JOURNAL OFVIROLOGY, June 2006, p. 5631–5636 Vol. 80, No. 11 0022-538X/06/$08.00⫹0 doi:10.1128/JVI.00219-06
Copyright © 2006, American Society for Microbiology. All Rights Reserved.
Two Surface-Exposed Elements of the B30.2/SPRY Domain as
Potency Determinants of N-Tropic Murine Leukemia Virus
Restriction by Human TRIM5
␣
Michel J. Perron,
1Matthew Stremlau,
1and Joseph Sodroski
1,2*
Department of Cancer Immunology and AIDS, Dana-Farber Cancer Institute, Department of Pathology, Division of AIDS, Harvard Medical School, Boston, Massachusetts 02115,1and Department of Immunology and Infectious Diseases,
Harvard School of Public Health, Boston, Massachusetts 021152
Received 31 January 2006/Accepted 7 March 2006
Human TRIM5␣(TRIM5␣hu) potently restricts N-tropic (N-MLV), but not B-tropic, murine leukemia virus
in a manner dependent upon residue 110 of the viral capsid. Rhesus monkey TRIM5␣(TRIM5␣rh) inhibits
N-MLV only weakly. The study of human-monkey TRIM5␣chimerae revealed that both the v1 and v3 variable regions of the B30.2/SPRY domain contain potency determinants for N-MLV restriction. These variable regions are predicted to be surface-exposed elements on one face of the B30.2 domain. Acidic residues in v3 complement basic residue 110 of the N-MLV capsid. The results support recognition of the retroviral capsid by the TRIM5␣B30.2 domain.
Primates express dominant restriction factors that block ret-rovirus infection soon after entry but prior to reverse transcrip-tion (1, 2, 4). Most early restrictranscrip-tion in primates is mediated by TRIM5␣(5, 6, 8, 10, 12). TRIM5␣is a member of the tripar-tite-motif family of proteins and contains RING, B-box 2, and coiled-coil (RBCC) domains (11). TRIM5␣ also contains a C-terminal B30.2/SPRY domain, which is essential for antiviral activity and has been implicated in interaction with the tar-geted viral capsid (9, 13, 16). Species-specific differences in TRIM5␣account for the patterns of restriction of retroviruses in Old World and New World primates. For example, TRIM5␣hu
from humans potently restricts N-tropic murine leukemia virus (N-MLV), whereas TRIM5␣rh from rhesus monkeys exhibits
much weaker activity against this virus (6, 7, 10, 18). On the other hand, TRIM5␣rhpotently blocks human immunodeficiency virus
type 1 (HIV-1), which is only weakly inhibited by TRIM5␣hu(15,
16). Four variable regions (v1 to v4) are found in the B30.2 domains of TRIM5␣proteins from different primates (12, 14). Differences in the v1 regions of TRIM5␣huand TRIM5␣rh
ac-count for the differences in anti-HIV-1 potency exhibited by these TRIM5␣variants (9, 17, 19).
The TRIM5␣region that dictates the potency of TRIM5␣hu
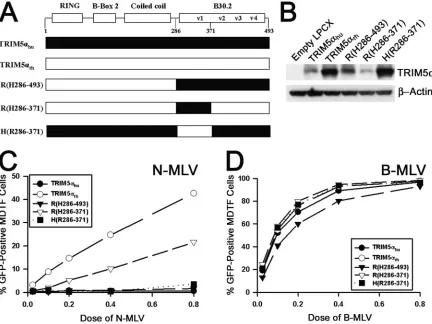
for N-MLV restriction has not been defined. To this end, we generated several chimerae between the human and rhesus TRIM5␣proteins (Fig. 1A) and examined the abilities of these chimerae to restrict N-MLV and B-tropic murine leukemia virus (B-MLV) infection in transduced MDTF cells. The ex-pression of each construct was confirmed by Western blot analysis (Fig. 1B). TRIM5␣hurestricted N-MLV significantly
more potently than TRIM5␣rh, even though the steady-state
levels of TRIM5␣hu expression were lower than those of
TRIM5␣rh(Fig. 1C). The R(H286-493) mutant, in which the
TRIM5␣rhB30.2 domain is replaced with that of TRIM5␣hu,
restricted N-MLV infection significantly more efficiently than TRIM5␣rh. Thus, the B30.2 domain of TRIM5␣hucontains the
determinant(s) for potent N-MLV restriction.
To define further the region(s) of the B30.2 domain involved in N-MLV restriction, two reciprocal chimerae, R(H286-371) and H(R286-371), were expressed in MDTF cells. The cells were challenged with N- and B-MLV (Fig. 1C). N-MLV infec-tion of the R(H286-371)-expressing cell line was reduced ap-proximately two- to threefold compared with that of the TRIM5␣rh-expressing cell line. The block to N-MLV infection
in MDTF cells expressing the H(R286-371) chimera was al-most as great as that in the TRIM5␣hu-expressing cells. No
difference in susceptibility to B-MLV infection was observed in any of the MDTF cell lines expressing TRIM5␣variants (Fig. 1D). These findings indicate that potency determinants for N-MLV restriction reside within multiple regions of the TRIM5␣ B30.2 domain, consistent with previous studies (19).
The v1 region in the TRIM5␣rhB30.2 domain is a major
determinant of anti-HIV-1 potency (9, 17, 19). To test whether any of the TRIM5␣huB30.2 variable regions are involved in
N-MLV restriction, we generated several chimerae in which a small segment of one TRIM5␣rhvariable region was replaced
by the corresponding sequence from TRIM5␣hu(Fig. 2A). We
assayed for a gain in N-MLV restriction in transduced MDTF cells compared to wild-type TRIM5␣rh. The steady-state levels
of expression of the TRIM5␣ variants are shown in Fig. 2B. The Rh(SYQ/PCK) and Rh(GSFA/SFSV) mutants exhibited only modest improvements in blocking N-MLV infection com-pared to TRIM5␣rh(Fig. 2C); apparently, these differences do
not account for the potency of TRIM5␣hu. By contrast, the
Rh(LFTFPSLT/RYQT–FV) and Rh(QYV/ECA) mutants re-stricted N-MLV infection as potently as TRIM5␣hu. B-MLV
infection was unaffected by expression of the TRIM5␣variants (Fig. 2D). We conclude that at least two potency determinants for N-MLV restriction reside within the TRIM5␣huB30.2
do-main, one within the v1 region between residues 335 and 340 and another in the N-terminal portion of v3.
* Corresponding author. Mailing address: Dana-Farber Cancer Insti-tute, 44 Binney Street, JFB 824, Boston, MA 02115. Phone: (617) 632-3371. Fax: (617) 632-4338. E-mail: [email protected].
5631
on November 8, 2019 by guest
http://jvi.asm.org/
The presence of a positively charged arginine residue at position 110 of the viral capsid renders N-MLV susceptible to TRIM5␣hurestriction (10). Alteration of this N-MLV capsid
residue from an arginine to the corresponding residue in B-MLV, glutamic acid, creates NBNN-B-MLV, which partially es-capes from TRIM5␣hurestriction (10). Conversely,
substitu-tion of arginine for the glutamic acid at residue 110 of the B-MLV capsid creates BNBB-MLV, which is susceptible to TRIM5␣hurestriction (10). The correlation between a positive
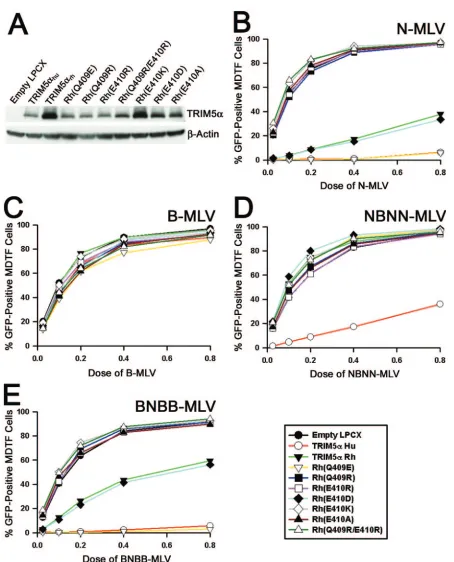
charge at position 110 of the MLV capsid and restriction sug-gested the hypothesis that the negative charges within the TRIM5␣v3 region (Fig. 2A) are involved in an electrostatic interaction with the viral capsid. To test this hypothesis, we generated several mutants with changes in residues 409 and 410 in the TRIM5␣rhv3 region and examined their abilities to
restrict N-MLV, B-MLV, NBNN-MLV,and BNBB-MLV. Mu-tant TRIM5␣constructs were expressed stably in MDTF cells (Fig. 3A). Introduction of a negatively charged glutamic acid at residue 409 of TRIM5␣rhcreated Rh(Q409E), which restricted
N-MLV infection to a level comparable to that of TRIM5␣hu.
However, alteration of residue 409 to a positively charged
arginine residue, Rh(Q409R), completely abrogated restric-tion of N-MLV. Likewise, alterarestric-tion of the adjacent TRIM5␣rh
residue 410 from a glutamic acid to either a positively charged residue, Rh(E410R) or Rh(E410K), or an uncharged residue, Rh(E410A), also abrogated restriction of N-MLV. On the other hand, the Rh(E410D) mutant, which retains the negative charge at residue 410, restricted N-MLV as efficiently as TRIM5␣rh. None of the TRIM5␣rhmutants restricted B-MLV
infection (Fig. 3C). These data suggest that acidic residues within the TRIM5␣v3 region enable potent N-MLV restriction.
To investigate whether the N-MLV restriction observed in the mutant TRIM5␣rh-expressing cell lines is dependent upon
the charge at position 110 of the viral capsid, we tested whether NBNN-MLV and BNBB-MLV are sensitive to restriction in the mutant TRIM5␣rh-expressing cell lines. NBNN-MLV was
able to completely escape restriction by TRIM5␣rh and all
TRIM5␣rh mutants; however, only a partial escape was
ob-served in the TRIM5␣hu-expressing cell line (Fig. 3D).
Con-versely, potent restriction of BNBB-MLV was observed in the TRIM5␣hu- and Rh(Q409E)-expressing cell lines, whereas
[image:2.585.72.504.70.394.2]only a partial restriction was observed in the TRIM5␣rh- and
FIG. 1. Contribution of the TRIM5␣huB30.2 domain to potent restriction of N-MLV. (A) The TRIM5␣chimerae generated to examine the role of the B30.2 domain in N-MLV restriction are depicted. The residue numbers of TRIM5␣huare shown, and TRIM5␣husegments are colored black. (B) Steady-state levels of expression of TRIM5␣variants in MDTF cells are shown. Lysates from MDTF cells transduced with either an empty LPCX vector or vectors expressing HA-tagged TRIM5␣variants were subjected to Western blotting using an anti-HA antibody. The lysates were also Western blotted for-actin to control for total protein. (C and D) MDTF cells transduced with LPCX vectors expressing TRIM5␣ variants were incubated with various amounts of N-MLV–green fluorescent protein (GFP) (C) or B-MLV-GFP (D). GFP-positive MDTF cells were counted by fluorescence-activated cell sorter. The results from a typical experiment are shown. Similar results were obtained in at least two independent experiments.
5632 NOTES J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
Rh(E410D)-expressing cells (Fig. 3E). There was no restric-tion of BNBB-MLV in the cells expressing Rh(Q409R), Rh(E410R), Rh(E410K), Rh(E410A), or Rh(Q409R/E410R). Thus, there is a correlation between the presence of negative charges at residues 409 and 410 of TRIM5␣rhand restriction of
an MLV containing a positively charged residue at position 110 of the viral capsid.
Our results indicate that two variable regions, v1 and v3, in the B30.2 domain of TRIM5␣hucontribute to the potent
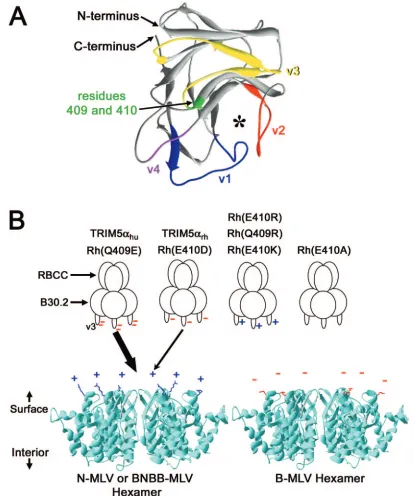
re-striction of N-MLV infection. Recently, the B30.2/SPRY do-main has been shown to assume a-sandwich, lectin-like fold (5). By analogy with this structure, the major variable regions
(v1 to v4) of the TRIM protein B30.2 domains (14) are pre-dicted to be surface-exposed elements on the same face of the domain (Fig. 4A); the variable loops surround a putative li-gand-binding site (5). Thus, the v1 and v3 regions of TRIM5␣ are well positioned to serve as determinants of interaction with the retroviral capsid.
The presence of negatively charged residues at positions 409 and 410 of the TRIM5␣rhB30.2 domain potentiates restriction
of MLVs that have an arginine at position 110 in the capsid. Alteration of either of these acidic TRIM5␣rh residues to a
positively charged arginine completely abrogated TRIM5␣ -mediated restriction of N-MLV. It is tempting to speculate that
FIG. 2. Potency determinants for N-MLV restriction in two B30.2 variable regions. (A) Differences between the primary sequences of TRIM5␣huand TRIM5␣rhin the B30.2 domain v1 to v3 variable regions are marked with asterisks. The locations of the changes in the v1, v2, and v3 regions of the B30.2 domain of the TRIM5␣rhmutants are shown beneath the alignment. Residues 409 and 410 in the v3 region of the TRIM5␣rh B30.2 domain are underlined. These residues, which are both acidic in TRIM5␣hu, are studied in greater depth in the experiments shown in Fig. 3. (B) Steady-state levels of expression of the TRIM5␣variants in MDTF cells were analyzed by Western blotting cell lysates, as in Fig. 1. (C and D) MDTF cells transduced with LPCX vectors expressing TRIM5␣variants were incubated with various amounts of N-MLV–green fluorescent protein (GFP) (C) or B-MLV-GFP (D). GFP-positive cells were counted by fluorescence-activated cell sorter. Panels C and D show data from a typical experiment. Similar results were obtained in at least two independent experiments.
VOL. 80, 2006 NOTES 5633
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 3. Contribution of charge to the potency determinant in the TRIM5␣B30.2 v3 region. (A) Steady-state levels of expression of TRIM5␣ variants in MDTF cells were measured by Western blotting cell lysates with an anti-HA antibody, as described in the legend to Fig. 1. TRIM5␣rh mutants are designated Rh, with the introduced amino acid change in parentheses. The number refers to the residue number in TRIM5␣rh. (B to E) MDTF cells transduced with either an empty LPCX vector or the LPCX vectors expressing TRIM5␣variants were incubated with N-MLV– green fluorescent protein (GFP) (B), B-MLV-GFP (C), NBNN-MLV-GFP (D), or BNBB-MLV-GFP (E). GFP-positive MDTF cells were counted by fluorescence-activated cell sorter. Panels B to E show the results of typical experiments. Similar results were obtained in at least two independent experiments.
5634 NOTES J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
FIG. 4. Models to explain the potency of N-MLV restriction by TRIM5␣variants. (A) The structure of the B30.2/SPRY domain of the human PRY-SPRY-19q13.4.1 protein (5) is shown, with the surface loops equivalent to the v1 to v4 variable regions of TRIM proteins colored (14). The strands corresponding to the v1 (blue), v2 (red), v3 (yellow), and v4 (purple) regions are labeled. The predicted locations of residues 409 and 410 on TRIM5␣rhare indicated. The asterisk marks a potential ligand-binding cleft (5). The N and C termini of the B30.2/SPRY domain are labeled. (B) Electrostatic interactions between the B30.2 domain v3 region of TRIM5␣and the surface of the viral capsid may modulate the potency of MLV restriction. The trimeric TRIM5␣variants, with either negative (red) or positive (blue) charges in the N terminus of the B30.2 domain v3 region, are depicted. The ribbon structure of the MLV capsid hexamers is shown in a lateral view (8). From this perspective, the surface of the assembled capsid faces upward, and the interior of the capsid faces downward. The surface-exposed side chain of residue 110 is shown (arginine [blue] in N-MLV and BNBB, glutamic acid [red] in B-MLV). The presence of a single negative charge in the TRIM5␣v3 N terminus allows modest restriction of N-MLV and BNBB (thin arrow). N-MLV and BNBB restriction is potentiated (thick arrow) by the addition of a second negative charge in this v3 region [as in TRIM5␣huor Rh(Q409E)]. Removal of these acidic v3 residues [in Rh(E410A)] or addition of a basic residue [in Rh(E410R), Rh(Q409R), and Rh(E410K)] completely abrogates TRIM5␣restriction of N-MLV and BNBB. The negatively charged B-MLV capsid is resistant to TRIM5␣-mediated restriction.
VOL. 80, 2006 NOTES 5635
on November 8, 2019 by guest
http://jvi.asm.org/
the N terminus of the TRIM5␣v3 region interacts electrostat-ically with the N-MLV capsid, perhaps involving capsid residue 110 (Fig. 4B). Electrostatics may also contribute to Fv-1n
re-striction in mice; for example, MLVs containing arginine 110 in the capsid are insensitive to Fv-1n restriction unless the
positively charged lysine at position 358 of Fv-1 is removed (3). Although alteration of both TRIM5␣rhresidues 409 and 410 to
basic residues abrogated N-MLV restriction, it did not confer the ability to restrict B-MLV. Additional studies should clarify how shape and bond formation govern the recognition of tar-geted retroviral capsids by TRIM5␣proteins.
We thank Yvette McLaughlin for manuscript preparation. We thank the National Institutes of Health (AI063987 and a Center for AIDS Research Award [AI60354]), the International AIDS Vac-cine Initiative, the Bristol-Myers Squibb Foundation, the William A. Haseltine Foundation for the Arts and Sciences, and the late William F. McCarty-Cooper for financial support. M.S. was supported by a National Defense Science and Engineering Fellowship and is a Fellow of the Ryan Foundation.
REFERENCES
1.Besnier, C., L. Ylinen, B. Strange, A. Lister, Y. Takeuchi, S. P. Goff, and G. J. Towers.2003. Characterization of murine leukemia virus restriction in
mam-mals. J. Virol.77:13403–13406.
2.Bieniasz, P. D.2003. Restriction factors: a defense against retroviral
infec-tion. Trends Microbiol.11:286–291.
3.Bishop, K. N., M. Bock, G. Towers, and J. P. Stoye.2001. Identification of the regions of Fv1 necessary for murine leukemia virus restriction. J. Virol.
75:5182–5188.
4.Goff, S. P.2004. Genetic control of retrovirus susceptibility in mammalian
cells. Annu. Rev. Genet.38:61–85.
5.Grutter, C., C. Briand, G. Capitani, P. R. Mittl, S. Papin, J. Tschopp, and M. G. Grutter.2006. Structure of the PRYSPRY-domain: implications for
autoinflammatory diseases. FEBS Lett.580:99–106.
6.Hatziioannou, T., D. Perez-Caballero, A. Yang, S. Cowan, and P. D. Bieniasz.
2004. Retrovirus resistance factors Ref1 and Lv1 are species-specific variants
of TRIM5␣. Proc. Natl. Acad. Sci. USA101:10774–10779.
7.Keckesova, Z., L. M. Ylinen, and G. J. Towers.2004. The human and African
green monkey TRIM5␣genes encode Ref1 and Lv1 retroviral restriction
factor activities. Proc. Natl. Acad. Sci. USA101:10780–10785.
8.Mortuza, G. B., L. F. Haire, A. Stevens, S. J. Smerdon, J. P. Stoye, and I. A. Taylor. 2004. High-resolution structure of a retroviral capsid hexameric
amino-terminal domain. Nature431:481–485.
9.Perez-Caballero, D., T. Hatziioannou, A. Yang, S. Cowan, and P. D. Bieniasz.
2005. Human tripartite motif 5␣domains responsible for retrovirus
restric-tion activity and specificity. J. Virol.79:8969–8978.
10.Perron, M. J., M. Stremlau, B. Song, W. Ulm, R. C. Mulligan, and J. Sodroski.2004. TRIM5␣mediates the postentry block to N-tropic murine
leukemia viruses in human cells. Proc. Natl. Acad. Sci. USA101:11827–
11832.
11.Reymond, A., G. Meroni, A. Fantozzi, G. Merla, S. Cairo, L. Luzi, D. Riganelli, E. Zanaria, S. Messali, S. Cainarca, A. Guffanti, S. Minucci, P. G. Pelicci, and A. Ballabio. 2001. The tripartite motif family identifies cell
compartments. EMBO J.20:2140–2151.
12.Sawyer, S. L., L. I. Wu, M. Emerman, and H. S. Malik.2005. Positive
selection of primate TRIM5␣identifies a critical species-specific retroviral
restriction domain. Proc. Natl. Acad. Sci. USA102:2832–2837.
13.Sebastian, S., and J. Luban.2005. TRIM5␣selectively binds a
restriction-sensitive retroviral capsid. Retrovirology2:40.
14.Song, B., B. Gold, C. O’Huigin, H. Javanbakht, X. Li, M. Stremlau, C. Winkler, M. Dean, and J. Sodroski.2005. The B30.2(SPRY) domain of the
retroviral restriction factor TRIM5␣exhibits lineage-specific length and
se-quence variation in primates. J. Virol.79:6111–6121.
15.Song, B., H. Javanbakht, M. Perron, D. H. Park, M. Stremlau, and J. Sodroski.2005. Retrovirus restriction by TRIM5␣variants from Old World
and New World primates. J. Virol.79:3930–3937.
16.Stremlau, M., C. M. Owens, M. J. Perron, M. Kiessling, P. Autissier, and J. Sodroski.2004. The cytoplasmic body component TRIM5␣restricts HIV-1
infection in Old World monkeys. Nature427:848–853.
17.Stremlau, M., M. Perron, S. Welikala, and J. Sodroski.2005. Species-specific
variation in the B30.2(SPRY) domain of TRIM5␣determines the potency of
human immunodeficiency virus restriction. J. Virol.79:3139–3145.
18.Yap, M. W., S. Nisole, C. Lynch, and J. P. Stoye.2004. Trim5␣protein restricts both HIV-1 and murine leukemia virus. Proc. Natl. Acad. Sci. USA
101:10786–10791.
19.Yap, M. W., S. Nisole, and J. P. Stoye.2005. A single amino acid change in
the SPRY domain of human Trim5␣leads to HIV-1 restriction. Curr. Biol.
15:73–78.
5636 NOTES J. VIROL.