Copyright © 1984, AmericanSocietyforMicrobiology
Cloning
and
Structural
Analysis of
Integrated
Woodchuck Hepatitis
Virus
Sequences from
a
Chronically Infected Liver
CHARLES E. ROGLERt* ANDJESSE SUMMERS
Institute for Cancer Research, Philadelphia, Pennsylvania 19111
Received 30 November 1983/Accepted14 February 1984
Wehaveisolated anddeterminedthestructureofarecombinant clone in lambda phage Charon 30 which contains woodchuck hepatitis virus sequences integrated in woodchuck genomic DNA sequences. This
clone, incontrasttopreviously reported clones (Ogstonetal., Cell 29:385-394, 1982),was isolated froma chronicallyinfected liver whichneverdeveloped hepatocellularcarcinoma. Southernblot analysis of viral sequences in the clone in conjunction with electron microscope heteroduplex analysis showed that the
integratedviralsequencesdidnotcontaininternalrearrangements,ashave thosefrom hepatomas, butwere
colinear with the cloned viral genome except for the deletion of approximately 500 base pairs of viral sequences (betweenpositions 1,000and 1,550ontheviral map). Therefore, the integrationwasprobablya defective genome incapable of supporting viral replication. However, the complete open reading frames
coding for the viral X, core, presurface, and surface antigengenes werepresent, indicatingthat the viral sequences could code for viral antigens. Southern blot analysis of the normal cellular flanking sequences, usingflankingsequenceprobes from the clone, showed thatnodetectablerearrangementsof cellularDNA
(less than 50 base pairs) had occurredatthe site of viral integration.
Woodchuck hepatitis virus (WHV) isoneoffourmembers of thenewly recognized "hepadna"groupof viruses (includ-ing also hepatitis B virus [HBV], ground squirrel hepatitis virus, and duck HBV) (16). The hepadna viruses havemany common characteristics,
including
the overall structure of thevirus,
similar size and unusual structure of the viral DNA, similar organization of viral genes, immunologicalcross-reactivity,
and thetendencytocause persistentinfec-tions (9, 10, 21,23). The cytopathiceffects of theseviruses,
however,
varygreatly
intheirrespective
hosts.WHVclose-ly resembles the human virus (HBV) in that it causes a
variety of cytopathic effects including chronic persistent hepatitis and chronic active hepatitis similartothat observed inhumans. Chronic active hepatitis in woodchucks hasbeen shownto lead, almost
invariably,
tohepatocellularcarcino-ma
(HCC)
(19; G. Tyler, Ph.D.thesis,
University
ofPenn-sylvania,
Philadelphia,
1984).Significant advances inourunderstanding ofthehepadna virus life cycle have recently been made. Summers and Mason (22) have isolated immature virion coresfrom duck
hepatocytes
infected with duck HBV. The cores contain agenome-sized RNA which is reverse transcribed by an
endogenus polymerase (reverse
transcriptase) activity.
Thefull-length
minus-strandDNAthusgeneratedisthenusedas atemplate for plus-strand DNA synthesis. Full-lengthRNAtranscripts coded by the DNA minus strand have been identifiedin duck liver(11).These RNAmolecules arelikely candidatestobe precursors of thosepackaged intocoresfor reversetranscription. Thisreplication mechanism, although similar to
retroviruses,
is notexactly
identical because a proteinbound to the 5' endoftheminusstrandis believedtofunction asaprimerfor reversetranscription (13) insteadof
a tRNA, as is used by retroviruses (7). All four of the
*Correspondingauthor.
tPresentaddress: Liver ResearchCenter, Departmentof Medi-cine, Albert Einstein College ofMedicine, Bronx,NY 10461.
hepadna
viruses have a protein bound to the5' end of thevirion minus strand
(21).
Integrated hepatitis
virus DNA has beenfound in almostallHCCsarisingfrom livertissuewhich has been
chronically
infectedwith either HBV or WHV.The existence of these viral integrations in hepatomas has implicated them in the
oncogenic process. However, the study of cloned viral
integrations (2, 8,
14)has notyetrevealedaspecific mecha-nism for oncogenesis induced by such integrations. The presence ofmultiple integrationsin tumors, which arepre-sumably of clonal origin, indicates that viral
integrations
may occur in cells before the cell becomes transformed. Theseintegrations would beclonally propagated once a cell
became transformed. Several investigators have used the
Southern blot technique to demonstrate the presence of
apparently integrated HBV DNA sequences in the
high-molecular-weight DNA from nontumorous chronically
in-fected liver tissue (1, 3, 18). In their experiments,
hybrid-ization was observed as either a diffuse smear in the
high-molecular-weight DNA region or, in some cases, as specific bands.A diffuse smear ofhybridizationcouldbedue
to the random integration of HBV in different cells, and weak bands could be generated by localized clonal propaga-tion of cells(focal clonal growths) containing such
integra-tions.
We have used molecular cloning in an effort to
directly
search for integrated WHV DNA in chronically infected hepatocytes.This approach has led to thediscoveryoflong formsof WHVDNAinchronicallyinfected nuclei whichwe
called "novel forms" (17). We have isolated similar novel
forms of ground squirrel hepatitis virus from persistently infectedground squirrel hepatocytes, using molecular clon-ing (P. L. Marion et al.,manuscriptinpreparation). We now reportthe structural characterization of an additional form of WHV for chronically infected hepatocytes in which WHV DNA is integrated in woodchuck cellular DNA sequences. In brief, our findings show that the integrated viral se-quences could not function as a provirus; however, they
832
on November 10, 2019 by guest
http://jvi.asm.org/
INTEGRATED WHV DNA FROM CHRONIC INFECTION 833
could support the
production
of viral core,surface,
and Xantigens during
persistent
infection.MATERIALSANDMETHODS
Animals and strains. Woodchuck HW197 (Marmota monax L.) was housed atthe PenroseResearch Laboratory ofthe Zoological Society of Philadelphia, Philadelphia, Pa. This animal had chronic active hepatitis for several years
before it died withouteverdevelopingahepatoma. DNAfor this study and a previous study was obtained from a liver
biopsy sample taken in October 1980. The tissue sample showedno signsofHCC,and the nuclear DNA containedan abundance ofcovalently closed circular and open circular
molecules.
Lambdaphagevector Charon 30(15) waskindly
supplied
byF. R. Blattner. Recombinant DNAwaspackaged in vitro
inEscherichia coli LE392
by
the method ofEnquist
and co-workers (4, 5).Cloning and Southern blot analysis and electron microsco-py. Nuclear DNA was extracted from liver HW197 as previously described (14). Lambda
phage
Charon 30 DNA and DNA from cloneHW197-2 were isolatedby themethodof VandeWoudeetal.
(24)
with minormodificationsreport-ed earlier (17). The
cloning
experiment
in whichpartial
EcoRI digests of HW197 nuclear DNA were cloned into Charon30 was
reported
earlier(17). ThreeWHV-containing
clones obtained by this
procedure
were reported earlierto contain novel forms ofWHV. Inthisreport,wedescribethe structural characterization ofafourth clone obtainedby
the same procedure which contains WHV sequencesintegrated
in cellularDNA.
Southern blot
analyses
oftheclonedDNAandwoodchuckgenomic
DNAs werecarriedout aspreviously reported
(17),
as was the electron microscopy of cloned DNA and
hetero-duplex
analysis.
The filamentousphage
recombinant M13-WHV wasused inheteroduplexes with HW197-2.Thisclone contains the entireWHV genomeclonedinto theEcoRI site ofvector M13mp7and waskindly
donatedby
W. Mason.RESULTS
Clone HW197-2 was isolated from a
library
ofrecombi-nant lambda phage
containing
nuclear DNAfrom a section ofliver HW197which showed noevidence of HCC. Animal HW197, from which the liver DNA wasobtained,
hadchronic active
hepatitis
withahigh
levelof virusproduction
and this animal died later without ever developing HCC. This is unusual forwoodchucks with chronicactive
hepatitis
in the Penrose Research Laboratorycolony.
Clone HW197-2 was
analyzed
for viral sequencesby
digesting
the cloned DNA with various restriction endonu-cleases which cut the viral genomeonly
one time.Restric-tion endonuclease digests were analyzed by Southern blot-ting and hybridization with cloned 3 P-labeled WHV DNA.
The initial Southern blot
(data
not shown) revealed that the clone contained two EcoRIfragments
which were 2.2 and6.3 kilobases
(kb)
inlength,
and bothhybridized
to WHV sequences.To map the position ofthe viral sequences in the clone,
electron microscope heteroduplex analysis of the cloned DNA wasconducted with an M13 clonecontainingtheentire WHV genome (3.3 kilobase pairs) inserted into M13 at the EcoRIsite. Wereasonedthathybridization ofclone HW197-2 to the WHV-M13 clone would cause the M13-WHV sequences to be circularized if the internal EcoRI site in HW197-2 was viral DNA. A structure containing circular-ized WHV sequences would be easily distinguishablewhen
analyzed by electron microscope
heteroduplex
analysis.
Aheteroduplex
was constructed between threemolecules,
clone HW197-2, M13-WHV, and Charon 30, and a
repre-sentative heteroduplex is shown in Fig. 1. Charon 30, the vector with itsstuffer fragment,wasincludedinthis
prepara-tion to establish the locations ofthe
junctions
between thecloned DNAand the vector arms.
Hybridization
of Charon30 to its homologous arms in clone HW197-2
clearly
delin-eated the ends ofthe cellular sequences in clone HW197-2
(see
Fig.
1, aand a'). Theunhybridized
stufferfragment
of Charon 30 is visible in Fig. 1, b and b'. Theheteroduplex
observed clearly demonstrated that the WHV sequences present inthe WHV-M13 clonewere circularizedwhenthey
hybridized
to clone HW197-2 (seeFig.
1, c, e,ande').
The brokenarms ofthe M13 vector are visible inFig.
1, d, and the heteroduplexregion
between the WHV sequences in cloneHW197-2and M13-WHVarevisible inFig.
1,eande'.This proved that the internal EcoRI site of cloneHW197-2 was viral, and measurement of the length ofthe heterodu-plexregionsoneither sideoftheinternalEcoRIsite revealed
that the totallength ofviral sequences in theclonewas2.75 kb. A single-stranded region of 550 base pairs (bp) was present in the circularized WHV sequences ofthe WHV-M13clone
(Fig.
1, c). Thisrepresented
the viral sequenceswhichweremissingin clone HW197-2
(approximately
550bp
between
positions
1,000and1,550
onthe viralmap) (6).
Thelengths of the left- and
right-hand
cellularflanking
sequences in clone HW197-2(Fig.
1, g andf,
respectively)
were determinedtobe 5.5and 0.6kb,
respectively, by
measuring
the
single-stranded regions
betweentheendoftheintegrated
WHVsequencesandthe
junction
withtheCharon30vectorarms. All ofthe
heteroduplexes observed,
including
broken molecules, were consistent with the structurepresented
in the intact molecule shown inFig.
1. No invertedduplica-tions, which would have been visualized as
hairpins
under the electronmicroscope, were observed.Further Southern blot
analysis
of clone HW197-2 DNAwasused as anindependent methodtoconfirmthe structure
of the viral sequences and to map additional restriction endonulease sites in the
flanking
cellular sequences. TheSouthern blot is
presented
inFig. 2,
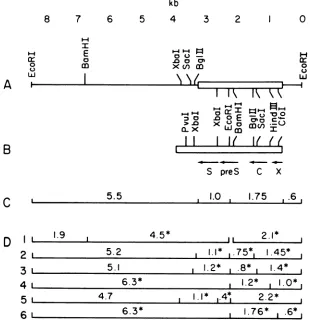
andthefinalrestriction endonuclease mapis showninFig.
3Aalong
witha map of thecomplete
WHVgenome(Fig. 3B)
and thelocation ofthe openreading
framesfor the viral surfaceantigen (S),
coreantigen
(C), X gene (X), andpre-S
sequences(preS).
Themeasurementsof viral and cellularsequencesfrom heterodu-plex analyses are
presented
inFig.
3C, and a summarydiagram illustrating
how theobserved restrictionendonucle-ase
fragments
confirmed the final mapisgiven
in Fig. 3D.Figure
2, lanes Sa andSb,
shows thatdoubledigestion
with EcoRIandCfoI
produced threefragments
whichhybridize
to WHV (6.3, 1.7 and 0.6
kb).
Thepublished
sequence for WHV shows aCfoI
site at 1,766bp
from the EcoRI site. Since the 0.6-kbfragment
hybridized weakly
to WHV, we conclude that theright-hand
virus-celljunction
is a short distancebeyond
the viralCfoI
site located 1,766 bp to theright of the
internal
viral EcoRI site in the clone. These resultswereconsistent withthe electronmicroscopehetero-duplex analysis which
positioned
the virus-cell junction 1,750bp tothe right ofthe internal EcoRI site.Theleft-hand virus-celljunctionoccurs veryclose to the 3' endofthesurface antigengeneaccordingtothe electron
microscope heteroduplex analysis.
Southern
blot analysis(Fig.
2)indicated that the Sacl andBglII
sites located 1.2 and 1.1 kb (respectively) to the left of the internalEcoRI
sitewere probably in cellular sequences because no hybridiza-VOL. 50, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
X
1
2
3
4
5
a
b
a
b
ab
ab
b
kb
23.8--9.6
-6.6
-4.4
-2.2
-1.9-0.5
-FIG. 1. Heteroduplex analysis ofWHVandcellularsequencesin clone HW197-2. DNA molecules usedwereCharon 30 (a,a,', b,b'),
clone HW197-2(wholelengthof heteroduplex), andM13-WHV (c, d, e, e'). Interpretation: (a and a')heteroduplex regions between
vector arms of Charon 30 and clone HW197-2; (b and b') single-stranded stufferfragment of Charon 30 vector;(c) single-stranded portion of WHV sequences in M13-WHV clone; (d) broken M13 vector arms from M13-WHV; (e and e') heteroduplex regions between WHV sequences in HW197-2 and M13-WHV clone; (f) single-stranded right-handwoodchuck cellularsequencein HW197-2; (g) single-stranded left-hand woodchuck cellular sequence in HW197-2.
tion of WHVwas observedtothe5.1-kb EcoRI-SacIorthe 5.2-kbEcoRI-BgIII fragments(Fig. 2, lanes 2aand band3a andb). Theseconclusions wereindependentlyconfirmed in
thenext experiment.
When DNA from clone HW197-2 was used to probe a Southern blot of EcoRI-digested, uninfected, woodchuck
genomic DNA,hybridization toasinglegenomic fragmentof approximately6.0 kbwasobserved. The size of thisgenomic fragment was approximately equal to the length of the
genomic sequences in clone HW197-2accordingtoelectron
microscopeanalysis. Thisobservation raised thepossibility
FIG. 2. Restriction endonuclease mapping and Southern blot analysis of the cellular and WHV sequences in clone HW197-2. Restriction endonuclease digests: (la) EcoRI; (2a)EcoRI + BglII;
(3a)EcoRI + Sacl; (4a)EcoRI +XbaI; (5a)EcoRI + CfoI.(Lanes lbtoSb)Southern blots of thecorresponding lanes, hybridized with
32P-labeled
WHV probe. A total of 10 x 106 cpm of 32P-labeled WHV DNA was prepared and hybridized as previously reported (17). Hybridization was for45min at 37°C. Vector DNA fragments in lanes la, 3a, and 4a are 33.5-, 21.5-, and 12-kb bands. Charon 30 vector fragments for lane 2a are the 21.0-, 7.0-, 4.0-, and 0.7-kb bands. Fainthybridization of the32P-labeledWHV probe to a 0.6-kb fragment inlaneSbwas stronger on longer exposureof the autora-diogram. The vector fragments for theEcoRI-CfoI digest were not mapped. Lambda,HindIlldigest of wild-type lambda phage; frag-ment sizesdesignated on the left.that theintegration ofWHV could haveoccurredin cellular sequencewithout any detectable rearrangement
(deletion
or duplication) of cellularsequences. In this case, insertion of 2.75 kbofviral DNA would haveresultedinthegeneration
ofanew fragment of8.75 kb which was
approximately
the size ofthe clonedDNA fragment.It was
possible
to test thishypothesis
in thefollowing
manner. Restriction endonuclease mappinghadestablished the presence of single BamHI,
BglII,
andSacl sites in theleft-handflanking cellular sequencesofclone HW 197-2. If nomajorrearrangementsof cellular sequences hadoccurred duringviralintegration,thenthecellularBamHI,BglII,and
Sacl restriction sites should be 4.2,
0.75,
and 0.85kb,
respectively,awayfrom theright-handEcoRIsite innormal uninfected woodchuck
genomic
DNA. A Southern blot of uninfected woodchuck DNA digested with EcoRI andBamHI, Sacl, or
BglII
wasprobed
with theright-hand
flanking sequences of clone HW197-2
(i.e.,
theright-hand
EcoRI fragment of clone HW197-2 was
used).
The results(Fig. 4A)showthatthisprobe
hybridized
tofragments
of thepredicted sizesfor all three double
digests
(4.2kbforon November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.68.302.78.485.2] [image:3.612.325.563.79.363.2]INTEGRATED WHV DNA FROM CHRONIC INFECTION 835
kb
8 7 6 5 4 3 2 1 0
E
I
0s
a]
.0 0
071
x>()
m\IaII\/.-F.
x
x
x oen(
II
I
(
I(
H
00
CC J
._O----pre-c
S preS C X
5.5 , 1.0 1.75 ,.6,
L 1.9 , 4.5* I
2.1*
l 5.2 1.1*
,.75*1
1.45*,5.1 , 1.2*, .8* 1.4* 1
1 6.3* , 1.2* , 1.0*,
6*4.7
1.1*6*4*
72.2*1
~~~6.
3* , 1.76*, .6*,1
FIG. 3. Restriction endonuclease mapof cloneHW197-2(A) and thecompleteWHVgenome(B). Symbolsfor(A): ( )woodchuck cellular sequences; (=El) WHV sequences. (B) Arrows indicate the location and direction of transcription of open reading frames correspondingtotheviralsurfaceantigengene(S),coreantigengene(C),Xgene(X),andpre-Ssequences(preS). (C) Measurementsof WHV and cellular sequences obtained from electron microscope heteroduplex analysis. (D) Schematic diagram of restriction endonuclease fragments produced by doubledigestions ofHW197-2 DNA with EcoRIplus (1)BamHI, (2) BglII, (3) SacI, (4) HindIII, (5) XbaI,and(6) CfoI.
* denotesfragments whichhybridize to32P-labeled WHVprobe.
BamHI, 0.75 kb forBglII-EcoRI,0.85 kb forSacI-EcoRI). A
probe containing the left-hand flanking sequences (the left-hand EcoRI fragment of HW197-2) was hybridized to a secondSouthern blot ofsimilarlydigested woodchuck DNA. Again the probe hybridizedtofragments of the sizes predict-ed fromtherestriction map of clone HW197-2 (Fig. 4B). It hybridized toEcoRI-BglIIandEcoRI-SacIfragments of 5.2 and 5.1 kb, respectively, and to two EcoRI-BamHI
frag-ments of 2.0 and 4.2 kb. The left-hand probe contains a mildly repeated sequence, andtherefore a smearof hybrid-ization was observed on Southern blots. However, intense hybridization to specific genomic bandswasclearly evident and distinguishable above the background smear. These results lead us to the conclusion that, within the level of resolution of thetechniques used (whichshouldbe 50to200 bp), there were no detectable rearrangements of cellular sequencesat thesite ofintegration of WHV DNA. Restric-tionmaps illustratingthe siteofintegrationof viral DNA in the cellular sequences, along witha schematic summary of theSouthern blotdatasupporting themaps,arepresentedin Fig. 4C and D. It is possible that short rearrangements (possibly directduplications)ofcellularsequences occurred
duringintegration and wouldnotbedetectedby the methods used.Thisquestioncanbe resolvedby sequencing the virus-celljunctions and the normal cell sequences at the site of integration. It is very clear, however, that large
rearrange-mentsofcellularsequencesdidnotoccuratthe site ofviral
integration ashas been observedfor the cellular sequences flanking WHV and HBV integrations from several HCCs
(manuscriptinpreparation).
DISCUSSION
Persistent infections with WHV share characteristics in common with other persistent virus infections, includinga
minimum cytopathic effect, limited immunopathological damage to persistently infected cells, and association of virus infections with tumors (12). In an attempt togain an
understandingofthe molecularbasis ofviral persistence in woodchuck hepatocytes, we began to study the forms of
viralDNAin the nuclei ofhepatocytes. The forms ofDNA described in a previous report (17) include (i) covalently
closed circularDNAwhich is themostabundantform,
being
present in approximately 50 copies per nucleus in liver
HW197; (ii) open circular DNA (possibly generated
by
nickingofcovalentlyclosed circular DNAduring isolation of nuclearDNA); and (iii) novelforms of WHV. These forms are composed oflong stretches of viral DNA (greater than
two genomes) which contain multiple rearrangements of viral sequences. Thesewere foundtobe presentin
approxi-matelyonecopypercell inliver HW197 butmaybepresent
inhighercopynumbers in other liver tissues.Theymayhave existed as large covalently closed circular moleculeswhich
I-I
0
u
Lu
A
I-B
C
D
2
3
4
5
6
VOL.50, 1984
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.150.460.78.399.2]836 ROGLER AND SUMMERS
A
B
X
1
2 3
4
X
1
2 3 4
kb
.3.8-9.6
-
6.6-4.4-
_
2.2
-_
I.9g__
S
0
01
_,
0
00:
0*.
C
1...
II.o-v..-Itt: I II
0 O C7
u EC to ac
D
L62XD
1 ,,
,,,
,6,2,,,,,,,,_4.2
i.7.5
lW^.
6.2
4.2 5.4 5.3
EcoRI
EcoRI+BomHI J Eco RI+Bgl1S
Eco RI+SacI
FIG. 4. Southern blot analysisof uninfected woodchuck cellularDNAdigestedwith various restriction endonucleases and hybridized with right-orleft-handflankingsequenceprobesfromclone HW197-2.(A)Southern blot of woodchuckDNA hybridized with the right-handEcoRI
fragment ofHW197-2. (B) Same as in (A) except the probe was the left-hand EcoRI fragment of HW197-2. Restriction endonuclease
digestions in (A) and(B)were:(1) EcoRIonly;(2) EcoRI +BamHI; (3) EcoRI+BglII;(4) EcoRI +SacI.(C) Proposed site of integration of WHV DNA in woodchuckcellular sequences: ( ) cellular DNA; (EOJ) WHV DNA. (D) Schematic diagram of results obtained from Southern blots (A) and (B) (above): restriction fragments hybridizingtotheright-hand probe (upper set) andleft-handprobe (lower set).
*Location of repetitioussequencein left-hand flankingsequences.
were linearized by cleavage withEcoRI, orthey may have beenexcisedfromlongerstretches ofrearrangedviralDNA
which was integrated in cellular DNA. In this report, we describe the structural characterization ofa fourth form of WHV DNAwhichisintegratedin cellular DNA in aboutone copy perthree cells.
Direct evidence for the function of any of the above molecules in the viral life cycle orin persistent infection is
completely lacking. At best, we can speculate as to their
possiblefunctions. Since theWHV lifecyclemayinvolvean RNA intermediate, similar to duck HBV, we have also
conductedexperimentstoattempttoisolateprovirusesfrom
acutelyinfected woodchuck and duck livers. These
experi-ments,conducted under thesameconditions in which inte-grated cloneswere isolated, have failed to identify aclone
containingacomplete provirus. Therefore,thenuclear cova-lently closed circular DNA present in high copy number in
hepatocytes is the most likely candidate to serve as the
templatefor theRNApregenomewhich wouldbenecessary
forreverse transcription.
The structural features of theintegratedWHV sequences
in thispapershow that thisintegratedmoleculemost
proba-2
EcoRI Eco RI+Bom HI EcoRI+BglIl
EcoRI+SocI
J.VIROL.
0
I
-1 -4
r-0
u
Ai
I --- 2.0
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.189.440.72.531.2]INTEGRATED WHV DNA FROM CHRONIC INFECTION 837
bly could not have supported viral replication because it
containedadefectivegenomewitha550-bpdeletion.Onthe other hand, the coding sequences for the pre-S, surface, core, and X genes are present in the integrated
molecule,
indicating thattranscription of thesesequences could result in production ofthose viral antigens. Whether this integra-tion significantly contributed to antigen production in the
infected hepatocyte in which it resided isunknown.Another
intriguing observation isthat the right-hand virus-cell
junc-tion of the clone is immediately adjacentto the proposed 5' end ofthe viral plus strand. Previous work (2. 8, 14) has
shown that virus-cell junctions in hepatomas frequently
occur in thevicinityof thecohesive endregion andoften in
the region of the genome which is single stranded in the
virionDNA.
Somegeneral characteristics ofWHV
integrations
presentin HCCs include: (i)integration atdifferent sites in boththe
viral and cellular sequences in independent hepatomas; (ii) variation in the amount of viral DNA in individual integra-tions from less than a genome length to greater than genome lengths; (iii)
complex
rearrangements of viralse-quences inthe
integrations, including
inverted duplications anddeletions; and(iv)
rearrangementsof cellularsequences at the site of viral integration(manuscript
inpreparation).
The integration described in this paper differs from those found in hepatomas in that the viral sequences did not contain any inverted
duplications
and there was nodetect-able rearrangement of cellular sequences at the site of
integration.
Isolation and structural characterizationofaddi-tional integrations from
chronically
infected tissues are in progress to determine whether these differences can begeneralized.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grants Al-15166, RR-05539, and CA-06927 from the National Cancer Institute andbyanappropriation from the Commonwealth ofPennsylvania.
LITERATURE CITED
1. Brechot,C.,M.Hadchouel, J.Scotto,M.Fonck,F.Potet,G. N. Vyas, and P.Tiollais. 1981. State ofHepatitisB virus DNAin hepatocytes of patients withHepatitis B surface
antigen-posi-tive andnegative liver diseases. Proc. Natl. Acad. Sci. U.S.A. 78:3906-3910.
2. Dejean, A.,C.Brechot, P.Tiollais,andS. Wain-Hobson. 1983. Characterization of integrated hepatitis B viral DNA cloned from a human hepatoma and the hepatoma derived cell line PLC/PRF/5. Proc.Natl. Acad.Sci. U.S.A. 80:2505-2509. 3. Dejean, A.,L.Vitvitski,C.Brechot, C. Trepo,P.Tiollais,and P.
Charnay. 1982. Presence andstateof woodchuckhepatitisvirus inliver andserumof woodchucks: furtheranalogieswith human hepatitis Bvirus. Virology121:195-199.
4. Enquist, L. W., M. J. Madden, P. Schrop-Stansly, and G. F. Vande Woude. 1979. Cloning of herpes simplex type I DNA fragments in bacteriophage lambda vector. Science 203:541-544.
5. Enquist, L., and N. Sternberg. 1979. In Vitro packaging of lambda Damvectors and theirusein cloningDNAfragments.
Methods Enzymol. 68:281-298.
6. Galibert, F., T. N. Chen, and E. Mandart. 1982. Nucleotide
sequence ofaclonedwoodchuckhepatitis virus genome:
com-parisonwiththehepatitis B virus sequence. J. Virol. 41:51-65. 7. Gilboa, E., S. W. Mitra, S. Goff, and D. Baltimore. 1979. A
detailed model of reverse transcription and tests of crucial aspects.Cell 18:93-100.
8. Koshy, R., S. Koch, A. von Loringhoven, R. Kahmann, K.
Murray,and P. H.Hofschneider. 1983.Integration ofHepatitis
B Virus DNA: evidencefor integration in the single-stranded gap. Cell34:215-223.
9. Marion,P.L.,L.S.Oshiro,D.C. Regnery, G. H. Scullard,and W. S.Robinson.1980.Avirus in Beechey ground squirrels that is relatedtohepatitisBvirus in humans. Proc.Natl. Acad.Sci. U.S.A. 77:2941-2945.
10. Mason,W.S., G. Seal, andJ. Summers. 1980. Virus of Pekin ducks with structural and biological relatedness to human hepatitis B virus. J. Virol. 36:829-836.
11. Mason,W. S., J. Taylor, G. Seal, andJ. Summers. 1981. An HBV-like virus of domestic ducks, p. 107-118. In W.Szmuness,
H.Alter, and J. Maynard (ed.), Viral hepatitis, 1981 Internation-alSymposium. Franklin Press,Philadelphia.
12. Mims, C. A.1974. Factors in themechanisms of persistence of viralinfections. Prog. Med. Virol. 18:1-14.
13. Molnar-Kimber, K. L., J. Summers, J. M. Taylor, andW. S. Mason. 1983. Protein covalently bound to minus-strand DNA intermediates of duck hepatitis B virus. J. Virol.45:165-172. 14. Ogston, C. W., G. J. Jonak, C. E. Rogler, S. M. Astrin, andJ.
Summers. 1982. Cloning and structural analysis of integrated woodchuck hepatitisvirussequences from hepatocellular carci-nomasofwoodchucks. Cell 29:385-394.
15. Rim, R. L., D. Horness, J. Kucera, and F. R.Blattner. 1980. Construction of coliphage lambda Charon vectors with BamHi cloning sites. Gene 12:301-309.
16. Robinson, W.,P. Marion,M. Feitelson, and A.Siddiqui. 1981. Thehepadna virus group: hepatitis Bandrelatedviruses, p. 57. In W. Szmuness, H. J. Alter, and J. E. Maynard (ed.), Viral
Hepatitis,1981InternationalSymposium. Franklin Press,
Phila-delphia.
17. Rogler, C. E., andJ. Summers. 1982. Novel forms of wood-chuck hepatitis virus DNA isolated from chronically infected woodchuck liver nuclei. J. Virol. 44:852-863.
18. Shafritz, D. A., D. Shouval, H. Sherman, S. J. Hadziyannis, and M. C.Kew. 1981. Integration of hepatitis B virus DNA into the genome ofliver cells inchronic liver disease andhepatocellular carcinoma. N.Engl. J. Med. 305:1067-1073.
19. Snyder, R. L., and L. Summers. 1980. Woodchuck hepatitis virus andhepatocellularcarcinoma. ColdSpring Harbor Conf. CellProlif. 7:447-458.
20. Summers, J. 1975. Physical map of polyoma viral DNA frag-ments produced by cleavage with a restriction enzyme from Haemophilus aegyptius, endonuclease R-HaeIII. J. Virol.
15:946-953.
21. Summers, J. 1981. Threerecentlydescribed animal virus models for humanhepatitis B virus. Hepatology 1:179-183.
22. Summers, J.,and W.S. Mason.1982.Replication of the genome ofa hepatitis B-like virus by reversetranscription ofanRNA intermediate. Cell29:403-415.
23. Summers, J., J.M.Smolec,and R.Snyder.1978. Avirus similar tohumanhepatitisBvirus associated withhepatitis and hepato-ma in woodchucks. Proc. Natl. Acad. Sci. U.S.A. 75:4533-4537.
24. VanWoude,G.F.,M.Oskarsson,L. W.Enquist, S. Nomura, M.
Sullivan, and P. J. Fischinger. 1979. Cloning of integrated Moloney sarcoma proviral DNA sequences in bacteriophage lambda. Proc. Natl. Acad. Sci. U.S.A.76:4464-4468. VOL. 50,1984