ob-tained from Bio-Rad Laboratories(Richmond, Calif.). The nonionic detergent Nonidet P-40 (NP40) was
obtained from Gallard-Schlesinger Manufacturing Corp. (Carle Place, N.Y.);cyanogenbromide-activated Sepharose 4B was from Pharmacia Fine Chemicals
(Piscataway, N.J.); bovine serum albumin (BSA),
chickeneggalbumin(OV),andproteaseV8 of Staph-ylococcusaureus werefromMilesLaboratories
(Elk-hart, Ind.); rabbit anti-BSA and anti-OV IgG (each
containing4 to6mgofspecific antibodyperml)were
fromCappelLaboratories (Downington, Pa.);pepsin, glucose oxidase, and lactoperoxidase were from
Worthington Biochemicals Corp. (Freehold, N.J.); chloramine-Twasfrom Sigma (St. Louis, Mo.); and
[35S]methionine (500 to 600Ci/mmol),
['4C]glucosa-mine(45to 60mCi/mmol),and[125I]Na (carrier-free,
17 Ci/mg) were from New England Nuclear Corp.
(Boston, Mass.).
Infectionormock infection of cells and incu-bation with radioactive precursors. Monolayer
cultures of cellswereinfected with virusat aninput
multiplicity of5 to 10PFU/cell.Afteradsorptionfor 2 h at370C,theinoculumwasremoved,and the cells wereincubatedat37°C in medium199supplemented
with1% fetal bovine serum (199-V). When continu-ously labeledextracts weredesired, medium contain-ing[3S]methionine(3,uCi/mlin199-Vpreparedwith one-tenth the usual concentration of unlabeled methi-onine) or['4C]glucosamine (1 ,uCi/ml in 199-V) was addedat 3hafterinfection,andincubationwas con-tinuedat370Cuntil 18 h afterinfection,atwhich time the cellswereharvested. Forpulse-labeled extracts, infected cellswere incubated for 5 minat 6h after infection with [nS]methionine (10l,iCi/ml in 199-V lacking unlabeled methionine), and the cells were
either harvested immediatelyor after continued in-cubation in nonradioactive 199-V.
Cellsurface iodination.Cellsurfaceproteinswere
iodinated with "2I accordingto the
lactoperoxidase-catalyzed procedure describedby Hubbard and Cohn (13).Briefly, cellmonolayerswereoverlaid with
phos-phate-buffered saline(PBS;10mMNa2HPO4,1.5 mM KH2PO4, 0.14 M NaCl, 3 mM KCI,0.5mM MgCl2.
6H20,1mMCaCl2), pH7.4,containing5mMglucose
and [251I]Naat400,uCi/ml.Lactoperoxidaseand
glu-coseoxidasewereaddedto give final concentrations of 20ug/mland0.1U/ml, respectively. After10min atroomtemperatureon ashaker, unreacted125Iwas
pouredoff and the cellswerewashed andharvestedas usual.
Preparation of cell extracts. After cell mono-layerswerewashed withPBS,thecellswerescraped witharubberpoliceman and collected by centrifuga-tion. For extraction ofproteins the cells were
sus-pendedinPBS containing1%NP40 (1.0ml/2 x 107 cells) andkeptonice,with occasionalmixing, for15 min.Nucleiwerethen removedby low-speed
centrif-ugation,and thecytoplasmicextracts werestoredat
-70°C. Immediately before usefor affinity
chroma-tography,theextractswerethawed andcentrifugedat 25,000 rpm in an SW 27.1 rotor for 2 h to remove insoluble material.
Preparation and use of Fc affinity columns.
BSA orOV wascovalently linkedtocyanogen bro-mide-activatedSepharose4Baccordingtoprocedures supplied byPharmacia. Typically, 15g ofcyanogen bromide-activatedSepharose4B wasrehydratedand washed with1mMHCIonasintered-glass filter and then equilibrated with coupling buffer (0.1 M NaHCO3, pH 8.3, containing 0.5 MNaCl).BSAor OV dissolved incoupling buffer was added to give a final concentration of 5 to 10 mg ofprotein per ml, and couplingwasallowedtoproceed at roomtemperature for2hwithshaking ofthemixture. Unreactedprotein wasthen washed out with coupling buffer, and the remaining active groups on the Sepharose were blocked with 1Methanolamine, pH 8.5 (2 h at room temperature). The Sepharose was washed several times with coupling buffer, alternating with 0.1 M sodiumacetatebuffer(pH 4.0) containing0.5 MNaCl,
on November 10, 2019 by guest
http://jvi.asm.org/
toremoveproteins that were adsorbed but not cova-lentlycoupled to the beads.Finally,the
protein-con-jugated Sepharose was equilibrated with PBS and stored at 4°C with 0.02% sodium azide.
Affinity columns were prepared by pouring suffi-cient BSA- or OV-conjugated Sepharose into 5-ml polypropylene columns toyield a bed volume of
ap-proximately1 ml. Thiswaswashed andequilibrated
with PBS-0.5% NP40 by passingatleast 25 ml of this bufferslowly through thecolumn,and then0.4 to 0.6 mgofspecific anti-BSAoranti-OVIgG (1.0to 3.0mg of totalIgG) in1.0ml of PBS-0.5%NP40wasadded. Thissolution wasallowed to enter the bed slowly over aperiodof1hat roomtemperature,andthe unbound
immunoglobulin was then washed out with excess
PBS-0.5% NP40. The columns were equilibrated at
370C, and NP40-solubilizedcellextractswere added
(3 ml/column). Every 15 min approximately 0.5ml wasallowedto enterthebeduntil all had beenapplied,
after which the columnswerewashedextensivelywith PBS-0.5% NP40.Finally,column-bound proteins
(in-cludingimmunoglobulin)wereeluted with 2to3mlof 3 Mpotassiumthiocyanate inPBS-0.5%NP40. The eluted macromoleculeswereprecipitatedwith 5% tri-chloroacetic acid, and theprecipitates were washed
sequentiallywith ethanol andacetoneand then solu-bilized foranalysis bysodiumdodecylsulfate
(SDS)-acrylamidegel electrophoresis (12).Greater than 70% of trichloroacetic acid-precipitable radioactivity was recovered for theanalysis.
Preparation of
F(ab')2
fragments from IgG. Rabbitanti-BSAIgGwastreated withpepsintoyield F(ab')2 fragments according to the procedure de-scribedbyNisonoff (20).Afterdialysisof theIgG (7 mg/ml) againstseveralchangesof0.1Msodium ace-tate,pH7.8, thepHwasadjustedtopH4.5with1M acetic acid andpepsinwasaddedtoyielda 1:50(wt/ wt)ratioofpepsintoIgG.This mixturewasincubatedovernightat370Candthenneutralized withNaOHto inactivatethepepsin.Asacontrol,anequalamount ofIgG wastreatedidenticallyexcept thatnopepsin
wasadded("mock-treated"IgG).Ouchterlony
double-diffusionprecipitationtestsindicated that the pepsin-treated, mock-treated, and untreated IgG had com-parable precipitating activity with BSA (data not shown).
Preparation and use of radiodinated
immu-noglobulin. RabbitIgG wasiodinated with "I ac-cordingtothe chloramine-Tmethoddescribedby Byrt
and Ada (3)and modifiedbyJensenius and Williams (14). A
10-Pd
amountof["lI]Na
(5mCi)wasaddedto 5mlofchilled rabbitIgGsolution (1.5mg/mlin0.05 M sodium phosphate buffer, pH7), followedbythe addition andrapidmixingof100pl
ofchloramine-T solution (1mg/ml).After10minonice the reaction wasstopped byaddition ofexcesstyrosine (100piat 0.4 mg/ml).TheIgG wasisolated from thereaction mixture bychromatography on Sephadex G-50 and hadaspecificactivityofapproxiimately 108 cpm/mg of IgG. This preparation was diluted 10-2 in PBS containing 5% heat-inactivated fetal bovineserumfor incubationwith intactcellmonolayerstomonitorthe appearance ofFc-bindingreceptors at varioustimes after infection. Incubationwasfor2hwithshakingat370C,
afterwhich thecells
were washed extensivelywith PBS-5% heat-inactivated fetal bovine serum, sol-ubilized in 1%SDS,and sonicated briefly; small sam-ples were taken for quantitation of bound radioactiv-ity.
Isolation and partial proteolysis of the Fc-bindingpolypeptides.Pulse-labeled or continuously labeled forms of the Fc-binding polypeptides were isolated byaffinity chromatography, followed by pre-parative acrylamide gel electrophoresis, for partial
proteolysis and analysis of the peptides. For each analysis extracts were prepared from approximately 1.2x108cells and applied to a single affinity column. Thepolypeptides eluted from the column were con-centrated by trichloroacetic acid precipitation as de-scribed above andfractionated byelectrophoresison
SDS-acrylamide gelslabs(1.5mmthick). Wide sample wells (4 cm) were used to accommodate the large amountofmaterial. Both thefast-migratingandmore
slowlymigrating Fc-binding polypeptides were local-ized in the gel by either punching out segments of the
gelforquantitationofradioactivityorestimatingtheir positions relative to BSA (the location of BSAwas determinedbystaining one ortwo vertical strips of the gel; the remainder of thegelwasnotstained or fixed).
Ahorizontal strip of gel containing the desired Fc-binding polypeptide was cut from the gel slab and divided into 5-mm segments for the peptide-mapping procedure described by Cleveland et al. (5). Briefly, the gel segments were placed in sample wells of a second acrylamide gel (15% separating gel prepared with aN,N-diallyltartardiamide cross-linker instead ofbisacrylamide) andoverlaid with buffer containing variousconcentrations of the staphylococcal V8 pro-tease.Theresultingproteolyticfragmentswere sepa-ratedbyelectrophoresis at constant current (12mA/ gel) for about 6h. The gels were fixed and stained according to the procedure of Fairbanks et al. (10), and the radioactive bandswerelocatedby fluorogra-phy (2).
RESULTS
Detection of an IgG-binding protein in
extracts from infected celils. The technique
ofaffinity chromatography was applied to the
isolation ofamolecule(s) responsible for
HSV-1-inducedFc-binding activity. The affinity
col-umnsconsistedof rabbitantibodyboundto
an-tigen which itself was covalently coupled to
Sepharose 4B.Inallexperimentsreportedhere,
the antigen was either OV or BSA and the
antibodies had been prepared against OV or
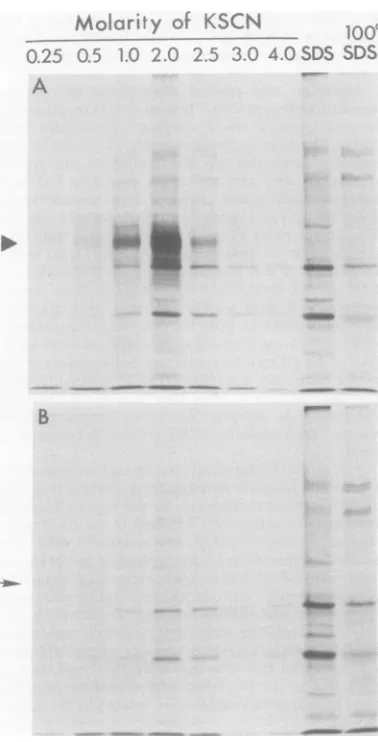
BSA,respectively. Inthe experimentshown in Fig. 1, extractspreparedfrom
[3S]methionine-labeled infected cells, byuse of the detergent
NP40 as described in Materials and Methods,
were applied to a pair of OV-conjugated
col-umnns, onlyoneof whichcontained boundIgG.
After being extensively washedto remove
un-bound proteins, the columnswere sequentially
elutedwith2-mlaliquotsofpotassium
thiocya-natesolution,ranginginconcentration from0.25
on November 10, 2019 by guest
http://jvi.asm.org/
782 BAUCKE AND SPEAR
A
FIG. 1. Elutionofpolypeptides bound Sepharose (A) orto controlOV-Sepharo
tractswereprepared from HEp-2cells in HSV-1(HFEM)synandincubated with[
ninefrom3to 18 hafter infection. Sam.
extractswereappliedtocolumns A and extensivewashingto removeunboundpi
columnswereelutedsequentially with 2-i ofthe solutionsindicated, asdescribed
The labeledpolypeptidespresentin each4 tionwereanalyzed by electrophoresison
amidegel slabs, autoradiograms ofwhich to 4 M, and then with 1% SDS. Fi Sepharosewassuspended inasecond
1% SDS and heatedto1000C for2mi
supernatant was recovered for ana
elutedproteins in each of these fraci precipitated by trichloroacetic acid,s
andanalyzed by SDS-acrylamide ele sis. Theautoradiogram shown in Fig.
stratesthespecific binding ofatleast peptide (indicated by the filled
triani
IgG-OV-Sepharose column(panel A) butnot to
I - the control OV-Sepharose column
(panel
B).SDS SDS Thisproteinwas
optimally
elutedbypotassium
thiocyanate in therangeof 1.0to2.5 M. Small
quantities ofaproteinwith similar
electropho-reticmobilityhad boundtothecontrol column
(indicated by the arrow), but its elution at a
lower molarity of potassium thiocyanate
sug-geststhat it is eitheradifferentprotein of
ap-proximately the same molecularweight or the
sameprotein capable of interacting inadifferent
fashion with the control column.
_mww
An experiment was done to determinewhether theIgG-binding protein wasproduced
only after infection orcould also be detected in
extracts from uninfected cells. Extracts were prepared from HSV-1-infected and from
mock-infected cells labeled with [3S]methionine for
15h(from3 to 18hafterinfection),andaliquots
wereappliedeithertoIgG-BSA-Sepharose col-umns or to control BSA-Sepharose columns.
After
being
washed,
thecolumn-boundproteins
wereelutedin asinglestepwith3Mpotassium
thiocyanate and prepared for electrophoresis.
Figure 2 shows that the IgG-binding protein
(indicated by the filled triangle) could be
iso-lated byaffinitychromatography fromextracts
of the infectedcells butnot from those of the
uninfected cells. It should be noted that this
IgG-bindingproteinisaminorcomponentof the
infectedcellextractsand isbarelydetectable in
electropherograms of unfractionated extracts.
For theelectrophoretic analysisshown in Fig.2,
theIgG-binding protein eluted from the affinity
column andappliedtothegelwasobtained from
- approximately80timesasmuchextract aswas alsoappliedtothegelforcomparative purposes. toIgG-OV- Results
indistinguishable
from those shown in se(B).
Ex.
Fig. 2 were obtained whether OV or BSA wastfectedwith
35simethio-
usedasthecovalently
coupled
antigen
(data
notwples
oftheshown),
indicating
that interaction of theIgG-,
and,
afterbinding protein
with theaffinity
column wasroteins, the
independent
of theantigen
used.mlaliquots Inpreliminary studies,wehave also identified inthetext. an
IgG-binding
protein induced after infec-elutedfrac- tion ofHEp-2 cells by an HSV-2 strain[HSV-SDS-acryl-
2(GP)].
Theresults
obtained by affinitychro-are
shown.
matographywereverysimilar
tothoseshown inFig. 2, and theIgG-binding protein inducedby
inally, the HSV-2(GP) was indistinguishable by
electro-aliquotof phoretic mobility from theprotein induced by
in,and the HSV-1(HFEM)syn.
lysis. The Glycosylationof the IgG-bindingprotein.
tions were To determinewhether the IgG-binding protein
;olubilized, is glycosylated,extractswere also preparedfrom
ctrophore- HSV-1-infected cellslabeled with
["4C]glucosa-1demon- mine. Column-bound proteins from these
ex-tonepoly- tracts are compared with
[3S]methionine-la-gle) tothe beled, column-bound proteins in Fig. 3. The
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.63.252.70.441.2]Uninfected Infected
Column-bound proteins
Ext C IgG IgG C Ext
- -130 K
gB-gA,
4C-glu-NH2 35S-meth
Column-bound proteins
Ext C IgG IgG C Ext
S.F
S 130 K
[image:5.504.248.451.60.265.2] [image:5.504.54.245.74.290.2]1 - 94 K
-° I D | - 68 K
...M 4
- 94 K
- 68 K gD- °
.-...4...
..- 43 K *5W
FIG. 2. Specific bindingtoIgG-BSA-Sepharoseof
a polypeptide produced by
HSV-1(HFEM)syn-in-fected HEp-2 cells, butnot byuninfected cells.
Ex-tractswereprepared from infectedormock-infected
HEp-2 cellsthathad beenincubated with
[rS]me-thioninefrom 3to 18 haftertheaddition ofvirus. Thisautoradiogram ofanacrylamide gelslabshowsthe labeledpolypeptides that were bound to and elutedfromeitherIgG-BSA-Sepharose(IgG)or
con-trolBSA-Sepharose (C)andalsothelabeled
polypep-tidespresent in unfractionated extracts (Ext) from the uninfectedor infected cells (the column-bound
proteinswereobtainedfromapproximately80times
asmuchextractas waspresentintheunfractionated sample appliedtotheacrylamide gel).Inthis andall
subsequentfigureselutionwasachieved inasingle
stepbytheapplicationof 3Mpotassium thiocyanate, after extensive washing ofthe affinity columns to
remove unbound proteins. The numbers along the
right edge indicate the positions of unlabeled poly-peptidesusedasmolecularweightstandards (/3-ga-lactosidase, 130,000; phosphorylase b, 94,000; BSA, 68,000; OV,43,000).
results show thataglucosamine-containing
poly-peptide binds specifically to
IgG-BSA-Sepha-rosecolumns and thatits
electrophoretic
mobil-ity is the same as thatof the [
S]methionine-labeled, IgG-binding protein (filled triangles). Another glycosylated polypeptide of lower ap-parent molecular weight (open triangles) also
appearstobindspecifically tothe IgG-contain-ing column. Variable quantities of this species
were detected in replicate experiments.
Evi-dence will be presented below which suggests
thatthesetwoIgG-binding speciesarerelated in aminoacidsequence.
Demonstration that theIgG-binding
pro-teininteracts with theFcregionofIgG.To
FIG. 3. Presenceof radiolabelfrom
['4C]glucosa-mine, as well as from [35Slmethionine, in HSV-1-inducedpolypeptidesthat boundspecificallyto
IgG-BSA-Sepharose. This autoradiogram of an
SDS-acrylamide gelslabshows thelabeledpolypeptides
presentinunfractionatedextracts (Ext)from
HSV-1(HFEM)syn-infected HEp-2cells and the
polypep-tidesboundtoandelutedfromeither
IgG-BSA-Seph-arose(IgG)orcontrolBSA-Sepharose (C). The
poly-peptideindicatedbythefilled triangleshas thesame
electrophoretic mobility as the major IgG-binding
species shown in Fig. 1 and2. In different experi-ments, variable quantities ofthepolypeptide indi-catedby the opentriangles werealsofoundtobind
specificallyto the IgG-containing affinitycolumns. Thesymbolsshownontheleft edgearedesignations ofthemajorHSV-1glycoproteins, which have
previ-ously beendescribed (6, 27).
determine whether binding of the glycosylated
polypeptidestoIgGdependsupon anintact Fc
region, rabbit anti-BSA IgG was treated with
pepsinto yield F(ab')2 fragments, as described
in Materials and Methods. The F(ab')2
frag-ments were demonstrated tohaveretained
an-tigen-bindingactivityanddivalency by
Ouchter-lony gel diffusion tests with BSA (data not
shown). Extracts from[3S]methionine-labeled,
HSV-1-infected cells were applied to F(ab')2-BSA-Sepharose columns. As controls, equal
amounts ofextract were also applied to
BSA-Sepharosecolumns andto
IgG-BSA-Sepharose
columns which had been prepared with either
untreated IgGorIgG subjected tothe reaction
conditionsused forpepsindigestion (inthe
ab-senceofprotease). The resultsshown inFig.4
illustratethatmarkedlylessof theIgG-binding
proteinwasretainedonthe
F(ab')2-BSA-Seph-arose column compared with retention by the
IgG-BSA-Sepharosecolumn.Thissuggests that
the bindingis mediated through the Fcregion
oftheIgGmolecule.
VOL. 32,1979
on November 10, 2019 by guest
http://jvi.asm.org/
784 AND SPEAR
.c>,
.> e i ..
9g
l.i 1 3C K
-Q4 K
68K
FIG. 4. Reduction in binding of the
HSV-1-in-ducedpolypeptidestotheaffinitycolumnsbyremoval
of the Fcregion ofIgG.Affinity columnswere pre-pared by the additiontoBSA-Sepharoseofuntreated
anti-BSA (IgG) orofanti-BSA which had been
di-gestedbypepsintoyieldF(ab')2 fragments[F(ab')2]
orhad beensubjectedtothesameconditions usedfor
proteolysis in the absenceofpepsin(IgG°).Anextract waspreparedfromHSV-I(HFEM)syn-infected
HEp-2cells that had been incubatedwith[35S]methionine from 3 to 18 h after infection, and aliquots were appliedtothe threeaffinitycolumnsandtoacontrol BSA-Sepharose (C)column. Thisautoradiogramof
anSDS-acrylamide gelslab shows thepolypeptides
boundtoand elutedfromeach columnalongwith the
polypeptidespresentin a sampleofthe
unfraction-ated extract (Ext). The filled and open triangles
indicate thepositionsoftwo IgG-binding
polypep-tides,asdescribed in thelegendtoFig.3.
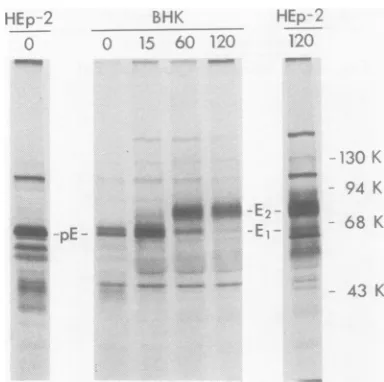
Newly synthesized and stable forms of
the Fe-binding glycoprotein. Affinity
chro-matographyofextractsfrompulse-labeled
HSV-1-infected cellswasdone todetermine whether
newlysynthesized forms of the Fc-binding
poly-peptide(s) could bind to the IgG-antigen
col-umns and might perhaps differ in
electropho-retic mobility from theprocessed, stable forms.
Infectedcellswerepulse-labeledfor 5 min with
[3S]methionineat 6 hafter infection and then
harvested immediately or after 15, 60, or 120 min of continued incubation in nonradioactive medium. BothHEp-2and BHK cells were used for thisexperimentinorder to compare the
Fc-binding polypeptides produced bydifferentcells.
The results shown in Fig. 5 demonstrate that
three electrophoretically distinct polypeptides bound specifically to the IgG-BSA-Sepharose columns (control columns without IgG not shown).
Two of these forms, designated pEand gE1,
were detected immediately after the
pulse.
[image:6.504.264.456.71.262.2]p~
FIG. 5. Newlysynthesizedandprocessed forms of
Fc-bindingpolypeptidesproducedinHEp-2cells and
BHKcells infected with HSV-1(HFEM)syn.At 6h after infection the cells were incubated with [35S
methioninefor 5min, and then extracts were pre-pared immediately (0) or afteran additional incu-bationofthe cellsin nonradioactive medium(for15, 60, or 120min). The extracts were applied to
IgG-BSA-Sepharosecolumns andtocontrol
BSA-Sepha-rosecolumns, and the boundproteinswereelutedfor electrophoretic analysis. The designations pE, E1,
andE2 indicate thepositionsofpolypeptides bound
specifically to theIgG-BSA-Sepharose columns but not tothecontrolBSA-Sepharosecolumns(controls
notshown).
Within 15minafter thepulseathirdform,gE2,
wasdetectable.By60 minpE had disappeared,
and by 120 min very little of gE1 remained.
These resultsareconsistentwiththepossibility
thatallthreeelectrophoretic formsarederived
from thesametranslationproduct and that
post-translationalprocessingoccurswith the
conver-sion ofpE to
gEI
andgEI
togE2. The resultsobtained with infected HEp-2 and BHK cells weresimilar, althoughpEand
gE,
producedin the BHK cellswere more easily recognized as distinct species. Because the apparent conver-sion ofgEltogE2is notcompleteeven after2 h, it seems likely that extracts of infected cellslabeled forlong periods of time wouldcontain
both ofthesespeciesand thatgEIisthe
faster-migrating Fc-binding glycoproteinseeninFig.3
and4(marked bytheopentriangles).
To testthepossibilitythat gE2 has amino acid sequences in common with pE and gE1, these
[3S]methionine-labeledspecieswereisolated by
affinity chromatography and preparative
elec-trophoresisandsubjectedtopartial proteolysis
by the procedure of Cleveland et al. (5). The
diffemoo
lo. k:...:..
4800ma so
on November 10, 2019 by guest
http://jvi.asm.org/
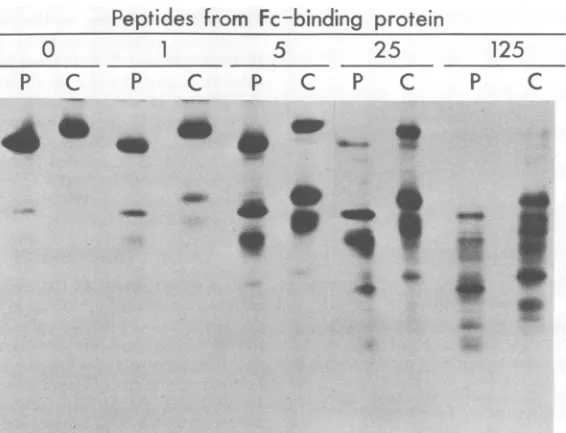
[image:6.504.87.231.74.252.2]partial proteolytic products obtained with
var-ious concentrations of the staphylococcal
pro-teaseV8areshown inFig.6. It canbe seen that,
at allconcentrations of enzyme tested, the
elec-trophoretic profile of peptides from the
pulse-labeled material (pE plus
gEj)
issimilar to thatofpeptides from the moreslowlymigrating
sta-ble material (gE2), except that the profiles are
displaced to the extent that the uncleaved
poly-peptides differ in mobility. This suggested the
possibility that all the [3S]methionine-labeled
peptides obtained fromgE2 contained the
mod-ification responsible for the slower
electropho-reticmobility of intact gE2. If this were true, and
ifthemodification includes glycosylation, then
all themethionine-containingpeptides from gE2
should containcarbohydrateand would
proba-blyincorporate label fromradioactive
glucosa-mine. Theresults shown inFig.7demonstrate,
infact, thatallthe[3S]methionine-labeled
pep-tides derived from gE2 also incorporated label
from
["4C]glucosamine.
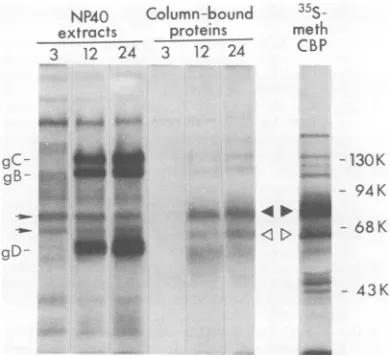
Expression of the Fc-binding
glycopro-tein on thesurfaces ofinfected cells. Other
investigators have quantitated the binding of
125I-labeled
IgG to HSV-infected cells toinves-tigate the kinetics of appearance ofFc-binding
activity in the infected cell surface (18, 31). Sim-ilarexperimentswereperformedinour
labora-tory(results shown in Fig. 8). Fc-binding activity
became detectable bythis assaybetween3 to6
hafter infection and reachedamaximumat 24
h. Todetermine whether the Fc-binding
glyco-protein, detected by affinity chromatography,
appeared in the cell surface withsimilarkinetics,
the following experiment was done. Intact
in-fected cellswereiodinated with
"2I
byalacto-peroxidase-catalyzed reaction at 3, 12, or 24 h
afterinfection.Detergentextractsof these
iodi-nated cells were then preparedand applied to
bothIgG-BSA-Sepharose columns and control
BSA-Sepharose columns. Figure 9 shows an
electropherogram of the
"MI-labeled
polypep-tides that were boundto and eluted from the
IgG-containing columns; the triangles identify
twospecies that boundspecificallyto the
IgG-containing columns butnot to the control
col-umns(controlsnotshown).Thesetwo
polypep-tides, whoseelectrophoretic mobilitiesare
indis-tinguishable from those of
gE,
and gE2(Fig.
5and 9), werevirtually undetectable in extracts
prepared from cells at 3 h after infection,
whereas significant quantities were present in
the extractspreparedat 12 and24h.Asis also
evident fromFig.9,sampleseluted from theIgG
affinity columns sometimes contained
hetero-geneous labeled material whose
electrophoretic
mobilitywasgreater than that of themajor
Fc-binding species.Therelationshipof this material
togE remainstobedetermined.
Peptides from
Fc-binding
protein
1
5
P
C
P C
44Ih~ ~ ~ ~ ~ ~~~4
FIG. 6. Peptidesobtainedfr-omnewly synthesized (P)andprocessed (C) forms oftheFc-bindingglycoprotein afterpartial proteolysiswith various concentrations(0 to125 pg/sample) ofS.aureusproteaseV8. The pulse-labeledforms ofthe Fe-binding glycoprotein (P)arethespecies designated pEandElin the kegendtoFig.5, andthepulse-chatsedform (C) isE2. Thesepolypeptideswere isolatedforthisanalysisasdescribed in the
text.
0
P C25
P
C
125
P
C
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.107.390.392.609.2]786 BAUCKE AND SPEAR
Peptides
from
Fc-binding
prore,
:.Fg aci g
4
_l
w-"I
FIG. 7. Peptidesobtainedfr-om[35S]methionine-labeled(aa)and[14C]glucosamine-labeled (g) Fc-binding glycoproteinE2afterpartial proteolysiswith various concentrations(0 to250pg/sample) ofS.aureusprotease
V8.
30
25[
x
20
0
-o0
02 15 U0
10
[
5
6 12 18 24 30
Hoursafterinfection
FIG. 8. Bindingof[125IJIgGto HSV-1(HFEM)syn-infected (filled circles) anduninfected (open circles) HEp-2 cells,as afunction oftimeafter infection or
mockinfection.Radioactivitywasquantitatedby
liq-uid scintillationspectrometry; the valuesgiven are
countsperminute boundper4x 1(1 cells.
The previously identified HSV-1 glycopro-teins gC, gB, and gD werereadily detected as
125I-labeled
bands in electropherograms of the unfractionated extractsprepared from the iodi-nated infected cells. The results presented inFig.9indicate thatsignificantlygreater
quanti-ties of theseproteinswerepresent inthesurfaces ofinfected cells at 12 and 24 h afterinfection than at 3h,as wasalso foundfortheFc-binding
polypeptides. Several other polypeptides
de-tected in the unfractionated extracts were
la-beledtoapproximatelythesameextent atboth
early and late times after infection andare
prob-ably cellular proteins, as they comigrate with
5I-labeled polypeptides from uninfected cells
(datanotshown).Two of theselabeled
polypep-tides (indicated bythe small arrows in Fig. 9)
are similar in electrophoretic mobility to the
virus-induced Fc-binding polypeptides gE, and
gE2 but are probably unrelated to them. The
contribution of
l2II-labeled
Fc-bindingpolypep-tidestoradioactivityin theextractsis too
sniall
tobedetected in electropherograms of the
un-fractionatedextracts.
DISCUSSION
Thisreport describes the isolation and char-acterization of an Fc-binding glycoprotein
pro-duced in HSV-1-infected cells. Several
proper-ties ofthisglycoprotein, designated gE, suggest
that itisresponsibleforthepreviouslydescribed
Fc-binding activity ofintactinfectedcells(9, 11,
18, 19, 26,28, 29, 31, 32). (This Fc-binding
gly-coprotein is thefifth antigenically or
function-allydistinct HSV-1-induced glycoproteintobe described. Assignment of thealphabetic desig-nationgEconforms tothe nomenclature agreed uponbyagroup ofparticipantsatthe Herpes-virus Workshop held in August 1978 in
Cam-bridge,England.)First, gEbinds toIgGandthis
0 0
/Z
/.
./
O O. Oon November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.123.409.75.274.2] [image:8.504.68.263.317.551.2]HERPESVIRUS Fc-BINDING GLYCOPROTEIN 787
NP4O Column-bound
35S-
pany posttranslational modifications (6,27).Theextracts proteins meth finding that newly synthesizedaswellasstable 3 12 24 3 12 24 CBP forms ofgE could bind to the affinitycolumns
indicates that Fc-binding activitydoes not
de-pend on complete processing or complete gly-cosylationof theprotein.It remainstobe
deter-mined whether allforms of theFc-binding
pro-gC-
" > -130K tein,including
the two forms detected on cellgB- surfaces,interact thesamewaywithIgG.
- 94 K
Although
thepossibility
existsthatgE
is an_U-A ."_
**4
induced cellular gene product, it seems moreM-=
= < D ^-
68Klikely
that thepolypeptide moiety
of thisFc-gD binding glycoprotein will prove to be of viral
43K genetic origin. First, gEisapparently produced
-43 K onlyafter infection of the cell linesanalyzedin
thisstudyand issynthesizedat atimewhen all
or mosthostprotein synthesisisinhibited.
Sec--- ond,theelectrophoreticmobilities of the various
FIG. 9. Presence and kineticsofappearanceofthe gE formsproducedinHEp-2cellsaresimilaror
Fc-binding glycoproteinon thesurfaces of infected identical to those produced in BHK cells.
Fi-cells. HEp-2 cells infected with HSV-1(HFEM)syn nally, recent studies (M. Para, R. Baucke, and
were labeled with
125I,
by aprocedure whichlabels p. G. Spear manuscript in preparation) haveonly proteins exposedtothe extracellularfluid soP
thatg
is a*constin
pofthenvirio
<¢v¢vsw- * r *
_rd,
at 3 shown thatgE
iS a constituent of the virion12, or24 hafter infection. Extracts were then
pre-pared andapplied toIgG-BSA-Sepharosecolumns envelope as well as of the
mfected
cell surface.and to control BSA-Sepharose columns. This auto- It can be concluded from the publications
radiogramofanSDS-acrylamidegelslab showsthe cited in the firstparagraphof this section that
'25I-labeledpolypeptides bound to andeluted from a number ofdifferent HSV strains caninduce
IgG-BSA-Sepharose,aswellasthelabeledpolypep- Fc-bindingreceptors,suggestingthatexpression
tidespresentinsmallsamples oftheunfractionated of this receptor is not a peculiarity of a few
extracts.Also shown isaprofileof[3Slmethionine- strains. There are differences among HSV
labeledpolypeptideswhich bound to and were eluted strains, however, in the number of receptors
from
anIgG-BSA-Sepharosecolumn. detectable by "2I-labeled IgG binding to thesurfaces of infected cells. We choseto use
HSV-binding depends
uponintegrity
of theFcregion. l(HFEM)syn for the studies reported here, onSecond,
gE
is a constituent of theinfected cell the basis ofscreening several 1 andHSV-surface.
Finally,
the time of its appearance in 2strains formaximal
expressionof cell surfacethe cellsurface correlates with the time ofap- receptors.Theresultsof this screening suggested
pearance of Fc receptors on intact cells. The that syncytial plaque variants of eitherHSV-1
association of
Fc-binding
receptoractivitywith orHSV-2expressedlarger numbers ofFc-bind-amembrane
glycoprotein
producedafter infec- ing receptors than did wild-type strains (R.tion is consistent with
previous
reportsthat this Baucke and P. G. Spear, unpublished data).Itreceptoris
trypsin
sensitive and that itsexpres- remainsto be determined whether theexpres-sion is
prevented
by deoxyglucose
andbyinhib- sion of Fc-binding activity is related tovirus-itorsof
protein synthesis
(9, 18, 31). induced cell fusion and whether the apparentAlthough
Fc-binding activity
wasfoundtobe differences in numbers ofreceptors areduepri-associated with three
electrophoretically
distinctmanly
tovariabilityin theamountofFc-bindingpolypeptides,
ranging
in apparent molecular glycoprotein produced, its intracellulardistri-weight
from 65,000 to 80,000, it appears that bution,oritsbindingcharacteristics. Inprelim-these
polypeptides
shareamino acidsequences inary studieswehavefound that theFc-bindingin common.Both the kinetics of appearance of glycoproteininducedby HSV-2(GP)is indistin-each
Fc-binding polypeptide
after a pulse of guishable from theHSV-1(HFEM)syn glycopro-radioactive amino acid and theresults ofcom- teinbytwocriteria,electrophoretic mobilityofparisons by partial
proteolysis suggest that a intactSDS-solubilizedproteinsand mobilities ofsingle
translationproduct
yieldsallthree forms peptides obtained afterdigestion with V8pro-by sequential
stages ofposttranslational
proc- tease (BauckeandSpear, unpublished data).essing.
Previous studies revealed that theHSV- The molecule reportedto beresponsible for1
glycoproteins gB,
gC, and gD are processed cytomegalovirus-induced Fc-binding activity similarly togE,
at least with respect to the (23) differs in at least two respects from thechanges
inelectrophoretic mobility
thataccom- HSV-1-inducedFc-binding gE. First,theappar-32,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.504.47.242.76.255.2]ent molecular weight of the
cytomegalovirus-inducedFc-bindingglycopolypeptide is43,000 as
comparedwith65,000to80,000forthe various
forms of gE. Second, the
cytomegalovirus-in-duced glycoprotein was isolated from cellular
extractsprepared without the use ofdetergents (and therefore may not have been membrane
bound), whereasHSV-1gEcouldnotbe
solubi-lized withoutdetergents.Itwill be of interestto
determine whether detergents can solubilize
other forms of the cytomegalovirus-induced
Fc-binding glycoprotein and to compare the
propertiesof theFc-binding proteinsinducedby
HSV,cytomegalovirus,andvaricella-zostervirus
(see referencecitations inIntroduction).
The function of the herpesvirus-induced
Fc-binding glycoprotein and its role in viral
repli-cation orpathogenesis,orinboth,remaintobe
determined. Thefinding thatFc-bindingactivity
canbe detectedonthe surfaces ofcells infected
by three different human herpesviruses
under-scores the possible importance of this activity.
The fact that detection ofthe binding activity
hassofar reliedontheuseofIgG,however, does
not necessarily imply that the purpose of the
virus-induced receptoris to interact with IgG.
Anunderstandingof the function of this
recep-tor,which in the caseof HSV-1 is expressedon
thesurfaces of both virions(Paraetal., in
prep-aration) and infected cells, will depend upon
additional experimentation, including
explora-tion of thepossibilitythat the receptor mayhave
affinity for molecules other than IgG.
ACKNOWLEDGMENTS
This workwassupported bygrant VC125 from the Amer-ican Cancer Society. P.G.S. is recipient of Public Health Service Research CareerDevelopmentAward5K04-CA00035 from the National Cancer Institute.
ADDENDUM IN PROOF
AttheHerpesvirusWorkshopheld in August1979
(ColdSpringHarborLaboratory,ColdSpring Harbor, N.Y.),agroupof theparticipants agreedupona con-ventionfor thenamingof the variousprocessed forms ofasingle HSVglycoprotein.Accordingto this con-vention,pEshould bedesignated pgE(64),gE,should bepgE(66), andgE2 should be gE. The numbers within parenthesesareapparentmolecularweights (x10-3),
usedtodifferentiate amongmultiple precursor forms.
LITERATURE CITED
1. Adler, R., J. C.Glorioso,J.Cossman,and M. Levine. 1978.Possible role ofFcreceptors oncellsinfectedand transformed by herpesvirus: escape from immune cy-tolysis. Infect.Immun.21:442-447.
2. Bonner, W.M., and R. A. Laskey. 1974. A film detection method fortritium-labeled proteinsand nucleic acids in polyacrylamidegels.Eur.J.Biochem. 46:83-88. 3. Byrt, P., and G. L. Ada. 1969. An in vitro reaction
betweenlabeledflagellinorhaemocyaninand
lympho-cyte-like cells from normalanimals.Immunology17: 503-516.
4. Cassai,E., R.Manservigi,A.Corallini,and M.Terni. 1975/76. Plaquedissociation of herpes simplex viruses: biochemical andbiologicalcharacters ofthe viral var-iants.Intervirology 6:212-223.
5.Cleveland, D. W., S. G. Fischer, M. W. Kirschner, andU. K. Laemmli. 1977.Peptide mapping bylimited proteolysis insodium dodecyl sulfate and analysis by gel electrophoresis. J. Biol. Chem.252:1102-1106. 6. Cohen, G.H.,M.Katze,C.Hydrean-Stern,and R. J.
Eisenberg. 1978.Type-commonCP-1antigenofherpes simplex virus is associated with a 59,000-molecular-weight envelopeglycoprotein. J. Virol.27:172-181. 7. Costa, J.C.,and A. S.Rabson. 1975.Roleof Fc
recep-torsinherpessimplexvirusinfection. Lanceti:77-78. 8. Costa, J., A. S.Rabson,C. Yee, and T. S.Tralka. 1977.Immunoglobulin binding to herpes virus-induced Fcreceptors inhibits virus growth. Nature (London) 269:251-252.
9. Costa,J., C. Yee, Y.Nakamura, and A. Rabson. 1978. Characteristics of the Fc receptor induced by herpes simplex virus.Intervirology 10:32-39.
10.Fairbanks, G.,T.L.Steck,and D. F. H.Wallach.1971. Electrophoreticanalysisof themajorpolypeptideof the humanerythrocytemembrane.Biochemistry 10:2606-2617.
11.Feorino,P.M.,S. L Shore,and C. B. Reimer. 1977. Detectionby indirectimmunofluorescenceof Fc recep-tors incells acutely infected with herpes simplex virus. Int. Arch.AllergyAppl.Immunol.53:222-233. 12.Heine,J.W.,R. W.Honess,E.Cassai,and B.
Roiz-man.1974.Proteinsspecified by herpes simplexvirus. XII. The virionpolypeptidesoftype 1 strains. J. Virol. 14:640-651.
13.Hubbard, A. L., and Z. A. Cohn. 1972. The enzymatic iodination of the red cell membrane. J.CellBiol.55: 390-405.
14.Jensenius, J.C.,and A. F.Williams.1974.The binding ofanti-immunoglobulin antibodies to rat thymocytes and thoracic ductlymphocytes.Eur.J. Immunol. 4:91-97.
15. Keller, R., R.Peitchel,J. N.Goldman,and M. Gold-man. 1976.AnIgG-Fcreceptorinducedin cytomega-lovirus-infected human fibroblasts. J. Immunol. 116: 772-777.
16. Lehner, T.,J. M. A.Wilton,and E. J.Shillitoe.1975. Immunological basis for latency, recurrences, and pu-tative oncogenicity ofherpessimplexvirus. Lancet ii: 60-62.
17. Manservigi,R., P. G.Spear,and A.Buchan. 1977. Cell fusion inducedbyherpessimplexvirus ispromoted and suppressed bydifferentviralglycoproteins. Proc. Natl. Acad. Sci. U.S.A. 14:3913-3917.
18. McTaggart, S. P.,W. H.Burns,D.0.White,and D. C. Jackson. 1978. Fc receptors induced by herpes simplexvirus. I.Biologic and biochemical properties. J. Immunol. 121:726-730.
19. Nakamura, Y., J.Costa,T.S. Tralka, C. L. Yee, and A.S. Rabson.1978.Propertiesofthecellsurface Fc-receptor inducedby herpessimplexvirus.J.Immunol. 121:1128-1131.
20. Nisonoff, A.1964.Enzymatic digestion of rabbit gamma globulin andantibody and chromatography of digestion products. Methods Med. Res. 10:134-141.
21. Rahman, A. A., M. Teschner, K. K. Sethi, andH. Brandis. 1976.AppearanceofIgG(Fc)receptor(s) on culturedhumanfibroblasts infected with human cyto-megalovirus. J. Immunol. 117:253-258.
22. Ruyechan,W.T., L. S. Morse, D. M. Knipe, and B. Roizman. 1979.Molecular genetics ofherpes simplex virus.II.Mapping ofthemajor viralglycoproteins and ofthe genetic loci specifying the social behavior of
on November 10, 2019 by guest
http://jvi.asm.org/
789
infectedcells. J. Virol. 29:677-697.
23. Sakuma, S., T. Furukawa, and S. A.Plotkin. 1977. Thecharacterization of IgGreceptor induced by human cytomegalovirus. Proc. Soc. Exp. Biol. Med. 155:168-172.
24. Sarmiento, M., M. Haffey, and P. G. Spear. 1979. Membrane proteins specified by herpes simplex viruses. III. Role of glycoprotein VP7(B2) in virion infectivity. J. Virol.29:1149-1158.
25. Sarmiento, M., and P. G. Spear. 1979. Membrane pro-teinsspecifiedbyherpessimplex viruses. IV. Confor-mationof the virion glycoprotein designatedVP7(B2). J. Virol. 29:1159-1167.
26. Shimizu, Y. 1971. Modification of host cell membrane after herpessimplex virus infection. Arch. Gesamte Virusforsch.33:338-346.
27. Spear, P.G. 1976. Membrane proteins specified by herpes simplexvirues.I. Identification of fourglycoprotein
precursors and their products in type1-infectedcells.J. Virol.17:991-1008.
28. Watkins,J. F. 1964.Adsorption of sensitizedsheep eryth-rocytes to HeLacells infected with herpessimplexvirus. Nature(London) 202:1364-1365.
29.Watkins, J. F. 1965. The relation ofherpes simplex haemadsorptionphenomenontothe virusgrowth cycle. Virology26:746-753.
30. Westmoreland,D., S. St. Jeor, and F.Rapp.1976.The development bycytomegalovirus-infectedcells of bind-ing affinity fornormalhumanimmunoglobulin.J. Im-munol. 116:1566-1570.
31. Westmoreland, D.,and J.F. Watkins. 1974. TheIgG receptor induced by herpessimplexvirus: studiesusing radioiodinatedIgG.J. Gen. Virol. 24:167-178. 32. Yasuda, J.,and F.Milgrom. 1968.Hemadsorptionby
herpessimplexinfectedcellcultures. Int. Arch.Allergy
33:151-170. VOL. 32,1979
on November 10, 2019 by guest
http://jvi.asm.org/