Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Human Immunodeficiency Virus Type 1 Nef Associates with Lipid
Rafts To Downmodulate Cell Surface CD4 and Class I Major
Histocompatibility Complex Expression and
To Increase Viral Infectivity
Melissa Alexander, Yeou-cherng Bor, Kodimangalam S. Ravichandran,
Marie-Louise Hammarskjo¨ld, and David Rekosh*
Myles H. Thaler Center for AIDS and Human Retrovirus Research, Carter Immunology Center, and the Department of Microbiology, University of Virginia, Charlottesville, Virginia 22908
Received 7 April 2003/Accepted 8 November 2003
Lipid rafts are membrane microdomains that are functionally distinct from other membrane regions. We have shown that 10% of human immunodeficiency virus type 1 (HIV-1) Nef expressed in SupT1 cells is present in lipid rafts and that this represents virtually all of the membrane-associated Nef. To determine whether raft targeting, rather than simply membrane localization, has functional significance, we created a Nef fusion pro-tein (LAT-Nef) containing the N-terminal 35 amino acids from LAT, a propro-tein that is exclusively localized to rafts. Greater than 90% of the LAT-Nef protein was found in the raft fraction. In contrast, a mutated form, lacking two cysteine palmitoylation sites, showed less than 5% raft localization. Both proteins were equally ex-pressed and targeted nearly exclusively to membranes. The LAT-Nef protein was more efficient than its nonraft mutant counterpart at downmodulating both cell surface CD4 and class I major histocompatibility complex (MHC) expression, as well as in enhancing first-round infectivity and being incorporated into virus particles. This demonstrates that targeting of Nef to lipid rafts is mechanistically important for all of these functions. Compared to wild-type Nef, LAT-Nef downmodulated class I MHC nearly as effectively as the wild-type Nef protein, but was only about 60% as effective for CD4 downmodulation and 30% as effective for infectivity enhancement. Since the LAT-Nef protein was found entirely in rafts while the wild-type Nef protein was distributed 10% in rafts and 90% in the soluble fraction, our results suggest that class I MHC downmodulation by Nef may be performed exclusively by raft-bound Nef. In contrast, CD4 downmodulation and infectivity enhancement may require a non-membrane-bound Nef component as well as the membrane-bound form.
Human immunodeficiency virus type 1 (HIV-1) Nef is a 27-kDa myristoylated protein that has been shown to be an important determinant of viral pathogenesis, since individuals infected with viruses carrying defective Nef alleles progress very slowly to disease (6, 15, 37). Three well-described func-tions of Nef in cell culture systems are the enhancement of virion infectivity (11, 45, 57) and downmodulation of CD4 (1, 24) and certain class I major histocompatibility complex (MHC) alleles (13, 58). Multiple other functions of Nef have also been reported. While there has been considerable specu-lation about the role that all of these functions might play in in vivo infection, the question of why Nef is required for viral pathogenesis remains unanswered (for reviews see references 4, 16, 18, and 25).
Several models of how Nef downmodulates CD4 and certain class I MHC alleles have been proposed. There is general agreement that Nef induces the internalization of CD4 via clathrin-coated pits, probably by binding directly to the cyto-plasmic tail of CD4 (53), and then directly (14, 49) or indirectly (26, 40) linking to a cellular adaptor complex, most likely either AP-1 (7, 17) or AP-2 (17, 26, 30). Nef has also been implicated in a second step that targets CD4 to lysosomes for
degradation (33, 50). Downmodulation of class I MHC by Nef is less well characterized, although it is thought to be clathrin independent (38) and utilize intracellular sorting molecules. It has been suggested that Nef downmodulates class I MHC via the Golgi and the PACS-1 sorting pathway (52, 63). Recently, a direct but weak interaction of Nef with the cytoplasmic tail of class I MHC alleles has been demonstrated (72), and it has also been suggested that Nef blocks the transport of newly synthe-sized class I MHC molecules to the cell surface (34).
Nef increases the infectivity of virus particles produced in its presence (12, 45, 57), but the manner by which it does this is incompletely understood. Downmodulation of CD4 by Nef is clearly a contributing factor to a form of infectivity enhance-ment (36, 56), but mutational analysis of Nef has shown that it is not the only mechanism (27), and particle infectivity en-hancement can be demonstrated even in cells lacking CD4 (10, 11, 45). Several groups have now shown that Nef is packaged into virus particles (9, 44, 47, 68, 69), probably as part of the viral core (20, 35). However, the inclusion of Nef in virus particles has not yet been directly linked to the increase of infectivity phenotype. Additionally, it is not known whether infectivity enhancement is the result of an alteration of the particle as it is produced or whether it is an effect of incoming Nef in the target cell after infection. Recently, it has been suggested that Nef may increase particle infectivity by altering the lipid and cholesterol composition of the virus particle as it is made (76).
* Corresponding author. Mailing address: University of Virginia, Department of Microbiology, Jordan Hall Rm. 7-87, P.O. Box 800734, Charlottesville, VA 22908-1386. Phone: (434) 982-1599. Fax: (434) 982-1590. E-mail: [email protected].
1685
on November 8, 2019 by guest
http://jvi.asm.org/
The traditional view of the plasma membrane as a homoge-neous entity has been called into question in recent years with the discovery of portions of the membrane termed lipid rafts (also known as detergent-insoluble glycolipid-enriched complexes and glycolipid-enriched membrane domains) (59). These regions are cholesterol- and sphingomyelin-rich mem-brane microdomains that are thought to play an important role in signal transduction, acting as a platform for the association of signaling molecules (8, 32, 69). Rafts can be readily isolated in a sucrose gradient by virtue of their insolubility in cold non-ionic detergent and low buoyant density (55). Proteins which target to rafts often have long, saturated fatty acid modifica-tions such as myristic acid or palmitic acid (65).
Previous studies have shown that a small fraction of Nef expressed in T-cell lines is membrane bound (3, 23, 68). Tar-geting of Nef to membranes is dependent on the cotransla-tional addition of myristic acid and a stretch of basic amino acids near the amino terminus of the protein (68, 74). A frac-tion of Nef expressed in T-cell lines targets to rafts, and it has been hypothesized that raft association may be critical for Nef function (66, 67, 76). Indeed, a mutation that changes the glycine myristate acceptor site of Nef to alanine (NefG2A)
pre-vents myristoylation and severely impairs the ability of Nef to perform most of its known functions (2, 12, 25, 48, 74). How-ever, since this mutation also completely abolishes the associ-ation of Nef with membranes, the resulting phenotype does not allow the conclusion that specific raft microdomain associa-tion, rather than bulk membrane associaassocia-tion, is the critical factor.
In the present study, we describe a strategy that has allowed us to alter the targeting properties of Nef in a controlled manner to evaluate the functional significance of Nef targeting to raft microdomains versus the nonraft membrane. We dem-onstrate that raft targeting by Nef is functionally important for CD4 and class I MHC downmodulation as well as infectivity enhancement.
MATERIALS AND METHODS
Cell culture.5BD.1 COS packaging cells that stably express HIV Gag-Pol and Env proteins were described previously (62). Cells were maintained with Iscove’s medium supplemented with 10% bovine calf serum and 200g of hygromycin per ml. HeLa CD4 cells were a kind gift from David Camerini (University of California, Irvine), and cells were maintained in Iscove’s medium supplemented with 10% bovine calf serum and 1.5 mg of G418 per ml. SupT1 cells were obtained from the AIDS Research and Reference Reagent Program (National Institutes of Health, Bethesda, Md.) and maintained in RPMI 1640 withL -glutamine and 10% fetal calf serum.
Expression vectors.Nef and LAT-Nef expression constructs were made in either a pCMV or pA vector background. The pCMV vector expressing Nef has been described previously (10, 62). pCMV-Nef (pHR1405) and its derivatives were used for the studies involving infectivity enhancement, which were carried out in 5BD.1 cells. This vector did not express well in SupT1 cells, and thus another vector, pA, was used for SupT1 cell studies. pA is a murine leukemia virus-based pBABE-derived vector which has the 5⬘long terminal repeat re-placed with the chicken actin promoter, and has been described previously (64).
Nef, LAT-Nef, and LATAANef constructs.Plasmids containing either the first
105 bases of the wild-type LAT or LATAAcoding sequence were used as PCR templates to create Nef fusion protein constructs. All constructs were confirmed by DNA sequencing. For constructs in the pCMV background, oligonucleo-tides 5⬘-GCGGGATCCGTCGACCCATGGAGGAGGCCATCCTGG-3⬘(top strand) and 5⬘-GCGCCATGGAGCCTGGCAGTCTGTG-3⬘ (bottom strand) were used. The resulting PCR fragments were then digested with NcoI and subcloned into theNcoI site 5⬘of the NL4-3 NefG2Agene in the pCMV-based vector pHR1864, which has been described previously (60).
For constructs in the pA background, pCMV LAT-Nef(pHR2458)and pCMVLATAA-Nef(pHR2462)were amplified with oligonucleotides 5⬘-GCCG AATTCTATACCATGGAGGAGGCCATCCTG-3⬘(top strand) and 5⬘-GCGT CGACGCATGCTCAGCAGTTCTTGAAG-3⬘ (bottom strand). The resulting products were then cut withEcoRI andSalI and cloned into the pA vector
(pHR2112)between those sites immediately 3⬘of the chicken actin promoter to create the vectors pA LAT-Nef(pHR2613)and pA LATAA-Nef (pHR2614).
The pA Nef vector (pHR2110) was constructed by amplification of NL4-3 Nef from pCMV-Nef with oligonucleotides 5⬘-GCGGATCCGAATTCTATACCAT GGGTGGCAAGTGGTC-3⬘(top strand) and 5⬘-GCGTCGACGCATGCTCA GCAGTTCTTGAAG-3⬘(bottom strand). The resulting PCR product was then cut withEcoRI andSalI and cloned into pA between those sites, following the chicken actin promoter.
The pA NefG2Avector (pHR2696) was constructed by amplification of NL4-3 NefG2Ain pCMV with oligonucleotides 5⬘-GCGGATCCGAATTCTATACCA TGGCTGGCAAGT-3⬘and the same bottom strand oligonucleotide as for the pA Nef construction above. The resulting PCR product was then cut withEcoRI andSalI and cloned into pA between those sites, following the chicken actin promoter.
Transfections.For each sample, 2⫻107SupT1 cells growing in log phase (5
⫻105to 7⫻105/ml) were harvested and resuspended in 250l of fetal calf serum and incubated in a microcentrifuge tube at room temperature for 10 min with the amount and type of plasmid indicated in the figure legends. The cells and DNA were then incubated with another 250l of serum-free RPMI medium for 10 minutes at room temperature. The resulting 500l was then transferred to a 4-mm electroporation cuvette. Electroporation was performed at 290 V and 960F with a Gene Pulser (Bio-Rad, Hercules, Calif.). After allowing the cells to recover for 10 minutes, the cells were resuspended in RPMI medium with L-glutamine containing 10% fetal calf serum at 106cells/ml and incubated until analyzed. When dose-response experiments were performed, the amount of DNA was held constant between samples with the appropriate empty vector.
Flow cytometry.SupT1 cells were cotransfected with 2g of pCMVGFP (BD Bioscience Clontech, Palo Alto, Calif.) and various amounts of the indicated Nef expression vectors. For staining, cells were harvested, resuspended in 50l of staining buffer (Dulbecco’s phosphate-buffered saline with 2% bovine calf serum and 0.1% sodium azide), and incubated for 1 h with phycoerythrin (PE)-conju-gated anti-CD4 (BD Biosciences PharMingen, San Diego, Calif.) or PE-conju-gated anti-HLA-A2 (MA2.1) antibody according to the manufacturer’s protocol. PE-MA2.1 was a kind gift from David Camerini (University of California, Irvine) and was described previously (43). It is specific for MHC antigens HLA-A and HLA-B. Cells were washed three times in staining buffer, fixed in 1% parafor-maldehyde, and analyzed with a FACSCalibur flow cytometer (Becton Dickin-son). Data were analyzed with CellQuest software (Becton DickinDickin-son). The CD4 or HLA-A2 mean fluorescence intensity of the green fluorescent protein (GFP)-positive cells was determined from the gated live cell population.
Membrane isolation.Membrane isolation by flotation centrifugation was per-formed with a modification of a protocol that has been described previously (61). Briefly, 1⫻108to 1.5⫻108SupT1 cells in 400l of flotation buffer (25 mM Tris-HCl, pH 7.5, 150 mM NaCl, 5 mM EDTA, 30 mM sodium pyrophosphate, and 10 mM-glycerophosphate) were lysed by freeze-thaw three times in liquid nitrogen and a 37°C water bath. Lysis was confirmed by trypan blue exclusion. Nuclei and unbroken cells were then spun out at 1,000⫻gat 4°C for 5 min. The pellet was washed once with 100l of flotation buffer, and the resulting super-natant was then added to the first supersuper-natant for a total volume of 400l. The supernatant was then adjusted to 80% sucrose, resulting in 2 ml volume, which was then layered at the bottom of an SW41 Beckman centrifuge tube.
Layers of 65% (5.5 ml) and 10% (4.5 ml) sucrose were then added on top, and the tubes were spun at 35,000 rpm for 18 h at 4°C; 1-ml fractions were harvested from the top of the gradient. The fractions were then made 2% in Triton X-100 and were either immunoprecipitated with a mouse anti-Nef monoclonal antibody prior to separation by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) or applied directly to an SDS gel. After SDS-PAGE, samples were analyzed by Western blotting with a rabbit polyclonal Nef serum, anti-transferrin receptor monoclonal antibody, or anti-Lck antibody. The H68.4 transferrin receptor (CD71) antibody was a kind gift from Samuel Green (Uni-versity of Virginia, Charlottesville) and has been described previously (70). Anti-Lck antibody was obtained from Santa Cruz Biotechnology (Santa Cruz, Calif.). The polyclonal Nef antiserum was made by immunizing rabbits with a glutathioneS-transferase (GST)-Nef fusion protein.
Lipid raft isolation.Lipid rafts were isolated from 108transfected SupT1 cells lysed in 0.5% Triton X-100-containing buffer as described previously (66). Briefly, 1 ml of lysates was adjusted to 40% sucrose and layered with 2 ml of 30% and 1 ml of 5% sucrose and centrifuged at 200,000g (40,000rpm in the SW55Ti
on November 8, 2019 by guest
http://jvi.asm.org/
rotor) for 16 h at 4°C. Fractions from the entire gradient were collected, solu-bilized with 2% Triton X-100 at room temperature for 15 min and were either immunoprecipitated with a Nef monoclonal antibody prior to separation by SDS-PAGE or applied directly to an SDS gel. After SDS-PAGE, samples were analyzed by Western blotting with a rabbit polyclonal Nef serum, anti-transferrin receptor monoclonal antibody, or anti-Lck antibody.
Immunoprecipitations and Western blotting.For experiments to determine expression level, 2⫻107transfected SupT1 cells were harvested 24 h posttrans-fection and lysed in 1 ml of lysis buffer (1% IGEPAL CA-630, 1% deoxycholate, 0.1% SDS, 150M NaCl, 10M Tris HCl, pH 7.4). Gradient fractions or lysates of cells were immunoprecipitated with a Nef-specific mouse monoclonal anti-body obtained from Bernhard Maier (University of Virginia) (41). Samples were incubated for 2 h with 10g of antibody followed by protein G-Sepharose beads for another 2 h. Both incubations were performed rotating at 4°C. The samples were then washed three times with flotation buffer (described above) or wash buffer (0.2% deoxycholate, 0.5% Tween 20, 150 mM NaCl, 10 mM Tris-HCl, pH 7.2, and 5 mM EDTA) and one time with 1⫻TNE (10 mM Tris-HCl, pH 7.3, 150 mM NaCl, 5 mM EDTA). The beads were then boiled for 5 min in 1⫻SDS sample buffer and the supernatant was loaded onto a 12% polyacrylamide gel. Proteins were separated by SDS-PAGE followed by transfer to Immobilon (Millipore, Bedford, Mass.) for Western blotting. Blots were probed with rabbit anti-Nef serum and analyzed by enhanced chemiluminescence (ECL) (Amer-sham Biosciences, Little Chalfont, United Kingdom) with a sheep anti-rabbit secondary antibody coupled to horseradish peroxidase or, for quantitation, [125I]protein A and phosphorimager scanning. In some experiments, anti-trans-ferrin receptor monoclonal antibody or anti-Lck antibody was used as the pri-mary antibody.
Infectivity enhancement assay.The effect of Nef on first-round infectivity was assayed as previously described (10) with packaging cells (62), an HIV-based vector, and HeLa-CD4 cells as targets. Briefly, a 100-mm plate containing about 8⫻1055BD.1 cells producing HIV Gag-Pol and HIV envelope proteins was transfected with 5g of HIV vector lacking Nef, pTR167nef(⫺), that conferred hygromycin resistance; 2g of a vector expressing Tat; and 5g of a vector expressing Nef, an LAT-Nef fusion protein, or an empty control vector. Super-natants were harvested at 72 h posttransfection and cleared of cells by centrif-ugation.
To perform the infection of HeLa CD4 cells with the HIV vector stocks, 10-fold serial dilutions of the cleared supernatant were made into complete medium. DEAE-dextran was then added to each dilution at a concentration of 8g/ml to facilitate viral absorption and 1 ml of each dilution was added to 2⫻ 105cells, which had been subcultured the previous day. The virus was allowed to absorb for 6 h at 37°C, at which time 3 ml of complete medium was added and the incubation was continued. The medium was replaced 2 days later, with medium containing 200g of hygromycin B per ml. This medium was changed every 2 to 3 days. After about 14 days, the resultant colonies were fixed and stained with 0.5% crystal violet in 50% methanol. All experiments were carried out in duplicate.
Production, purification, and analysis of virions containing Nef, LAT-Nef, or LATAANef.Preparations of virions were generated by calcium phosphate
trans-fection (28) of 7.5⫻106293T cells in 15-cm dishes with 25g of pNL4-3 lacking a functionalnefgene, pNL4-3nef, and 10 or 15g of plasmids expressing Nef, NefG2A, LAT-Nef, and LATAANef. Six hours posttransfection, cells were washed and fresh medium containing the HIV-1 protease inhibitor saquinavir was added to the final concentration of 10M. Saquinavir was obtained from the NIH AIDS Research and Reference Reagent Program, Rockville, Md. At 36 h post-transfection, an additional 1M saquinavir was added to the medium.
Virus was harvested at 72 h after transfection by collection of the medium (20 ml), which was centrifuged at 2,500 rpm for 10 min to remove cellular debris. The supernatant was then subjected to ultracentrifugation through a cushion of 5 ml of 20% (wt/vol) sucrose (in phosphate-buffered saline) at 25,000 rpm in an SW28 rotor (Beckman) for 1.5 h. Pelleted virions were resuspended in 1 ml of phos-phate-buffered saline at 4°C overnight and further purified on a 20 to 60% sucrose gradient by ultracentrifugation at 36,000 rpm in an SW41Ti rotor (Beck-man) for 18 h. Fractions of 600l each were collected and analyzed for viral content by reverse transcription assay as previously described (71).
After identifying peak fractions of virus, the fractions were pooled and diluted with phosphate-buffered saline pelleted at 36,000 rpm for 2 h. Pelleted virions were resuspended in 100l of 10 mM Tris-HCl (pH 7.6) containing 0.5% Triton X-100, and stored at⫺20°C. The levels of p24 in concentrated virion stocks were determined by enzyme-linked immunosorbent assay (ELISA). Six nanograms of p24 equivalents was added to SDS loading buffer and used to analyze the levels of Nef proteins in purified virions by Western blot. As part of this analysis, cell lysates were also prepared at 36 h posttransfection. Total protein content of the
lysates was measured with the Bradford dye-binding assay (Bio-Rad, Hercules, Calif.), and the lysates were analyzed for Nef protein expression on Western blots as described above.
RESULTS
A small fraction of the Nef protein expressed in SupT1 cells targets to cellular membranes and it is entirely in the lipid raft
domain. Previous studies have shown an association of Nef
with cell membranes and that at least some of the protein associates with the lipid raft fraction (66, 67). This association is known to be dependent on N-terminal myristoylation, as well as basic amino acids residues found within the first 22 amino acids of Nef (68, 74). However, although few reports have quantitated the amount of Nef that is associated with the membrane fraction, in most cases it appears that a greater fraction of Nef was found associated with the cytoskeleton, or free in the cytosol, than the fraction found in membranes (3, 19, 23).
As a prelude to directly examining the role played by lipid raft association for Nef function, we initially wished to quan-titatively determine the fraction of Nef that was actually asso-ciated with membrane and lipid rafts in the SupT1 cells. To do this, vectors expressing Nef or a mutant form of Nef lacking the myristoylation signal were transfected into SupT1 cells by elec-troporation and extracts were analyzed on sucrose gradients.
Initially, non-detergent-treated extracts from cells transfect-ed with vectors expressing Nef or the NefG2Amyristoylation
mutant were analyzed by centrifugation in sucrose gradients with conditions which allowed for the flotation of membranes. Gradient fractions were collected and quantitatively immuno-precipitated with a Nef monoclonal antibody. The amount of Nef in each fraction was then determined by a quantitative Western blotting procedure with a Nef rabbit polyclonal anti-serum and [125I]protein A (Fig. 1A). As a control, we also
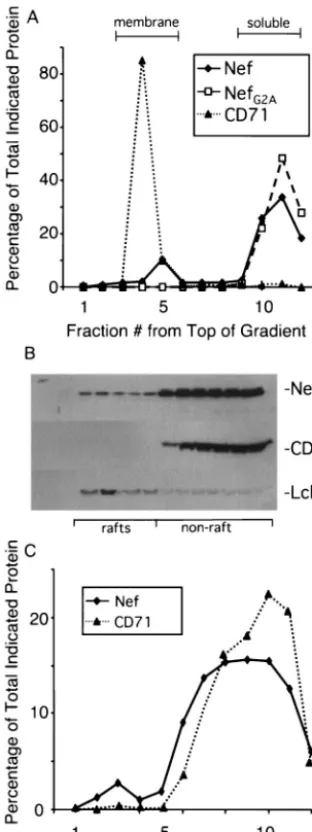
examined the distribution of the transferrin receptor (CD71), an integral membrane protein. The analysis revealed that ap-proximately 10% of wild-type Nef targeted to the membrane fraction, whereas the myristoylation mutant, NefG2A, was found
exclusively in the nonmembrane fractions. Virtually all of CD71 was found in the membrane fraction. This result confirms the notion that only a small amount of Nef targets to the mem-brane and that this targeting is dependent on N-terminal my-ristoylation.
We next determined the fraction of Nef present in the de-tergent insoluble raft fraction. Cells were lysed in Triton X-100-containing buffer at 4°C and the extract was centrifuged in a sucrose gradient in the cold leading to flotation of the raft fraction. Fractions were analyzed for Nef as before with quan-titative immunoprecipitation-Western blotting (Fig. 1B and C). Fractions were also probed directly for CD71 and Lck as markers of the solubilized membrane fraction and raft fraction, respectively. The analysis showed that approximately 8 to 10% of Nef targeted to the raft fractions. This result, taken together with the data from Fig. 1A, indicates that the majority of membrane-associated Nef is, more specifically, associated with lipid rafts.
Construction and expression of LAT-Nef fusion proteins.
Virtually all described functions of Nef are dependent on its myristoylation. This suggests that membrane association of Nef
on November 8, 2019 by guest
http://jvi.asm.org/
is critical for its activity. However, since lack of myristoylation completely abolishes the association of Nef with both raft and nonraft membrane domains, experiments with myristate-neg-ative Nef mutants have shed no light on whether the lipid raft association, rather than simply association with any portion of the plasma membrane by Nef, is critical for its function.
To directly address this issue, we designed vectors to express LAT-Nef fusion proteins containing 35 amino acids from the amino terminus of the linker of activated T-cell (LAT) protein. The T-cell LAT protein is an example of a protein that con-stitutively targets to rafts (75). The first 35 amino acids of LAT contain a classical type III transmembrane domain as well as two cysteines which, when palmitoylated, localize LAT primar-ily to the lipid raft membrane fraction. Mutation of these two cysteines to alanines causes LAT to be dissociated from rafts and to localize only to the nonraft membrane fraction (75). Other investigators have used this domain to target heterolo-gous T-cell proteins to raft domains (5).
Two fusion protein expression plasmids (LAT-Nef and LATAANef) were made by fusing either the wild-type LAT
N-terminal targeting domain or the LAT domain mutated in the two critical cysteines to a full-length Nef sequence (Fig. 2A). In each case, a Nef sequence lacking the myristoylation signal (NefG2A) was used as the fusion partner to ensure that
[image:4.603.84.240.85.501.2]any protein that might arise from an internal initiation would not become myristoylated.
Figure 2B shows a Western blot of extracts of SupT1 cells transfected with the fusion protein plasmids and a plasmid expressing wild-type Nef. The blot was developed with an anti-Nef serum. In the case of LAT-anti-Nef, two protein species were seen. By analogy with what has been shown for the full-length LAT protein, the upper band is likely the singly palmitoylated species, while the lower band is likely the doubly palmitoylated form of the protein (75). The unpalmitoylated fusion protein generated by the mutant, migrates at the same position as the singly palmitoylated form. This is consistent with the behavior of the unpalmitoylated LAT protein. Quantitation of the blot with [125I]protein A shows that expression levels of the
wild-type and mutated LAT fusion proteins are roughly equivalent (Fig. 2C). In comparison, there appears to be approximately half as much native Nef protein expressed under these condi-tions.
LAT-Nef and LATAANef target to the membrane
equiva-lently but only LAT-Nef targets to rafts.The localization of the
two fusion proteins was then determined by separating the membrane fractions of transiently transfected cells away from other components with sucrose gradient centrifugation. As controls, the localization of NefG2Aand CD71 was also
ana-lyzed. For each Nef protein, fractions from the gradient were first immunoprecipitated and then analyzed by Western blot. CD71 was analyzed by direct Western blotting. Quantitation of each blot was carried out with [125I]protein A. The results are
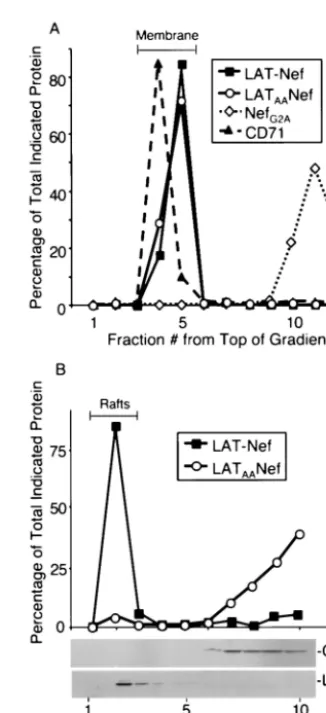
plotted in Fig. 3A, which demonstrates that virtually 100% of each fusion protein was found in the membrane fraction.
We next analyzed the fraction of each fusion protein that was present in lipid rafts with the same conditions described above for wild-type Nef (see Fig. 1B and C). As controls the distribution of Lck and CD71 was also analyzed. The results presented in Fig. 3B show that LAT-Nef was found almost exclusively in the raft fractions, whereas LATAANef was found
FIG. 1. Subcellular targeting of Nef. (A) Membrane localization; 75g of pA-Nef or pA-NefG2Aexpression vector was transfected by
electroporation into 1.5⫻108SupT1 cells. Cells were lysed after 48 h
by freeze-thawing, and lysates were layered at the bottom of a sucrose gradient and spun overnight as described in Materials and Methods. Fractions were then solubilized and subjected to immunoprecipitation with a mouse anti-Nef antibody, followed by Western blot analysis with rabbit anti-Nef serum. For CD71 detection, fractions were analyzed by direct Western blot with a monoclonal anti-CD71 antibody. In both cases, development was performed with [125I]protein A. The
percent-age of the total protein in each fraction was determined by dividing the phosphorimager units of the band in that fraction by the combined total value of that band in all fractions. (B and C) Raft localization; 108
SupT1 cells were transfected with 50 g of the pA-Nef expression vector as in panel A and harvested after 48 h. Cells were lysed in Triton X-100-containing buffer, and lysates were layered at the bottom of a sucrose gradient and spun overnight as described in Materials and Methods. The gradient was fractionated and analyzed for Nef by im-munoprecipitation and Western blot or by direct Western blot for Lck and CD71. Panel B is a Western blot probed for the indicated protein and developed by ECL. Panel C shows the same gel quantitated with [125I]protein A as described for panel A, and fractions were analyzed
for Nef and CD71 as in panel A.
on November 8, 2019 by guest
http://jvi.asm.org/
predominantly in the nonraft soluble fractions. The controls, Lck and CD71, distributed as expected in the raft and nonraft fractions, respectively. The results of Fig. 4A and B taken together show that the LAT targeting strategy worked as an-ticipated.
LAT-Nef is more efficient than LATAANef at
downmodulat-ing CD4 and class I MHC.We then evaluated the functional
activity of LAT-Nef or LATAANef for CD4 and class I MHC
downmodulation in both time course and dose-response ex-periments. For comparison, the activities of Nef and NefG2A
were also evaluated.
For the time course experiment, 10 g of an expression plasmid for either the LAT-Nef, LATAANef, Nef, NefG2A, or
control empty vector was transiently transfected into 2⫻107
SupT1 cells, together with 2g of a plasmid expressing green fluorescent protein (GFP). Cells were then harvested and stained with phycoerythrin (PE)-labeled anti-CD4 or anti-class I MHC (HLA A2.1) monoclonal antibodies at various times after transfection. The mean fluorescence intensity of CD4 and class I MHC in the GFP-positive population (usually about 30 to 40% of the live cells) was then measured by flow cytometry, as this population represented the successfully transfected cells (see Fig. 4A and B). Cells were also transfected with various amounts of the various Nef expression plasmids (2.5 to 20g) and stained at 24 h posttransfection with CD4-PE or anti-class I MHC (HLA 2.1)-PE monoclonal antibodies. These cells were analyzed similarly (Fig. 4C and D).
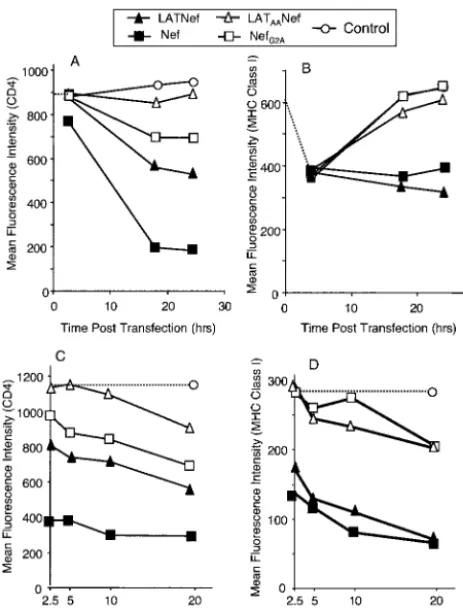
[image:5.603.102.231.61.464.2]CD4 was very efficiently downmodulated over time in cells transfected with wild-type Nef in comparison to those trans-fected with the control vector in which CD4 expression re-mained unchanged. (Fig. 4A). We consistently observed CD4 downmodulation by Nef as early as 2 to 4 h posttransfection.
FIG. 2. LAT-Nef and LATAA-Nef fusion proteins: construction
and expression. (A) Schematic representation of LAT-Nef and LATAA
-Nef and their expected subcellular destination. These constructs were built into vectors with expression driven by either the simian cytomeg-alovirus immediate-early promoter or the chicken actin promoter; 2⫻ 107SupT1 cells were transfected with 10g of the pA-based vectors
expressing Nef, LAT-Nef, or LATAA-Nef. Lysates were then made and
normalized for total protein content before being subjected to immu-noprecipitation with a monoclonal anti-Nef antibody and Western blot analysis with a rabbit anti-Nef serum. The Western blot was developed by ECL (B) or [125I]protein A (C). In panel C the amount of protein
shown for LAT-Nef is a summation of both bands representing the partially and fully palmitoylated forms of the protein.
FIG. 3. LAT-Nef and LATAA-Nef target to rafts and nonraft
mem-brane, respectively; 108SupT1 cells were transfected with 40g of
pA-NefG2A, pA-LAT-Nef, or pA-LATAANef expression vector.
Ly-sates were made and subjected to membrane (A) and raft (B) isolation and gradient analysis as described for wild-type Nef in Fig. 1.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.603.337.500.322.679.2]By 24 h posttransfection, the mean fluorescence intensity of the GFP-positive population was decreased to 20% of the value of the control. LAT-Nef also downmodulated CD4, al-though somewhat less well. In this case, the mean fluorescence intensity of the transfected cell population was 55% of the control value by 24 h posttransfection. In contrast to these two proteins, 10 g of LATAANef failed to downmodulate CD4
(Fig. 4A). In this case, the mean fluorescence intensity of the transfected cell population was similar to the mock-transfected population over the entire 24-h course. The same relative func-tionalities of these proteins in downmodulating CD4 were seen in the dose-response experiment (Fig. 4C), although at the highest concentration of LATAANef, a slight downmodulation
of CD4 was apparent.
Surprisingly, some CD4 downmodulation activity was also observed with NefG2A, although this protein clearly failed to
target to membranes (Fig. 1A). A decrease of the mean fluo-rescence intensity to 74% of the control values was observed. This is consistent with results from other laboratories which
have demonstrated a residual activity for CD4 downmodula-tion by this protein (1).
Class I MHC proteins were also clearly downmodulated by wild-type Nef and LAT-Nef, though less dramatically than CD4 (Fig. 4B). In the time course experiment there was an initial apparent decrease in signal for class I MHC following electroporation with all of the plasmids. This is likely due to the transfection procedure, since it was also observed with the empty vector. For the cells transfected with Nef and LAT-Nef, the mean fluorescence intensity remained at this low level out to 24 h. In contrast, the mean fluorescence intensity value re-bounded in cells transfected with either LATAANef or NefG2A,
suggesting that these two proteins are ineffective at down-modulating class I MHC. This was further confirmed in the dose-response experiment (Fig. 4D). Mean fluorescence inten-sity values measured at 24 h posttransfection showed clearly that LAT-Nef functioned more efficiently than LATAANef in
class I MHC downmodulation. Interestingly, in contrast to the results for CD4 downmodulation, LAT-Nef displayed activity similar to that of the wild-type Nef protein whereas the LATAANef and NefG2Aproteins showed equal minimal
activ-ity.
To evaluate the reproducibility of these results, 10 experi-ments for class I MHC downmodulation and 9 for CD4 down-modulation at 24 h posttransfection were performed. The re-sults are compiled in Fig. 5, where the activity of each fusion protein is compared to the activity of wild-type Nef, which has been set at 100%. For class I MHC downmodulation, the LAT-Nef fusion protein behaved nearly as well as wild-type Nef (95%), while LATAANef only worked 38% as well. For
CD4, LAT-Nef downmodulated the molecule to 59%, while LATAANef worked only 16% as well as the wild-type Nef
protein.
Mutations in Nef that affect CD4 or class I MHC down-modulation have the same phenotype in the context of LAT-Nef.LAT-Nef is completely membrane associated while only a small fraction of wild-type Nef exists in membranes. This raises
FIG. 4. LAT-Nef downmodulates CD4 and class I MHC more ef-ficiently than LATAA-Nef; 2⫻107SupT1 cells were transfected with
Nef, NefG2A, LAT-Nef, or LATAA-Nef, expression vectors along with
[image:6.603.48.280.71.375.2]2 g of a GFP expression vector. In panels A and B, cells were transfected with 10g of each vector and incubated with PE-labeled antibody at the indicated times. In panels C and D, cells were trans-fected with the indicated amount of expression vector and incubated with PE-labeled antibody after 24 h. Cells were stained with either CD4-PE (A and C) or class I MHC A2.1-PE (B and D) antibodies and analyzed by flow cytometry. In each case, the data plotted are the CD4 or class I MHC mean fluorescence intensity of the GFP-positive gated population.
FIG. 5. Functional differences between LAT-Nef and LATAA-Nef
are highly reproducible. For CD4 and class I MHC downmodulation, 2g of GFP expression vector was cotransfected together with 10g of pA, pA-Nef, pA-LAT-Nef, or pA-LATAA-Nef expression vector
into 2⫻107SupT1 cells. Cells were stained with antibodies against
CD4 or class I MHC after 24 h. The LAT-Nef and LATAA-Nef CD4
and class I MHC activities of the GFP-positive populations were de-termined by flow cytometry. Results from nine measurements of CD4 downmodulation and 10 for class I MHC downmodulation are com-piled.
on November 8, 2019 by guest
http://jvi.asm.org/
the possibility that the pathways used for downmodulation of CD4 and class I MHC by LAT-Nef and Nef are different. To examine this, we utilized a series of well-characterized muta-tions in Nef that previously had been shown to differentially affect the two pathways (51, 54). The mutations chosen were D123G, previously shown to inhibit both CD4 and class I MHC downmodulation (39); L164, L165/AA (LL/AA), previously shown inhibit only CD4 downmodulation (7, 29); and E62, E63, E64, and E65/AAAA (EEEE/AAAA), previously shown to inhibit only class I MHC downmodulation (31).
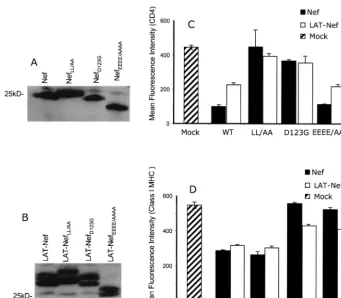
All of the above mutations were introduced into plasmids expressing either Nef, LAT-Nef, or LATAANef. These were
then transfected into SupT1 cells and analyzed for protein expression. Figure 6A and B show that good levels of expres-sion were obtained for all of the proteins containing the mu-tations in the context of Nef or LAT-Nef. Equivalent expres-sion was also obtained with the LATAANef constructs (data
not shown). We also noted that each of the different mutations caused a shift in the migration position of the Nef or LAT-Nef protein on the SDS-polyacrylamide gel. They were also seen in the LATAANef constructs (data not shown). The reason for
these shifts is not known. They may reflect conformational changes or differential posttranslational modifications due to the mutations. Interestingly, both Nef and LAT-Nef proteins are shifted similarly for each mutation.
Figures 6C and D show the results obtained for CD4 and class I MHC downmodulation by the mutations in the context of the Nef and LAT-Nef proteins. The data demonstrate that the mutations behaved as expected and similarly in both pro-teins. In each case, LL/AA nearly completely abolished CD4 downmodulation but had no effect on class I MHC downmodu-lation; D123G dramatically reduced both CD4 and class I MHC downmodulation; and EEEE/AAAA primarily affected only class I MHC downmodulation. In the context of the non-raft-associated LATAANef protein, all of the mutations had
little effect, in that all of the LATAANef proteins were still
severely impaired for downmodulation of either CD4 or class I MHC (data not shown). These results strongly suggest that the pathways of downmodulation for both CD4 and class I MHC by LAT-Nef are the same as those used by Nef.
LAT-Nef is more efficient than LATAANef in promoting
in-fectivity.It has been well described by several laboratories that
[image:7.603.118.459.66.364.2]virus particles produced in the presence of Nef have a greater infectivity than particles produced in the absence of Nef (11, 45, 57). This effect of Nef appears to be independent of the increase in infectivity that is attributable to CD4 downmodu-lation, because it can be observed in cells lacking CD4 (10, 11, 45). We have previously described a packaging cell and HIV vector system where this infectivity enhancement can be readily measured in a single round infectivity assay (10).
FIG. 6. Analysis of CD4 and class I MHC downmodulation by LAT-Nef and Nef containing mutations that affect these functions. (A and B) Vectors expressing the indicated proteins were transfected into SupT1 cells. Lysates were made and analyzed by Western blotting as described in Materials and Methods. The blots were probed with a rabbit polyclonal serum directed against Nef and developed by ECL. (C and D) SupT1 cells were transfected with Nef or LAT-Nef expression vector carrying the indicated mutation, together with 2g of a GFP expression vector. At 24 h posttransfection, cells were harvested and stained with either an anti CD4-PE antibody (C) or an anti-class I MHC A2.1-PE antibody (D). Samples were then analyzed by flow cytometry. In each case, the data plotted are the CD4 or class I MHC mean fluorescence intensities of the GFP-positive gated population. The sample marked mock underwent the transfection procedure with empty pA vector.
on November 8, 2019 by guest
http://jvi.asm.org/
To measure infectivity enhancement of the LAT-Nef fusion proteins, we performed an experiment in which 8⫻1055BD.1
packaging cells (62) stably expressing HIVBH10 Gag-Pol and
HIVHXB2envelope protein were cotransfected with 5g of an
HIV vector containing the hygromycin resistance gene and 5, 10, or 15g of the appropriate Nef expression plasmid. Su-pernatant was harvested at 72 h posttransfection and HIV p24 was measured by ELISA. p24 values in the supernatant were all similar and within a standard deviation of about 5%.
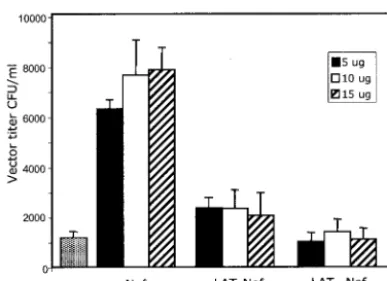
Equal amounts of the supernatants were then used to infect HeLa CD4 cells. Cells were selected for 14 days with hygro-mycin and colonies were counted. The experiment was per-formed in duplicate, and the results are plotted in Fig. 7. The results show that wild-type Nef increased infectivity about 6- to 7-fold at all concentrations of Nef plasmid transfected, while LAT-Nef increased infectivity about 2.5-fold. The LATAANef
mutant did not increase infectivity at all. Although the effects seen with the LAT Nef construct were small (only 2.5 times background compared to 7-fold for Nef), the results were highly reproducible and at the concentrations of DNA used in these experiments appeared to be independent of variations in DNA concentration, suggesting that saturating amounts of Nef or LAT-Nef were present.
LAT-Nef is incorporated in virions more efficiently than LATAANef.Several reports have shown that Nef is present in
HIV particles (9, 44, 47, 68, 69) and that incorporation is dependent on the N-terminal myristate residue (68). Thus, raft association by Nef might be a determinant involved in directing the protein to the virion. Raft association by Nef would likely place it at the site of viral assembly, since studies have shown that Gag and Gag-Pol localize to rafts and appear to initiate viral assembly there (46). However, it is also likely that Nef must leave the membrane at some point during viral assembly to be internalized, since Nef ultimately ends up in the viral core (35), possibly in association with the viral ribonuclear protein complex (20). Thus, if raft association is a determinant
in-volved in directing Nef to the particle, other factors must also be involved in determining the final localization of Nef in articles.
To analyze the ability of LAT-Nef and LATAANef to be
incorporated into particles, we cotransfected plasmids express-ing these proteins together with a proviral clone (pNL4-3) lacking Nef in 293T cells. As controls, plasmids expressing wild-type Nef and NefG2A were also cotransfected with the
proviral clone. Since Nef is readily cleaved by the viral pro-tease, the experiment was performed in the presence of sa-quinavir, a viral protease inhibitor. This allowed us to readily determine primary expression and incorporation levels without subsequent alterations that might be due to protease cleavage. Cell lysates were prepared at the optimal time of protein syn-thesis (36 h posttransfection) to determine relative expression levels of the various Nef proteins. Viral particles were isolated from the supernatant at 72 h posttransfection. After normal-ization for total protein or p24, lysates and viral particles were analyzed by quantitative Western blot with [125I]protein A to
[image:8.603.66.260.67.208.2]determine the level of Nef or LAT-Nef protein present. Figure 8 shows the results of this analysis.
Figure 8A shows the levels of expression of the various Nef proteins in the transfected cell. As before, LAT-Nef migrated as a double band, reflective of the singly and doubly palmitoy-lated species (75). For all Nef proteins expressed, each indi-vidual protein band was present at approximately equal levels (within 1.4-fold). Analysis of the virions showed that all sam-ples contained approximately equal levels of Pr55gagas
expect-ed (Fig. 8B, upper panel); however, incorporation of NefG2A
and LATAANef was severely impaired relative to Nef. On the
other hand, LAT-Nef was incorporated to a much greater extent, with the fully palmitoylated form being incorporated better than the singly palmitoylated form. Thus, these results strongly suggest that raft association, and not just membrane association, by Nef is an important determinant for particle association.
DISCUSSION
In this study, we have shown that about 10% of the wild-type Nef protein produced in SupT1 cells is membrane-associated and that virtually all of the membrane-associated protein is found in raft microdomains. The amount of Nef associated with the total membrane fraction is similar to the previously published estimates from other laboratories (3, 23, 68). Previ-ous reports have also found Nef associated with rafts, but these studies did not attempt to determine the distribution of Nef between the nonraft membrane fraction and rafts, as we have done (66, 67, 76). By retargeting Nef, with LAT-Nef chimeras, to either the raft or nonraft membrane fraction, we have now been able to conclude that raft association, rather than simply membrane association, is critical for Nef to efficiently perform three of its well recognized functions.
Our study showed that the LAT-Nef chimera that targeted to rafts was more efficient at CD4 and class I MHC down-modulation and first-round infectivity enhancement than the chimera that targeted to the nonraft membrane. In addition, differences in how well the raft-targeted chimera worked com-pared to wild-type Nef were also observed. For the same amount of transfected DNA, LAT-Nef had an efficiency
sim-FIG. 7. LAT-Nef enhances virion infectivity greater than LATAA
-Nef. Virus vector was harvested at 3 days posttransfection from 5BD.1 cells transfected with pTR167 Nef(⫺) and 5, 10, or 15g of pCMV-Nef, pCMV LAT-pCMV-Nef, or pCMVLATAA-Nef as indicated. For the
control sample 5g of pCMV containing no insert was utilized. In all cases a vector that expressed Tat (pCMVTat) was also added. The supernatant was then used to infect HeLa CD4 cells, and hygromycin-resistant colonies were counted after 14 days. Vector titer was calcu-lated as the number of hygromycin-resistant HeLa CD4 colonies formed per milliliter of supernatant. Each bar shows the data from independent duplicate experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
ilar to that of wild-type Nef for class I MHC downmodulation, about 60% that of wild-type Nef for CD4 downmodulation, and about 30% that of wild-type Nef for infectivity enhance-ment.
In considering the functional differences between the chi-meric LAT-Nef protein and wild-type Nef, we cannot rule out the possibility that attaching an extension at the amino termi-nus affects protein conformation in a way that differentially impacts its ability to perform each function. However, we be-lieve that a more likely explanation for the different efficiencies observed relates to the fact that the LAT-Nef protein is essen-tially 100% membrane associated. The LAT-Nef protein is firmly anchored in membranes via a transmembrane domain, compared to wild-type Nef, which is only 10% membrane as-sociated and has the presumed flexibility to go in and out of the membrane. Following this logic, our data would suggest that non-membrane-associated Nef may play some role in CD4 downmodulation and enhancement of viral infectivity, while class I MHC downmodulation by Nef may be an exclusively membrane-bound process. In this regard, it is interesting that the nonmyristoylated form of Nef (NefG2A), although highly
impaired, still downmodulated CD4 in a dose- and time-de-pendent manner, but had no effect on class I MHC downmodu-lation.
It has been shown previously that CD4 and class I MHC downmodulation by Nef occur through different pathways and that different regions of Nef mediate these functions and in-fectivity enhancement (27, 42). While our data clearly support raft involvement in all three processes, at this time we can only speculate on the role played by rafts for each of these func-tions. The reasons for the importance of raft targeting could include the fact that such targeting would allow a close geo-graphic proximity between Nef and its target molecules, a clustering of Nef with intermediary adaptor molecules, or the maintenance of a Nef structure that would allow it to interact appropriately with other molecules.
It has been well established that the CD4 protein is found associated with lipid rafts (22), and there is an extensive liter-ature which suggests that CD4 downmodulation by Nef occurs primarily at the cell surface by endocytosis through clathrin-coated pits (21, 73). In this model it has been suggested that Nef interacts directly with the cytoplasmic tail of CD4 and links it to the endocytosis machinery via adaptins and/or the vacu-olar ATPase (40, 49). Since internalization of raft-associated proteins may involve a clathrin-independent route, it may be that a key role of Nef is to target the clathrin-mediated endo-cytosis machinery to a location normally devoid of clathrin. Although nuclear magnetic resonance studies have shown that a full-length recombinant Nef protein interacts directly with a synthetic peptide derived from the cytoplasmic tail of CD4, Nef-CD4 binding has been difficult to demonstrate (53). It may be that a combination of CD4 binding and raft association by Nef is key to the forming of a stable Nef-CD4 complex.
[image:9.603.81.247.68.506.2]Recent data have suggested that downmodulation of class I MHC by Nef in T cells may primarily be the result of a direct binding of Nef to the class I MHC cytoplasmic tail and a blockage of transport to the cell surface of newly synthesized class I MHC molecules (34, 63, 72). Other mechanisms such as increased clathrin-independent endocytosis and retargeting through such molecules as PACS-1 could also play some role in
FIG. 8. Incorporation of Nef proteins into virus particles produced in 293T cells. 293T cells were contransfected with the pNL4-3 proviral clone lacking Nef and vectors expressing either Nef, or NefG2A(10g)
and LAT-Nef or LATAA-Nef (15g). (A) Cells were harvested at 36 h
posttransfection, and lysates were made and analyzed by Western blotting as described in Materials and Methods. The blots were probed with a rabbit polyclonal serum directed against Nef and developed with [125I]protein A. The amount of radioactivity in each band was then
measured with a phosphorimager. The intensity of each band relative to the band from the sample transfected with the Nef expression vector is shown. (B) Virus particles were harvested, as described in Materials and Methods, at 72 h posttransfection from duplicate cultures that were transfected at the same time as those in part A. The particles were then lysed and subjected to Western blotting after normalizing for p24 content as measured by ELISA. The upper panel shows a portion of the Western blot that was probed with an anti-p24 mono-clonal antibody capable of recognizing Pr55gag. The blot was developed
with ECL. The bottom panel shows a portion of the blot probed with a rabbit polyclonal serum directed against Nef. The blot was developed with [125I]protein A. The intensity of each band relative to the band
from the sample transfected with the Nef expression vector is shown after normalization for the slight differences in expression that were determined in part A.
on November 8, 2019 by guest
http://jvi.asm.org/
T cells (52). Whatever the mechanism, accumulation of class I MHC molecules in the Golgi seems to result. Interestingly, this is the cellular compartment where rafts are first thought to form. Thus, Nef association with rafts in the Golgi could trap class I MHC molecules there, either alone or in conjunction with other proteins.
Our results demonstrate that LATAANef is very poorly
in-corporated into viruses particles relative to either LAT-Nef or Nef. Since LATAANef fails to target to rafts while the other
two proteins do, this indicates a role for raft targeting in Nef particle incorporation. However, the association of Nef with rafts cannot be the only factor that mediates this incorporation. If this were the case, more LAT-Nef should be incorporated into particles than Nef, since 100% of it is raft associated compared to only a small fraction of Nef (see Fig. 3). In actuality, after normalization for expression, we found that the two species of LAT-Nef were only incorporated about 40% and 17% as well as Nef. One possible explanation for this finding might be that tight anchoring to the membrane through the transmembrane domain of LAT-Nef is actually detrimental for incorporation and that Nef may have to come off of the membrane at some point, perhaps to interact with Gag or Gag-Pol, to be packaged efficiently. The finding that both pro-tease-cleaved and uncleaved Nef is associated with the viral ribonucleoprotein in virions (20), rather than membrane, is consistent with this idea.
It should also be noted that a role for particle-associated Nef in infectivity enhancement has yet to be directly established, so it is not clear that the infectivity differences seen between raft-associated LAT-Nef and non-raft-associated LATAANef
can be explained simply by the incorporation differences. Ex-isting data do not allow distinction between models in which Nef increases particle infectivity by acting in the producer cell to alter the viral particle so that it functions more efficiently in the subsequent round of infection and mechanisms by which Nef itself functions as part of the incoming preintegration complex. In either case, however, raft association by Nef would place it at the site of viral assembly, since studies have shown that Gag and Gag-Pol localize to rafts and appear to initiate viral assembly there (46).
One study has suggested that particle infectivity enhance-ment by Nef may be related to Nef-induced alterations of cholesterol and ganglioside GM1 levels in particles (76). The study also showed that infectivity enhancement by Nef was abolished when cells were depleted of cholesterol, through the use of either biosynthetic inhibitors or cells defective in cho-lesterol biosynthesis. The authors interpreted these results to mean that under conditions of cholesterol depletion, Nef could no longer induce a change in the composition of the virus membrane. However, these results are also consistent with the view that rafts function as staging areas for assembly and that if they are disrupted, Nef is no longer able to localize to regions where there is a high concentration of newly assembling virus particles and exert its effect.
In recent years, lipid rafts have been recognized as central organizational platforms in the T-cell membrane, yet many details of how they form and function are still lacking. Con-tinuing studies of how HIV Nef interacts with these domains will likely generate more knowledge about this subject. Thus, studies of HIV and its accessory genes such asnefcontinue to
provide insight into fundamental processes operating within the cell. Understanding these processes in more detail will also provide important information for the discovery and design of future anti-HIV drugs.
ACKNOWLEDGMENTS
We thank Joy Morgenegg for expert technical assistance, Greg Singer for participating in some of the early infectivity experiments, and Bernhard Maier for Nef reagents, helpful discussions, and helping to establish the T-cell line transfection protocol.
This work used material from the AIDS Research and Reference Reagent Program (National Institutes of Health, Bethesda, Md.) and was supported by NIH grants AI34721 to M.-L.H. and AI47008 to D.R. Salary support for M.-L.H. and D.R. was provided by the Charles H. Ross, Jr., and Myles H. Thaler endowments at the University of Virginia.
REFERENCES
1. Aiken, C., J. Konner, N. R. Landau, M. E. Lenburg, and D. Trono.1994. Nef induces CD4 endocytosis: requirement for a critical dileucine motif in the membrane-proximal CD4 cytoplasmic domain. Cell76:853–864.
2. Aldrovandi, G. M., L. Gao, G. Bristol, and J. A. Zack.1998. Regions of human immunodeficiency virus type 1nefrequired for function in vivo. J. Virol.72:7032–7039.
3. Allan, J. S., J. E. Coligan, T. H. Lee, M. F. McLane, P. J. Kanki, J. E. Groopman, and M. Essex.1985. A new HTLV-III/LAV encoded antigen detected by antibodies from AIDS patients. Science230:810–813. 4. Arold, S. T., and A. S. Baur.2001. Dynamic Nef and Nef dynamics: how
structure could explain the complex activities of this small HIV protein. Trends Biochem. Sci.26:356–363.
5. Boerth, N. J., J. J. Sadler, D. E. Bauer, J. L. Clements, S. M. Gheith, and G. A. Koretzky.2000. Recruitment of SLP-76 to the membrane and glyco-lipid-enriched membrane microdomains replaces the requirement for linker for activation of T cells in T-cell receptor signaling. J. Exp. Med.192:1047– 1058.
6. Brambilla, A., L. Turchetto, A. Gatti, C. Bovolenta, F. Veglia, E. Santa-gostino, A. Gringeri, M. Clementi, G. Poli, P. Bagnarelli, and E. Vicenzi.
1999. Defective nef alleles in a cohort of hemophiliacs with progressing and nonprogressing HIV-1 infection. Virology259:349–368.
7. Bresnahan, P. A., W. Yonemoto, S. Ferrell, D. Williams-Herman, R. Geleziu-nas, and W. C. Greene.1998. A dileucine motif in HIV-1 Nef acts as an internalization signal for CD4 downregulation and binds the AP-1 clathrin adaptor. Curr. Biol.8:1235–1238.
8. Brown, D. A., and E. London.1998. Functions of lipid rafts in biological membranes. Annu. Rev. Cell Dev. Biol.14:111–136.
9. Bukovsky, A. A., T. Dorfman, A. Weimann, and H. G. Gottlinger.1997. Nef association with human immunodeficiency virus type 1 virions and cleavage by the viral protease. J. Virol.71:1013–1018.
10. Chazal, N., G. Singer, C. Aiken, M. L. Hammarskjold, and D. Rekosh.2001. Human immunodeficiency virus type 1 particles pseudotyped with envelope proteins that fuse at low pH no longer require Nef for optimal infectivity. J. Virol.75:4014–4018.
11. Chowers, M. Y., M. W. Pandori, C. A. Spina, D. D. Richman, and J. C. Guatelli.1995. The growth advantage conferred by HIV-1 nef is determined at the level of viral DNA formation and is independent of CD4 downregu-lation. Virology212:451–457.
12. Chowers, M. Y., C. A. Spina, T. J. Kwoh, N. J. Fitch, D. D. Richman, and J. C. Guatelli.1994. Optimal infectivity in vitro of human immunodeficiency virus type 1 requires an intactnefgene. J. Virol.68:2906–2914.
13. Collins, K. L., B. K. Chen, S. A. Kalams, B. D. Walker, and D. Baltimore.
1998. HIV-1 Nef protein protects infected primary cells against killing by cytotoxic T lymphocytes. Nature391:397–401.
14. Craig, H. M., T. R. Reddy, N. L. Riggs, P. P. Dao, and J. C. Guatelli.2000. Interactions of HIV-1 nef with the mu subunits of adaptor protein complexes 1, 2, and 3: role of the dileucine-based sorting motif. Virology271:9–17. 15. Deacon, N. J., A. Tsykin, A. Solomon, K. Smith, M. Ludford-Menting, D. J.
Hooker, D. A. McPhee, A. L. Greenway, A. Ellett, C. Chatfield, et al.1995. Genomic structure of an attenuated quasi species of HIV-1 from a blood transfusion donor and recipients. Science270:988–991.
16. Doms, R. W., and D. Trono.2000. The plasma membrane as a combat zone in the HIV battlefield. Genes Dev.14:2677–2688.
17. Erdtmann, L., K. Janvier, G. Raposo, H. M. Craig, P. Benaroch, C. Berlioz-Torrent, J. C. Guatelli, R. Benarous, and S. Benichou.2000. Two indepen-dent regions of HIV-1 Nef are required for connection with the endocytic pathway through binding to the mu 1 chain of AP1 complex. Traffic1:871– 883.
18. Fackler, O. T., and A. S. Baur.2002. Live and let die: Nef functions beyond HIV replication. Immunity16:493–497.
on November 8, 2019 by guest
http://jvi.asm.org/
19. Fackler, O. T., N. Kienzle, E. Kremmer, A. Boese, B. Schramm, T. Klimkait, C. Kucherer, and N. Mueller-Lantzsch.1997. Association of human immu-nodeficiency virus Nef protein with actin is myristoylation dependent and influences its subcellular localization. Eur. J. Biochem.247:843–851. 20. Forshey, B. M., and C. Aiken.2003. Disassembly of human
immunodefi-ciency virus type 1 cores in vitro reveals association of nef with the subviral ribonucleoprotein complex. J. Virol.77:4409–4414.
21. Foti, M., A. Mangasarian, V. Piguet, D. P. Lew, K. H. Krause, D. Trono, and J. L. Carpentier.1997. Nef-mediated clathrin-coated pit formation. J. Cell Biol.139:37–47.
22. Foti, M., M. A. Phelouzat, A. Holm, B. J. Rasmusson, and J. L. Carpentier.
2002. p56Lck anchors CD4 to distinct microdomains on microvilli. Proc. Natl. Acad. Sci. USA99:2008–2013.
23. Franchini, G., M. Robert-Guroff, J. Ghrayeb, N. T. Chang, and F. Wong-Staal.1986. Cytoplasmic localization of the HTLV-III 3⬘orf protein in cultured T cells. Virology155:593–599.
24. Garcia, J. V., and A. D. Miller.1991. Serine phosphorylation-independent downregulation of cell-surface CD4 by nef. Nature350:508–511. 25. Geyer, M., O. T. Fackler, and B. M. Peterlin.2001. Structure–function
relationships in HIV-1 Nef. EMBO Rep.2:580–585.
26. Geyer, M., H. Yu, R. Mandic, T. Linnemann, Y. H. Zheng, O. T. Fackler, and B. M. Peterlin.2002. Subunit H of the V-ATPase binds to the medium chain of adaptor protein complex 2 and connects Nef to the endocytic machinery. J. Biol. Chem.277:28521–28529.
27. Goldsmith, M. A., M. T. Warmerdam, R. E. Atchison, M. D. Miller, and W. C. Greene.1995. Dissociation of the CD4 downregulation and viral infectivity enhancement functions of human immunodeficiency virus type 1 Nef. J. Virol.69:4112–4121.
28. Graham, F. L., and A. J. van der Eb.1973. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology52:456–467.
29. Greenberg, M., L. DeTulleo, I. Rapoport, J. Skowronski, and T. Kirch-hausen.1998. A dileucine motif in HIV-1 Nef is essential for sorting into clathrin-coated pits and for downregulation of CD4. Curr. Biol.8:1239–1242. 30. Greenberg, M. E., S. Bronson, M. Lock, M. Neumann, G. N. Pavlakis, and J. Skowronski.1997. Co-localization of HIV-1 Nef with the AP-2 adaptor protein complex correlates with Nef-induced CD4 down-regulation. EMBO J.16:6964–6976.
31. Greenberg, M. E., A. J. Iafrate, and J. Skowronski.1998. The SH3 domain-binding surface and an acidic motif in HIV-1 Nef regulate trafficking of class I MHC complexes. EMBO J.17:2777–2789.
32. Hoessli, D. C., S. Ilangumaran, A. Soltermann, P. J. Robinson, B. Borisch, and D. Nasir Ud.2000. Signaling through sphingolipid microdomains of the plasma membrane: the concept of signaling platform. Glycoconj. J.17:191– 197.
33. Janvier, K., H. Craig, S. Le Gall, R. Benarous, J. Guatelli, O. Schwartz, and S. Benichou.2001. Nef-induced CD4 downregulation: a diacidic sequence in human immunodeficiency virus type 1 Nef does not function as a protein sorting motif through direct binding to beta-COP. J. Virol.75:3971–3976. 34. Kasper, M. R., and K. L. Collins.2003. Nef-mediated disruption of HLA-A2
transport to the cell surface in T cells. J. Virol.77:3041–3049.
35. Kotov, A., J. Zhou, P. Flicker, and C. Aiken.1999. Association of Nef with the human immunodeficiency virus type 1 core. J. Virol.73:8824–8830. 36. Lama, J., A. Mangasarian, and D. Trono.1999. Cell-surface expression of
CD4 reduces HIV-1 infectivity by blocking Env incorporation in a Nef- and Vpu-inhibitable manner. Curr. Biol.9:622–631.
37. Learmont, J., B. Tindall, L. Evans, A. Cunningham, P. Cunningham, J. Wells, R. Penny, J. Kaldor, and D. A. Cooper.1992. Long-term symptomless HIV-1 infection in recipients of blood products from a single donor. Lancet
340:863–867.
38. Le Gall, S., F. Buseyne, A. Trocha, B. D. Walker, J. M. Heard, and O. Schwartz.2000. Distinct trafficking pathways mediate Nef-induced and clath-rin-dependent major histocompatibility complex class I down-regulation. J. Virol.74:9256–9266.
39. Liu, L. X., N. Heveker, O. T. Fackler, S. Arold, S. Le Gall, K. Janvier, B. M. Peterlin, C. Dumas, O. Schwartz, S. Benichou, and R. Benarous.2000. Mutation of a conserved residue (D123) required for oligomerization of human immunodeficiency virus type 1 Nef protein abolishes interaction with human thioesterase and results in impairment of Nef biological functions. J. Virol.74:5310–5319.
40. Lu, X., H. Yu, S. H. Liu, F. M. Brodsky, and B. M. Peterlin.1998. Interac-tions between HIV1 Nef and vacuolar ATPase facilitate the internalization of CD4. Immunity8:647–656.
41. Lucchiari, M., G. Niedermann, C. Leipner, A. Meyerhans, K. Eichmann, and B. Maier.1994. Human immune response to HIV-1-Nef. I. CD45RO-T lymphocytes of non-infected donors contain cytotoxic T lymphocyte precur-sors at high frequency. Int. Immunol.6:1739–1749.
42. Mangasarian, A., V. Piguet, J. K. Wang, Y. L. Chen, and D. Trono.1999. Nef-induced CD4 and major histocompatibility complex class I (MHC-I) down-regulation are governed by distinct determinants: N-terminal alpha helix and proline repeat of Nef selectively regulate MHC-I trafficking. J. Vi-rol.73:1964–1973.
43. McMichael, A. J., P. Parham, N. Rust, and F. Brodsky.1980. A monoclonal
antibody that recognizes an antigenic determinant shared by HLA A2 and B17. Hum. Immunol.1:121–129.
44. Miller, M. D., M. T. Warmerdam, S. S. Ferrell, R. Benitez, and W. C. Greene.1997. Intravirion generation of the C-terminal core domain of HIV-1 Nef by the HIV-1 protease is insufficient to enhance viral infectivity. Virology234:215–225.
45. Miller, M. D., M. T. Warmerdam, K. A. Page, M. B. Feinberg, and W. C. Greene.1995. Expression of the human immunodeficiency virus type 1 (HIV-1) nef gene during HIV-1 production increases progeny particle in-fectivity independently of gp160 or viral entry. J. Virol.69:579–584. 46. Nguyen, D. H., and J. E. Hildreth.2000. Evidence for budding of human
immunodeficiency virus type 1 selectively from glycolipid-enriched mem-brane lipid rafts. J. Virol.74:3264–3272.
47. Pandori, M. W., N. J. Fitch, H. M. Craig, D. D. Richman, C. A. Spina, and J. C. Guatelli.1996. Producer-cell modification of human immunodeficiency virus type 1: Nef is a virion protein. J. Virol.70:4283–4290.
48. Peng, B., and M. Robert-Guroff.2001. Deletion of N-terminal myristoylation site of HIV Nef abolishes both MHC-1 and CD4 down-regulation. Immunol. Lett.78:195–200.
49. Piguet, V., Y. L. Chen, A. Mangasarian, M. Foti, J. L. Carpentier, and D. Trono.1998. Mechanism of Nef-induced CD4 endocytosis: Nef connects CD4 with the mu chain of adaptor complexes. EMBO J.17:2472–2481. 50. Piguet, V., F. Gu, M. Foti, N. Demaurex, J. Gruenberg, J. L. Carpentier, and
D. Trono.1999. Nef-induced CD4 degradation: a diacidic-based motif in Nef functions as a lysosomal targeting signal through the binding of beta-COP in endosomes. Cell97:63–73.
51. Piguet, V., O. Schwartz, S. Le Gall, and D. Trono.1999. The downregulation of CD4 and MHC-I by primate lentiviruses: a paradigm for the modulation of cell surface receptors. Immunol. Rev.168:51–63.
52. Piguet, V., L. Wan, C. Borel, A. Mangasarian, N. Demaurex, G. Thomas, and D. Trono.2000. HIV-1 Nef protein binds to the cellular protein PACS-1 to downregulate class I major histocompatibility complexes. Nat. Cell Biol.
2:163–167.
53. Preusser, A., L. Briese, A. S. Baur, and D. Willbold.2001. Direct in vitro binding of full-length human immunodeficiency virus type 1 Nef protein to CD4 cytoplasmic domain. J. Virol.75:3960–3964.
54. Riggs, N. L., H. M. Craig, M. W. Pandori, and J. C. Guatelli.1999. The dileucine-based sorting motif in HIV-1 Nef is not required for down-regu-lation of class I MHC. Virology258:203–207.
55. Rodgers, W., and J. K. Rose.1996. Exclusion of CD45 inhibits activity of p56lck associated with glycolipid-enriched membrane domains. J. Cell Biol.
135:1515–1523.
56. Ross, T. M., A. E. Oran, and B. R. Cullen.1999. Inhibition of HIV-1 progeny virion release by cell-surface CD4 is relieved by expression of the viral Nef protein. Curr. Biol.9:613–621.
57. Schwartz, O., V. Marechal, O. Danos, and J. M. Heard.1995. Human immunodeficiency virus type 1 Nef increases the efficiency of reverse tran-scription in the infected cell. J. Virol.69:4053–4059.
58. Schwartz, O., V. Marechal, S. Le Gall, F. Lemonnier, and J. M. Heard.1996. Endocytosis of major histocompatibility complex class I molecules is induced by the HIV-1 Nef protein. Nat. Med.2:338–342.
59. Simons, K., and E. Ikonen.1997. Functional rafts in cell membranes. Nature
387:569–572.
60. Smith, A. J., M. I. Cho, M. L. Hammarskjold, and D. Rekosh.1990. Human immunodeficiency virus type 1 Pr55gagand Pr160gag-polexpressed from a
simian virus 40 late replacement vector are efficiently processed and assem-bled into viruslike particles. J. Virol.64:2743–2750.
61. Spearman, P., R. Horton, L. Ratner, and I. Kuli-Zade.1997. Membrane binding of human immunodeficiency virus type 1 matrix protein in vivo supports a conformational myristyl switch mechanism. J. Virol.71:6582– 6592.
62. Srinivasakumar, N., N. Chazal, C. Helga-Maria, S. Prasad, M. L. Ham-marskjold, and D. Rekosh.1997. The effect of viral regulatory protein ex-pression on gene delivery by human immunodeficiency virus type 1 vectors produced in stable packaging cell lines. J. Virol.71:5841–5848.
63. Swann, S. A., M. Williams, C. M. Story, K. R. Bobbitt, R. Fleis, and K. L. Collins.2001. HIV-1 Nef blocks transport of MHC class I molecules to the cell surface via a PI 3-kinase-dependent pathway. Virology282:267–277. 64. Takata, M., H. Sabe, A. Hata, T. Inazu, Y. Homma, T. Nukada, H.
Yama-mura, and T. Kurosaki.1994. Tyrosine kinases Lyn and Syk regulate B cell receptor-coupled Ca2⫹mobilization through distinct pathways. EMBO J.
13:1341–1349.
65. van’t Hof, W., and M. D. Resh.2000. Targeting proteins to plasma mem-brane and memmem-brane microdomains by N-terminal myristoylation and pal-mitoylation. Methods Enzymol.327:317–330.
66. Walk, S. F., M. Alexander, B. Maier, M. L. Hammarskjold, D. M. Rekosh, and K. S. Ravichandran.2001. Design and use of an inducibly activated human immunodeficiency virus type 1 Nef to study immune modulation. J. Virol.75:834–843.
67. Wang, J. K., E. Kiyokawa, E. Verdin, and D. Trono.2000. The Nef protein of HIV-1 associates with rafts and primes T cells for activation. Proc. Natl. Acad. Sci. USA97:394–399.
on November 8, 2019 by guest
http://jvi.asm.org/
68. Welker, R., M. Harris, B. Cardel, and H. G. Krausslich.1998. Virion incor-poration of human immunodeficiency virus type 1 Nef is mediated by a bipartite membrane-targeting signal: analysis of its role in enhancement of viral infectivity. J. Virol.72:8833–8840.
69. Welker, R., H. Kottler, H. R. Kalbitzer, and H. G. Krausslich.1996. Human immunodeficiency virus type 1 Nef protein is incorporated into virus parti-cles and specifically cleaved by the viral proteinase. Virology219:228–236. 70. White, S., K. Miller, C. Hopkins, and I. S. Trowbridge.1992. Monoclonal
antibodies against defined epitopes of the human transferrin receptor cyto-plasmic tail. Biochim. Biophys. Acta1136:28–34.
71. Willey, R. L., J. S. Bonifacino, B. J. Potts, M. A. Martin, and R. D. Klausner.
1988. Biosynthesis, cleavage, and degradation of the human immunodefi-ciency virus 1 envelope glycoprotein gp160. Proc. Natl. Acad. Sci. USA
85:9580–9584.
72. Williams, M., J. F. Roeth, M. R. Kasper, R. I. Fleis, C. G. Przybycin, and
K. L. Collins.2002. Direct binding of human immunodeficiency virus type 1 Nef to the major histocompatibility complex class I (MHC-I) cytoplasmic tail disrupts MHC-I trafficking. J. Virol.76:12173–12184.
73. Xavier, R., T. Brennan, Q. Li, C. McCormack, and B. Seed.1998. Membrane compartmentation is required for efficient T-cell activation. Immunity8:723– 732.
74. Yu, G., and R. L. Felsted.1992. Effect of myristoylation on p27 nef subcel-lular distribution and suppression of HIV-long terminal repeat transcription. Virology187:46–55.
75. Zhang, W., R. P. Trible, and L. E. Samelson.1998. LAT palmitoylation: its essential role in membrane microdomain targeting and tyrosine phosphory-lation during T-cell activation. Immunity9:239–246.
76. Zheng, Y. H., A. Plemenitas, T. Linnemann, O. T. Fackler, and B. M. Peterlin.2001. Nef increases infectivity of HIV via lipid rafts. Curr. Biol.
11:875–879.