0022-538X/93/063481-08$02.00/0
Copyright © 1993, American
Society
forMicrobiologyIdentification and Characterization
of
lef-1,
a
Baculovirus
Gene
Involved in Late and Very Late Gene Expression
A. LORENA PASSARELLI'ANDLOIS K.
MILLER'12*
Department of Genetics' andDepartmentofEntomology,2 The University of Georgia,
Athens,
Georgia
30602Received30 October 1992/Accepted5March 1993
An Autographa californica nuclear polyhedrosis virus (AcMNPV) gene required in transient expression
assays for late andverylate viral geneexpressionwasidentified, sequenced, and transcriptionally mapped.
This gene, designated late expression factor 1 (lef-1), was located between 7.4 and 8.7 map units of the
AcMNPVphysicalmap.Itwasidentified by cotransfecting Spodopterafrugiperda cultured cells withacollection ofoverlappingcloned DNA fragmentscovering theentire AcMNPVgenomeandareportergenecontrolledby
anearly, late,orverylate AcMNPVpromoter.Omissionof the DNAfragmentcontaininglef-lcurtailedmost
late andverylategeneexpressionbutnotearlygeneexpression.lef-1 wasfoundtobeanearlygenetranscribed
as a 1.8-kb RNA in the presence of the protein synthesis inhibitor cycloheximide. The C terminus of the
predicted polypeptide product,LEF-1, contained asequence motifcharacteristicof nucleoside
triphosphate-binding sites.
Upon infectionofpermissive culturedcells with the bac-ulovirusAutographa californica nuclear polyhedrosis virus (AcMNPV), three major classes of viral transcripts are detected: early, late, andverylate (forarecentreview, see reference23). Earlygenes aretranscribedbyan a-amanitin-sensitive RNA polymerase (8, 14), apparently host RNA polymerase II (13), andtranslated toprovidefactors
neces-saryfor DNAreplicationandtransactivation of lategenes.
Basal transcription of early genes does not require active
viralproteinsynthesis (6,13, 28, 34), althoughtheproductof the ie-1 gene isrequiredforexpressionofsomeearlygenes intransient expressionassays(9, 10).Following theonsetof
DNAreplicationat approximately6 hpostinfection (p.i.),a
drasticchangeingeneexpression is observed. Lateandvery
late gene transcription involving an ax-amanitin-resistant
RNApolymerase activity (8, 14)commences,whereaslevels
ofhosttranscripts arereduced (18). The latetranscriptional phase provides structural proteins for nucleocapsid assem-bly and budded virus production. Thevery late
transcrip-tionalphaseisdelayeduntilapproximately18to24 hp.i. (33)
and provides gene products involved in occlusion suchas polyhedrin.
We are interested inidentifying andcharacterizing bacu-lovirus genes involved in the transitions betweenearly and late and between late andvery late viralgene expression. Thus,wedevelopedatransientexpressionassaytoidentify
viralgenesinvolved inexpressionfrom thelatecapsid (vp39)
and the very late polyhedrin (polh) promoters (25). This assay has already been successfully applied in assigning a
role to the 630-nucleotide (nt) open reading frame (ORF) (26),renamedlef-2,in late andverylategeneexpression (25)
and inidentifyingadditional roles fortwobaculovirus
regu-latory proteins,IE-1 and IE-N(3, 9-11, 25).Inthisstudy,we
have used the transientexpressionassayto identifyanovel
earlygene, lef-1, involved in expressionofa reporter gene driven by a late or very late baculovirus promoter. The
predictedLEF-1polypeptidecontains266amino acids with
* Correspondingauthor.
an estimated molecular mass of 31 kDa and a C-terminal nucleoside triphosphate-binding motif.
MATERIALS ANDMETHODS
Virus and cells. The Spodoptera frugiperda (fall
army-worm) IPLB-SF-21 (SF-21) cell line (35)wasgrownat27°C
in TC-100 medium (GIBCO/BRL, Gaithersburg, Md.)
sup-plemented with 10% fetal bovine serumand0.26% tryptose brothasdescribedpreviously(23). Wild-type AcMNPVL-1 (16), usedto obtainviral DNA fortransfections,was
prop-agated in SF-21 cells or Trichoplusia ni (cabbage looper)
larvae infected peros (23).
Recombinant plasmids. The threereporterplasmids used, phcwt, pCAPCAT, and pETCAThr5, have been previously
described(20,25, 33). Briefly,thereporterplasmidscontain the AcMNPV promoter from the polyhedrin gene (for phcwt), the major capsid protein gene (forpCAPCAT), or
the proliferating cell nuclear antigen homologous gene, etl (for pETCAThr5), controlling the reporter gene encoding
chloramphenicol acetyltransferase (CAT). The enhancer-likeelementorhomologous region (hr5)ispresentupstream ofeachpromotersequence.
Subclones ofpBamC, a pBR322-based (2) plasmid con-taining the AcMNPV BamHI Cfragment (4.8 to 11.1 map units [m.u.]),were cloned into pBluescript (Stratagene, La
Jolla, Calif.). pBCPsB contains the PstI-BamHI fragment (7.35 to 11.1 m.u.), pBCBPscontains the BamHI-PstI
frag-ment (4.8 to 7.35 m.u.), pBCPsE contains the PstI-EcoRI
fragment (7.35 to8.65m.u.), pBCPsScontains the PstI-SalI
fragment (7.35 to 8.3 m.u.), and pBCSEcontains the
SalI-EcoRI fragment (8.3 to 8.65 m.u.). pBCNE contains a
NruI-EcoRIfragment (7.5 to8.65m.u.) comprisingthelef-1 ORFandtwosmall antisenseORFs. pBCHE, containingthe
HpaI-EcoRIfragment (7.9to8.65m.u.),wasconstructedto
synthesize a lef-1-specific probe used in Northern (RNA)
blot analysis.
DNA cotransfections and transient expression assays. Ap-proximately 2 x 106 SF-21 cells per 60-mm-diameter dish
were used per cotransfection by the calcium phosphate coprecipitation method (23). Each cotransfection mixture
3481
on November 9, 2019 by guest
http://jvi.asm.org/
fresh complete TC-100 medium was added to each mono-layer, and the cells were incubated at 27°C for 24 h for
cotransfections containing pETCAThr5 or48 h for cotrans-fections containing pCAPCATor phcwtbefore harvesting.
CATreactions were performed as previously described (7, 27), using 1/10 of theharvested cell lysate perreaction.
Nucleotide sequencing. pBluescript plasmids containing
theregionbetweenPstI(7.35 m.u.) andSall (8.3 m.u.)were
used togenerate nested setsofdeletions, usingexonuclease IIIand mungbeannuclease asdescribedby Stratagene (La Jolla, Calif.)based ontheoriginalmethodby Henikoff(12).
Selected deletion clones, providing sequence information
from bothDNAstrands across the entireregion, wereused as double-stranded DNA templates and sequenced by the dideoxyribonucleotide chaintermination method (30), using
T7 and T3 primers purchased from Stratagene. Since the
sequence from the Sall (8.3 m.u.) site to the EcoRI (8.65 m.u.) site had been previously determined (22), only one
DNA strand of thisregionwas sequenced, and thejunction
at theSalIsite (8.3 m.u.) wasverified.
RNAisolation. For RNA studies, SF-21 monolayers (5 x 106 cells per 100-mm-diameter dish) were infected with
AcMNPVatamultiplicityof20 PFUpercell. The viruswas
allowedtoadsorb for1 hatroomtemperature,and timezero was defined as the time when the virus was removed and
incubation at 27°C started. The protein synthesis inhibitor cycloheximidewasaddedat100p,g/mltothecells for 30min
before infection and maintained thereafter. The DNA
syn-thesis inhibitor aphidicolin wasadded at5 ,ug/ml following adsorption.TotalRNAwasisolatedatselectedhoursp.i. by
theguanidinium isothiocyanate method(4).
Northern blotanalysis. Samples oftotal RNAl,g(15 per
lane) were denatured by glyoxalation (17), separated by agarose gel electrophoresis, transferred to a nylon mem-brane, and hybridized to a strand-specific DNA probe at
42°C. The probe, which was complementary to the lef-1 transcript, was made from single-stranded pBCHE DNA. The T7 oligonucleotide primer was annealed to pBCHE single-stranded DNA to synthesize a radiolabeled DNA
strandcomplementarytothelef-1 transcript, usingthelarge fragment ofDNApolymeraseI to extend the primer in the
presenceof[Ot-32P]dCTP (29).
Si nuclease protection oflef-l transcripts. Total RNA(30 ,ugperlane)wasusedforS1nucleaseprotectionassays(36).
To construct the plasmidused as aprobe for mapping the
3' end(s) of the lef-1 transcript(s), pBamC was digested
with NdeI, radiolabeled exclusively at one end with T4
DNA polymerase and [a-32P]dCTP, and redigested with
MscI togenerateafragment3.9kbpinlength with onlyone
radiolabeled end. The labeled fragment of interest was
purified by agarose gel electrophoresis and annealed to
total RNA in
formamide-piperazine-N,N'-bis(2-ethanesulfon-ic acid) (PIPES) buffer (80% deionized formamide, 40 mM PIPES [pH 6.4], 0.4 M EDTA) at 52°C. After S1 nuclease
digestion, theprotected fragmentsweredenatured, separated on a polyacrylamide-urea gel, and visualized by autoradi-ography.
Primer extension analysis. The oligodeoxyribonucleotide primer 5'-TCGACGCGCTTCTGCG-3', complementary to sequencesstartingattheSall site (8.3 m.u.) downstreamof thefirstmethioninecodon ofthelef-1 ORF,waslabeledwith [_y-32P]ATP (New England Nuclear, Boston, Mass.) andT7
BRL). A sequencing ladder of this regionwas generated by using the labeled primer in dideoxyribonucleotide chain termination DNA sequencing reactions.The extended prod-ucts were denatured, separated on a polyacrylamide-urea gel, and visualized by autoradiography.
Nucleotide sequence accession number. The GenBank
ac-cession number for the AcMNPV lef-I gene sequence is
L09723.
RESULTS
Experimental
strategy.
To identify genesinAcMNPVthat regulate different temporal classes of promoters, a plasmid containing the CAT reporter gene controlled by an early,late, orvery late AcMNPV promoter was cotransfectedinto SF-21 cells with a series of cloned, overlapping fragments
comprising allof the AcMNPV DNA genome (Fig. 1D)(25)
toprovide all of theviral factors required for transcription.
Upon cotransfection of SF-21cells with the entirecollection
ofcloned viral DNAs and areporter plasmid, reporter gene
expression was expected from each temporal class of
pro-moter. However, omission of any one of the AcAMNPV
fragments which provides a function necessary for
expres-sion from the promoter tested should eliminate CAT gene
expression.
Identification of agenein the region between 4.9 and 14.7
m.u. required for vp39 andpolh promoter-mediated expres-sion.The reporterplasmids used, pETCAThr5, pCAPCAT, andphcwt, contain the CAT reporter gene fused to the early
etl(6), the late vp39 (32), and the very latepolh promoters, respectively. The etl promoter is dependent solely on the IE-1 gene product for expression in transient expression assays,but IE-1 is not sufficient for expressionfrom the vp39 or polh promoter (25).
Transfection of each reporter plasmid alone into SF-21
cells gave no CAT activity, but highlevels were observed
whentheplasmids werecotransfectedwith intactAcMNPV
DNA (Fig. 1A to C; compare lanes 1 and 2).Cotransfection
of each reporter plasmid with the overlapping library of
AcMNPV clones gave equal or slightly lower levels of
expression fromtheearly andlatepromoterstested (Fig. 1A
and B, lanes 3). CATexpressiondrivenby thepolh promoter
was much lower in cotransfections with the library than those with AcMNPV (Fig. 1C; compare lanes 2 and 3). Higher expression from thepolh promoter was observed if cells were harvested at 72 h postcotransfection, but the levels of expression with the library of clones were still lower than thatobserved with intact AcMNPV DNA under these assayconditions.
Removal of theIE15 clone, which containsie-1 from the AcMNPV clone library, abolished expression from all three temporal classes of promoter, as expected (Fig. 1, lane 16) (25). However, omission of several clones (i.e., BC5, PstH5, and HC10) from the library greatly diminished expression from the late and very late promoters but not the early promoter (Fig. 1, lanes 4, 11, and 13). Although omission of the IE15 clone gave no detectable levels of CAT activity, omission of BC5, PstH5, orHC10 gave very slight levels of CAT activity when autoradiograms were overexposed. Omission of several other library clones (i.e., PstHl,
pXmaB, and HK5) also diminished expression from the late andvery late promoters (Fig.1B and C, lanes 10, 14, and 15).
on November 9, 2019 by guest
http://jvi.asm.org/
Clones
Omitted
I
> _
Z- >1 I-~
z c to w I q)C_ I I CZE t t U U)0 -Jo Tcor tot 0 0
uc
LO2:
cm
I I ualf: CLaCL
CL I I a I LLA
pETCAThr5
AcCm
* * . * , 0 - c
99999999494 99.99
99-Cm
B*_
pCAPCAT
AcCm
*.
*..-*-
.
41.
@41
*410414*
9 99999999999999-* **
*Cm
C_
phcwt
Ac Cm
*~~..
*
*
99999999999999j
j
j
*
*
*-Cm
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
EcoRi
hrl hr2 hr3 hr4L hr4R hr5
I
|tRO A I J K T M hJ F VU C aW G D:>
as
L E HS
P BDJ * go1 A kJK MNIF1 VU C Gl DOL I
le,
B151 57 865 181 25400 32194 434 57 601 1321 [4] 97 8887 903 1000
00 69 194-199 301 348 428 527161533 698728 87916]883
BC5 PStH4 HC9
- I.--- I 4
IE15 HL8 PstHl HC10 1E15
...4 - -4 - - --4 4
-HL5 pBSP-PstH pXmaB
,,, .. 4 v
ETL7 PstH5 HK5
1--- - ...4
---FIG. 1. CAT expression from early (A), late (B), and very late (C)AcMNPVpromoters in thepresence or absence ofdifferent regions of the AcMNPV genome(D). SF-21 cells were transfected withpETCAThr5 (A), pCAPCAT (B), or phcwt (C), and lysates were collected after 24hforpETCAThr5or 48 h forpCAPCAT and phcwt. Transfections werecarriedout with the reporter plasmid only (lane 1), intact wild-type AcMNPV DNA(lane 2), the entire overlapping AcMNPV clone library (lane 3), or theAcMNPV library lacking the specified clones (lanes 4 to 16). The acetylated products (Ac Cm) and unacetylated substrate (Cm) of each enzymatic reactionwere separated by thin-layer chromatography and visualized byautoradiography. (D) Locations of the cloneswithrespect to the AcMNPV genome are schematically presented aslines below theEcoRIrestriction map of theAcMNPV genome as reported by O'Reilly et al. (23). The solid lines indicate the regionknown to be covered by the clone; the broken lines at the ends of the solid lines denote the ambiguity of the endpoints of each clone.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.105.515.97.628.2]Ps
Ps
r-
'~~~~~~~~~~~-Ps E
IP
Ps S
pBCSE
B
0
r-l
S E
0
1kb
AcCm
** -Cm
BC5 clone BC5 subclone
1 2 3 4 5 6 7 8
BamC BCPsB BCBPs BCPsE BCPsS BCSE
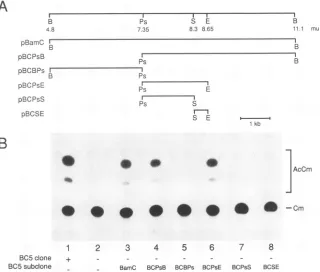
FIG. 2. CAT expression from thepCAPCATreporterplasmid, usingsubclones of the AcMNPVgenomein theregionfrom 4.8to11.1m.u.
toreplace the BC5 cloneof theoverlappingAcMNPVclonelibraryincotransfections.(A)The subclones used intransientexpressionassays.
The numbers indicateapproximatemapunits in theAcMNPVgenome;restrictionenzymesitesareabbreviatedasfollows: B, BamHI; Ps, PstI;S,Sall;E, EcoRI. Onlyoneof theSalland EcoRI sites inthisregionis shown.(B)AssayinwhichSF-21 cellswerecotransfected with
pCAPCATand theentire AcMNPVlibrary (lane 1)orcellsweretransfected withpCAPCATandthe AcMNPVlibraryexcept for theBC5
clone andaBC5 subclonewasaddedasindicated(lanes2to8).Theacetylated products (AcCm)andunacetylatedsubstrate(Cm)ofeach enzymaticreactionwereseparated by thin-layerchromatographyandvisualizedby autoradiography.
This study focused on determining the gene(s) within the
BC5 clonerequiredforexpressionfrom the late andverylate promoters(Fig. 1,lane4). TheBC5 clonemapsbetween 4.9 and 14.7m.u. ontheAcMNPVgenome.
A clonecontaining the BamHI Cfragment ofAcMNPV, pBamC,which extends from4.8to11.1m.u. onthegenome,
was foundto substitute forBC5 in the transient expression
assay (Fig. 2B, lane 3). pBamC was then subcloned into
smaller fragments, which were tested by similar transient
expression assays for the ability to restore CAT activity
from thevp39promoterinthe absence of BC5. The subclone
pBCPsE (7.35 to 8.65 m.u.) was the smallest subclone capable ofsubstituting for BC5 (Fig. 2). Subclones ofthis plasmidcut attheSalI site, pBCPsS (7.35 to 8.3 m.u.) and pBCSE (8.3to8.65 m.u.),wereunabletoreplace BC5 (Fig. 2B, lanes 7 and 8). This finding suggested that a gene
necessary forexpression from the vp39 promoter spanned
the SalI site at 8.3 m.u. The same results were obtained
when thevery latepromoter reporter plasmid, phcwt, was used in transient expression assays with these subclones (datanotshown).
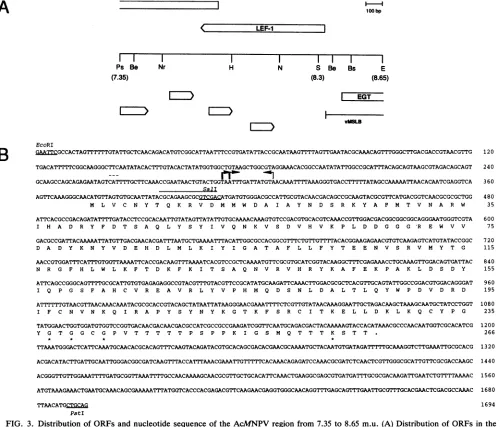
Nucleotide sequence of thelef-) gene. The nucleotide
se-quencecorresponding to theregion between 7.35 and 8.65
m.u. was determined in both directions. This region con-tainedseveral putative ORFs larger than 50 amino acids,as indicated in Fig. 3A. The largest ORF, designatedlef-1 for
late expression factor 1,was 266 codons and spanned the Sall site at 8.3 m.u.; since disruption of this site reduced
CATactivity (Fig. 2B,lane 7),thepolypeptideencodedby
this ORFwaslikelytoberequiredforvp39 expression. The three smallORFs contained withinpBCPsSwerenotable to reconstitute CAT activity in the absence of BC5 (Fig. 2B,
lane 7). To rule out the possibility that the partial ORF downstream oflef-1 wasnecessaryforexpressionfromthe vp39promoter, asubclone(NruIat7.5m.u.toEcoRIat8.65
m.u.)disruptingthispartial ORFwasusedto substitute for
BC5 in transient expression assays. This clone, pBCNE,
was able to complement CAT activity (data not shown), indicating that the lef-1 ORF and not the flanking and
overlappingORFswasinvolved invp39- and polh-mediated
expression.
The nucleotide sequence of lef-1 is shown in Fig. 3B.
Comparison ofthe predicted polypeptide sequenceoflef-1 with those in published data bases revealed no obvious homologyto anyother knownpolypeptide. Analysis of the
derived amino acid sequence of lef-I for known protein
sequence motifs revealed the presence of a nucleoside triphosphate-binding site similarto the ATP-binding site of
severalproteinkinases (15). Itconsists ofatriad of appro-priately spaced glycines followed by a lysine 21 residues
downstreamneartheC terminus(Fig. 3B, amino acids 237to
263).
Northernblotanalysisoflef-1 transcription.The temporal regulation oflef-I transcriptionwasexamined by Northern
blot analysis. SF-21 cells were infected with AcMNPV,
harvested at various times p.i., and lysed, and total RNA wasextracted. Aprotein synthesis inhibitor, cycloheximide,
anda DNAsynthesis inhibitor, aphidicolin, wereadded to pBamC rB
B pBCPsB
pBCBPs rF B pBCPsE
pBCPsS
B
B
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.146.466.76.348.2]A
Ps Be (7.35)
Nr H
l
100bp
< LEF-1
Bs
I
N S Be
(8.3)
E (8.65)
ILD
IED
i EGT
vMLB
EcoRI
TGACATTTTTCGGCAAGGGCTTCAATATACACTTTGTACACTATATGGTGGCTGTAAGCTGGCGTAGGAAACACGGCCAATATATTGGCCGCATTTACAGCAGTAAGCGTAGACAGCAGT
GCAAGCCAGCAGAGAATAGTCATTTTGCTTCAAACCGAATAACTGTACTGGTAATTTGATTATGTAACAAATTTTAAAGGGTGACCTTTTTATAGCCAAAAATTAACACAATCGAGGTCA
SallI
AGTTCAAAGGGCAACATGTTAGTGTGCAATTATACGCAGAAGCGCGTCGACATGATGTGGGACGCCATTGCGTACAACGACAGCCGCAAGTACGCGTTCATGACGGTCAACGCGCGCTGG
M L V C N Y T Q K R V D M M W D A I A Y N D S R K Y A F M T V N A R W
ATTCACGCCGACAGATATTTTGATACCTCCGCACAATTGTATAGTTATATTGTGCAAAACAAAGTGTCCGACGTGCACGTCAAACCGTTGGACGACGGCGGCGGCAGGGAATGGGTCGTA
I H A D R Y F D T S A Q L Y S Y I V Q N K V S D V H V K P L D D G G G R E W V V
GACGCCGATTACAAAAATTATGTTGACGAACACGATTTAATGCTGAAAATTTACATTGGCGCCACGGCGTTTCT'GTTGTTTTACACGGAAGAGAACGTGTCAAGAGTCATGTATACCGGC
D A D Y K N Y V D E H D L M L K I Y I G A T A F L L F Y T E E N V S R V M Y T G
AACCGTGGATTTCATTGTGGTTAAAATTCACCGACAAGTTTAAAATCACGTCCGCTCAAAATGTTCGCGTGCATCGGTACAAGGCTTTCGAGAAACCTGCAAAGTTGGACAGTGATTAC
N R G F H L W L K F T D K F K I T S A Q N V R V H R Y K A F E K P A K L D S D Y
ATTCAGCCGGGCAGTTTTGCGCATTGTGTGAGAGAGGCCGTACGTTTGTACGTTCCGCATATGCAAGATTCAAACTTGGACGCGCTCACGTTGCAGTATTGGCCGGACGTGGACAGGGAT
I Q P G s F A H C V R E A v R L Y V P H M Q D S N L D A L T L Q Y W P D V D R D
ATTTTTTGTAACGTTAACAAACAAATACGCGCACCGTACAGCTATAATTATAAGGGAACGAAATTTTCTCGTTGTATAACAAAGGAATTGCTAGACAAGCTAAAGCAATGCTATCCTGGT 10180
I F C N V N K Q I R A P Y S Y N Y K G T K F S R C I T K E L L D K L K Q C Y P G 235
TATGGAACTGGTGGATGTGGTCCGGTGACAACGACAACGACGCCATCGCCGCCGAAGATCGGTTCAATGCAGACGACTACAAAAAGTACCACATAAACGCCCAACAATGGTCGCACATCG
Y G T G G c G P v T T T T T P S P P K I G S M Q T T T K S T T .
TTAAATGGGACTCATTCAAATGCAACACGCACAGTTTCAAGTACAGATACGTGCACAGCGACACGAACGCAAAATGCTACAATGTGATAGATTTTGCAAAGGTCTTGAAATTGCGCACG
1200 266
1320
ACGACATACTTGATTGCAATTGGGACGGCGATCAAGTTTACCATTTAAACGAAATTGTTTTTCACAAACAGAGATCCAAACGCGATCTCAACTCGTTGGGCGCATTGTTCGCGACCAAGC 14140
ACGGGTTGTTGGAAATTTTGATGCGGTTAAATTTTGCCAACAAAAGCAACGCGTTGCTGCACATTCAAACTGAAGGCGAGCGTGATGATTTGCGCGACAAGATTGAATCTGTTTTAAAAC 11560
ATGTAAAGAAACTGAATGCAAACAGCGAAAAATTTATGGTCACCCACGAGACGTTCAAGAACGAGGTGGGCAACAGGTTTGAGCAGTTTGAATTGCGTTTGCACGAACTCGACGCCAAAC 168 0
TTAACATGCTGCA 1694
PstI
FIG. 3. Distributionof ORFsand nucleotide sequenceof theAcMNPV region from 7.35 to8.65m.u. (A) Distribution of ORFs in the
PstI-EcoRI (7.35to8.65 m.u.) subfragment of AcMNPV. EGT is theecdysteroid UDP-glucosyltransferasegene(21); the barlabeled vMSLB
indicatesone endpoint of the deletionpresentinthe mutantvirus vMSLB (24).Abbreviations for restriction enzyme sites:Ps, PstI; Be,
BstEII; Nr, NruI; H, HpaI; N, NarI;S, SalI; Bs, BstXI; E, EcoRI. (B) Nucleotidesequenceoflef-I from 8.65to7.35m.u. and the derived polypeptidesequence.The thickand thinarrowsindicate positions of the 5' ends mapped for thelef-I andegttranscripts,respectively. The
brokenoverline shows thepositionof thetranslationalstartcodon for EGT. Thenucleotides complementarytotheoligonucleotide used in primerextensionanalysisareoverlined. Theamino acidsconstituting the putative nucleosidetriphosphate-binding site haveanasterisk. Key
restriction endonuclease sitesareunderlined and indicated.
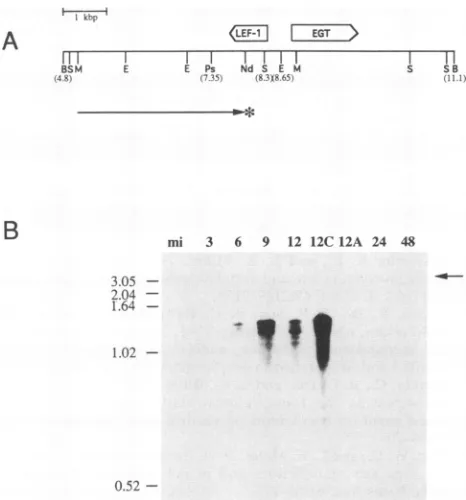
somecellstoassesstherequirementsforproteinand DNA
synthesis forlef-1 transcription. The strand-specific probe
used is represented byan arrow inFig. 4A. Atranscript of
approximately 1.8 kbwas observedat 3, 6, 9,and 12 h p.i. (Fig. 4B). Thetranscriptwasalso observed in thepresence ofcycloheximide. In thepresenceofaphidicolinat12hp.i.,
thelevels of thetranscriptwerelow. Thistranscriptwasnot observed at very late times (e.g., 24 and 48 h p.i.) or in mock-infected cells.Thus,lef-1 appearstobeanearlygene, and lef-1 transcription occurs in the absence of protein synthesisorDNAreplication. Upon longerexposuresof the
autoradiogram,aminortranscriptofapproximately2.8 kb in lengthwas observed at early times and at 12 h p.i. in the
presenceofcycloheximide;wehavenotfurtherexploredthe natureof this early transcript.
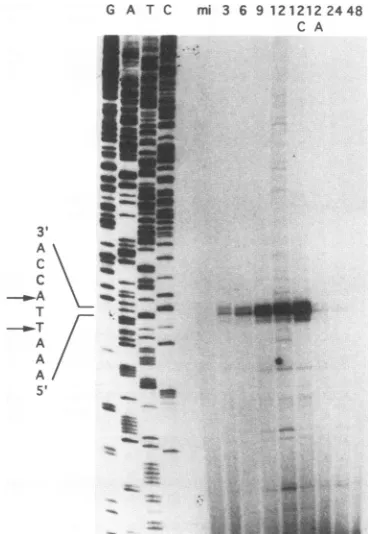
Mapping the 5' termini oflef-l transcripts. The 5' termini of the major lef-1 RNAwere mapped by primer extension analysis. A16-ntoligomer complementaryto thelef-1
tran-scriptandcoincidingwith theSallsiteat8.3m.u. (Fig. 3B) was5' end labeled and annealedtototal RNA isolated from uninfected SF-21 cells or cells infected with AcMNPV harvestedatvarious hoursp.i.Theannealedoligonucleotide was then extended with reverse transcriptase, and the
ex-tended products were analyzed and compared with a
se-quenceladder of thesameregioninadenaturing
polyacryl-amidegel.Twomajorextensionproductsof117 and 119 nt
B
120240 360
480 35 600 75 720 115 840 155 960 195
I I
I I
I I I I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.69.572.79.506.2]J
H N S Be
(8.3)
Bs E
(8.65)
mi 3 6 9 12 12C12A 24 48
11.20-
9.16-
7.13-
5.09-4.07-_
3.05-2.04'- it
t
1.640
1.0
FIG. 4. Northern blot analysis of lef-1 transcripts. (A) The
strand-specific DNAfragment used for Northern blot analysis is
illustrated byan arrowunder theopen arrowrepresentingthelef-l
ORF. Numbers indicateapproximate mapunits, and letters show
key restriction sites as designatedinFig. 3A. (B)Total RNAwas
extracted from SF-21 cells either mock infected(mi)orinfected with AcMNPVatthe timesindicated inhoursabove the lanes. RNA from
samples treated with cycloheximide(12C)oraphidicolin(12A)was
extractedat12hp.i. Molecular weightmarkersareindicatedtothe left inkilobasepairs.
inlength, correspondingto nt292and294ofFig. 3B,were
observed (Fig. 5). These virus-specific extension products wereobserved forRNA isolatedat3, 6, 9,and 12 hp.i.inthe presenceor absenceofcycloheximide. The sameextension
products werealso observed at 12hp.i. in thepresence of
aphidicolin, albeit atmuch lowerlevels. Atvery latetimes (24 and 48 h p.i.), the extension products of these RNAs
weregreatly reduced.Usinganoligonucleotide
complemen-tary to sequences downstream of the fourth methionine of thelef-I ORF (i.e.,aprimerspanningnt473to488;Fig. 3B),
the only extension products observed also mapped to the samesitesasdescribed above,thusrulingoutthepossibility
that thelef-1 transcript(s) initiated within sequences corre-sponding to the predicted LEF-1 N terminus (data not
shown).
The 5' ends of the transcripts were also mapped by S1 nuclease protection analysis using a probe uniquely end labeledatthe 5' endofthe Sall site shown in Fig.3(datanot
shown). Twofragments 117 and 119 ntlong, corresponding againto nt292 and294 in Fig.3B,wereprotected byRNA
frominfected butnotfrom mock-infected cells.
Mappingthe 3' termini oflef-I transcripts. The 3' end of
themajorlef-1 transcriptwaslocalized by using afragment
3'end labeledattheNdeI siteandextendingtosequencesin
theplasmidvector(Fig.6A). Fragments 1.4to1.25kbp long wereprotected at6through 12 h p.i. andat 12 hp.i. in the presence of cycloheximide (Fig. 6B). The amount of
pro-3'
A
am
_A
Ut___T --__a
_Tm _-
-A / :Ct
-A M
A /"a
5' _
JL..
.._
3=__'..
FIG. 5. Primer extension analysis of lef-1 transcripts.An oligo-nucleotideprimer (overlined in Fig. 3B) coinciding with theSallsite at8.3m.u. was5' endlabeled, annealedtototal RNAisolatedfrom mock-orAcMNPV-infected SF-21cells, andextended withreverse
transcriptase. The same primerwas used to derive a sequencing ladder(G, A,T, C)fromaplasmidcontainingtheprimer-extended region. Lanedesignationsarethesameasdescribed forFig.4B.The
sequencespanningthetranscriptionalstartsite isrepresentedtothe leftof the sequencing ladder, and the mapped 5' ends of the RNAs
areindicatedbyarrows.
tectedfragmentwasreduced at 12 hp.i. in thepresenceor absence ofaphidicolin.Similarexperimentsusingafragment 3' end labeledattheSalI site (8.3 m.u.)andextendingtothe
PstI site(7.35 m.u.)and then intoplasmidvectorsequences confirmed thatlef-1 transcriptsextendbeyond the PstI site at 7.35 m.u. (data not shown). Extension beyond thePstI site is also consistent with the lack of a polyadenylation
signalinthesequenced regiondownstream of thelef-1ORF. The 5' and 3' mapping dataare consistent with the 1.8-kb lef-1 RNAobservedin Northernblots.
DISCUSSION
Werecently developedanassaydesignedtoidentifygenes
necessaryfor late andverylateAcAMNPVgene expression (25). Ourassay uses alibrary ofclonescovering the entire
AcMNPV genome that can activate expression from a
re-porterCATgeneunderthe controlof thelatevp39promoter
or the very late polh promoter. The assay is based on deletinganindividualclone from thelibrary anddetermining
the smallest region(s) which can functionally substitute to activatereportergeneexpression. Thus,theassaymarkedly differs fromaprevious approachwhichusedaCATreporter gene under the control of a putative late promoter of a relativelyuncharacterizedgeneandmonitoredCAT expres-sion after adding only limited regions of the AcAMNPV
genome (11). One of the two ORFs mapped by this latter
Ps Be
(735)
Nr
B
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.60.295.69.343.2] [image:6.612.339.523.78.345.2]khp
A
11I
BSM ,4-8
B
@LEF Zii1EGT >
l
I I
E E Ps Nd S E M S
f7e.351 8.311.651
mi 3 6 9 12 12C 12A 24 48
3.05 2.04
-I.64
I.02
-0.52
-FIG. 6. Mapping the 3' end of the lef-1 transcript. (A) The probe
usedtomapthe3' endof thelef-I transcript. Thearrowindicates
the 5'-to-3'directionoftheprobe, and the asteriskrepresentsthe position of the label. Key restriction sitesareabbreviatedasin Fig. 3A.Additional sites: M, MscI; Nd, NdeI. Keymapunitsareshown
inparentheses belowtherestrictionsite.The size anddirection of
thelef-1 andegtORFsareindicated above therestrictionmap. (B) S1 protection analysis ofthe3'end of thelef-l RNA. Total RNA
was isolated from uninfected and AcMNPV-infected SF-21cells,
annealedtotheend-labeledprobe,and treated withS1 nuclease,and
protected fragmentswereseparated in a polyacrylamide-urea gel.
Lanedesignationsarethesame asdescribed for Fig. 4B. Thearrow
totheright indicatesthemigration of the untreated labeled probe.
Molecularweightmarkersareindicatedtothe leftinkilobasepairs.
technique was subsequently found to be nonessential for
virus replication in vivo and in vitro (24). The use of well-characterized promotersofgenes ofknown functional importance provides assurancethatwe are locating
neces-sarylate expressionfactors.
We have found that expression from the vp39 andpolh
promoters inourassayrequiresanumber of AcMNPVgenes distributedthroughoutthegenome. Inaprevious study, we defined threegenesinthe 83.7-to7.5-m.u.regioninvolved in lategene expression (25), and in thisstudy, we extendour analysistoa clone from the library thatflanksthis region.
One gene, which we have named lef-1, was found to be
required for substantial levels ofexpression from the late
vp39and theverylatepolhpromoters.
The derived polypeptide sequence of lef-1 contains 266 amino acids andpredictsa31-kDapolypeptide. TheLEF-1 sequence has no apparent homology to other published
sequences. The C terminus contains a nucleoside triphos-phate-bindingsite motif similartothetype found inprotein
kinases (G-X-G-X2-G-X15_20-K). The regioncontaining this sequence motif is rich in serine, threonine, andproline.
lef-1istranscribedas anearly1.8-kbRNA whichappears
to be a bicistronic mRNA, with a second ORF located
downstream andoverlapping thelef-1 ORF. Levels of the 1.8-kb transcript begin to decline during the late phase of
infection (e.g., 12 h p.i.) and are very low by 24 h p.i. This
declinemaydepend on early protein synthesis; at 12 h p.i.,
lef-l RNA is more abundant in the presence than in the absence of cycloheximide. The very low abundance of lef-1 RNAin the presence of aphidicolin at 12 h p.i. may be due to amorerapid decline oflef-IRNAin the absence of viral DNA replication and late gene products or the failure to expresslef-1 efficiently in the absence of DNA replication.
Using primer extension andSi nuclease protection anal-yses, we mapped the putative 5' ends of the lef-1 transcript
to sites 82 and 84 nt upstream of the translational start codon. The adjacent egt gene is transcribed in the opposite
direction from lef-1 (22), and the 5' ends of the egt RNA overlap with the 5' end(s) of the lef-1 RNA (Fig. 3B).
Overlappingantisense RNAs may result in inaccurate
map-ping of 5' termini by S1 nuclease protection and primer extension analyses (19). If this is also the case here, then the
lef-1 5' end(s) may extend further upstream than mapped here. Closer examination of the region within the egt ORF reveals the presence of two CAGT sequences (nt 221 and 237;Fig. 3B) with a TATA box at nt 201. A number of other earlybaculovirus genes have been reported to initiate at a
CAGT sequence usually located approximately 20 to 30nt
downstream ofaTATAbox(1, 5, 9, 10, 31). There areno
ATGcodons between the two CAGT sequences and the lef-I 5' endsthatwehave mapped. However, thereisanATG in the RNA leader atnt302 which is immediately followed by
aterminationcodon.
Atthe presenttime, we assume thatlef-1is essential to the virusin vivo. However, anAcMNPVmutant isolatedafter
serial passage through Manduca sexta larvae, vMSLB,
containsadeletion thatstarts at nt360 upstream of thelef-1
ORF(Fig. 3) and extendsthroughegt (24). Thus, thelef-1
ORF remains intact in vMSLB, but the promoter region including the putative transcriptional start sites has been
deleted;itispossiblethatlef-1is stilladequatelytranscribed invMSLB. Theprecise role of lef-Iin late geneexpression
remainstobe determined; genesidentifiedinthisassay may
include genesrequired forexpressionfromearly promoters other than the etl promoter, genesinvolvedin DNA replica-tion, and genesdirectlyinvolved in late gene expression.
This transient expression assay is clearly apowerful tool fordefiningtheAcMNPVgenesinvolved inexpressionfrom thevp39andpolh promotersand may be extendedtostudy
other viral gene promoters as well. We are presently at-tempting to
identify
all ofthe AcMNPV genesrequired forexpressionfrom thevp39 andpolh promoters.
ACKNOWLEDGMENTS
We thank D. R. O'Reilly for providing plasmids pBCPsE and pBCPsB, andwethankS.Kumar forpBamC.
This workwassupportedin partbyPublicHealth Servicegrant
Al 23719 from the National Institute of Allergy and Infectious Diseases.
REFERENCES
1. Blissard, G. W., and G. F. Rohrmann. 1989. Location,
se-quence, transcriptional mapping, and temporal expression of thegp64envelopeglycoproteingeneof theOrgyia
pseudotsug-atamulticapsid nuclearpolyhedrosis virus. Virology 170:537-555.
2. Bolivar,F.,R. L.Rodriguez,P.J.Greene,M.C.Betlach,H.L.
Heyneker, and H. W.Boyer.1977. Constructionand character-ization of new cloning vehicles. II. A multipurpose cloning
system.Gene2:95-113.
3. Carson, D. D., L. A. Guarino, and M. D. Summers. 1988. FunctionalmappingofanAcNPVimmediatelyearlygenewhich
f
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.67.300.78.328.2]5299.
5. Chisholm, G. E., and D. J. Henner. 1988. Multiple early tran-scripts and splicing of the Autographa califomica nuclear polyhedrosis virusIE-1 gene. J.Virol. 62:3193-3200.
6. Crawford,A.M., and L. K. Miller. 1988.Characterization ofan earlygeneaccelerating expression of lategenesofthe baculo-virus Autographa californica nuclear polyhedrosis virus. J. Virol. 62:2773-2781.
7. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinantgenomeswhich expresschloramphenicol acetyl-transferase in mammalian cells. Mol.Cell. Biol. 2:1044-1051. 8. Grula, M. A., P. L. Buller, and R. F. Weaver. 1981.
Alpha-amanitin-resistant viral RNAsynthesisin nuclei isolated from nuclear polyhedrosis virus-infected Heliothis zea larvae and Spodopterafrugiperda cells. J.Virol.38:916-921.
9. Guarino,L.A.,and M.D.Summers. 1986.Functionalmapping ofatrans-activating generequired forexpression ofa baculo-virusdelayed-earlygene.J. Virol. 57:563-571.
10. Guarino,L.A., and M. D.Summers. 1987. Nucleotidesequence and temporal expression of abaculovirus regulatorygene. J. Virol.61:2091-2099.
11. Guarino, L. A., and M. D. Summers.1988. Functional mapping ofAutographa califomica nuclear polyhedrosis virus genes requiredforlategeneexpression. J.Virol.62:463-471. 12. Henikoff, S. 1984.Unidirectional digestion with exonuclease III
createstargetedbreakpoints forDNAsequencing. Gene 28:351-359.
13. Hoopes, R. R., and G. F. Rohrmann. 1991.In vitro transcription ofbaculovirus immediateearlygenes: accuratemessengerRNA initiationby nuclearextractsfrom bothinsect and human cells. Proc. Natl.Acad. Sci. USA 88:4513-4517.
14. Huh, N. E., and R. F. Weaver. 1990. Identifying the RNA polymerases that synthesize specific transcripts of the Au-tographacalifornica nuclear polyhedrosis virus. J. Gen. Virol. 71:195-202.
15. Hunter, T. 1987. A thousand and one protein kinases. Cell 50:823-829.
16. Lee, H. H., and L. K. Miller. 1978. Isolation of genotypic variantsofAutographacalifornica nuclear polyhedrosisvirus. J. Virol.27:754-767.
17. McMaster, G. K., and G. G. Carmichael. 1977. Analysis of
single- and double-stranded nucleic acids on polyacrylamide and agarose gels by using glyoxal and acridineorange. Proc. Natl.Acad. Sci. USA 74:4835-4838.
18. Ooi, B. G., and L. K. Miller. 1988. Regulation ofhost RNA levels during baculovirus infection. Virology166:515-523. 19. Ooi, B. G., and L. K. Miller. 1991.The influenceof antisense
RNA ontranscriptionalmapping ofthe5' terminus of a bacu-lovirusRNA.J.Gen. Virol. 72:527-534.
20. Ooi, B. G., C. Rankin, and L. K. Miller. 1989. Downstream sequences augment transcription from the essential initiation
ofabaculovirusecdysteroid UDP-glucosyltransferasegene.J. Virol. 64:1321-1328.
23. O'Reilly,D.R.,L.K.Miller,and V. A.Luckow. 1992. Baculo-virusexpressionvectors: alaboratorymanual. W. H. Freeman &Co.,New York.
24. O'Reilly, D. R., A. L. Passarelli, I. F. Goldman, and L. K. Miller. 1990.Characterization of the DA26genein a hypervari-ableregionof theAutographa californicanuclearpolyhedrosis virusgenome.J. Gen. Virol. 71:1029-1037.
25. Passarelli, A. L., and L. K. Miller. 1993. Three baculovirus genes involved in late and very late geneexpression:ie-1, ie-n, andlef-2. J. Virol. 67:2149-2158.
26. Possee, R. D., T. P. Sun, S. C. Howard, M. D. Ayres, M. Hill-Perkins, and K. L. Gearing. 1991. Nucleotidesequenceof the Autographa californica nuclear polyhedrosis 9.4 kbp EcoRI-I and -R(polyhedrin gene) region. Virology185:229-241. 27. Rankin,C.,B. G.Ooi,and L. K.Miller.1988. Eightbasepairs encompassing the transcriptional start point are the major determinant forbaculoviruspolyhedrin gene expression.Gene 70:39-50.
28. Rice,W.C.,and L.K.Miller.1986. Baculovirustranscriptionin the presence of inhibitors and in nonpermissive Drosophila cells.Virus Res. 6:155-172.
29. Sambrook,J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratorymanual. ColdSpringHarborLaboratory Press,ColdSpring Harbor,N.Y.
30. Sanger, F.,S.Nicklen,and A.Coulson. 1977. DNAsequencing withchain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
31. Theilmann, D. A., and S. Stewart. 1991. Identification and characterization of the IE-1 gene of Orgyia pseudotsugata multicapsid nuclear polyhedrosis virus.Virology180:492-508. 32. Thiem, S.M.,and L.K.Miller. 1989.Identification,sequence,
andtranscriptionalmapping of the major capsid proteingeneof the baculovirusAutographa califomica nuclear polyhedrosis virus. J.Virol. 63:2008-2018.
33. Thiem, S. M., and L. K. Miller.1990. Differentialgene expres-sion mediated by late, very late, and hybrid baculovirus pro-moters. Gene 91:87-94.
34. Tomalski, M. D., J. Wu, and L. K. Miller. 1988.The location, sequence, transcription and regulation ofa baculovirus DNA polymerasegene.Virology 167:591-600.
35. Vaughn,J.L., R.H.Goodwin,G. J.Tompkdns,and P. McCaw-ley. 1977. The establishment oftwocell lines from the insect Spodoptera frugiperda (Lepidoptera: Noctuidae). In Vitro 13: 213-217.
36. Weaver, R. F., and C. Weissmman. 1979.MappingofRNA bya modification of theBerk-Sharp procedure: the 5' termini of 15S beta-globin mRNA precursor and mature 10 S beta-globin mRNA have identical map coordinates. Nucleic Acids Res. 7:1175-1193.