JOURNALOFVIROLOGY, Jan.1978,p.374-383 Copyright © 1978 American Society forMicrobiology
Vol. 25, No. 1 Printed in U.S.A.
Synthesis
and
Processing
of
Precursor
Polypeptides
to
Murine
Mammary Tumor
Virus Structural
Proteins
JANIS RACEVSKIS* andNURUL H. SARKAR
Memorial Sloan-Kettering Cancer Center, New York, New York 10021
Received for publication24June1977
Biosynthesisofmurinemammary tumorvirus (MuMTV) proteinswasstudied
inthechronicallyMuMTV-infectedepithelialcellline MuMT-73by using
mon-ospecificantiseratothemajorMuMTVcoreproteinp27 and themajorenvelope
glycoproteingp47. Inpulse-labeling experiments using[35S]methionine,
monospe-cificantiseratop27precipitateda75,000-molecular-weightproteinasthemajor
intracellularcomponent.Analysis of thesamecellularextractswithmonospecific
antiserato gp47 revealed that the gp47 precursor was a 70,000-dalton protein. After chaseperiods,therewas aloss of label from theprecursorsanda
concom-itant increase oflabeled extracellular mature viral proteins. The glycoprotein
precursorincorporated labeled glucosamine and seemed to be processed more
rapidlythan the p27precursor. Considerableamounts ofapparently
nonvirion-associated gp47 andglycoproteinprecursorcould be detected in the extracellular culture fluid.
Themurinemammary tumorvirus(MuMTV) isatype BRNAtumorviruscomposed offive major structural polypeptides (6, 11, 18, 25,30)
andsomeadditionalminorpolypeptides,
includ-ing the reverse transcriptase (14). Two of the five majorpolypeptidesareglycoproteins (gp47 andgp34), which are associated with the viral membrane (20),whereasthe other three major
polypeptides (p27, p16, and p12) are core
con-stituents (20). (Inour gelsystem, which shows
alinear rangefor calibration proteins between molecular weights 25,000 and 200,000 on a log
scale,wefind that themajorMuMTV
glycopro-teinmigratesas awide bandjustbehind cellular
actin [molecular weight 42,000] in a molecular weight range of between 45,000 and 50,000.) Mostof theinvestigationsinto thestructureand
composition of MuMTV have been done by
usingvirus isolated from the milk of mice from
strains withahighmammary tumorincidence.
However, thesuccessfulcultivation of
MuMTV-producing cell lines (13, 15, 19, 25) has made possible the study of intracellular viral protein
synthesis, processing,andassembly.
A number of laboratories have established
that the structural proteins of type C RNA tumorvirusesare firstsynthesizedas high-mo-lecular-weightprecursors that aresubsequently cleavedandprocessedtogiverisetothemature viralproteins(9, 24, 27).Thesestudies havealso indicatedthat themajorviralglycoprotein and
majorinternalcoreproteinsof thetypeC viruses
are initially synthesizedas part oftwo distinct
separate precursors,referredto astheenvand
gaggeneproducts (1), respectively.Tworecent reports provide evidence suggesting that the
synthesisoftype B RNAtumorvirus proteins
mightoccurinananalogous fashiontothose of typeC viruses.By usingananti-MuMTVserum,
Dickson and co-workers (5) reported a
73,000-dalton precursor to MuMTV glycoproteins in
extracts of
primary
cell cultures derived from murine mammary gland tumors. The proteinsynthesis inhibition studies carried out by
Schochetman and Schlom (22), usinga MuMTV-producing cellline, haveprovided evidence for
thenoncoordinate synthesis of the glycoproteins and thecoreproteinof MuMTV.
In thispaper wepresent acharacterizationof
theintracellular synthesis of MuMTVproteins
and the identification of theprecursor polypep-tides of the MuMTV structural proteins gp47
and p27byusing MuMTV-producing epithelial celllineMuMT-73. These studieswere carried
outby the application ofimmunoprecipitation
techniques by usingmonospecific antiseratothe major MuMTVglycoproteingp47 andthe major
coreproteinp27.
MATERIALS AND METHODS
Cell cultures. Thesourceand characterization of theepithelialcelllineMuMT-73, which continuously produces MuMTV, have been described previously (19). The cellswerepropagated inasolution of Eagle minimumessential medium supplemented with 10% (vol/vol) fetal calf serum, 100 U of penicillin per ml and 100 ug ofstreptomycin per ml. The Rauscher virus-infected JLSV-9 cells were a gift from P. J. Gomatos of thisInstitute.
374
on November 10, 2019 by guest
http://jvi.asm.org/
Labeling of cells and extraction ofproteins. Cellswere labeled a day afterchanging the growth medium in cultures that had grown toconfluence in 75-cm2T flasks. The celllayerwaswashedoncewith
5mlofwarmmethionine-deficient minimum essential medium and then incubated withanadditional7ml of thesamemediumfor1h. Afterthispreincubation period, the mediumwaspouredoff andreplacedwith 3 ml of methionine-free medium containing 200yCi of[3S]methionine (NewEnglandNuclear).After the pulseperiod, eitherthecellswereimmediately lysed orthelabelwaschasedbythe addition of 0.5 ml of 1 mMmethionine ingrowthmediumplus0.4ml of fetal calfserum.
Cellular proteins were extracted according tothe method ofShapiro andAugust (23).Inthismethod, the cellsarewashed three times with ice-cold Earle balanced salt solution and thenscrapedinto2ml of
anextractionbuffercontaining5mM Tris-hydrochlo-ride (pH9.2),1mMEDTA,400mMKCI, 1% Triton X-100, 1 mM TPCK
(L-1-tosylamido-2-phenylethyl-chloromethyl ketone),and1mMPMSF (phenylmeth-ylsulfonyl fluoride). The protease inhibitors TPCK andPMSFwereobtained fromSigmaChemical Co. and added to the extraction bufferjust before use
from a 100mMstocksolution made up indimethyl sulfoxide. Theextracts werecentrifugedat25,000xg for 10min, and the pellets werereextracted with 1
ml ofKCl-freeextractionbuffer.Theywerethen
cen-trifugedat 25,000xgfor 10min.Thesupernatants
were pooled and centrifuged at high speed (1 h at
100,000 xg)beforeimmunoprecipitation.
Forlabelingof cells with[3H]glucosamine, growth medium fromaconfluent culture ina75-cm2Tflask
wasreplacedwith 10 ml of minimumessential medium containing25% of the normal concentration ofglucose and supplemented with 50
ACi
of [3H]glucosamine(NewEnglandNuclear)perml. The[3H]glucosamine
solution was concentrated by evaporation under a stream ofnitrogen toreduce the amountof ethanol (fromtheglucosaminesolution) addedtotheculture medium. Cellswereincubated for 24 h and then
ex-tracted asdescribed above.
Inexperimentswhere the extracellularculture fluid
wasanalyzedfor the presence of virion- and nonvirion-associatedproteins,thegrowthmediumwasclarified bylow-speed centrifugation (10minat 500xg) and then layered onto a discontinuous sucrose gradient made up of1 ml of 60% and 3ml of 20% sucrosein TEN buffer (20 mM Tris-hydrochloride [pH 7.4]-1
mMEDTA-100 mM NaCl) and spun for90min at
38,000 rpm inanSW41rotor.Thesupernatantculture fluid was removed, and Triton X-100 and sodium deoxycholatewereaddedtoafinal concentration of1
and0.5%, respectively. Virusatthe interface of the discontinuous gradient was removed, diluted with
TENbuffer,anddisruptedbytheadditionof Triton X-100sodiumdeoxycholatetoconcentrations of1and 0.5%,respectively.
Immunoprecipitation. The antisera used were
monospecific goat anti-p27 and anti-gp47 sera-(21) and rabbit anti-wholeMuMTVsera.Thepreparation andadsorption of antiserahave been described(18). Rabbitanti-LETS antiserumwas agiftfrom Lan Bo Chen of ColdSpringHarborLaboratory. Portions of
0.5to1.0 mlofcentrifuged cell extracts were incubated
at 4°C with 3 to 8,ul of antisera, depending on the titer ofthe antiserum. After 20min, 20to40
pld
ofa10%suspensionofstaphylococcal proteinA-antibody adsorbent(10)wasadded andincubated for an addi-tional 20 min at 4°C. Staphylococcus aureus strain Cowan I waskindly provided by H. Oppermann of theUniversityofCalifornia,SanFrancisco. The prep-aration of thestaphylococcal antibody adsorbent and theisolation of theimmunoglobulins were carriedout
accordingtotheprocedure of Kessler (10). The follow-ing modifications to the techniques of Kesslerwere
introduced toreducenonspecific cellular contamina-tionin theimmunoprecipitates: the washed staphylo-cocciweresuspended toa10%suspension in unlabeled cellextract, and cell extracts were preadsorbed once
withcontrol serum andstaphylococcaladsorbent be-foreimmunoprcipitation. After the 20-min incubation withstaphylococcal adsorbent,thesuspensionswere layeredon 1 mlof1 Msucrose-0.5% sodium deoxy-cholate TEN solution and centrifuged at 3,000 rpm for 10min. Theadsorbent antibody pellets were then washed threetimes with0.5% NP-40 TEN buffer. The antigen-immunoglobulin complexes were dissociated from theadsorbentby heating at 100°C for 3minin 50
pil
of 1.5x concentrated electrophoresis sample buffer(12).Polyacrylamide gel electrophoresis. Gel elec-trophoresis was done in 20-cm-long, 5 to 20% exponen-tialgradient sodium dodecyl sulfate (SDS)-polyacryl-amide slab gels by the discontinuous Tris-glycine buffer system of Laemmli (12). The exponential gra-dientpolyacrylamide slab gels were prepared as de-scribed by Van Blerkom and Manes (26). Protein molecularweights were determined with the Combi-thekmolecular-weightmarker kit(Boehringer Mann-heim), which includes soybean trypsin inhibitor (21,-500), bovine serum albumin (68,000), and Escherichia coli RNApolymerase subunit a (39,000), subunit ,B (155,000), and subunit if (165,000). Myosin heavy chain (molecular weight 200,000) was also used as a marker. Inaddition, a mixture of [3S]methionine-la-beledvesicular stomatitis virus- and reovirus-infected cell extracts was used as amolecular-weight marker on the autoradiograms, as previously described by Racevskis and Koch (16). [3H]amino acid- or ['4C] amino acid-labeled MuMTV marker virus was pre-paredasdescribed by Sarkar et al. (20). After electro-phoresis,gels were treated by the procedure of Bonner and Laskey (2), dried, and then exposed to Kodak RR Royal X-Omat film at -700C.
RESULTS
Immunoprecipitation of intracellular
viralprecursorproteins. The MuMT-73
ep-ithelial cellline, derivedfromspontaneous
mam-marytumorsofBALB/cfC3H mice (19),which continuously produces MuMTV (B particles),
was used throughout these studies. Although
virusproductioncanbeincreasedinthe
MuMT-73 cells by glucocorticoid hormone treatment (19), the studies described in this report were
doneby using untreated culturessoon after they
on November 10, 2019 by guest
http://jvi.asm.org/
376 RACEVSKIS SARKAR
reachedconfluency andadayafter fresh media
had beenadded. Since these epithelial cellsare
fairlyslow-growing cells, and the viral proteins
account for only avery small fraction of total cellular protein synthesis, itwasfoundnecessary
to pulse-label cells for periods of30 to 60 min
with about
200,uCi
of[3S]methionineper T75flasktoeasily detectviral-specific polypeptides intheimmunoprecipitates.
By using monospecific antisera tothe major glycoprotein gp47and major internalcore
pro-teinp27, wedetectedtwo closely migratingyet
distincthigh-molecular-weightintracellular
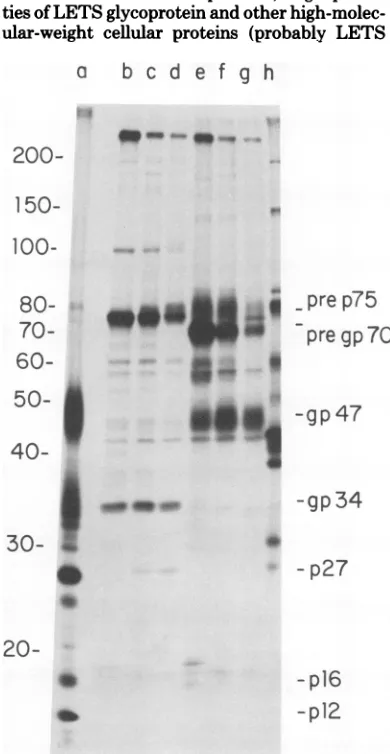
pro-teinsbeing specifically immunoprecipitated with the antisera (Fig. lb and c). Figure lb showsa
60-min [3S]methionine-labeled MuMT-73 cell
extractprecipitatedwithanti-gp47serun.This
serumprecipitatedapolypeptideof70,000
mo-lecularweight (pre gp7O). Some gp47 was also precipitated,ascan beseenbycomparing itto
the [3H]amino acid-labeled marker MuMTV pattern in column a.Figure lcshows thesame extract asin lb, but immunoprecipitated with anti-p27serum,which,as canbeseen, detected
a75,000-daltonspecies (pre p75).For
compari-son purposes, Fig. ld and e are 60-min
[35S]_
methionine-labeled cellextractsof Rauscher
vi-rus-producing JLS-V9 cells precipitated with anti-Rauscher p30and gp69/71 serum,
respec-tively. Figure ld shows the 67,000-dalton
pre-cursor to p30, and Fig. le shows the
80,000-dalton glycoprotein precursor, as well as the envelope glycoprotein gp69/71, which
comi-grateswith the MuMTV glycoprotein precursor.
Analysisof the MuMT-73cellextractwith
anti-Rauscher virus antisera showed no detectable viralproteins beingprecipitated (not shown).
Proteins of cellular origin. In both the MuMT-73 and the JLS-V9cellextracts(Fig. 1),
a very high-molecular-weight species (250,000
daltons) wasprecipitatedwithall the different antisera. There was less of thisprotein in the MuMT-73 cellextractsthan in the JLS-V9 ex-tract because the MuMT-73 extracts were
preadsorbedwithcontrolgoat serumand
staph-ylococcal antibody adsorbent before
immuno-precipitation, as described above. We found it necessary to preadsorb the MuMT-73 cell ex-tractssincetheseepithelialcell linessynthesize
great quantities of cytoskeletal-type proteins,
whichinterferewith theresolutionof
viral-spe-cificpolypeptides.Twoadditional high-molecu-lar-weight proteins are seen in the nonpread-sorbed JLS-V9cell extracts.
Anotherprominent proteinband that is pres-ent in all immunoprecipitates in variable
amounts is a species of about 42,000 daltons. Thecellular originof the42,000-dalton protein
M
UMTV
pre
p(75bm
-_r~~~~~~0
pre
gp(C.gp47
.--,1
C
b
c,
d eR
LV
*
-pregp8O
;
-gp69/7l
-Lc,r)e p67
*--
(1ct
riFIG. 1. Autoradiograph of SDS-gel electrophore-sisofimmunoprecipitatesfrom MuMT-73cellsand JLS-V9cellspulse-labeledwithf5SJmethionineand
run on a20-cm-long5 to20% exponentialgradient polyacrylamide gel. (a)
I3H]amino
acid-labeled MuMTV marker virus. (b) 60-min f5S]methioninepulse-labeledMuMT-73 cellextract immunoprecipi-tated withanti-MuMTV-gp47serum.(c) Sameas(b)
but immunoprecipitated with anti-MuMTV-p27
se-rum. (d) 60-min
f6S]methionine
pulse-labeled JLS-V9cellextractimmunoprecipitated withanti-RLV-p30serum. (e) Same as(d) but immunoprecipitated withanti-RLV-gp69/71serum.
and the nonspecificity of the precipitation is indicated by the observation that the quantity
present in the immunoprecipitates is inversely
proportional tothe extent that the precipitates
are washed. This cellular protein is probably actin, which has been estimated to make up some5to10% of theprotein of nonmuscle cells
(3) and identified as being present in purified preparations ofseveral different enveloped vi-ruses (28). Onthebasis of thereport that
non-muscle actinshave thesamemobility on SDS-acrylamide gels as muscle actin (3), we have
identifiedthe42,000-daltonproteininour
prep-aration as actin bythe observation that it
co-migratedwith mouseskeletal muscle actin(Fig. 2b). MuMTV purified from RIII mouse milk
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.271.458.72.356.2]a
b
-
myosin
gp47-
*- -
actin
p27-FIG. 2. Coomassie brilliant blue-stained
SDS-polyacrylamide gel. (a) Purified RIII mouse milk MuMTV. (b)RIIImouseskeletalmuscleextract.
alsocontainedaprotein bandwhichcomigrated with actin extracted from RIII mouse skeletal muscle(Fig. 2a).
Specificityofthe antisera. The
monospec-ificity of the antisera used in this study was demonstrated by Sarkarand Whittington (21) by the application of the double antibody tech-nique against purified ['4C]amino acid-labeled MuMTV. Wehaverepeated the testfor speci-ficity by employingthestaphylococcal antibody adsorbenttechnique against purified [3S]methi-onine-labeled MuMTV(Fig.3c).Goat antiserum against p27 precipitated only p27 (Fig. 3a), and goat antiserum against gp47 precipitated only gp47 (Fig. 3b) when tested against [3S]methio-nine-labeled, purified MuMTV (Fig. 3c). It is
also quite clear from Fig. 3d and e that the
intracellularviralprotein species designatedas
prep75 andpregp7O precipitatedfrom 90-min
pulse-labeled cell extracts by each antiserum
werequite distinct and thatnocross-reactivity
was observed. The anti-p27 serum also seems tohaveprecipitatedaless prominent protein of about 100,000 daltons (Fig. 3d), whichwas not
observed in the sample precipitated with
anti-gp47 serum (Fig. 3e). Analysisof the same la-beled cellextractwith nonimmune goat serum
(Fig. 3f) reveals that only the
high-molecular-weight (250,000 daltons) species was
precipi-tated. No other prominent species were ob-served.
Identification of the 250,000-dalton
cel-lular protein. On the basis of a number of
observations, includingmolecularweight,release
into tissue culture medium, and labeling with glucosamine, wesuspected that the large,
250,-000-molecular-weightprotein observed in all the
immunoprecipitates might be thelargeexternal
transformation-sensitiveglycoproteinknownby
theacronym LETS(7).Immunoprecipitationof the
[3S]methionine-labeled
cell extract with anti-LETS antiserum (Fig. 3g) confirmed oursuspicion. All theimmunoprecipitations shown
in Fig. 3 d (anti-p27), e (anti-gp47), f (control serum), andg(anti-LETS)weredone with iden-tical aliquots of the same [3S]methionine-la-beled cellextractusing identical volumes of the respective antiserum andstaphylococcal
adsorb-ent.Toquantitate theamountof
high-molecu-lar-weight (250,000 dalton) protein in each of
the above immunoprecipitates, densitometer
scans of the top half of these four lanes (d
through g) were done (Fig. 4). The amount of
250,000-dalton protein in the sample
immuno-precipitatedwith anti-LETSserum(Fig.4a)was
abouteightfoldgreater than that inany oneof the other three samples (Fig. 4b through d),
thusestablishingitsidentityasLETS
glycopro-tein. It may also be noted that in additionto
the LETS glycoprotein, anti-LETS serumalso
brought down many other
high-molecular-weight proteins (Fig. 3g). This observation is
not surprising in view ofthe fact that LETS
has been shown to be extensively disulfide bondedtomanyotherproteinsonthe
mamma-lian cell surface (8). Since the cell extraction
andimmunoprecipitation arecarriedoutunder
nonreducingconditions, thedisulfide bonds
be-tweenLETSandothercellularproteinsremain
intact,resultingin thecoprecipitationof various LETSaggregates.
Pulse-chase studies-kinetics of
precur-sorprocessing.Theresultsofpulse-chase
ex-periments usingthe MuMT-73 cell lineare
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.78.215.66.450.2]378
a b c d e f g
200
-100
-
80-70
-60
-50
-40
-30
-a -a m
#
-LETS
.
rpre
p75
doft
-Lpre
gp7O
-
gp47
-
gp34
[image:5.505.151.382.68.366.2]-
p27
FIG. 3. Analysis of antiserum specificity. (a)P5S]methionine-labeledpurifiedMuMTVimmunoprecipitated with monospecific anti-gp27serum. (b) Same as (a) but immunoprecipitated with monospecific anti-gp47 serum. (c) 65S]methionine-labeledpurified MuMTV. (d) 90-minPS]methioninepulse-labeledcell extract immunoprecipitated with anti-p27 serum. (e) Same as (d) but immunoprecipitated with anti-gp47 serum.
(f)
Same as (d)butprecipitatedwith controlnonimmune goat serum. (g) Same as (d) but immunoprecipitated withanti-LETS serum.
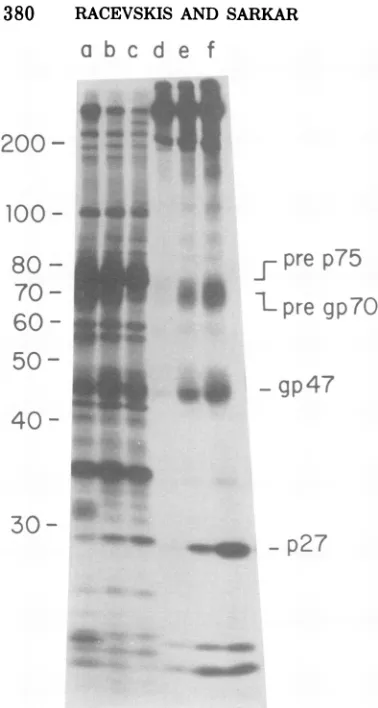
picted in Fig. 5 and 6. Figures 5b through d
show immunoprecipitates obtained with
anti-p27 sera incellextractsof60-min
[35S]methio-nine-pulsed and 90- and 180-min-chased
cul-tures, respectively. It canbeseenthat the
pre-sumed75,000-molecular-weight polypeptide pre-cursorofp27isfairly stable and thata consid-erablequantityofprecursoris stillpresentafter
a chaseperiod of3h.Inotherexperiments, we
have found that the precursor protein is still
detectable after a chase period of 6 h. During thechaseperiod,the pre p75appearedtodisplay
aslightlyloweredmobilityintheSDS-gel (Fig.
5candd). Wehave found that thisslowed
mi-gration of pre p75 after the chase periods is caused by a post-translational modification of theprecursorinvolving phosphorylation (N.H.
Sarkar, E. S.Whittington, J.Racevskis, andS. L. Marcus, manuscript in preparation). The
100,000-molecular-weight protein,whichis
con-siderablylessprominentthan prep75and seems to be p27 specific, is chased as well (Fig. 5b
through d).Amoreprominentband in theregion
of 34,000daltons ispresentin allthreesamples shown inFig.5bthrough d andmightrepresent anintermediate precursor of p27. The amount
of intracellularp27 isbarelydetectable.Analysis ofadditionalportions ofthe same cellextracts
withanti-gp47serumrevealed that the glycopro-teinprecursor pregp7Oisprocessedmorerapidly
than prep75,and thatlargequantities ofmature
gp47 are present intracellularly after a60-min
pulse and after ensuing chase periods (Fig. 5e
through g). Some protein speciesthat seem to be gp47 specific are visible in the molecular-weight region of about 60,000 (Fig. 5e through
g). The fact that theseproteins decreasein con-centration after chase periods and display smearing, which is characteristic of
glycopro-teins electrophoresed on SDS-gels, indicates
thattheymay beprocessing intermediatesfrom pregp7O.Thediffuse dark arearunningbehind pre gp7O (Fig. 5e) mightrepresent fuly glyco-sylated speciesoftheprecursor(Fig. 7k).These
species, however, make up only a very small
fraction of total pregp70, asshownbythe
on November 10, 2019 by guest
http://jvi.asm.org/
lected and likewise immunoprecipitated with whole MuMTV antisera (Fig. 6d through f). After a90-minchaseperiod (Fig. 6e),five viral proteins were recovered from the media: the majorglycoprotein gp47, the majorcoreprotein
p27, twosmall viral proteins of less than 20,000
molecularweight, and one diffuseband that
co-migrated with glycoprotein precursor pre gp7O. Inadditiontothese viralproteins, large
quanti-ties of LETS glycoproteinand other
high-molec-ular-weight cellular proteins
(probably
LETSa
b c d e f
g
h
200-
150-
100-
80-
70-
60-
50-200 100 80 70
FIG. 4. Densitometerscansofthe upper portion of columnsd, e,f,and gofthegelshown inFig.3. (a) Column (g) of Fig. 3. (b) Column (D of Fig. 3. (c) Column(e)ofFig.3.(d)Column(d)ofFig.3. Approx-imatemolecularweightsin kilodaltonsareshownat
thebottomofthefigure.
sitometer scan (Fig. 4c). The glycoprotein
het-erogeneityisespeciallynoticeable on these
20-cm-long high-resolution
gradient
gels,
and itmay be noted that gp34 of the marker virus (Fig. 5a) isactuallyaheterogeneouspopulation ofsomethreeorfour discrete bandsrunningin
amolecular-weightrangeof between32,000and
37,000. Wecanalso seefromFig. 5thatLETS
proteinis decreased in concentration after the
chase periods, indicating that it is
probably
transportedtotheoutersurface of thecell
mem-branefrom where it isshedinto themedia. Extracellulartransportof viralproteins. Analysis of anti-MuMTV immunoprecipitates obtainedfromextractsofcellspulsedfor60min
and eithernotchasedorchasedfor either90 or 180 minis shown in Fig. 6a throughc. Allthe
species precipitated with the individual
mono-specificantisera inFig.5 arealsorecognized by
the antisera to disruptedwhole MuMTV. The
culturemedia from the three cultureswere
col-40-
I
was fI
3-,-ma
m-30-9
20-FIG. 5. Autoradiograph of SDS-gel
electrophore-sis of immunoprecipitates frompulse-chase studies
in MuMT-73 cells. (a) '4C]amino acid-labeled
MuMTVmarker virus. (b) 60-min f5SJmethionine
pulse-labeled cell extract immunoprecipitated with
anti-p27serum.(c)Sameas(b)butchasedfor90min
with excess cold methionine. (d) Same as (b) but chased 180min. (e)Sameas(b) but
immunoprecipi-tated withanti-gp47serum.(/)Sameas(e) but chased
for90min. (g) Sameas (e) but chasedfor 180min.
(h)Mixtureofvesicularstomatitis virus-and
reovi-rus-infected cell extracts labeled with f5SJmethio-nine(molecular-weight markers).
a LETS
b
C pregp7O
d prep75
A
pre
p75
pre
gp70
-g
p
47
-gp34
S
-
p27
-p16
-pl2
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.84.216.62.392.2] [image:6.505.252.447.161.538.2]a
b
c
d e f
200
-100 - etoe
80-70--
*
60
-^2
50-40-
*430
-pre
p75
1-pregp7O
gp47
"N i"4U
p27
FIG. 6. Polyacrylamide gelelectrophoresis analy-sis of the immunoprecipitates oftheMuMT-73 cell extractsand extracellular media reacted with
anti-whole MuMTVantisera. (a) 60-minI5SJmethionine
pulse-labeledcellextractsimmunoprecipitated with
anti-MuMTVserum. (b)Sameas(a), but cellswere
chasedfor90 minwith excess coldmethionine. (c)
Same as (a), butcellswerechasedfor 180min. (d) Cell-free culture media from same cells as in (a) immunoprecipitated with anti-MuMTV serum. (e)
Culturemediafrom 90-min-chasedcells immunopre-cipitated withanti-MuMTVserum.(f) Culturemedia
from 180-min-chased cellsimmunoprecipitated with anti-MuMTVserum.
aggregates) were found adhered to the immu-noprecipitates (Fig. 6d through f). Two factors that probably contributed to the high level of these cellular contaminantswere(i)the cell-free
mediainthe experimentwerenotpreadsorbed before immunoprecipitation, and (ii) relatively large volumes ofanti-MuMTV sera as well as staphylococcal adsorbentwere used to precipi-tateviralproteinsfromthe media.
Virion- and nonvirion-associated extra-cellular viral proteins. Inview ofthe
unex-J. VIROL.
pected detection, with anti-MuMTV sera, of what seems to be pre gp7O in cell-free media (Fig. 6e and f), we attempted to identify the
origin of the pre gp70 and determine whether
the other extracellular viral proteins were, or were not, virus associated. Cells were labeled
with [3S]methionine in media containing
one-twentieth thenormal concentration of methio-nine for12h and thenchased foranadditional
12 hby the addition ofexcess,cold methionine.
The media from these cultures was collected
andlayeredontopofa20to60%discontinuous
sucrosegradient andcentrifugedat100,000 x g for 2 h. The remaining virus-free media were
saved, and the viral fraction at the 20 to 60% interface was removed and treated with lysing buffer. Immunoprecipitation of thecell extract
with anti-p27 and anti-gp47 sera is shown in Fig. 7b and c,respectively. Itcanbe seenthat thepregp7O andprep75precursors werebarely detectable after the long chase period. There
was a verysmallamountof p27 inthe cell (Fig. 7b);however, largeamounts ofgp47 were pres-ent(Fig.7c). It should be noted thatnolabeled cell-associated LETSproteinwasobserved after the long chase period. Immunoprecipitation of the virus-free media withanti-p27sera (Fig. 7d) revealed thatsomenonvirion-associatedp27 was presentinthe mediatogether withaprotein of less than 20,000 molecular weight. Analysis of thesame mediawithanti-gp47 serum (Fig. 7e) revealedthat, in additionto alarge amountof nonvirion-associated gp47, precursor gp7O was
also present in the media. The
low-molecular-weight proteindetected withanti-p27serum was
also observed. This protein does notcomigrate withanyof thesmallviral proteins in the marker MuMTV (Fig. 7n) and probably represents a
cellularspecies whichmaybeabreakdown
prod-uct or anassociated protein ofthe
high-molec-ular-weight cellular contaminants. Ananalysis
of the viral fraction withmonospecific antisera revealed the presenceof p27 and gp47 (Fig. 7f
andg,respectively).In contrast tothe virus-free
media, nopre gp7Owasfound in the viral
frac-tion. Some p27 was unexpectedly observed in
the viral fraction precipitated with anti-gp47 serum (Fig. 7g). We have observed that when
immunoprecipitation is done in concentrated
viral extracts, some p27 will be present in the precipitates unless theyarewashedextensively
andcentrifugedthroughasucrosedeoxycholate
solution asdescribedabove. Thesampleshown inFig.7g was notcentrifugedthroughasucrose
deoxycholate solution. The p27 probably
ad-herestothestaphylococcusadsorbentsincep27 wasalso detected insamples precipitated with nonimmune sera, which were not washed
thor-oughly. We have observed that Rauscher
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.67.256.49.403.2]200
-a b c d e f g h j k I m n
*4.0
150
-80-
- 70-60- w
50-
40-* -p27
20
-% -p16
- - p12
FIG. 7. Long-term labeling of MuMT-73 cells with f'S]methionine andI3H]glucosamine. (a) Mixtureof vesicular stomatitis virus- and reovirus-infected cell extracts, P3S]methionine labeled (molecular-weight markers). (b) MuMT-73 cell extract labeledfor 24 h with f35S]methionine andimmunoprecipitated with anti-p27serum. (cJ Sameas (b) butimmunoprecipitated withanti-gp47serum. (d)Mediafromcells in (b) after spinningoutvirus andimmunoprecipitatedwithanti-p27serum.(e)Sameas(d),butimmunoprecipitated
withanti-gp47serum. (f) Spun down, solubilized virus isolated from media of 24-h[fS]methionine-labeled
cellsimmunoprecipitated with anti-p27serum.(g) Sameas(f)butimmunoprecipitated with anti-gp47serum.
(h) MuMT-73 cell extract labeledfor 24 h with P3Higlucosamine and immunoprecipitated with anti-p27
serum.(i) Sameas(h)butimmunoprecipitatedwithanti-gp47serum.
OF)
Virus-freemediaofsamecellsasin(h), immunoprecipitatedwithanti-p27serum. (k)Sameas(j) butimmunoprecipitated withanti-gp47serum.
(1)pH]glucosamine-labeledsolubilized virusisolatedfrommediaofsamecellsasin(h),immunoprecipitated withanti-p27serum.(m)Sameas(1) butimmunoprecipitatedwithanti-gp47serum.(n) "4C-labeledMuMTV
marker virus.
kemia virus p30 also hasan affinity to staphy-lococcus adsorbent (Racevskis and Sarkar, un-published data). LETS glycoproteinis present in the media as well as in the virus samples. Additional, higher-molecular-weight bands are seeninthe viralsamplesas well (Fig. 7f and g).
Glucosamine labeling of viral proteins. Tofurthercharacterize and identify the glyco-sylated viralproteins, [3H]glucosamine labeling of the MuMT-73 cells was done. Cells were labeled for 24 h with[3H]glucosamine in media containing 25% of the normalamountofglucose. Cellswereextracted,mediawerecollected, and viruswasspunoutasdescribedabove.Analysis of the three fractions-cellular extracted (Fig.
7i), virus-freemedia (Fig.7k),and virus fraction (Fig. 7m)-with anti-gp47 serum shows that boththe70,000-molecular-weightprecursorand gp47 were heavily glycosylated and that the glycoproteinsinthe various fractionswere iden-ticaltothoseseenpreviouslywith the [3S]me-thionine-labeled cells. After 24 h oflabeling in low-glucose media,large quantitiesofgp47were present, but not much intracellularpregp70was observed (Fig. 7i).The virus-free medium (Fig. 7k) contains large quantities ofprecursor and matureglycoprotein,whereas the virus fraction (Fig. 7m) contains only mature glycoprotein gp47 associated with it. As seen in Fig. 7h, j, and 1, anti-p27 serum did not precipitate any
30- l _
-pre gp70
-
gp47
-
gp34
I
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.505.128.370.72.367.2][3H]glucosamine-labeled proteins from any of
the three fractions, except for LETS protein,
which isglycosylated.
DISCUSSION
By the application of immunoprecipitation techniques using monospecific antisera and high-resolution polyacrylamide gels, we have found that the MuMTVmajorglycoproteingp47
andmajorcoreprotein p27areinitially synthe-sized as part oftwo separate high-molecular-weightprecursorpolypeptides (pre gp7O andpre p75,respectively).
Dickson et al. (5) described the detection of
a 73,000-dalton precursor to the two MuMTV glycoproteins (gp47 and gp34) by the use of antiserumfor whole MuMTV inpulsedcell ex-tracts. However, these authors failed todetect the major core protein precursor even though their anti-MuMTVserumhadastronganti-p27 activity when tested against purified virus (4). When used in labeled cell extracts, therefore, this same antiserum should have precipitated the p27 precursor,butbecause of therelatively small difference in molecularweights, the p27
precursor was probably not resolved from the glycoproteinprecursorin theirgelsystem.
The major intracellular protein that is precip-itated withmonospecific anti-p27serumaftera
shortpulseisaspeciesof 75,000daltons,which
has arelatively longhalf-life of about 3to4 h
inthe MuMT-73 cells.Duringthe chaseperiods,
the 75,000-dalton protein displayed a slightly
altered mobility in the SDS-gels. We have de-termined that the alteredmigration results from
post-translational phosphorylation of the
pre-cursor (unpublished data). Two additional
pro-teinsareprecipitatedwithanti-p27 serumfrom the MuMT-73 cells. One isaminor protein of about100,000 molecularweight,which isnearly undetectable aftera 180-minchaseperiod.The otherproteinismoreprominentandhas a
mo-lecular weight of 34,000 daltons. This 34,000-dalton species is observed after the 60-min
la-beling periodaswellasafter the 180-min chase
period,andmightrepresent aprocessing inter-mediate precursor of p27. This polypeptide is not to be confused with the viral glycoprotein
gp34, which showsabsolutelynocross-reactivity
withouranti-p27serum.The34,000-dalton
pro-tein, which isprecipitatedwithanti-p27serum, hasalso beenfoundtobe phosphorylated (un-published data). Very little p27 can be found
intracellularly, even after the chase periods
whenlabeledvirion-associatedp27canbe
read-ily detected in the culture medium, suggesting that the final cleavage step to mature p27 isa very late event in virus maturation.
The major glycoproteingp47 isinitially
syn-thesized as a polypeptide of 70,000 daltons, which is processed about twice as fast as the corresponding p27 precursor pre p75. Large quantities of gp47 are found to be associated with thecells and in the extracellular medium,
both in the viral fraction and in the virus-free media. In addition, largeamountsof the glyco-proteinprecursor pre gp70 are found extracel-lularly in the virus-free media, butno glycopro-teinprecursoris found associated with the iso-lated virus fraction. This observation suggests
that thepre gp7O found in the media doesnot
result from the breakdown of virus, but is prob-ably shed from the cell membrane in a free form. Thematuregp47might also be shed from cell membranes, although it is possible that it derives from breakdown of virus. Our observa-tion of the in vitrosheddingof viral glycoprotein by tissue culture cells might explain the obser-vation of elevated levels of MuMTV glycopro-teinantigensintheplasma of mammary
tumor-bearing mice as reported by Ritzi et al. (17).
Both gp47 and pre gp70 readily incorporate [3H]glucosamine, indicating that both species areheavily glycosylated.
Inspection of the gel profile of the
immuno-precipitated cell extracts reveals that they all containaprominent high-molecular-weight
pro-tein of about 250,000. This protein, therefore,
seems to bea "sticky" cellular contaminant of
the immunoprecipitates since it isalso
precipi-tatedby controlserumandcanbe quantitatively reduced in the cellextracts by a preadsorption
procedure with nonimmune serumand
staphy-lococcal adsorbent. The fact that thislarge cel-lular protein incorporates [3H]glucosamine, is released from cells into the media, and has a
molecular weight of about 250,000 led us to
consider that it is perhaps the large external transformation-sensitive glycoprotein referred
toby theacronymLETS(7).Analysis ofcellular
extracts with anti-LETS serum confirmed its
identityastheLETSglycoprotein.The
MuMT-73epithelialcellssynthesizeconsiderable
quan-tities of LETS and continuously shed it, along
with otheraggregated species, into the culture
medium.
Another protein ofcellular origin, actin, has been identified both in theimmunoprecipitates
ofcellextractsand in purifiedMuMTV
prepa-rations,onthe basis ofcomigrationinSDS-gels
with actin extracted from RIII mouse skeletal
muscle.
Ourfindings oftwo distinct precursor forms
for the two main MuMTV structural proteins,
one for the major envelope constituent and a secondfor themajorinternalcorepolypeptide,
isconsistent with thestudyofSchochetmanand
on November 10, 2019 by guest
http://jvi.asm.org/
Schlom
(22), which indicated noncoordinate synthesisof these two MuMTV proteins. A sim-ilar mechanism of noncoordinate synthesis of viral structural protein has been well established by a number of laboratories for the better-char-acterized typeC RNA tumor viruses (9, 24, 27). The findings presented in this report, together with the previous findings with MuMTV and type C RNA tumor viruses, suggest that non-coordinate synthesis of viral structural proteins, by way of independently synthesized high-mo-lecular-weight precursors, might be a general characteristic ofall
RNA tumorviruses.Further studies using antisera to the other MuMTV proteins as well as tryptic peptide anal-yses
will
be required to firmly establish the precursor-product relationship between the var-ious MuMTV proteins.ACKNOWLEDGMENTS
We are indebted to H. Oppermann of the University of
Californiaat San Francisco for providing us with the Staphy-lococcus aureus Cowan I strain. We thank Lan Bo Chen of the Cold Spring Harbor Laboratory for his gift of anti-LETS serum. We also thank E. Whittington forhis assistance in preparing the labeled, purified MuMTV marker virus.
This work was supported byPublicHealth Service grants CA-08748 and CA-17129 from the National Cancer Institute.
LITERATURE CIED
1. Baltimore, D. 1975. Tumorviruses: 1974. Cold Spring Harbor Symp. Quant.Biol.39:1187-1200.
2. Bonner, M. W., and R.Laskey. 1974. A film detection method for tritium-labeled proteins and nucleic acids in polyacrylamide gels. Eur. J. Biochem.46:8388.
3. Clarke, M., and J. A.Spudich. 1977. Nonmuscle con-tractile proteins: the role of actin and myosin incell
motility and shape determination. Annu. Rev. Biochem. 46:797-822.
4. Dickson, C., J. P. Puma, and S. Nandi. 1975. Intracel-lular synthesis of mousemammary tumorvirus poly-peptides: indication of a precursor glycoprotein. J. Virol. 16:250-258.
5. Dickson, C., J. P.Puma,and S. Nandi. 1976. Identifi-cation of a precursor protein to the major glycoproteins of mousemammary tumor virus. J. Virol. 17:275-282. 6. Dickson, C., and J. J.Skehel. 1974. The polypeptide composition of mousemammarytumorvirus.Virology 58:387-395.
7. Hynes, R.0.1976. Cell surface proteins and malignant transformation.Biochim.Biophys. Acta 458:73-107. 8. Hynes, R.O.,and A. Destree. 1977.Extensivedisulfide
bonding at themammalian cell surface. Proc. Natl. Acad. Sci. U.S.A. 74:2855-2859.
9. Jamjoom, G. A., R. B. Naso,andR.B. Arlinghaus. 1977. Further characterization of intracellular precursor polyproteins of Rauscher leukemia virus. Virology 78:11-34.
10. Kessler, S. W. 1975. Rapid isolation of antigens from cells with a staphylococcal protein A-antibody adsorb-ent: parameters of the interaction of antibody-antigen complexes with protein A. J. Immunol. 15:1617-1624. 11. Kimball, P. C., M. Boehm-Truitt, G. Schochetman, and J.Schlom.1976. Characterization of mouse mam-mary tumorvirusesfrom primary tumorcell cultures. I. Immunological and structural studies. J.Natl. Cancer Inst. 56:111-117.
12. Laemmli, U. K. 1970. Cleavage of structural proteins
duringtheassembly oftheheadofbacteriophageT4. Nature(London) 227:680-682.
13. Lasfargues,E.Y., B. Kramarsky,N.H. Sarkar, J. C.Lasfargues,and D.H.Moore.1972. Anestablished RIII mousemammary cell line:kinetics of mammary tumorvirus (MTV) production. Proc. Soc. Exp. Biol. Med. 139:242-247.
14. Marcus, S.L.,N. H.Sarkar,and M. J. Modak. 1976. Purification andproperties of murinemammary tumor virus DNApolymerase. Virology 71:242-254. 15. Owens,R.B., and A. J.Hackett. 1972. Tissue culture
studies of mouse mammary tumor cells and associated viruses. J. Natl. Cancer Inst.49:1321-1332.
16. Racevskis, J., and G.Koch. 1977. Viral protein synthe-sis in Friend erythroleukemia cell lines. J. Virol. 21:328-337.
17. Ritzi, E.,D.S.Martin,R. L Stolfi, and S. Spiegeiman. 1976. Plasma levels of a viral protein as a diagnostic signalfor the presence of tumor: the murinemammary tumormodel.Proc. Natl. Acad. Sci.U.S.A.73:4190-4194. 18. Sarkar,N.H., and A. S. Dion. 1975. Polypeptides of the mouse mammary tumor virus. I. Characterization oftwogroup-specific antigens. Virology64:471-491. 19. Sarkar, N.H.,A. A. Pomenti, and A. S. Dion. 1977.
Replicationofmousemammary tumorvirus intissue culture. L. Establishment of a mousemammary tumor cell line, virus characterization, and quantitation of virus production. Virology 77:12-30.
20. Sarkar, N.H., N. E. Taraschi, A. A. Pomenti, and A. S. Dion. 1976. Polypeptides of the mouse mammary tumorvirus.II. Identification of two major glycopro-teinswith the viral structure. Virology 69:677-690. 21. Sarkar,N.H.,and E.S.Whittington.1977.
Identifica-tion ofthe structuralproteins of the murine mammary tumorvirusthat are serologicallyrelated to theantigens
ofintracytoplasmictype-Aparticles.Virology8l:91-106.
22. Schochetman, G., and J. Schlom. 1976. Independent polypeptide chain initiation sites for the synthesis of different classes of proteins for an RNAtumor virus: mousemammarytumor virus.Virology73:431-441. 23. Shapiro, S. Z., and J. T. August. 1976. The use of
immunoprecipitation tostudy the synthesis and cleav-age processing of viral proteins. J.Immunol. Methods 13:153-159.
24. Shapiro, S. Z., M. Strand, and J. T. August. 1976. High molecular weightprecursorpolypeptidesto struc-tural proteins of RauscherMurineLeukemiaVirus. J. Mol.Biol. 107:459-477.
25. Teramoto, Y. A.,M. J.Puentes,L J.Young,and R. D. Cardiff. 1974. Structure of the mouse mammary tumor virus: polypeptides andglycoproteins. J. Virol. 13:411418.
26. Van Blerkom, J., and C.Manes. 1974. Development ofpreimplantationrabbitembryos invivo and invitro. II.Acomparison of qualitativeaspectsofprotein syn-thesis. Develop.Biol. 40:40-51.
27. VanZaane, D., M. J. A.Dekker-Michielsen, and H. P. J.Bloemers. 1976.Virus-specific precursor polypep-tides incelLs infected with Rauscher leukemia virus: synthesis, identification, and processing. Virology 75:113-129.
28. Wang, E., B. A. Wolf,R. A.Lamb, P. W. Choppin, and A. R. Goldberg. 1976. The presence of actin in enveloped viruses, p. 589-599. In R.Goldman,T. Pol-lard, and J.Rosenbaum (ed.), ColdSpringHarbor con-ferences on cellproliferation, vol. 3. Cell motility.Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y. 29. Yagi, M. J. 1973. Cultivation and characterization of BALB/cf C3H mammary tumor cell lines. J. Natl. Cancer Inst. 51:1849-1860.
30. Yagi, M. J., and R. W. Compans. 1977. Structural components of mousemammary tumor virus. I. Poly-peptides of the virion.Virology76:751-766.
383
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 3.serum.Samewithimmunoprecipitatedwith Analysis ofantiserum specificity. (a) P5S]methionine-labeled purified MuMTV immunoprecipitated monospecific anti-gp27 serum](https://thumb-us.123doks.com/thumbv2/123dok_us/1535482.106126/5.505.151.382.68.366/samewithimmunoprecipitatedwith-analysis-ofantiserum-specificity-methionine-purified-immunoprecipitated-monospecific.webp)
![FIG. with anti-gp47markerwithcellswith(1)(h), pH]glucosamine-labeled immunoprecipitated anti-p27 Mixturemarkers)](https://thumb-us.123doks.com/thumbv2/123dok_us/1535482.106126/8.505.128.370.72.367/fig-with-anti-markerwithcellswith-glucosamine-labeled-immunoprecipitated-mixturemarkers.webp)