0022-538X/86/080479-07$02.00/0
Copyright © 1986, American Societyfor Microbiology
Relationship between
Poliovirus Neutralization and Aggregation
ADRI A. M. THOMAS, RAF VRIJSEN, ANDALBERT BOEYE*
Dep(aiittUient o
Miclrobiology
antid Hygiene, Vr-ije Universiitit Brulssel, B-1090Bruiissels, Belgiuimon Received 9October 1985/Accepted 5 May 1986Theinteraction ofmono-and polyclonal neutralizing antibodies with poliovirus was studied. In allcases,
neutralization wasduetoantibody-mediated virus aggregation, and the unpolymerized virions accounted for
the residual infectivity. Theeffect of papainonpreviouslyneutralized viruswastodeaggregate the virustofully infective single virions. With some antibodies, the amount ofaggregated virus regressed in the region of greatestantibodyexcess, even though thevirus remained fullyneutralized. Under theseconditions, noninfec-tive, unaggregated immune complexes were formed. Amutant resistant tooneof the monoclonal antibodies was selected. The mutant virions were still bound but no longer aggregated or neutralized by the selecting
antibodies.
In previous papers, we studied the neutralization of poliovirus by a single hybridoma antibody (MoAb). This
MoAb, 35-1f4, reduced the number of infective units through antibody-mediated virion aggregation (4). After partial
neu-tralization with MoAb 35-1f4, the residual infectivity was
proportional to the remaining amount of unpolymerized virions. The aggregated virions were not intrinsically dam-aged, for they regained theiroriginal sedimentation value of 160S and full infectivity upondispersion of the aggregates by papaintreatment (21). The reduction ofinfectivity (i.e., by definition, neutralization) brought about by MoAb 35-1f4 clearly resulted from nondestructive, antibody-mediated virion aggregation. The question which will be addressed in the present paper is whether this conclusion can be gener-alized.
There exists a vast literature dealing with poliovirus neutralization (e.g., see references 9 and 14). The mecha-nism proposed by Mandel (13), which is based on an
antibody-induced shift in the isoelectric pH of the virus, has been widely accepted but was also recently challenged (7). Eminietal. (11)showed that bivalentbindingof theantibody caused neutralization and pl shift. It was claimed without supporting evidence that nonsaturating, neutralizing
amounts ofantibody failed to cause virus aggregation (10). Neutralization associatedwith stable, monogamous binding of theantibodywasdemonstratedonly for MoAbF7.12(12). and the residual infectivity of the virions was shown to
decrease exponentially as the average number of bound antibody molecules increased. However, MoAb F7.12 also causedaggregation, and itcanbe seen(12: seeFig. 4and 6) thatthismechanism wasthe maincause of neutralization at
the medium range ofantibody/virion ratios.
Baxt et al. (1) showed that at least some monoclonal antibodies directed against foot-and-mouth disease virus
neutralized by an aggregation mechanism.
The analysis originally performed with MoAb 35-1f4was
extended to all the other neutralizing MoAbs in our own panel (5) and to the MoAbs lc (10) and F7.12 (12). The
antibodies oftwoconventional antisera, previously shown to differ in theirability to shift the isoelectric pH of poliovirus (7), were also included in this study. All of the antibodies tested caused polio virions toaggregate, and the relation of proportionality originally derived for 35-1f4 between the
* Corresponding author.
residual infectivity and theremaining amount of unpolymer-ized (160S) virions was generally valid. In the region of
extreme antibody excess, however, noninfective, unaggre-gated immune complexes wereformed.
MATERIALS ANDMETHODS
Virusneutralization assay.Type 1 poliovirus strains la/S3 and Mahoney were used (21). The culture of 3H- or
"S-labeled virus was as previously described (19, 21). Mutantsresistant to MoAb 35-1c3 were isolated as previously de-scribed(17). Neutralization wasdeterminedby plaque assay (3).
Antibodies. Allof theantibodies usedarelisted in Table1.
The sera and ascitic fluids were assayed for neutralizing activity as previously described (3). The dilutions causing 50% plaque reduction fell between
10-2.5
to 10-5. . Noplaque reduction with preimmune sera or control ascitic fluids (induced by nonsecreting SP2/0 cells) was observed.
MoAbs were prepared from ascitic fluids by
(NH4)2SO4
precipitation, labeled by reductive methylation with 14C]for-maldehyde, and tested for possible inactivation (21); only preparations retaining full activity after '4C labeling wereused. Sodium dodecyl sulfate-gel electrophoresis, followed by fluorography, showed that all of the antibody prepara-tions contained at least 80% immunoglobulin. The percent-age of
14C
binding to excess poliovirus was considered torepresent specific antibody content (Table 1). The input ratios mentioned in this paper were corrected for the
pres-ence ofnonspecific proteins in ascitic fluids and sera.
Polyclonal rabbit and guinea pig antibodies (7) were puri-fied and labeled as described above.
RESULTS
Analysisof neutralization mixtures. In previous work, we showed thesedimentationprofiles of neutralizationmixtures made of 3H-poliovirus and
14C-labeled
MoAb 35-1f4. The formation of virion oligomers (antibody-bridged dimers,trimers, etc.) and large aggregates was documented. The
analysis was extended to the 12 other neutralizing MoAbs listed in Table 1andelsewhere (5). Representative
sedimen-tation profiles were selected to document the formation of virion oligomersby using an adequate antibody/virion ratio for each antibody (Fig. 1). In all cases, higher
anti-479
on November 10, 2019 by guest
http://jvi.asm.org/
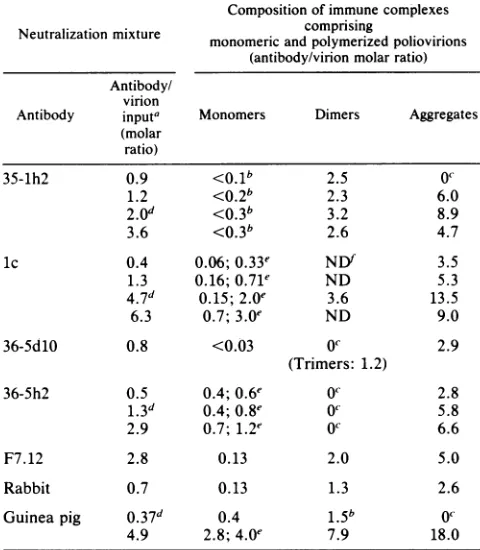
TABLE 1. Properties of the antibodies
Specific Seii
Antibody Isotype" Reference p1 shift"
antiendy
radio-activity"35-1h2 IgG 5 + 50 3.400
lc' IgG 8 NT' 50 24,900
36-5d1O IgM-K 5 + 9( 10,740
36-5h2 IgG3-K 5 + 20 4,350
F7.12v IgG 12 NT 50 5,740
35-1f4 IgG2a-K 4, 21 - 50 5,260
35-1c3" IgG 5 NT NT NT
Rabbit NT 7 - 50 6,000
Guinea pig NT 7 + 5 10,740
DeterminedbytheOuchterlonydoublediffusiontest.
+ Meansthat theantibodyathighconcentraltioncaused theisoelectric pl
of poliovirus toshift to approximatelypH 4.5. These results were hitherto unpublished or takenfromreference 7.
SeeMaterials and Methods.
d 14 C counts per minute per microgram of p-otein (for the labeling
procedure, seeMaterialsandMethods).
This antibody, also known as 1BM 55.6, wais kindly provided by R.
Crainic andB. Blondel.
NT, Not tested.
Kindly provided byR. Rueckert.
MoAb 35-1c3 bound to N aind H aintigens. aindthis double specificity subsistedafterrecloningby single-cellisolation(21)). Thisaintibodvwas not 14Clabeled.
body/virion ratios brought all virusradioactivity down in the pellet. The composition ofvirus-antibodycomplexesisgiven in Table 2. Thebehavior encountered with all immunoglob-ulin G (IgG) antibodies except lc, 36-5h2, and F7.12 (see below) is exemplified by MoAb 35-1h2 (Fig. 1A). Oligomers and aggregates were formed, and the residual 160S particles remained free of antibody. MoAb 36-Sd10, the only IgM in
ourpanel, formed so few oligomers thattheir presence had tobeinferred from the antibody profile (Fig. 1C). No dimers were formed by this antibody, whereas they were predomi-nant among the oligomers formed by the IgG antibodies (e.g., Fig. 1A and B). Only one antibody (36-5h2, Fig. 1D) failed to form any oligomers, although it formed large aggregates. Stable association of the antibody with mono-meric virions was observed only with MoAbs Ic (Fig. iB), 36-5h2(Fig. ID), and F7.12(Table 2) and polyclonalguinea pig antibodies (Fig. 1F).
With MoAb lc (Fig. 1B), the peaks in the antibody and virion radioactivity profiles near the 160S position did not coincide exactly, as theantibodies sedimented more slowly than the virions. As a result, the relative antibody content
varied from 2.0 molecules of MoAb lc per virion in the antibody peak fraction to only 0.15 at the virus peak (Fig. 1B; Table2). The slower sedimentation of virionscarryinga
few antibody molecules was probably due to increased friction.
Table 2 shows the composition of the virus-antibody complexes sedimenting as single virions, oligomers, or ag-gregates. Therelativeantibody content of the aggregates (as defined in thelegend to Fig. 1) alwaysexceeded that ofthe
dimers, as well as the input antibody/virion ratio. The oligomers contained at least twice the minimum number of antibodymoleculesrequiredtolink thevirionstoeachother (e.g., one divalent antibodymolecule foradimerortwofor
atrimer), aswas also found with MoAb 35-If4(21).
It was previously reported (12) that MoAb F7.12formed virion oligomers and that it also bound stably to single virions (two properties not seen together with the MoAbs generated in this laboratory). We confirmed these
observa-tions. The results with F7.12were similartothose obtained with MoAb Ic (Fig. 1B).
Thefact that 36-5h2 antibody molecules remained bound to 160S particles did not mean that this MoAb neutralized poliovirus by inactivating single virions. Ataninputratioof 2.5antibodies pervirion, the infectivity of the mixture was
reduced to 10% of that of untreated virus. After ultracen-trifugation, it was found that only circa 10% of the virus input sedimented as 160S material. These single virions carried an average of one antibody molecule and retained 80% of the specific infectivity of untreated virus. The re-mainder of the virus input was recovered as aggregates of low (9%) specific infectivity. Thus, even though MoAb 36-5h2 formed stable complexes with single virions,
aggre-gation was still the main factor in neutralization by this antibody. For MoAb F7.12, the slope of residualinfectivity
versus the antibody/virion ratio in monomeric complexes indicated that infectivitywasnotsignificantly reduced when only one antibody per virion was bound (12). Ourfindings with MoAb 36-5h2 therefore do not contradict those of Icenogle etal. (12)
Figure 1 and Table 2 also show data on neutralization
(I
0
z
i
0
a
7
;5l3
0 5 10 15 0 5 10 15
TOP VOLUME ( ml )
FIG. 1. Sedimentation profiles of mixtures of 3H-labeled poliovirus and 14C-labeled antibodies. 3H-labeled poliovirus strain la/S3was used,except for MoAb lc (B), where it was replaced by Mahoney. The final virus concentration was 24 nM. '4C-antibody wasadded to the virus in the molar ratio shown below. The buffer was PBS-A (140 mM NaCI. 6.5 mM KCI, 5.5 mM phosphate. 0.5 mM MgCIl. 0.1 mM CaCI2 containing 0.1 mg of bovine serum albumin per ml, final pH7.2). Themixtures,in afinalvolumeof 100
.1l,wereincubated for1h at37TCand overnight at 4°C. Aftertaking a 10-p.I sample for plaque assay. we layered the remainder onto 14.5-ml 15 to 30% sucrose gradients in phosphate-buffered saline and centrifuged it for 2.5 h at 85,000 x g.a at4°C in an SW30 rotor (Measuring and Scientific Equipment, Ltd.). The pellet was sus-pended in 300 ,ul of PBS-A. and all fractions were analyzed for radioactivity as previously described (21). Under the conditions of centrifugation used, complexes larger than hexamers reach the bottom of the gradient. These complexes are referred to as aggre-gates. The amounts of virus and antibody in aggregates are repre-sented by closed and open bars, respectively. The input anti-body/virion molarratios(Ab/V) and residual infectivities (RI)ofthe mixtures before centrifugation were as follows: A, 2.0 Ab/V, RI 90%: B. 4.7 Ab/V. RI 85%: C, 0.5 Ab/V, RI45%: D, 1.3 Ab/V, RI 55%: E. 0).7Ab/V, RI 65%: F, 0.4Ab/V, RI 95%.
on November 10, 2019 by guest
http://jvi.asm.org/
mixtures made with antibodies purified from conventional rabbit and guinea pig antisera. The rabbit antibodies (Fig. 1E) behaved like themajority of the MoAbs; oligomers and large aggregates were formed, and little antibody remained bound to single virions. In contrast, the guinea pig antibodies (Fig. 1F) yielded few oligomers, and more antibody re-mained bound to 160S virions.
Relation of residual infectivity to the amount of 160S virions remaining after neutralization. Although the amount of virus in oligomers was variable, depending on the antibody used, it never exceeded 5 to 10% of the total virus input. The specific infectivity oftheoligomers was reduced but not nil, as was also reported for 35-1f4 (21) and F7.12 (12). There-fore, incorporation of virions into oligomers contributed only slightly to neutralization. With all of the antibodies listed in Table 1, the specificinfectivity of aggregated virus was between 1 and 10% of that of untreated controls (data notshown).
The residual infectivity of virus-MoAb 35-1f4 mixtures correlated well with the percentage of160S virus remaining afterneutralization, as shownby sucrose gradient centrifu-gation (21).Allavailable data on antibodies otherthan35-1f4 are shown in Fig. 2. In all ofthe cases studied, the same linear relationwasfound asfor 35-1f4. Thus, at least for all of the antibodies presently studied, the loss ofinfectivity was mainly dueto virion aggregation.
Effect of papain on previously neutralized poliovirus. As
TABLE 2. Composition of virus-antibody complexes isolated aftersucrosecentrifugationof neutralization mixtures
Composition of immune complexes Neutralization mixture comprising
monomericandpolymerizedpoliovirions (antibody/virionmolarratio) Antibody/
virion
Antibody inputa Monomers Dimers Aggregates (molar
ratio)
35-1h2 0.9 <0.b 2.5 0C
1.2 <0.2b 2.3 6.0
2.0d <0.3b 3.2 8.9
3.6 <0.3b 2.6 4.7
lc 0.4 0.06;0.33e NDf 3.5
1.3 0.16;0.71e ND 5.3
4.7d 0.15; 2.0e 3.6 13.5
6.3 0.7; 3.0e ND 9.0
36-5dlO 0.8 <0.03 OC 2.9
(Trimers: 1.2)
36-5h2 0.5 0.4;0.6e 0o 2.8
1.3d 0.4;0.8e 0o 5.8
2.9 0.7;1.2e OC 6.6
F7.12 2.8 0.13 2.0 5.0
Rabbit 0.7 0.13 1.3 2.6
Guineapig 0.37d 0.4 1.5b 0
4.9 2.8;4.0e 7.9 18.0
aType1poliovirus of strain Mahoneywasused with MoAbs F7.12 and lc;
strain la/S3wasused in allothercases.
bOwingtotrailing of14C-antibody,theseratiosareroughestimates.
' Nosufficient dimers(oraggregates)werepresent. dDatafrom theprofileshowninFig. 1.
eThe first figure refersto the virus peak, and the secondrefers to the
antibody peak (seethe textfor furtherdetails).
fND,Notdetermined.
1001-x
'U
U1)
0
to
z
U)
n
:
5
LL
0
z
uJ
C.)
501
0
.
0 0
.
0 0
100 50 0
RESIDUAL INFECTIVITY (PERCENT)
FIG. 2. Relationship between residual infectivity andamountof residual 160S virions after neutralization. Residual infectivity of neutralization mixtureswas measuredas describedpreviously (3). Themixtureswereanalyzedasdescribed in thelegendtoFig. 1, and theamountofunpolymerized virionswascalculated.O,36-5d1O; *, 36-5h2; *, 35-1c3; 0, rabbitantibodies. -, Theoretical
expecta-tion assuming infectivity to be exclusively associated with unpolymerized virions.---,Theoretical expectation assumingthat neutralization wasunaccompanied bypolymerization.
reported elsewhere (21), papain acting on poliovirus
previ-ously neutralized and aggregated by MoAb 35-1f4 caused deaggregation, restoration of infectivity, and release of fully infective, monomeric virions.
Werepeatedtheexperimentwith MoAbslc,36-5d1O,and
36-5h2 and polyclonal rabbit antibodies. Enough antibody (fourtosevenmolecules ofantibodypervirion)wasusedto aggregate the virus completely (Fig. 3) and to lower the
infectivity of the mixturesto8%of theoriginalorless (Table
100DA B C D
1 36-5d0 36-5h2 Rabbit
,,50-<
0 5 10 15 0 5 10 15 0 5 10 15 0 5 10 15
TOP
VOLUME (ml)
FiG. 3. Deaggregation of neutralized poliovirus by papain. To 2.4 pmol (50,000 cpm) of 35S-Mahoneyvirus (Ato C and E to G) or 0.8 pmol (50,000 cpm) of3H-laIS3 virus (D and H) the following amounts of antibody were added in a final volume of 100 p.1 of phosphate-buffered saline minus divalent cations, supplemented with 2 mML-cysteine and 10 mM EDTA. A and E, 18 pmol of1c;
BandF. 10pmol of
36-Sd10;
C and G, 16 pmol of36-5h2; D and H, 60 pmol of polyclonal rabbitantibodies. After1h at37°C, 12.5 u1lof buffer (A to D) or buffer containing 2.5 ,ug papain (E to H) was added. After 4 h at37°Cand overnight at20°C,10pu1
was taken to determine the residual infectivity, and the remainder was layered onto 15 to 30% sucrose gradients made in phosphate-buffered saline. Centrifugation was for 3 h(AtoC and EtoG)orfor 2.5 h (D and H) at85,000x at 4C in anpm MSESW3Orotor. Forfurther details, see the legend to Fig. 1. The arrows indicate positions to which untreated virus sedimented.on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.349.515.80.238.2] [image:3.612.54.294.394.669.2] [image:3.612.314.556.445.590.2]TABLE 3. Reactivation ofpoliovirus by papain Residualinfectivity Specificinfectivity Addition" ofmixture %) of 160Sparticlesb
None 100 100
Monoclonal antibodies
1c' 2 0
lc + papain' 50 106
36-5dlOc 6 0
36-5dlO + papain' 33 114
36-5h2' 4 0
36-5h2 + papain' NDe 80
Rabbitantibodies' 8 0
Rabbit antibodies + 86 68
papainc
aSeethelegendtoFig.3for details.
Ratio of PFUtoradioactivity, assayed aftersucrosegradient isolation. Sedimentation profile shown in Fig. 3.
d No 160S material available. ND,Notdone.
3). After papain treatment, infectivity was restored to at least 33%, and the
bulk
of the virus again sedimented as single virions (Fig. 3) of full specific infectivity (Table 3).Effectofantibody concentration on the formation of large aggregates. The experiments reported above were carried out at high virus concentration (7 x
109
PFU/ml). In this concentration region, the degree of neutralization depends on theantibody/virion ratio. At lower virus concentrations (i.e., less than 1.5 x 108PFU/ml), thedegree of neutraliza-tion is determined solely by the antibody concentration (percentage law; reference3). We examined whether aggre-gation also occurred when the percentage law applied. Therefore,we used theminimumvirusconcentration allow-ing radioactivity measurements, i.e., 5 x107
PFU/ml or 0.016 nMvirus.Toavoid laborious sucrose gradientanalyses, the amount of large aggregates was estimated by a simpler method. When neutralized mixtures were centrifuged in conical 1.5-ml tubesfor 10min at 10,000 x g, aggregated virus was pelleted, whereas monomeric virions and oligomers
re-mained in the supernatant. Using this procedure, we mea-sured the percentage of virus in the pellet as a function of antibody concentration. In some cases, the residual infectiv-ity of the mixtures was also determined before centrifuga-tion. Figure 4 shows the results. Most MoAbs and sera yielded athree-legged curve. At first, the amount of aggre-gated virus increased withincreasing antibodyconcentration until total aggregation was reached withmostantibodies. At a large antibody excess, however, the amount ofpelleted virus tended to regress as the antibody input was further increased. The findings confirmed that allneutralizing anti-bodies caused virus aggregation. However, whereas some MoAbs caused totalaggregation over a wideantibody con-centration range (Fig. 4A, D, E, and H), this range was narrower with other MoAbs (Fig. 4B and C), and in two cases the virus was incompletely pelleted at any anti-body/virusratio(Fig.4FandG). Ourfindings of incomplete aggregation with antibody F7.12 may seem at variance with those ofIcenogle et al. (12), who reported complete aggre-gation. However, when we applied the same centrifugal conditions as used by them (and by ourselves [Fig. 1]), the virus was indeed fully aggregated. The low-speed centrifu-gation method evidently yielded a lower estimate of the extent of virus aggregation.
Neutralization curveswere determinedfor four antibodies (Fig. 4B, D, G, and H). They paralleledthe risingleg of the aggregationcurves; in three cases(Fig. 4D,G, and H), the neutralization curve preceded the aggregation curve by about 0.5log10. This does notcontradictthe resultsinFig. 2,
80 30
20V
1--i
LU CL -J LU
0.
0 LU
z U)
100
50
0 100
50
0
A B C D
35-1h2 lc 36-5d10
36-5h2-E F G H
Rabbit Ginea pig F7.12 35-lf44
-2 -1 0 1 -2 -1 0 1 -2-1 0 1 -2-1 0 1 2
pmol Ab (loglo)
100
z
50 m
c --4 m
0
r-N
100 >
I z
50
-0
wo
I 10
0 80 30'
20
10
0
A IB
=I
I.~~~I.
141
- ;
-,1-0 2.5 5.0 0 2.5
TOP
VOLUME (ml)
5.0
[image:4.612.356.526.434.627.2]4N....
FIG. 4. Formation oflarge poliovirusaggregates bymono- and polyclonal antibodies. To 1,000 cpm of3H-poliovirus, i.e., 0.003 pmol of Mahoney (0) or 0.005 pmol of la/S3 (0), the indicated amountofspecific antibodywasaddedinafinalvolumeof100p.lof PBS-A(seethelegendtoFig. 1).After16hat23°C,10 p.1wastaken for the plaque assay, and the remainder was centrifuged as de-scribed in thetext. Theradioactivity ofthe supernatant andpellet (suspended by heating at 56°C for 5 min in 1% sodium dodecyl sulfate)wasdeterminedandexpressedas apercentageof theinput. Preimmuneseraneveraggregatedmorethan10%of theinputvirus (data not shown). Neutralization experiments were done with Mahoney virus.
FIG. 5. Formationofaggregates and single-virioncomplexesas
a,function ofantibody concentration. To 0.003 pmol ofMahoney virus (3,300 cpm) thefollowingamountsofMoAb lcwereadded in afinalvolumeof100p.1of PBS-A(seethelegendtoFig.1):A,none; B,0.5pmol; C,S pmol; D, 50 pmol.After 16 hat 23°C, 10pI. was
taken for plaqueassay, and the remainderwascentrifuged for 45 min inanMSESW60rotor at205,000 X gavina15to30% sucrose gradientmadeinphosphate-buffered saline. Fractionsof 300p.lwere
collected and counted for radioactivity. The bar represents the radioactivityin thesolubilizedpellet.Theresidualinfectivityof the mixtures beforecentrifugationwas:A, 100%; B,56%; C,0.6%;D, 0.5%.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.64.302.486.626.2]as the supernatants of low-speed centrifugation contained small aggregates in addition to single virions. The virus remained fully neutralized, even when aggregation re-gressed.
Inimmunoprecipitation assays mediated by Staphylococ-cus aureus Cowan I (23), MoAb 35-1h2 bound to laIS3 but not to Mahoney virus. The reverse was true of F7.12. All of theotherantibodiesbound to both strains of poliovirus. To assess the possible influence of the virus strain on the results, a comparison was made between the la/S3 and Mahoney viruses with someantibodies that recognized both (Fig. 4B, E, and F). Similar aggregation curves were ob-tained withboth virus strains.
Formation of single-virion immune complexes in the region of high antibody excess. Theaggregation curve of MoAb Ic (Fig. 4B)
illustrated
the negative effect of great antibody excessesonaggregationparticularlywell. Thisantibodywas therefore chosen to study the physical condition of the virions after reacting with a great antibody excess.Labeled poliovirus (0.030 nM) was incubated with anti-body lcin concentrations ranging from5to 500nM,andthe mixtures were centrifuged in 15- to 30% sucrose gradients (Fig. 5). Antibody at aconcentration of50 nMcaused all of the input virusto aggregate and neutralized the mixture by more than 99%. With 500 nM antibodies, only 10% ofthe virus remained in the pellet, and most ofthe virus radioac-tivity sedimented slightly slower than native virions (com-pare Fig. 5, panels A and D). These particles were by no means nativevirions,however, as theresidual infectivityof themixture was only0.5% oftheoriginal.
Tofindoutwhether theseparticles carried antibody,0.030 nM3H-labeled virus wasincubated with250 nM
"4C-labeled
antibodylc, and the mixturewascentrifugedin a 15 to30% sucrosegradient. The result is showninFig. 6B. The pellet containedonly anegligible amount of virus orantibody. As
30- A 30-c
20 2020
10 _ 10 |
30 0
30-
~~B
30-D -20-10 10 >
20 ~ jO.8 20 ".40 co
10 0.4 10 -0.2
0 5 10 15 0 2.5 5.0
TOP TOP
VOLUME (ml)
FIG. 6. Sedimentation of native poliovirus and single-virion complexes in two different sucrose gradients. To 0.003 pmol of
3H-Mahoney virus either no antibody (A and C) or 25 pmol of
'4C-labeledantibodylc(BandD)wasaddedinafinal volume of100 ,ul of PBS-A (see thelegendtoFig. 1). After 18 h at 23°C, 10 ,u was taken forplaqueassay, and the remainder wascentrifuged in 15 to 30% (A and B) or46 to52% (Cand D) sucrosegradients made in phosphate-buffered saline. GradientsAand B were centrifuged for 2.5 h at 4°C at 85,000 X gav and fractionated into400-pdfractions. Gradients C and D were runfor 17 h at4°Cat 205,000 X ga,and fractionatedinto200-pulfractions. Bars represent theradioactivityin solubilized pellets. Owing tothe largeadmixture of14C in the top fractions,the3Hradioactivitycould not be determined accurately in thesefractions(B and D).
I-LU
a
-J
-J
LUI 0.
0
z
C,)
100
50
0
X
I
Ie
I I1073
100
50
0
z m c
~-q
--4
0 z
1lo1
DILUTION OF ASCITIC FLUID
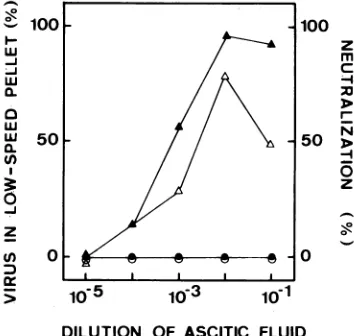
FIG. 7. Neutralization and aggregation of poliovirus by MoAb 35-1c3: comparison of the wild type and aneutralization-resistant mutant. Ascitic fluid of 35-1c3 was serially diluted 10-fold in phosphate-buffered saline.Thedilutionsweremixed with1,000 cpm of 35S-labeled la/S3 or mutant virus(resistant toneutralization by 35-1c3). Incubationwasfor1h at37°C. Theamountof viruspelleted was determinedasdescribed in the legendtoFig.4.Virus inpellet: wild type, A; mutant, 0. Neutralization: wildtype, A; mutant, 0.
in the preceding experiment, the main peak ofvirus
radio-activity sedimented slightly slower than untreated virions
(compare Fig. 6A and B). As might be expected from the
7,500-fold molarexcessofantibody,the bulkof theantibody remainedfree. However, some antibodywasassociatedwith
the virion peak as well as with minor,
faster-sedimenting
virus material (presumably virion di-andtrimers). From the known specific radioactivities ofthe virus and antibody, it wascalculated that eachmonomeric virion carriedabout 100 moleculesofIgG antibody.
It seemedincredible thatamassincrease of 15 million (150,000 x 100) daltons would leave the sedimentation
velocity
ofthevirions almostunchanged. However, Icenogle et al. (12) also found no significant change in the sedimentation velocity of virions when 30 molecules of F7.12 antibody were bound to each virion. It waspossible thattheeffect oftheincreasedmass wasoffset by thelower density andhigher frictional coefficient ofthe immune complexes; if that were the case, increasing thedensity
andviscosity
ofthe mediummight
beexpected
toslow the immunecomplexesmorethan thenativevirions.To
testthis,amixtureofvirus andantibody lcwas preparedas
described above,but
centrifugation
wascarriedoutina46to52% sucrose gradient instead of in the usual 15 to 30% gradient. Inthis
highly
viscous medium, the immunecom-plexes sedimented about 20% slower than native virions (compare Fig. 6C and D), thus confirming the theoretical expectation.Thecomposition oftheimmunecomplexeswas
unchanged, i.e., about 100 antibody molecules per virion. Relationship between binding, aggregation, and neutraliza-tionofamutant virus. Mutantsresistanttoneutralization by
MoAb 35-1c3 were selected within the la/S3 virus strain.
Constant amounts of the wild type orthe resistant mutant
were incubated with various concentrations of MoAb
35-1c3,
and aggregation and residual infectivity weremea-sured (Fig. 7). The neutralization curve of the wild-type virus preceded the upward leg of the aggregation curve by 0.5
log1o
in antibody concentration, as also observed with other MoAbs (see above). The sedimentation profile (datanot shown) andaggregation of the wild type were
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.345.522.74.242.2] [image:5.612.55.297.437.598.2]ble to those shown in Fig. 1A for MoAb 35-1h2. On the other hand, the mutant was neither measurably neutralized nor aggregated by the selecting MoAb 35-1c3 at any concentra-tion (Fig. 7). Remarkably, protein A-aided immunoprecipi-tation experiments showed thatantibody 35-1c3 still bound to the mutant, although with less affinity than to the wild type. The immunoprecipitation titer of antibody 35-1c3, as measured against the mutant virus, was 1.3 log10 lower than thetitermeasuredagainst the wild-type antigen. Thus, at the higher antibody concentrations (e.g., at the 10-1 dilution [Fig. 7]), the virions were still recognized by the selecting antibody, but they were neither aggregated nor neutralized. Inconclusion, the ability to bind tovirionsis a necessary but not a sufficient condition of neutralization; virions are neu-tralizedonly iftheantibodyalso causes them to aggregate.
DISCUSSION
Two situations were distinguished by the antibody con-centration. Either the concentration was just sufficient to neutralizethevirusby about 2 log10, orit wasmuch greater than that.
The region of moderate antibody excess. The ability to
cause poliovirus aggregation, formerly documented for MoAb35-1f4, was found to be a general property of all of the neutralizing MoAbs we examined, which weregenerated in threedifferentlaboratories. All antibodies studied converted 160S virionsto aggregates of high antibodycontentand low residualinfectivity. The infectivity loss of partly neutralized mixtures waswell correlated with the loss of160S particles to aggregation. It was concluded that aggregation was the basis of neutralization by all of theantibodies studied. This conclusion was strengthened when a mutant resistant to
neutralization by one of these antibodies was studied. Even though the antibody still recognized the mutant virions in immunoprecipitation tests, it no longer aggregated or neu-tralizedthem. Resistant mutants thatwerestill boundby the selecting antibody were also reported by Blondel et al. (2).
It mightbe argued that aggregation is secondary to neu-tralization, i.e., that aggregation occurs only after antibodies bind to single virions and inactivate them. If this were the case, one would expect a substantial contribution of unag-gregated, noninfective immune complexes. At leastat
mod-eratedegrees of neutralization, atwhich theamountof 160S materialcould still be measured,this was not so (Fig. 2).
When single virions were released from aggregates by papain digestion, theywerefullyinfective. Toreconcile this fact with the hypothesis ofpreaggregation inactivation
re-quirestheadditionalhypothesisthat thedamagewas
revers-ible by papain. The least contrived explanation is that the virionssufferednointrinsic inactivation, being simply linked into aggregates by the antibody and released by papain.
Aggregationwasnotrestrictedtoexperimental conditions involving huge virus concentrations suchasthoseusedwhen neutralization mixturesare tobeanalyzed bysucrose
gradi-entcentrifugation. To showthis, the relatively low
concen-tration of 5 x 107 PFU/ml (0.016 nM) was used, and a
simplified techniquewasused, in which the aggregateswere collected by low-speed centrifugation. Without exception, all antibodies again caused virus aggregation. With most
antibodies, the amount ofpelleted virus regressed in the region ofextreme antibodyexcess (seebelow).
Itwas claimed that some MoAbs (including lc) failed to causepoliovirusaggregation(10). However,nomentionwas
madeof either the virus orantibody concentration. When a
single antibody concentrationisused,and thishappenstobe
outside the rangeatwhich virusaggregation occurs,onemay erroneously conclude that the antibody does not cause
aggregation. This possibility is particularly evident for MoAb lc(Fig. 4B).
In conclusion, (i) for all antibodies studied, aggregation wasthe mostimportant and, inmost cases, theonlyknown mechanism ofneutralization, and (ii) direct inactivation of unpolymerized 160S particles (12) contributed little to
neu-tralization.
The region of great antibody excess. Neutralization may
occur without aggregation, but only in the presence of an enormousantibodyexcess(Fig. 5). Whereasantibody Icat a concentration of 50 nM ensured full aggregation and neutralization(Fig. SC),a10-times-higherconcentrationwas
required for full conversionto single-virion complexes. Recently, McCullough (16) reported aggregation of foot-and-mouthdisease virus by amonoclonal antibody andthe formation of monomeric complexes when 100 times more
antibody
was used.Our findings are in agreement with the immune lattice theory (15, 18). The theory
predicts
formation of virion oligomersin theregionofantigenexcess,oflargeaggregates in balanced virus-antibody mixtures, and ofsingle virions, surroundedby
asmanyantibody
moleculesastheirvalence allows, when the antibody is in great excess. Poliovirus is expected tocarry a maximum of 60molecules ofantibody, provided each is attached by a single paratope, as wasobserved with tobacco mosaicvirus in the
region of
extremeantibody excess (22). That the actual number of antibody molecules per virion was even higher suggests that unspe-cific
binding
occurred. Theantibody-saturated
virions re-mainednoninfective.In
conclusion,
theoccurrenceof virusaggregationand the contributionofaggregation
tolossofinfectivity oftheviruspopulation (i.e., neutralization)
depends on at least three kinds of factors: (i)the physicochemical environment: low-ering the ionic strength may replace an aggregative by adisruptive
mechanismofneutralization [6];(ii) theantibody
concentration,
asdiscussedabove;
and(iii)
thepreference
of each monoclonal antibody for mono- or bigamousbinding.
Monogamous, bivalentbinding
of MoAbs 35-1f4 [21],35-1h2,
and36-5d1O
was not observed; on the otherhand,
MoAbs lc, 36-5h2, and F7.12 were capable of bothmono-and
bigamous
binding;
antibodiesincapable
ofbigamous
binding may well exist but have not yet beenencountered.ACKNOWLEDGMENTS
Wethank R. Crainic,B. Blondel,and R. Rueckert fortheir kind giftsofasciticfluids, B. Rombautfor criticalcomments, and M. De Peismacker, A. De Rees,and S. Peeters fortechnical assistance.
Thisinvestigation is part ofaconcerted research programofthe Vrije UniversiteitBrussel and theDepartmentofScientific Program-ming (Belgium).
LITERATURE CITED
1. Baxt,B.,D.0. Morgan,B. H. Robertson,and C. A.Timpone. 1984. Epitopes on foot-and-mouth disease virus outercapsid protein VP,involved in neutralization and cell attachment. J. Virol. 51:298-305.
2. Blondel, B.,R.Crainic,0.Fichot,G.Dufraisse,A.Candrea,D. Diamond, M. Girard,and F. Horaud. 1986. Mutations confer-ringresistanceto neutralization with monoclonal antibodies in type 1polioviruscanbelocated outsideorinside the antibody-bindingsite.J. Virol. 57:81-90.
3. Brioen, P.,and A.Boeye.1985. Poliovirusneutralizationand the percentage law. Arch.Virol. 83:105-111.
4. Brioen, P., D. Dekegel, and A. Boeye. 1983. Neutralization of
on November 10, 2019 by guest
http://jvi.asm.org/
poliovirus by antibody-mediated polymerization. Virology 127:463-468.
5. Brioen, P., R. J. Sijens, R. Vrijsen, B. Rombaut, A. A. M. Thomas,A. Jackers, and A.Boeye. 1982. Hybridoma antibodies topoliovirus N and H antigen. Arch. Virol. 74:325-330. 6. Brioen, P., B. Rombaut, and A. Boeye. 1985. Hit-and-run
neutralization of poliovirus. J. Gen. Virol. 66:2495-2499. 7. Brioen, P., A. A. M. Thomas, and A. Boeye. 1985. Lack of
quantitative correlation between the neutralization of poliovirus and theantibody-mediated pl shift of the virions. J. Gen. Virol. 66:609-613.
8. Crainic, R., B.Couillin, B. Blondel, N. Cabau, A. Boue, and F. Horodniceanu. 1983. Natural variation of poliovirus neutraliza-tion epitopes.Infect. Immun. 41:1217-1225.
9. Dimmock, N. J. 1984. Mechanisms of neutralization of animal viruses. J. Gen. Virol.65:1015-1022.
10. Emini, E. A., S.-Y. Kao, A. J. Lewis, R. Crainic, and E. Wimmer. 1983. Functional basis of poliovirus neutralization determinedwithmonospecific neutralizing antibodies. J. Virol. 46:466-474.
11. Emini, E. A., P. Ostapchuk, and E. Wimmer. 1983. Bivalent attachmentofantibodyontopoliovirusleadstoconformational alterationandneutralization. J. Virol. 48:547-550.
12. Icenogle, J., H. Shiwen, G. Duke, S. Gilbert, R. Rueckert, and J. Anderegg. 1983. Neutralization ofpoliovirus by a monoclonal antibody: kineticsand stoichiometry. Virology 107:412-425. 13. Mandel, B. 1976. Neutralization ofpoliovirus: ahypothesis to
explainthe mechanism and one-hit character of theneutralizing reaction. Virology69:500-510.
14. Mandel, B. 1985. Virusneutralization,p. 53-70. In M. H. V.van
Regenmortel and A. R. Neurath (ed.), Immunochemistry of viruses. Elsevier Biomedical Press. Amsterdam.
15. Marrack, J. R. 1938.Thechemistry of antigensandantibodies. p. 151. HisMajesty's StationeryOffice. London.
16. McCullough, K. C. 1986. Monoclonal antibodies: implications for virology. Arch. Virol. 87:1-36.
17. Minor, P. D., G. C. Schild, J. Bootman, D. M. A. Evans, M. Ferguson, P. Reeve, M. Spitz, G. Stanway, A. J. Cann, R. Hauptmann, L. D. Clarke, R. C. Mountford,and J. W. Almond. 1983. Location and primary structure of a major antigenic site forpoliovirus neutralization. Nature (London) 301:674-679. 18. Pauling, L. 1940. A theory of the structure and process of
formationof antibodies. J. Am. Chem. Soc.62:2643-2657. 19. Rombaut, B., R. Vrijsen, P. Brioen, andA. Boeye. 1982.A pH
dependentantigenic conversion of empty capsids of poliovirus studied with the aid of monoclonal antibodies to N and H antigen.Virology22:215-218.
20. Sijens, R. J., A. A. M. Thomas, A. Jackers, and A.Boeye. 1983. Clonal isolation ofhybridomasby manual single-cell isolation. Hybridoma 2:231-234.
21. Thomas,A. A.M.,P.Brioen,and A.Boeye.1985. Amonoclonal antibody that neutralizes poliovirus by cross-linkingvirions. J. Virol. 54:7-13.
22. Van Regenmortel, M. H. V., and G. Hardie. 1976. Immuno-chemical studies of tobacco mosaic virus. Immunochemistry 13:503-507.
23. Vrijsen, R.,B.Rombaut, andA. Boeye.1983. A simple quanti-tative protein A micro-immunoprecipitation method: assay of antibodies to the N and H antigens ofpoliovirus. J. Immunol. Methods 59:217-220.